A Trypsin Inhibitor from Tamarind Reduces Food Intake and Improves Inflammatory Status in Rats with Metabolic Syndrome Regardless of Weight Loss

 , , , , and
, , , , and
Abstract
:
1. Introduction
2. Materials and Methods
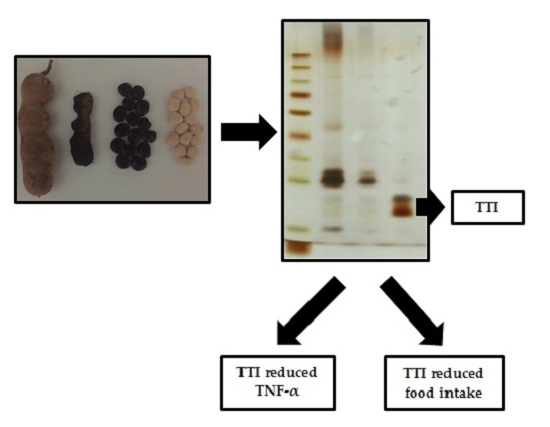
2.1. Extraction, Fractioning and Isolation of the Trypsin Inhibitor
2.2. In Vivo Experiment
2.2.1. Animals and Experiment Design
2.2.2. Diets
2.2.3. Obesity and Metabolic Syndrome (MetS) Confirmation
2.2.4. Food Intake and Weight Gain
2.2.5. Biochemical and Inflammatory Parameters
2.3. Statistical Analysis
3. Results
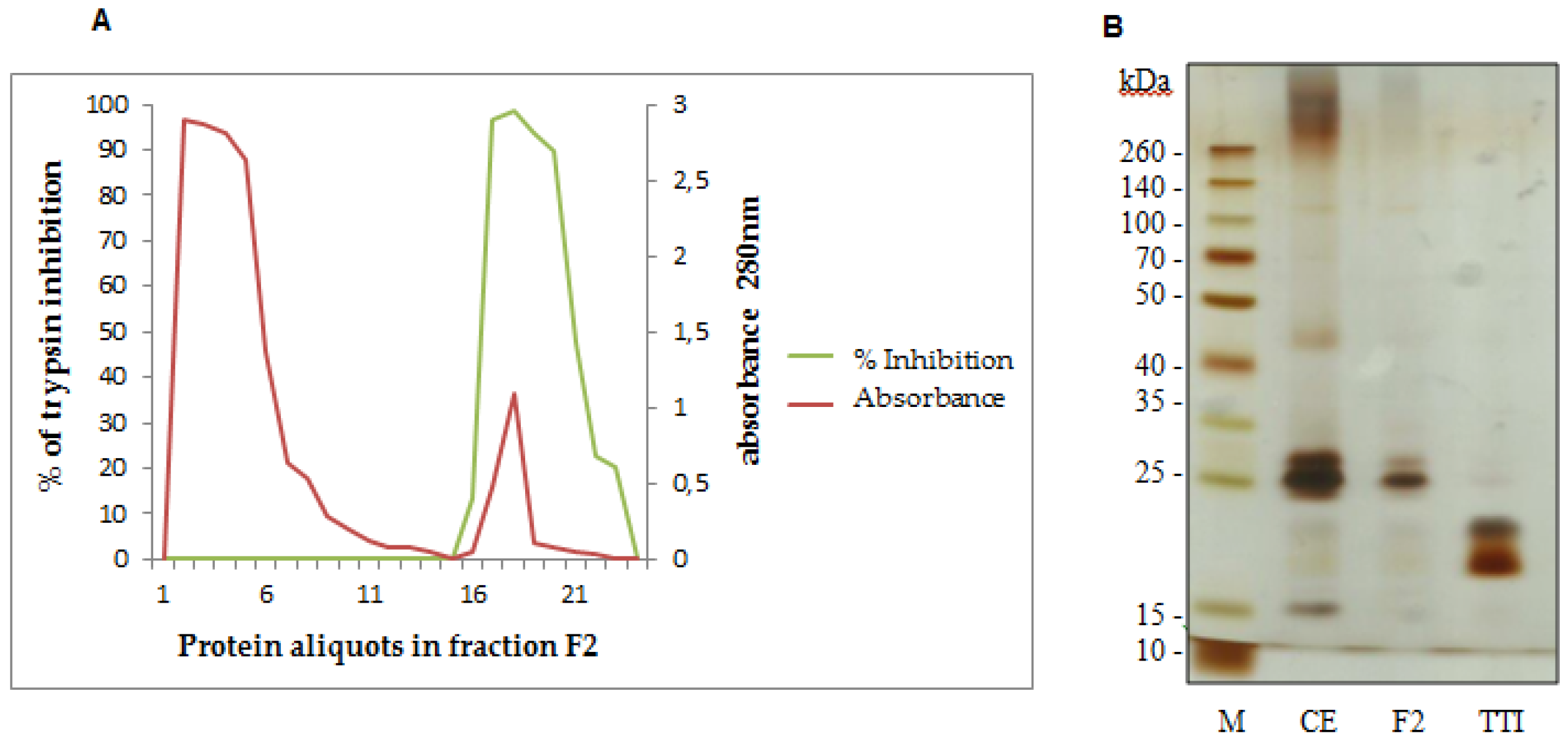
3.1. TTI Isolation in Trypsin-Sepharose Affinity Chromatography
3.2. Metabolic Syndrome (MetS) Confirmation
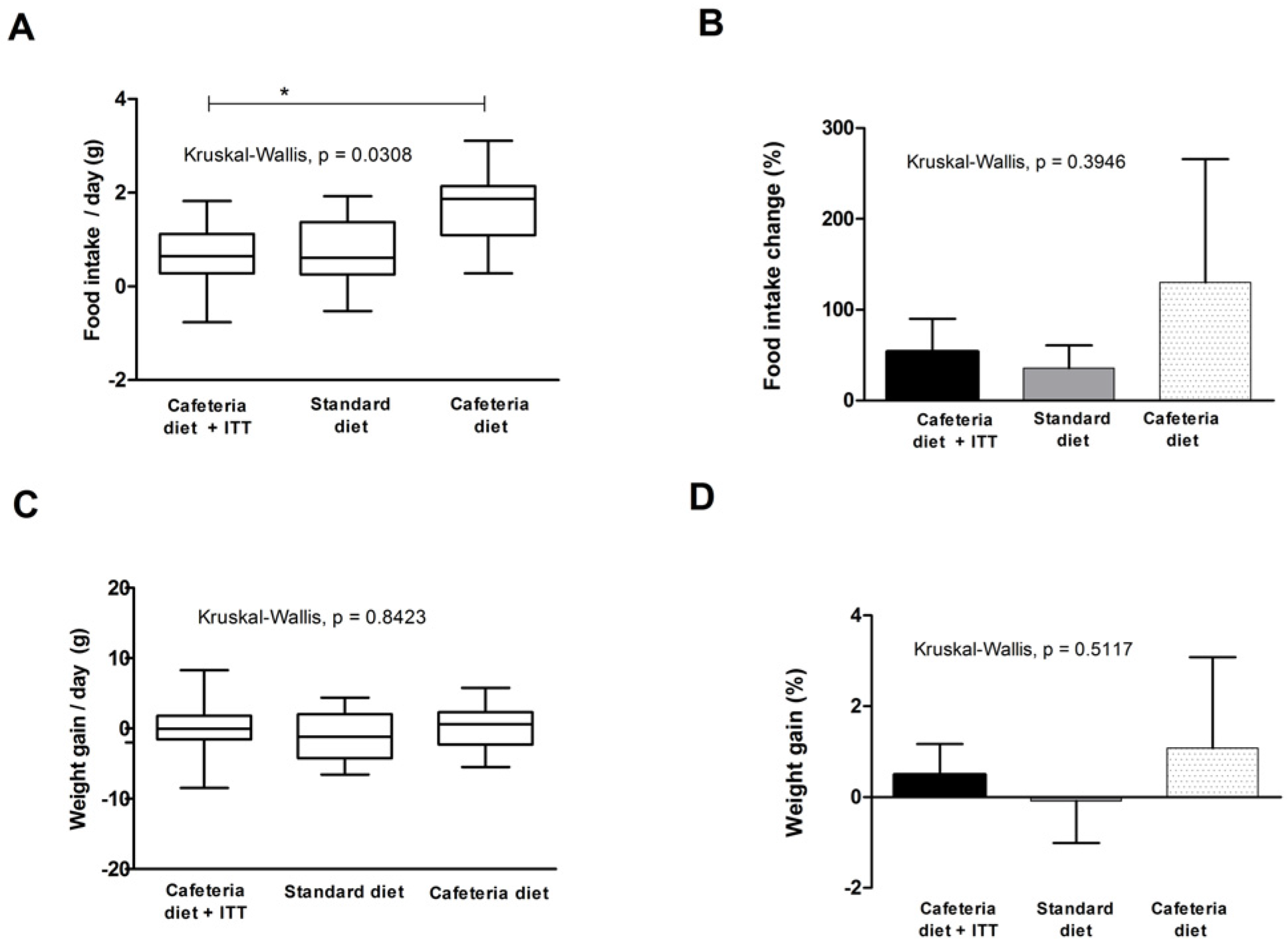
3.3. Food Intake and Weight Gain
3.4. Biochemical and Inflammatory Parameters
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kaplan, N.M. The deadly quartet. Upper-body obesity, glucose intolerance, hypertriglyceridemia, and hypertension. Arch. Intern. Med. 1989, 149, 514–520. [Google Scholar] [CrossRef]
- World Health Organization. Global Action Plan for the Prevention and Control of NCDs 2013–2020. Available online: http://apps.who.int/gb/ebwha/pdf_files/WHA66/A66_R10-en.pdf (accessed on 1 February 2016).
- Shoelson, S.E.; Herrero, L.; Naaz, A. Obesity, inflammation, and insulin resistance. Gastroenterology 2007, 132, 2169–2180. [Google Scholar] [CrossRef] [PubMed]
- Krinninger, P.; Brunner, C.; Ruiz, P.A. Role of the adipocyte-specific NF-ĸB activity in the regulation of IP-10 and T cell migration. Am. J. Physiol Endocrinol. Metab. 2011, 300, E304–E311. [Google Scholar] [CrossRef] [PubMed]
- Ouchi, N.; Parker, J.L.; Lugus, J.J.; Walsh, K. Adipokines in inflammation and metabolic disease. Nat. Rev. Immunol. 2011, 11, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Kanneganti, T.D.; Dixit, V.D. Immunological complications of obesity. Nat. Immunol. 2012, 13, 707–712. [Google Scholar] [CrossRef] [PubMed]
- Weisberg, S.P.; McCann, D.; Desai, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W., Jr. Obesity is associated with macrophage accumulation in adipose tissue. J. Clin. Investig. 2013, 112, 1796–1808. [Google Scholar] [CrossRef]
- Machado, R.J.A.; Monteiro, N.K.V.; Migliolo, L.; Silva, O.N.; Pinto, M.F.S.; Oliveira, A.S.; Franco, O.L.; Kiyota, S.; Bemquerer, M.P.; Santos, E.A.; et al. Characterization and pharmacological properties of a novel multifunctional kunitz inhibitor from Erythrina velutina seeds. PLoS ONE 2013, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Lalloyer, F.; Staels, B. Fibrates, glitazones, and peroxisome proliferator-activated receptors. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 894–899. [Google Scholar] [CrossRef] [PubMed]
- Barroso, I.; Gurnell, M.; Crowley, V.E.F.; Agostini, M.; Schwabel, J.W.; Soos, M.A. Dominant negative mutations in human PPARg associated with severe insulin resistance, diabetes mellitus and hypertension. Nature 1999, 402, 880–883. [Google Scholar] [PubMed]
- Shah, A.; Mehta, N.; Reilly, M.P. Adipose inflammation, insulin resistance, and cardiovascular disease. J. Parenter. Enteral Nutr. 2008, 32, 638–644. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.F.; Plutzky, J. Peroxisome proliferator activated receptors as transcriptional nodal points and therapeutic targets. Circulation 2007, 115, 518–533. [Google Scholar] [CrossRef] [PubMed]
- Fang, E.F.; Wong, J.H.; Bah, C.S.F.; Lin, P.; Tsao, S.W.; Ng, T.B. Bauhinia variegate var. variegate trypsin inhibitor: From isolation to potencial medicinal applications. Biochem. Biophys. Res. Commun. 2010, 396, 806–811. [Google Scholar] [CrossRef] [PubMed]
- Scoott, C.J.; Taggart, C.C. Biologic protease inhibitors as novel therapeutic agents. Biochimie 2010, 92, 1681–1688. [Google Scholar] [CrossRef] [PubMed]
- Serquiz, A.C.; Machado, R.J.A.; Serquiz, R.P.; Lima, V.C.O.; Carvalho, F.M.C.; Carneiro, M.A.A.; Maciel, B.L.L.; Uchôa, A.F.; Santos, E.A.; Morais, A.H.A. Supplementation with a new trypsin inhibitor from peanut is associated with reduced fasting glucose, weight control, and increased plasma CCK secretion in an animal model. J. Enzyme Inhib. Med. Chem. 2016, 29, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Santos, E.A.; Oliveira, A.S.; Rabêlo, L.M.A.; Uchôa, A.F.; Morais, A.H.A. Affinity Chromatography as a Key Tool to Purify Protein Protease Inhibitors from Plants (2012). Affinity Chromatography. Available online: http://www.intechopen.com/download/get/type/pdfs/id/33057 (accessed on 1 February 2016).
- Fook, J.M.; Macedo, L.L.P.; Moura, G.E.; Teixeira, F.M.; Oliveira, A.S.; Queiroz, A.F.S. A serine proteinase inhibitor isolated from Tamarindus indica seeds and its effects on the release of human neutrophil elastase. Life Sci. J. 2005, 76, 2881–2891. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Hira, T.; Nakajima, S.; Tomozawa, H.; Tsubata, M.; Yamaguchi, K. Suppressive effect on food intake of a potato extract (Potein®) involving cholecystokinin release in rats. Biosci Biotechinol Biochem. 2012, 76, 1104–1109. [Google Scholar] [CrossRef] [PubMed]
- Lima, V.C.O.; Machado, R.J.A.; Monteiro, N.K.V.; Lyra, I.L.; Camilo, C.S.; Serquiz, A.C.; Oliveira, A.S.; Rufino, F.P.S.; Maciel, B.L.L.; Uchôa, A.F.; et al. Gastroprotective and antielastase effect of protein inhibitors from Erythrina velutina seeds in na experimental ulcer model. Biochem. Cell Biol. 2016. [Google Scholar] [CrossRef]
- Ribeiro, J.A.; Serquiz, A.C.; Silva, P.F.; Barbosa, P.B.; Sampaio, T.B.; Araujo Junior, R.F.; Oliveira, A.S.; Machado, R.J.A.; Maciel, B.L.L.; Uchôa, A.F.; et al. Trypsin inhibitor from Tamarindus Indica L. seeds reduces weight gain and food consumption and increases plasmatic cholecystokinin levels. Clinics 2015, 70, 136–143. [Google Scholar] [CrossRef]
- Nakajima, S.; Hira, T.; Tsubata, M.; Takagaki, K.; Hara, H. Potato extract (Potein) suppresses food intake in rats through inhibition of luminal trypsin activity and direct stimulation of cholecystokinin secretion from enteroendocrine cells. J. Agric. Food Chem. 2011, 59, 9491–9496. [Google Scholar] [CrossRef] [PubMed]
- Komarnytsky, S.; Cook, A.; Raskin, I. Potato protease inhibitors inhibit food intake and increase circulating cholecystokinin levels by a trypsindependent mechanism. Int. J. Obes. 2011, 35, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Raybould, H. Mechanisms of CCK signaling from gut to brain. Curr. Opin. Pharmacol. 2007, 7, 570–574. [Google Scholar] [CrossRef] [PubMed]
- Duca, F.A.; Covasa, M. Current and emerging concepts on the role of peripheral signals in the control of food intake and development of obesity. Br. J. Nutr. 2012, 108, 778–793. [Google Scholar] [CrossRef] [PubMed]
- Sayegh, A.I.; Washington, M.C.; Johnson, R.E.; Johnson-Rouse, T.; Freeman, C.; Harrison, A.; Lucas, J.; Shelby, M.; Fisher, B.; Willis, W.; et al. Celiac and the cranial mesenteric arteries supply gastrointestinal sites that regulate meal size and intermeal interval length via cholecystokinin-58 in male rats. Horm. Behav. 2015, 67, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Gomez, G.; Englander, E.W.; Greeley, G.H., Jr. Nutrient inhibition of ghrelin secretion in the fasted rat. Regul. Pept. 2004, 117, 33–36. [Google Scholar] [CrossRef] [PubMed]
- Peters, J.H.; Karpiel, A.B.; Ritter, R.C.; Simasko, S.M. Cooperative activation of cultured vagal afferent neurons by leptin and cholecystokinin. Endocrinology 2004, 145, 3652–3657. [Google Scholar] [CrossRef] [PubMed]
- Araújo, C.L.; Bezerra, I.W.L.; Oliveira, A.S.; Moura, F.T.; Macedo, L.L.P.; Gomes, C.E.M.; Barbosa, A.E.A.D.; Macedo, F.P.; Souza, T.M.S.; Franco, O.L.; et al. In vivo bioinsecticidal activity toward Ceratitis Capitata (fruit fly) and Callosobruchus Maculatus (cowpea weevil) and in vitro bioinsecticidal activity toward different orders of insect pests of a trypsin inhibitor purified from tamarind tree (Tamarindus Indica) seeds. Agric. Food Chem. 2005, 53, 4381–4387. [Google Scholar]
- Nascimento, A.F.; Sugizaki, M.M.; Leopoldo, A.S.; Lima-Leopoldo, A.P.; Luvizotto, R.A.; Nogueira, C.R. A hypercaloric pellet-diet cycle induces obesity and comorbidities in Wistar rats. Arq. Bras. Endocrinol. Metabol. 2008, 52, 968–974. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.; Collingwood, T.N.; Rebar, E.J.; Wolfe, A.P.; Camp, H.S. PPARg knockdown by engineered transcription factors: exogenous PPARg2 but not PPARg1 reactivates adipogenesis. Gene Dev. 2002, 16, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Tontonoz, P.; Hu, E.; Spiegelman, B.M. Stimulation of adipogenesis in fibroblasts by PPARg2, a lipid activated transcription factor. Cell 1994, 79, 1147–1156. [Google Scholar] [CrossRef]
- Schoonjans, K.; Watanabe, M.; Suzuki, H. Induction of the acyl-coenzyme A synthetase gene by fibrates and fatty acids is mediated by a peroxisome proliferators response element in the C promoter. J. Biol. Chem. 1995, 270, 19269–19276. [Google Scholar] [PubMed]
- Schoonjans, K.; Staels, B.; Auwerx, J. Role of the peroxisome proliferators activated receptor (PPAR) in mediating effects of fibrates and fatty acids on gene expression. J. Lipid Res. 1996, 37, 907–925. [Google Scholar] [PubMed]
- Sfeir, Z.; Ibrahimi, A.; Amri, E. Regulation of FAT/CD36 gene expression: further evidence in support of a role of the protein in fatty acid binding/transport. Prostaglandins Leukot. Essent. Fatty Acids 1997, 57, 17–21. [Google Scholar] [CrossRef]
- Kakade, M.L.; Simons, N.; Liener, I.E. An evaluation of natural vs. synthetic substrates for measuring the antitryptic activity of soybean samples. Cereal Chem. 1969, 46, 518–526. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Oakley, B.R.; Kirsch, D.R.; Morris, N.R. A simplified ultrasensitive silver stain for detecting proteins in polyacrylamide gels. Anal. Biochem. 1980, 105, 361–363. [Google Scholar] [CrossRef]
- Committee for the Update of the Guide for the Care and Use of Laboratory Animals. Guide for the Care and Use of Laboratory Animals, 8th ed.; Institute for Laboratory Animal Research. Division on Earth and Life Studies; The National Academies Press: Washington, DC, USA, 2011; p. 246. [Google Scholar]
- Naderali, E.K.; Pickavance, L.C.; Wilding, J.P.H.; Williams, G. Diet-induced endothelial dysfunction in the rat is independent of the degree of increase in total body weight. Clin. Sci. 2001, 100, 635–641. [Google Scholar] [CrossRef] [PubMed]
- Novelli, E.L.B.; Diniz, Y.S.; Galhardi, C.M.; Ebaid, G.M.X.; Rodrigues, H.G.; Mani, F. Anthropometrical parameters and markers of obesity in rats. Lab. Anim. 2007, 41, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Expert Panel on Detection, Evaluation and Treatment of High Blood Cholesterol in Adults. Executive summary of the Third Report of the National Cholesterol Education Program (NCEP) Expert Panel on Detection, Evaluation and Treatment of High Cholesterol. JAMA 2001, 285, 2486–2497. [Google Scholar]
- Guimarães, M.A.; Mazáro, R. Princípios éticos e Práticos do uso de Animais de Experimentação; UNIFESP: São Paulo, Brazil, 2004. [Google Scholar]
- Vendramea, S.; Daughertya, A.; Kristoa, A.S.; Risob, P.; Klimis-Zacasa, D. Wild blueberry (Vaccinium Angustifolium) consumption improves inflammatory status in the obese Zucker rat model of the metabolic syndrome. J. Nutr. Biochem. 2013, 24, 1508–1512. [Google Scholar] [CrossRef] [PubMed]
- Vanzela, E.C.; Ribeiro, R.A.; Oliveira, C.A.M.; rodrigues, F.B.; Bonfleur, M.L.; Carneiro, E.M.; Souza, K.L.A.; Boschero, A.C. Pregnancy restores insulin secretion from pancreatic islets in cafeteria diet-induced obese rats. Am. J. Physiol. 2010, 298, R320–R328. [Google Scholar] [CrossRef] [PubMed]
- Macedo, I.C.; Medeiros, L.F.; Oliveira, C.; Oliveira, C.M.; Rozisky, J.R.; Scarabelot, V.L.; Souza, A.; Silva, F.R.; Santos, V.S.; Coioato, S.G.; et al. Cafeteria diet-induced obesity plus chronic stress alter serum leptin levels. Peptides 2012, 38, 189–196. [Google Scholar] [CrossRef] [PubMed]
- White, P.A.S.; Cecato, L.M.; Araújo, J.M.D.; Souza, L.A.; Soares, A.F.; Barbosa, A.P.O.; Neto, J.M.R.; Marçal, A.C.; Machado, U.F.; Camargo, E.A.; et al. Model of high-fat diet-induced obesity associated to insulin resistance and glucose intolerance. Arq. Bras. Endocrinol. Metab. 2013, 57, 339–345. [Google Scholar] [CrossRef]
- Castro, H.; Pomar, C.A.; Picó, C.; Sánchez, J.; Palou, A. Cafeteria diet over feeding in Young male rats impairs the adaptive response to fed/fasted conditions and increases adiposity independent of body weight. Int. J. Obes. 2015, 39, 430–437. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, C.L.; Peikin, S.R.; Baile, C.A. Trypsin inhibitor effects on food intake and weight gain in Zucker rats. Physiol. Behav. 1983, 31, 487–491. [Google Scholar] [CrossRef]
- Li, Z.; Maglione, M.; Tu, W.; Mojica, W.; Arterburn, D.; Shugarman, L.R.; Hilton, L.; Suttorp, M.; Solomon, V.; Shekelle, P.G.; et al. Meta-analysis: pharmacologic treatment of obesity. Ann. Intern. Med. 2005, 142, 532–546. [Google Scholar] [CrossRef] [PubMed]
- Yanovski, S.Z.; Yanovski, J.A. Obesity. N. Engl. J. Med. 2002, 346, 591–602. [Google Scholar] [PubMed]
- Gomez-Zorita, S.; Fernandez-Quintela, A.; Lasa, A.; Hijona, E.; Bujanda, L.; Portillo, M.P. Effects of resveratrol on obesity-related inflammation makers in adipose tissue of genetically obese rats. Nutrition 2013, 29, 1374–1380. [Google Scholar] [CrossRef] [PubMed]
- Reis, T.A.; Goulart, P.F.P.; Oliveira, R.M.E.; Oliveira, L.; Abreu, P.S.; Azevedo, A.O. Metabolic parameters of Wistar rats subjected to diet supplemented with stevia and sugar. Semin. Ciênc. Agrar. 2011, 32, 1477–1488. [Google Scholar] [CrossRef]
- Roy, D.M.; Schneeman, B.O. Effect of soy protein, casein and trypsin inhibitor on cholesterol, bile acids and pancreatic enzymes in mice. J. Nutr. 1981, 111, 878–885. [Google Scholar] [PubMed]
- Gregor, M.F.; Hotamisligil, G.S. Inflammatory Mechanisms in Obesity. Annu. Rev. Immunol. 2011, 29, 415–445. [Google Scholar] [CrossRef] [PubMed]
- Petros, W.P.; Peters, W.P. Cytokines. In Therapeutic Immunology, 1st ed.; Austen, K.F., Burakoff, S.J., Rosen, F.S., Strom, T.B., Eds.; Blackwell Science, Inc.: Malden, MA, USA, 1996; pp. 229–279. [Google Scholar]
- Durum, S.K.; Muegge, K. Cytokines linking the immune and inflammatory systems. In Clinical Immunology, 1st ed.; Rich, R., Ed.; Mosby-Year book, Inc.: St Louis, MO, USA, 1996; pp. 350–362. [Google Scholar]
- Sarker, M.R.; Franks, S.; Sumien, N.; Thangthaeng, N.; Filipetto, F.; Forster, M. Curcumin Mimics the Neurocognitive and Anti-Inflammatory Effects of Caloric Restriction in a Mouse Model of Midlife Obesity. PLoS ONE 2015, 10, e0140431. [Google Scholar] [CrossRef] [PubMed]
- Cohen, A.N.; Veena, M.S.; Srivatsan, E.S.; Wang, M.B. Suppression of interleukin 6 and 8 production in head and neck cancer cells with curcumin via inhibition of Ikappa beta kinase. Arch. Otolaryngol. Head Neck Surg. 2009, 135, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, B.B.; Shishodia, S. Molecular targets of dietary agents for prevention and therapy of cancer. Biochem. Pharmacol. 2006, 71, 1397–1421. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Ingredients | Cafeteria Diet | Standard Diet |
|---|---|---|
| Labina® (g) | 45.2 | 100.0 |
| Condensed milk (mL) | 45.2 | - |
| Sugar (g) | 9.6 | - |
| Total | 100.0 | 100.0 |
| Lee Index * (g/cm3) | Waist Circumference * (cm) | Fasting Glucose * (mg/dL) | TG * (mg/dL) | HDL * (mg/dL) |
|---|---|---|---|---|
| 0.280 ± 0.0 | 14.1 ± 0.2 | 68.0 ± 13.5 | 44.6 ± 19.6 | 29.8 ± 4.7 |
| Rats | Lee Index (g/cm3) | Waist Circumference (cm) | Fasting Glucose (mg/dL) | TG (mg/dL) | HDL-C (mg/dL) |
|---|---|---|---|---|---|
| R1 | 0.303 * | 16.0 * | 127 * | 260 * | 43 |
| R2 | 0.314 * | 17.0 * | 129 * | 192 * | 40 |
| R3 | 0.312 * | 16.0 * | 158 * | 58 | 35 |
| R4 | 0.303 * | 16.0 * | 105 * | 45 | 33 |
| R5 | 0.313 * | 16.5 * | 186 * | 53 | 35 |
| R6 | 0.308 * | 16.5 * | 167 * | 192 * | 44 |
| R7 | 0.302 * | 17.0 * | 112 * | 191 * | 33 |
| R8 | 0.310 * | 16.5 * | 184 * | 147 * | 66 |
| R9 | 0.315 * | 16.0 * | 151 * | 42 | 22 * |
| R10 | 0.306 * | 16.0 * | 115 * | 74 * | 28 |
| R11 | 0.307 * | 16.0 * | 151 * | 106 * | 30 |
| R12 | 0.304 * | 16.0 * | 123 * | 49 | 25 |
| R13 | 0.304 * | 16.0 * | 113 * | 42 | 24 * |
| R14 | 0.305 * | 16.0 * | 99 * | 61 | 29 |
| R15 | 0.306 * | 17.0 * | 173 * | 44 | 23 * |
| Parameters | Cafeteria Diet + TTI Mean ± SD | Standard Diet Mean ± SD | Cafeteria Diet Mean ± SD |
|---|---|---|---|
| Fasting glucose (mg/dL) | 114.4 ± 34.6 | 84.0 ± 34.7 | 203.3 ± 122.0 |
| Total cholesterol (mg/dL) | 127.6 ± 62.3 | 84.6 ± 45.3 | 102.6 ± 32.8 |
| HDL-C (mg/dL) | 31.8 ± 6.4 | 26.4 ± 8.4 | 23.4 ± 3.9 |
| LDL-C (mg/dL) | 71.2 ± 62.9 | 45.8 ± 36.4 | 50.2 ± 31.8 |
| VLDL-C (mg/dL) | 24.58 ± 6.9 | 12.4 ± 3.6 * | 29.0 ± 7.1 |
| TG (mg/dL) | 123.2 ± 34.6 | 62.2 ± 18.0 * | 145.0 ± 35.3 |
| SGOT (mg/dL) | 17.00 ± 1.41 | 15.80 ± 3.35 | 19.00 ± 5.34 |
| SGPT (mg/dL) | 28.50 ± 17.94 | 31.40 ± 9.56 | 43.20 ± 19.20 |
| GGT (mg/dL) | 19.25 ± 2.99 | 22.80 ± 17.11 | 18.00 ± 5.10 |
| IL-6 (pg/mL) | 1.68 ± 0.24 | 2.25 ± 1.17 | 1.85 ± 0.13 |
| TNF- α (pg/mL) | undetectable | 6.53 ± 0.96 # | 5.86 ± 0.43 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carvalho, F.M.C.; Lima, V.C.O.; Costa, I.S.; Medeiros, A.F.; Serquiz, A.C.; Lima, M.C.J.S.; Serquiz, R.P.; Maciel, B.L.L.; Uchôa, A.F.; Santos, E.A.; et al. A Trypsin Inhibitor from Tamarind Reduces Food Intake and Improves Inflammatory Status in Rats with Metabolic Syndrome Regardless of Weight Loss. Nutrients 2016, 8, 544. https://doi.org/10.3390/nu8100544
Carvalho FMC, Lima VCO, Costa IS, Medeiros AF, Serquiz AC, Lima MCJS, Serquiz RP, Maciel BLL, Uchôa AF, Santos EA, et al. A Trypsin Inhibitor from Tamarind Reduces Food Intake and Improves Inflammatory Status in Rats with Metabolic Syndrome Regardless of Weight Loss. Nutrients. 2016; 8(10):544. https://doi.org/10.3390/nu8100544
Chicago/Turabian StyleCarvalho, Fabiana M. C., Vanessa C. O. Lima, Izael S. Costa, Amanda F. Medeiros, Alexandre C. Serquiz, Maíra C. J. S. Lima, Raphael P. Serquiz, Bruna L. L. Maciel, Adriana F. Uchôa, Elizeu A. Santos, and et al. 2016. "A Trypsin Inhibitor from Tamarind Reduces Food Intake and Improves Inflammatory Status in Rats with Metabolic Syndrome Regardless of Weight Loss" Nutrients 8, no. 10: 544. https://doi.org/10.3390/nu8100544
APA StyleCarvalho, F. M. C., Lima, V. C. O., Costa, I. S., Medeiros, A. F., Serquiz, A. C., Lima, M. C. J. S., Serquiz, R. P., Maciel, B. L. L., Uchôa, A. F., Santos, E. A., & Morais, A. H. A. (2016). A Trypsin Inhibitor from Tamarind Reduces Food Intake and Improves Inflammatory Status in Rats with Metabolic Syndrome Regardless of Weight Loss. Nutrients, 8(10), 544. https://doi.org/10.3390/nu8100544