Effects of Different Processing on miRNA and Protein in Small Extracellular Vesicles of Goat Dairy Products

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Acquisition and Pretreatment of Goat Milk Samples
2.2. Extraction of Milk Small Extracellular Vesicles
2.3. Western Blot
2.4. Nanoparticle Tracking Analysis (NTA)
2.5. Transmission Electron Microscope (TEM)
2.6. Extraction of miRNA, Small RNA Library Construction, and Sequencing
2.6.1. Extraction and Detection of Milk Small Extracellular Vesicle miRNA
2.6.2. Small RNA Library Construction and miRNA Sequencing
2.7. Mass Spectrum and Proteomic Analysis of Milk Small Extracellular Vesicle Proteins
2.7.1. Extraction, Enzymatic Hydrolysis, and Peptide Acquisition of Milk Small Extracellular Vesicle Proteins
2.7.2. Mass Spectrum Detection and Database Search
2.8. Statistical Analysis
3. Results
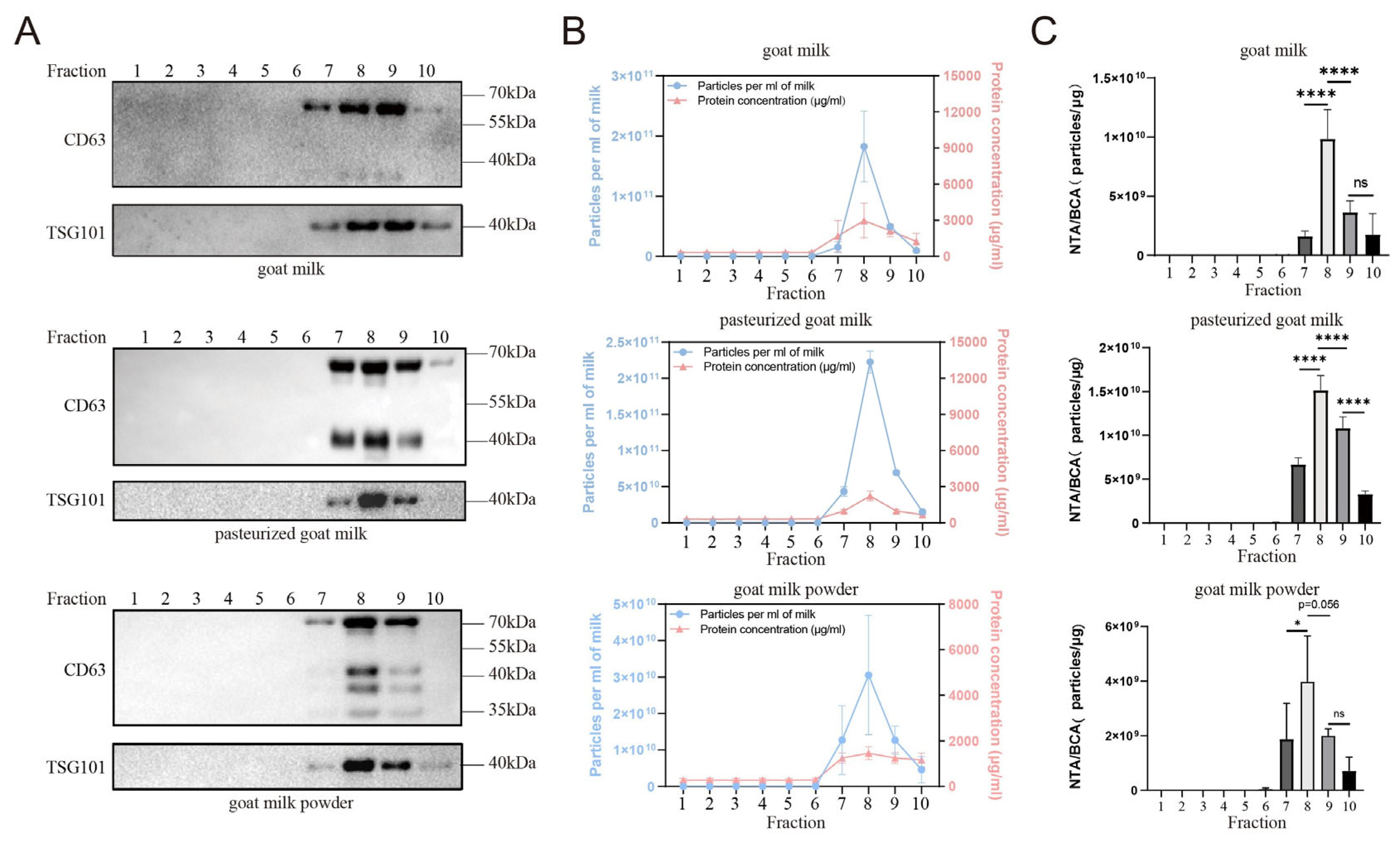
3.1. Extraction of sEVs from Goat Milk Samples
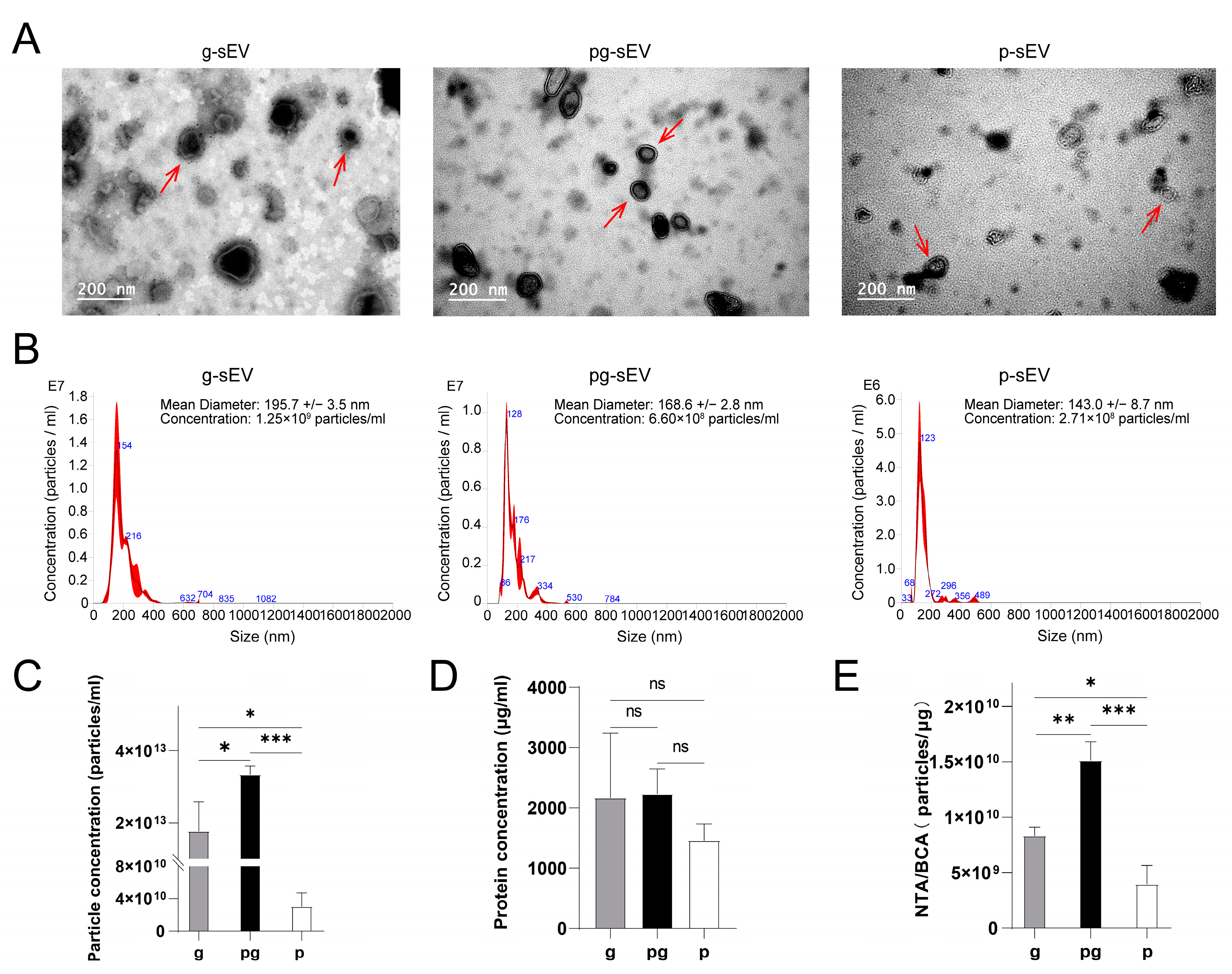
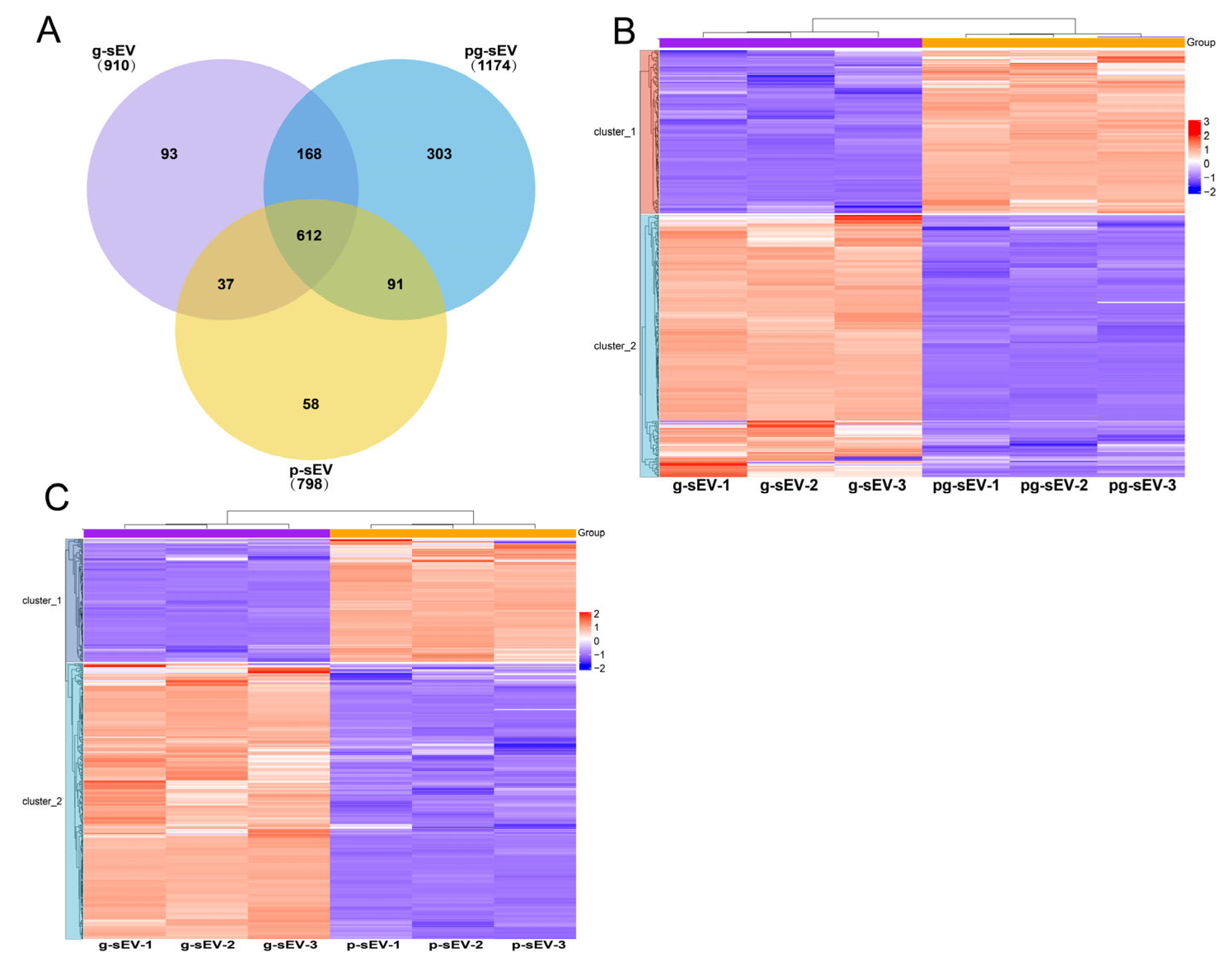
3.2. Identification and Comparison of sEVs from Goat Milk Products
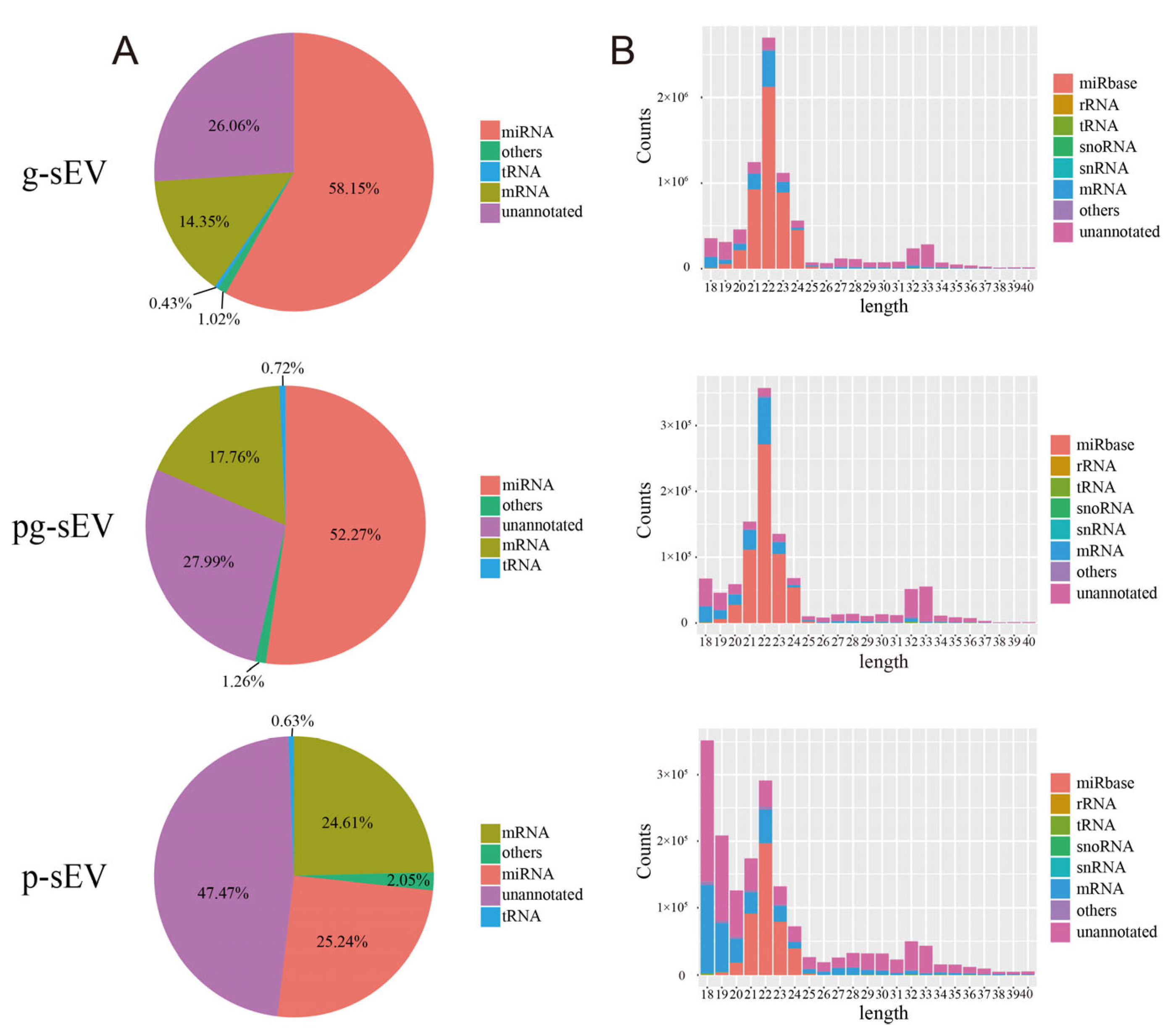
3.3. Comparison of Small RNA in sEVs of Goat Milk Products
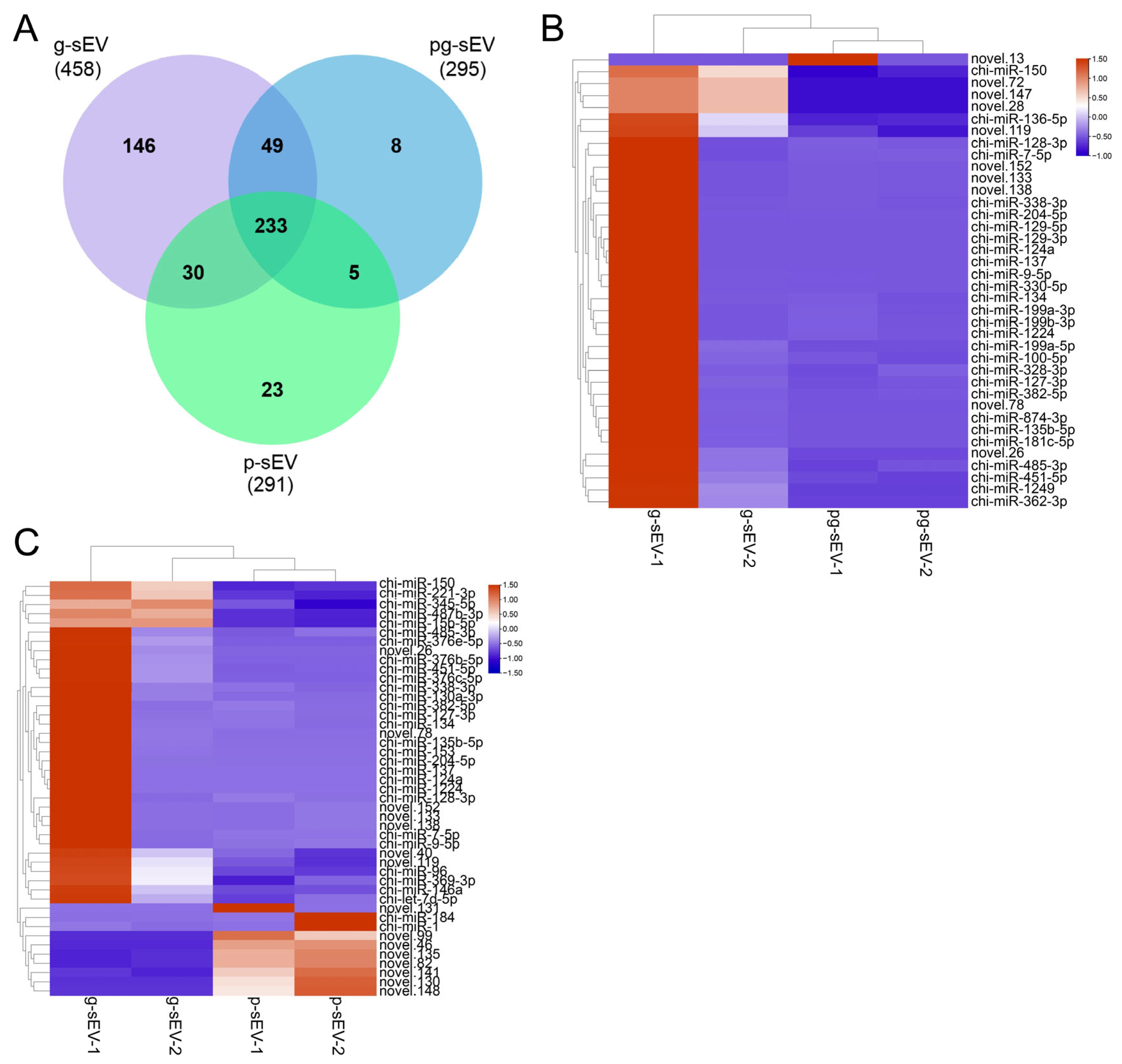
3.4. Comparison of miRNA Abundance in sEVs of Different Goat Milk Products
3.5. Function Analysis of the Top 10 Differentially Expressed miRNAs
3.6. Comparison of Small Extracellular Vesicle Protein in Different Goat Milk Products
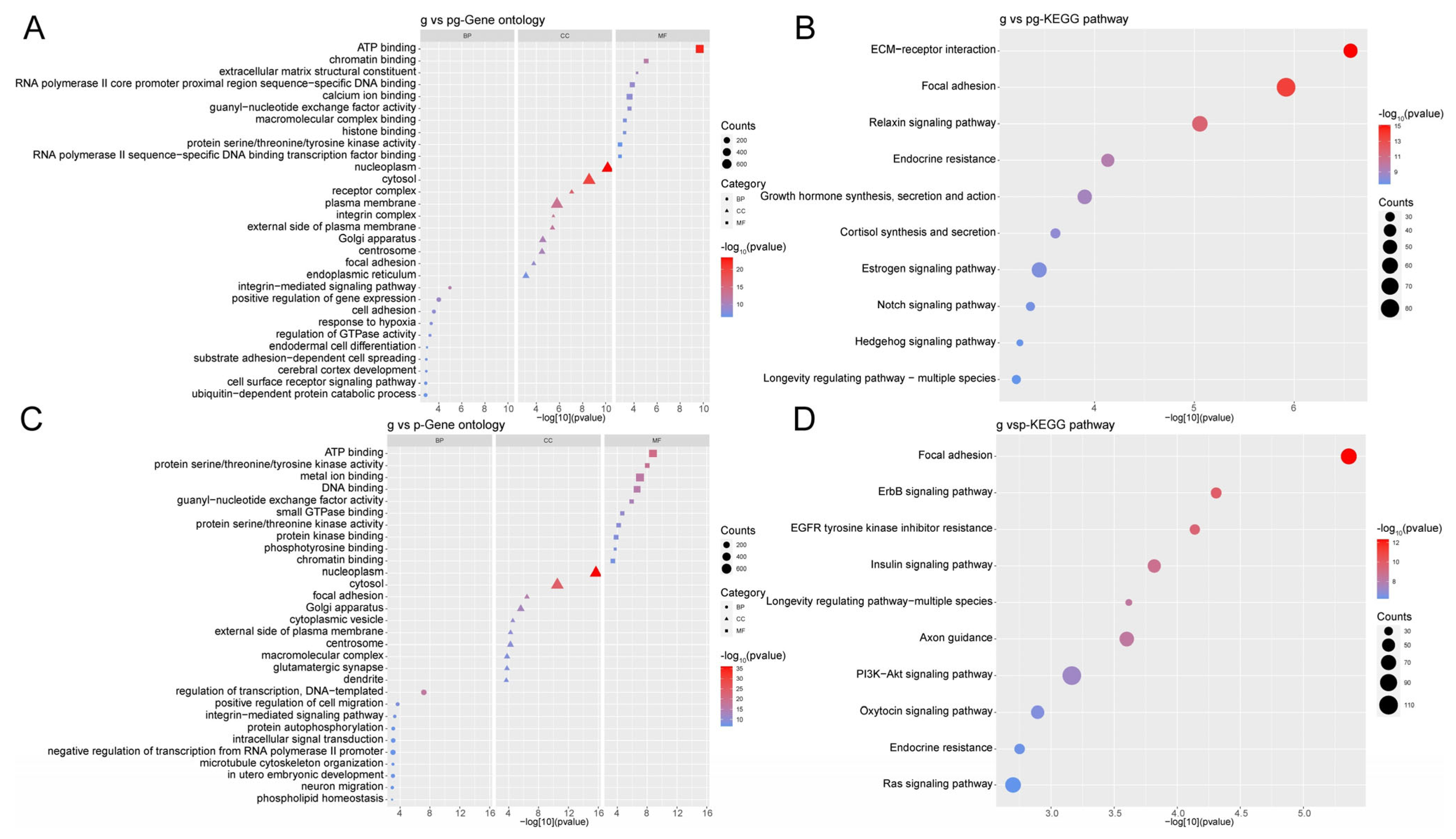
3.7. Functional Analysis of Differentially Expressed Proteins
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dalziel, J.E.; Dunstan, K.E.; Dewhurst, H.; Van Gendt, M.; Young, W.; Carpenter, E. Goat milk increases gastric emptying and alters caecal short chain fatty acid profile compared with cow milk in healthy rats. Food Funct. 2020, 11, 8573–8582. [Google Scholar] [CrossRef] [PubMed]
- Pereda, J.; Jaramillo, D.P.; Quevedo, J.M.; Ferragut, V.; Guamis, B.; Trujillo, A.J. Characterization of volatile compounds in ultra-high-pressure homogenized milk. Int. Dairy J. 2008, 18, 826–8345. [Google Scholar] [CrossRef]
- Fox, P.F.; Uniacke-Lowe, T.; Mcsweeney, P.; O’Mahony, J. Heat-Induced Changes in Milk; Springer International Publishing: Cham, Switzerland, 2015. [Google Scholar]
- Almiñana, C.; Corbin, E.; Tsikis, G.; Alcântara-Neto, A.S.; Labas, V.; Reynaud, K.; Galio, L.; Uzbekov, R.; Garanina, A.S.; Druart, X.; et al. Oviduct extracellular vesicles protein content and their role during oviduct-embryo cross-talk. Reproduction 2017, 154, 153–168. [Google Scholar] [CrossRef]
- Ambrosini, G.; Ebert, B.; Carvajal, R.D.; Schwartz, G.K.; Rai, A.J. Use of antibody arrays to probe exosome and extracellular vesicle mediated functional changes in cells. Methods Enzymol. 2020, 645, 43–53. [Google Scholar] [CrossRef]
- Li, X.; Su, L.; Zhang, X.; Chen, Q.; Wang, Y.; Shen, Z.; Zhong, T.; Wang, L.; Xiao, Y.; Feng, X.; et al. Recent Advances on the Function and Purification of Milk Exosomes: A Review. Front. Nutr. 2022, 9, 871346. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Zitvogel, L.; Amigorena, S. Exosomes: Composition, biogenesis and function. Nat. Rev. Immunol. 2002, 2, 569–579. [Google Scholar] [CrossRef]
- Benmoussa, A.; Lee, C.H.; Laffont, B.; Savard, P.; Laugier, J.; Boilard, E.; Gilbert, C.; Fliss, I.; Provost, P. Commercial Dairy Cow Milk microRNAs Resist Digestion under Simulated Gastrointestinal Tract Conditions. J. Nutr. 2016, 146, 2206–2215. [Google Scholar] [CrossRef]
- Liao, Y.; Du, X.; Li, J.; Lönnerdal, B. Human milk exosomes and their microRNAs survive digestion in vitro and are taken up by human intestinal cells. Mol. Nutr. Food Res. 2017, 61, 1700082. [Google Scholar] [CrossRef]
- Lönnerdal, B. Human Milk MicroRNAs/Exosomes: Composition and Biological Effects. Nestle Nutr. Inst. Workshop Ser. 2019, 90, 83–92. [Google Scholar] [CrossRef]
- Jiang, X.; You, L.; Zhang, Z.; Cui, X.; Zhong, H.; Sun, X.; Ji, C.; Chi, X. Biological Properties of Milk-Derived Extracellular Vesicles and Their Physiological Functions in Infant. Front. Cell Dev. Biol. 2021, 9, 693534. [Google Scholar] [CrossRef]
- Munagala, R.; Aqil, F.; Jeyabalan, J.; Gupta, R.C. Bovine milk-derived exosomes for drug delivery. Cancer Lett. 2016, 371, 48–61. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Li, M.; Wang, X.; Li, Q.; Wang, T.; Zhu, Q.; Zhou, X.; Wang, X.; Gao, X.; Li, X. Immune-related microRNAs are abundant in breast milk exosomes. Int. J. Biol. Sci. 2012, 8, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Admyre, C.; Johansson, S.M.; Qazi, K.R.; Filén, J.J.; Lahesmaa, R.; Norman, M.; Neve, E.P.; Scheynius, A.; Gabrielsson, S. Exosomes with immune modulatory features are present in human breast milk. J. Immunol. 2007, 179, 1969–1978. [Google Scholar] [CrossRef] [PubMed]
- Webber, J.; Clayton, A. How pure are your vesicles? J. Extracell. Vesicles 2013, 2, 19861. [Google Scholar] [CrossRef]
- Li, X.Y.; Cheng, M.; Li, J.; Zhao, X.; Qin, Y.S.; Chen, D.; Wang, J.M.; Wang, C.F. Change in the structural and functional properties of goat milk protein due to pH and heat. J. Dairy Sci. 2020, 103, 1337–1351. [Google Scholar] [CrossRef]
- González, M.I.; Gallardo, B.; Cerón, C.; Aguilera-Jiménez, E.; Cortes-Canteli, M.; Peinado, H.; Desco, M.; Salinas, B. Isolation of goat milk small extracellular vesicles by novel combined bio-physical methodology. Front. Bioeng. Biotechnol. 2023, 11, 1197780. [Google Scholar] [CrossRef]
- Izumi, H.; Kosaka, N.; Shimizu, T.; Sekine, K.; Ochiya, T.; Takase, M. Purification of RNA from milk whey. Methods Mol. Biol. 2013, 1024, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Kleinjan, M.; van Herwijnen, M.J.; Libregts, S.F.; van Neerven, R.J.; Feitsma, A.L.; Wauben, M.H. Regular Industrial Processing of Bovine Milk Impacts the Integrity and Molecular Composition of Extracellular Vesicles. J. Nutr. 2021, 151, 1416–1425. [Google Scholar] [CrossRef]
- Baddela, V.S.; Nayan, V.; Rani, P.; Onteru, S.K.; Singh, D. Physicochemical Biomolecular Insights into Buffalo Milk-Derived Nanovesicles. Appl. Biochem. Biotechnol. 2016, 178, 544–557. [Google Scholar] [CrossRef]
- Izumi, H.; Kosaka, N.; Shimizu, T.; Sekine, K.; Ochiya, T.; Takase, M. Bovine milk contains microRNA and messenger RNA that are stable under degradative conditions. J. Dairy Sci. 2012, 95, 4831–4841. [Google Scholar] [CrossRef]
- Smyczynska, U.; Bartlomiejczyk, M.A.; Stanczak, M.M.; Sztromwasser, P.; Wesolowska, A.; Barbarska, O.; Pawlikowska, E.; Fendler, W. Impact of processing method on donated human breast milk microRNA content. PLoS ONE 2020, 15, e0236126. [Google Scholar] [CrossRef]
- Kirchner, B.; Pfaffl, M.W.; Dumpler, J.; von Mutius, E.; Ege, M.J. microRNA in native and processed cow’s milk and its implication for the farm milk effect on asthma. J. Allergy Clin. Immunol. 2016, 137, 1893–1895.e13. [Google Scholar] [CrossRef]
- Howard, K.M.; Jati Kusuma, R.; Baier, S.R.; Friemel, T.; Markham, L.; Vanamala, J.; Zempleni, J. Loss of miRNAs during processing and storage of cow’s (Bos taurus) milk. J. Agric. Food Chem. 2015, 63, 588–592. [Google Scholar] [CrossRef] [PubMed]
- Kontopodi, E.; Boeren, S.; Stahl, B.; van Goudoever, J.B.; van Elburg, R.M.; Hettinga, K. High-Temperature Short-Time Preserves Human Milk’s Bioactive Proteins and Their Function Better Than Pasteurization Techniques With Long Processing Times. Front. Pediatr. 2021, 9, 798609. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Wu, S.; Zhang, K.; Xu, Z.; Quan, F. Goat milk exosomal microRNAs alleviate LPS-induced intestinal inflammation in mice. Int. J. Biol. Macromol. 2024, 268, 131698. [Google Scholar] [CrossRef] [PubMed]
- Shome, S.; Jernigan, R.L.; Beitz, D.C.; Clark, S.; Testroet, E.D. Non-coding RNA in raw and commercially processed milk and putative targets related to growth and immune-response. BMC Genom. 2021, 22, 749. [Google Scholar] [CrossRef]
- Na, R.S.; E, G.X.; Sun, W.; Sun, X.W.; Qiu, X.Y.; Chen, L.P.; Huang, Y.F. Expressional analysis of immune-related miRNAs in breast milk. Genet. Mol. Res. GMR 2015, 14, 11371–11376. [Google Scholar] [CrossRef]
- Zhao, X.; Ji, Z.; Xuan, R.; Wang, A.; Li, Q.; Zhao, Y.; Chao, T.; Wang, J. Characterization of the microRNA Expression Profiles in the Goat Kid Liver. Front. Genet. 2021, 12, 794157. [Google Scholar] [CrossRef]
- Manoharan, P.; Basford, J.E.; Pilcher-Roberts, R.; Neumann, J.; Hui, D.Y.; Lingrel, J.B. Reduced levels of microRNAs miR-124a and miR-150 are associated with increased proinflammatory mediator expression in Krüppel-like factor 2 (KLF2)-deficient macrophages. J. Biol. Chem. 2014, 289, 31638–31646. [Google Scholar] [CrossRef]
- Xiao, C.; Calado, D.P.; Galler, G.; Thai, T.H.; Patterson, H.C.; Wang, J.; Rajewsky, N.; Bender, T.P.; Rajewsky, K. MiR-150 controls B cell differentiation by targeting the transcription factor c-Myb. Cell 2007, 131, 146–159. [Google Scholar] [CrossRef]
- Bezman, N.A.; Chakraborty, T.; Bender, T.; Lanier, L.L. miR-150 regulates the development of NK and iNKT cells. J. Exp. Med. 2011, 208, 2717–2731. [Google Scholar] [CrossRef] [PubMed]
- Chapnick, D.A.; Warner, L.; Bernet, J.; Rao, T.; Liu, X. Partners in crime: The TGFβ and MAPK pathways in cancer progression. Cell Biosci. 2011, 1, 42. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Li, X.; Zhao, X.; Qin, Y.; Wang, J.; Wang, C. Comparative proteomics of goat milk during heated processing. Food Chem. 2019, 275, 504–514. [Google Scholar] [CrossRef] [PubMed]
- Brick, T.; Ege, M.; Boeren, S.; Böck, A.; von Mutius, E.; Vervoort, J.; Hettinga, K. Effect of Processing Intensity on Immunologically Active Bovine Milk Serum Proteins. Nutrients 2017, 9, 963. [Google Scholar] [CrossRef]
- Xiong, L.; Li, C.; Boeren, S.; Vervoort, J.; Hettinga, K. Effect of heat treatment on bacteriostatic activity and protein profile of bovine whey proteins. Food Res. Int. 2020, 127, 108688. [Google Scholar] [CrossRef]
- Wei, Z.; Kang, J.; Liao, M.; Ju, H.; Fan, R.; Shang, J.; Ning, X.; Li, M. Investigating changes of proteome in the bovine milk serum after retort processing using proteomics techniques. Food Sci. Nutr. 2022, 10, 307–316. [Google Scholar] [CrossRef]
- Meltretter, J.; Wüst, J.; Dittrich, D.; Lach, J.; Ludwig, J.; Eichler, J.; Pischetsrieder, M. Untargeted Proteomics-Based Profiling for the Identification of Novel Processing-Induced Protein Modifications in Milk. J. Proteome Res. 2020, 19, 805–818. [Google Scholar] [CrossRef]
- Milkovska-Stamenova, S.; Mnatsakanyan, R.; Hoffmann, R. Protein carbonylation sites in bovine raw milk and processed milk products. Food Chem. 2017, 229, 417–424. [Google Scholar] [CrossRef]
- Zou, Z.; Duley, J.A.; Cowley, D.M.; Reed, S.; Arachchige, B.J.; Bhandari, B.; Shaw, P.N.; Bansal, N. Physicochemical Properties and Whey Proteomes of Camel Milk Powders Produced by Different Concentration and Dehydration Processes. Foods 2022, 11, 727. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, W.; Han, B.; Zhang, L.; Zhou, P. Changes in bioactive milk serum proteins during milk powder processing. Food Chem. 2020, 314, 126177. [Google Scholar] [CrossRef]
- Colella, A.P.; Prakash, A.; Miklavcic, J.J. Homogenization and thermal processing reduce the concentration of extracellular vesicles in bovine milk. Food Sci. Nutr. 2024, 12, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Bionaz, M.; Loor, J.J. Gene networks driving bovine milk fat synthesis during the lactation cycle. BMC Genom. 2008, 9, 366. [Google Scholar] [CrossRef] [PubMed]
- Smith, I.A.; Knezevic, B.R.; Ammann, J.U.; Rhodes, D.A.; Aw, D.; Palmer, D.B.; Mather, I.H.; Trowsdale, J. BTN1A1, the mammary gland butyrophilin, and BTN2A2 are both inhibitors of T cell activation. J. Immunol. 2010, 184, 3514–3525. [Google Scholar] [CrossRef]
- Yang, Y.; Zheng, N.; Zhao, X.; Zhang, Y.; Han, R.; Ma, L.; Zhao, S.; Li, S.; Guo, T.; Wang, J. Proteomic characterization and comparison of mammalian milk fat globule proteomes by iTRAQ analysis. J. Proteom. 2015, 116, 34–43. [Google Scholar] [CrossRef]
- Collard, C.D.; Montalto, M.C.; Reenstra, W.R.; Buras, J.A.; Stahl, G.L. Endothelial oxidative stress activates the lectin complement pathway: Role of cytokeratin 1. Am. J. Pathol. 2001, 159, 1045–1054. [Google Scholar] [CrossRef]
- Satyam, A.; Graef, E.R.; Lapchak, P.H.; Tsokos, M.G.; Dalle Lucca, J.J.; Tsokos, G.C. Complement and coagulation cascades in trauma. Acute Med. Surg. 2019, 6, 329–335. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | g-sEV | pg-sEV | p-sEV |
|---|---|---|---|
| Diameters (nm) | 195.7 | 168.6 | 143.0 |
| Particle concentration (particles/mL) | 1.77 × 1013 | 3.34 × 1013 * | 3.06 × 1010 * |
| Particles per mL milk (particles) | 1.18 × 1011 | 2.2 × 1011 * | 1.53 × 108 *** |
| Protein concentration (μg/mL) | 2169.47 | 2231.41 ns | 1458.3 ns |
| NTA/BCA (particles/μg) | 8.34 × 109 | 1.51 × 1010 ** | 3.98 × 109 *** |
| Ordinal | g-sEV | pg-sEV | p-sEV |
|---|---|---|---|
| 1 | chi-let-7b-5p | chi-let-7f-5p | chi-let-7b-5p |
| 2 | chi-let-7f-5p | chi-let-7b-5p | chi-let-7f-5p |
| 3 | chi-miR-141 | chi-miR-16a-5p | chi-miR-148a-3p |
| 4 | chi-miR-29a-3p | chi-miR-141 | chi-miR-141 |
| 5 | chi-miR-16a-5p | chi-miR-223-3p | chi-miR-30a-5p |
| 6 | chi-miR-148a-3p | chi-miR-30a-5p | chi-miR-16a-5p |
| 7 | chi-miR-30a-5p | chi-miR-148a-3p | chi-miR-21-5p |
| 8 | chi-miR-26b-5p | chi-miR-146b-5p | chi-miR-29a-3p |
| 9 | chi-miR-223-3p | chi-miR-26b-5p | chi-miR-200a |
| 10 | chi-miR-146b-5p | chi-miR-29a-3p | chi-miR-103-3p |
| Ordinal | g-sEV | pg-sEV | p-sEV |
|---|---|---|---|
| 1 | BTN1A1 | BTN1A1 | BTN1A1 |
| 2 | CD36 | H-FABP | LALBA |
| 3 | XDH | CD36 | H-FABP |
| 4 | H-FABP | TSPAN1 | CD36 |
| 5 | MFGE8 | CSN2 | CD9 |
| 6 | MUC1 | ABCG2 | XDH |
| 7 | CD9 | MFGE8 | MUC1 |
| 8 | GLYCAM1 | GLYCAM1 | MFGE8 |
| 9 | SLC34A2 | MUC1 | SLC34A2 |
| 10 | ABCG2 | SLC34A2 | MUC15 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, Y.; Li, Z.; Hou, Y.; Tan, C.; Xiong, S.; Zhong, J.; Xie, Q. Effects of Different Processing on miRNA and Protein in Small Extracellular Vesicles of Goat Dairy Products. Nutrients 2024, 16, 4331. https://doi.org/10.3390/nu16244331
Fan Y, Li Z, Hou Y, Tan C, Xiong S, Zhong J, Xie Q. Effects of Different Processing on miRNA and Protein in Small Extracellular Vesicles of Goat Dairy Products. Nutrients. 2024; 16(24):4331. https://doi.org/10.3390/nu16244331
Chicago/Turabian StyleFan, Yuqin, Zhikang Li, Yanmei Hou, Chumin Tan, Sheng Xiong, Jinjing Zhong, and Qiuling Xie. 2024. "Effects of Different Processing on miRNA and Protein in Small Extracellular Vesicles of Goat Dairy Products" Nutrients 16, no. 24: 4331. https://doi.org/10.3390/nu16244331
APA StyleFan, Y., Li, Z., Hou, Y., Tan, C., Xiong, S., Zhong, J., & Xie, Q. (2024). Effects of Different Processing on miRNA and Protein in Small Extracellular Vesicles of Goat Dairy Products. Nutrients, 16(24), 4331. https://doi.org/10.3390/nu16244331