Effects of Lactiplantibacillus plantarum GUANKE on Diphenoxylate-Induced Slow Transit Constipation and Gut Microbiota in Mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
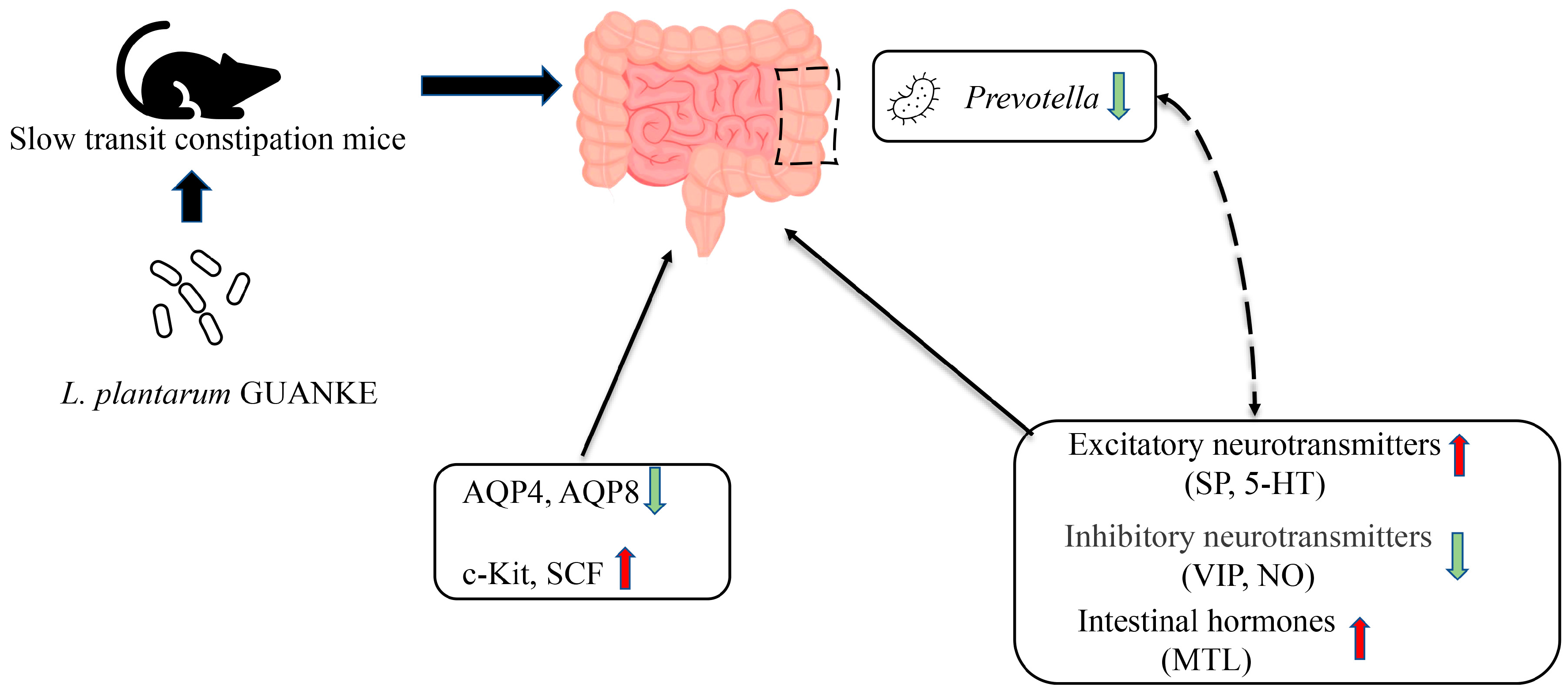
:1. Introduction
2. Materials and Methods
2.1. Animals and Strain
2.2. Establishment of Constipation Mouse Model and L. plantarum GUANK Intervention
2.3. Measurement of Intestinal Transit Ratio in Mice
2.4. Immunohistochemistry Examination of AQP4, AQP8, C-Kit, and SCF
2.5. Measurement of SCFAs
2.6. Quantification Measurement of Serum Neurotransmitters and Hormones
2.7. 16S rRNA Sequencing and Gut Microbiota Analysis
2.8. Statistical Analysis
3. Results
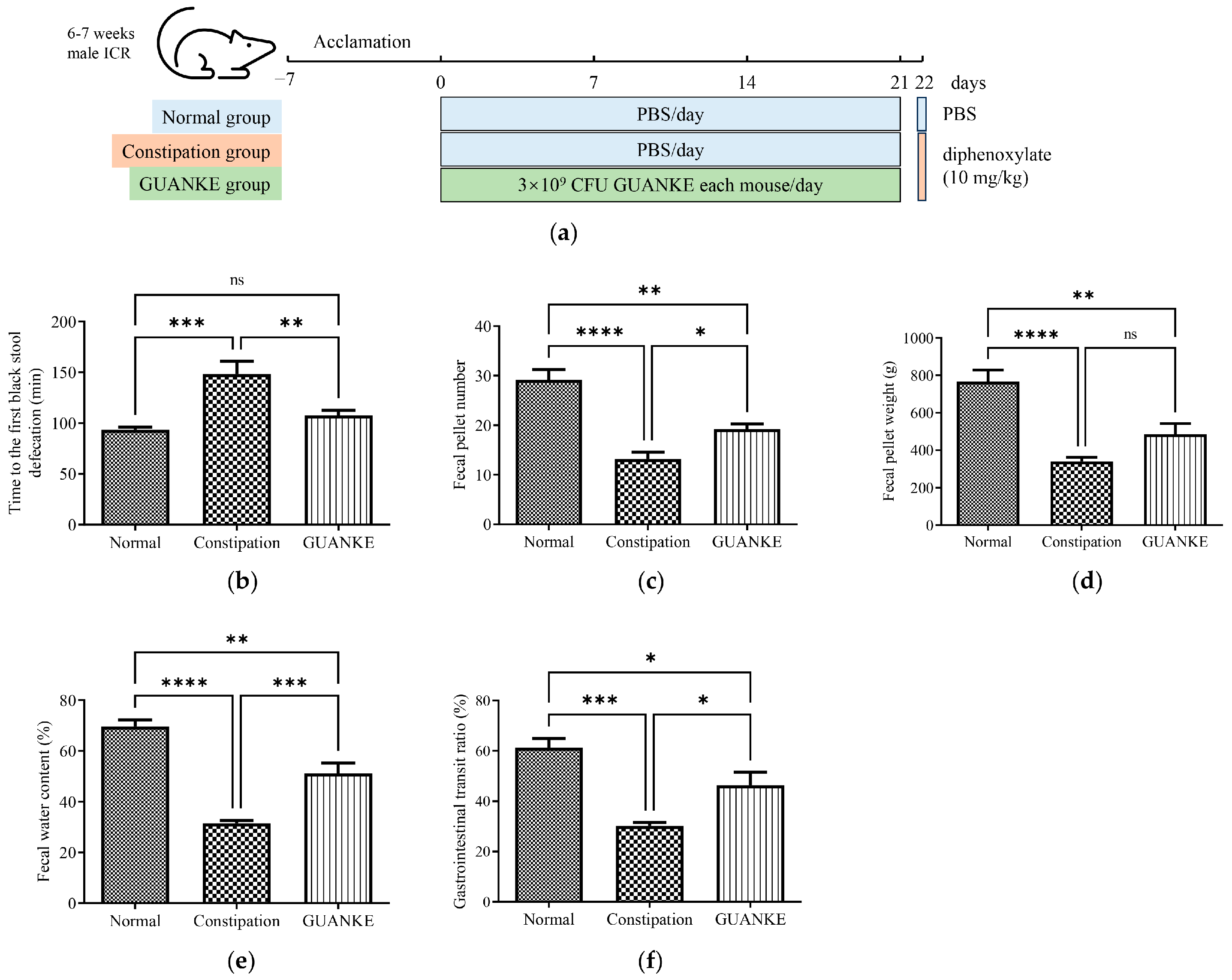
3.1. Effects of L. plantarum GUANKE on Fecal Parameters and Gastrointestinal Transit
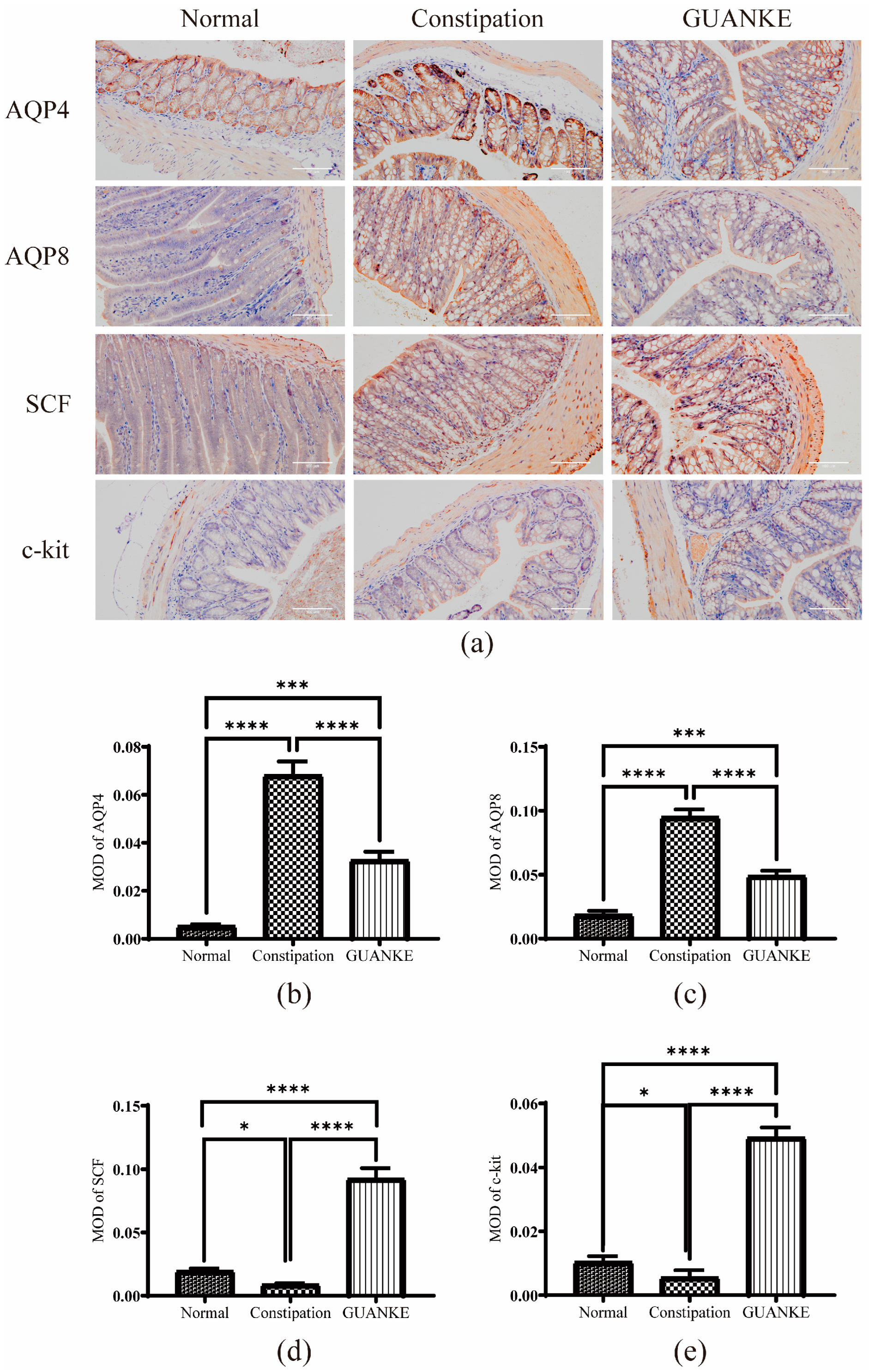
3.2. Effects of L. plantarum GUANKE on Protein Expression Levels of Constipation-Related Intestinal Factors
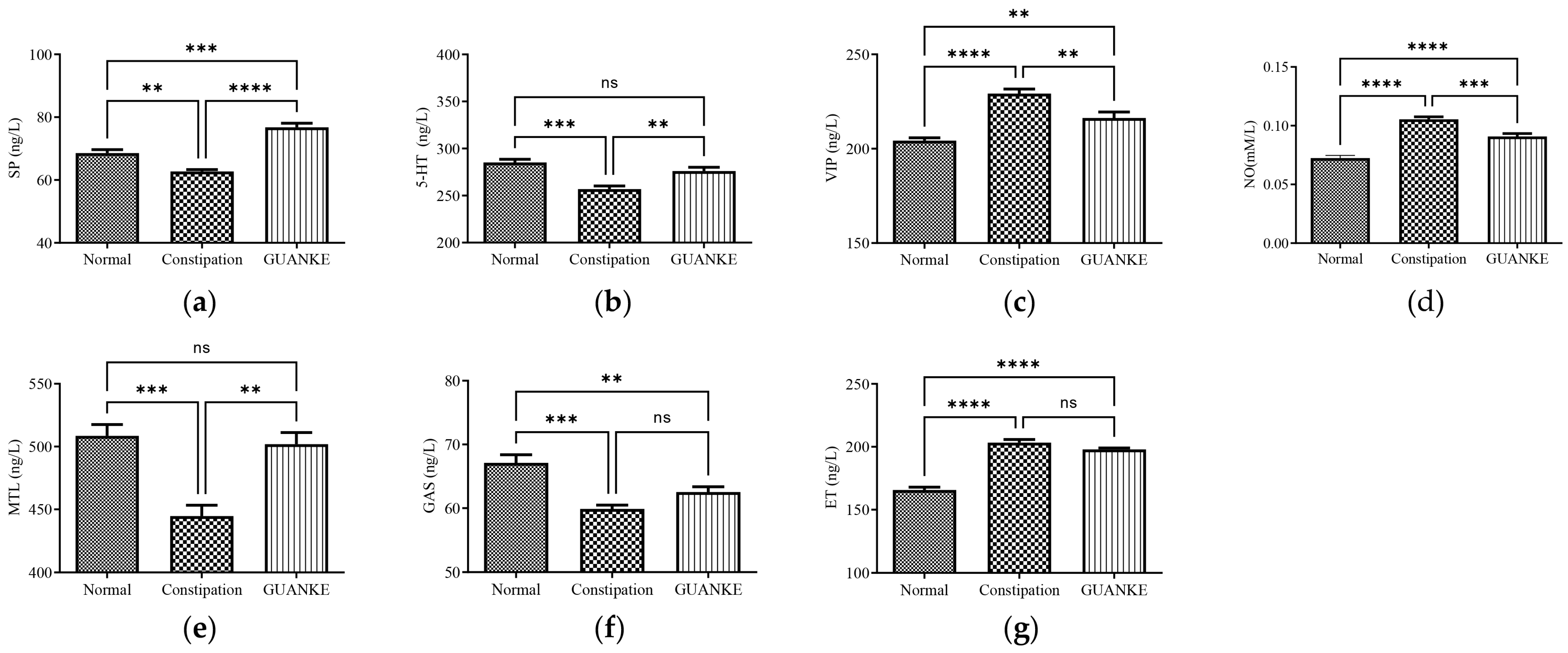
3.3. Effects of L. plantarum GUANKE on Serum Neurotransmitter and Hormone Levels
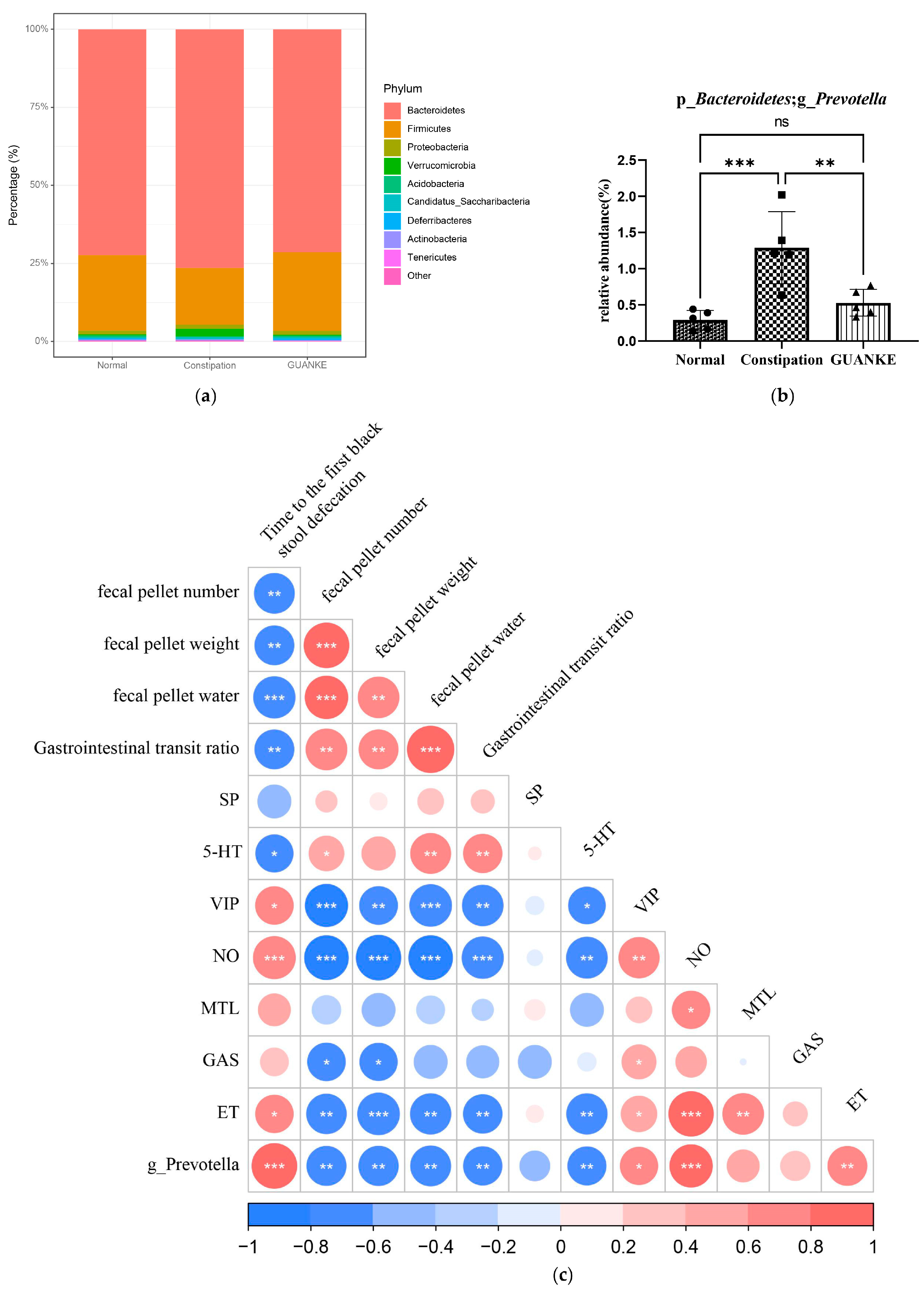
3.4. Effects of L. plantarum GUANKE on the Diversity of Intestinal Microbiota
3.5. Effects of L. plantarum GUANKE on SCFA Profile in Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wong, S.W.; Lubowski, D.Z. Slow-Transit Constipation: Evaluation and Treatment. ANZ J. Surg. 2007, 77, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Tillou, J.; Poylin, V. Functional Disorders: Slow-Transit Constipation. Clin. Colon Rectal Surg. 2017, 30, 76–86. [Google Scholar] [CrossRef]
- Sbahi, H.; Cash, B.D. Chronic Constipation: A Review of Current Literature. Curr. Gastroenterol. Rep. 2015, 17, 47. [Google Scholar] [CrossRef]
- Tian, H.; Chen, Q.; Yang, B.; Qin, H.; Li, N. Analysis of Gut Microbiome and Metabolite Characteristics in Patients with Slow Transit Constipation. Dig. Dis. Sci. 2021, 66, 3026–3035. [Google Scholar] [CrossRef] [PubMed]
- Aziz, I.; Whitehead, W.E.; Palsson, O.S.; Törnblom, H.; Simrén, M. An Approach to the Diagnosis and Management of Rome IV Functional Disorders of Chronic Constipation. Expert. Rev. Gastroenterol. Hepatol. 2020, 14, 39–46. [Google Scholar] [CrossRef]
- Black, C.J.; Ford, A.C. Chronic Idiopathic Constipation in Adults: Epidemiology, Pathophysiology, Diagnosis and Clinical Management. Med. J. Aust. 2018, 209, 86–91. [Google Scholar] [CrossRef]
- Tomita, R.; Tanjoh, K.; Fujisaki, S.; Ikeda, T.; Fukuzawa, M. Regulation of the Enteric Nervous System in the Colon of Patients with Slow Transit Constipation. Hepatogastroenterology 2002, 49, 1540–1544. [Google Scholar] [PubMed]
- Bassotti, G.; Villanacci, V.; Maurer, C.A.; Fisogni, S.; Di Fabio, F.; Cadei, M.; Morelli, A.; Panagiotis, T.; Cathomas, G.; Salerni, B. The Role of Glial Cells and Apoptosis of Enteric Neurones in the Neuropathology of Intractable Slow Transit Constipation. Gut 2006, 55, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Shang, F.; Morgan, M.J.; King, D.W.; Lubowski, D.Z.; Burcher, E. Cyclooxygenase-Dependent Alterations in Substance P-Mediated Contractility and Tachykinin NK1 Receptor Expression in the Colonic Circular Muscle of Patients with Slow Transit Constipation. J. Pharmacol. Exp. Ther. 2009, 329, 282–289. [Google Scholar] [CrossRef]
- Jung, H.-K.; Kim, D.-Y.; Moon, I.-H. Effects of Gender and Menstrual Cycle on Colonic Transit Time in Healthy Subjects. Korean J. Intern. Med. 2003, 18, 181–186. [Google Scholar] [CrossRef]
- Björnsson, E.S.; Chey, W.D.; Hooper, F.; Woods, M.L.; Owyang, C.; Hasler, W.L. Impaired Gastrocolonic Response and Peristaltic Reflex in Slow-Transit Constipation: Role of 5-HT(3) Pathways. Am. J. Physiol. Gastrointest. Liver Physiol. 2002, 283, G400–G407. [Google Scholar] [CrossRef]
- Dimidi, E.; Christodoulides, S.; Scott, S.M.; Whelan, K. Mechanisms of Action of Probiotics and the Gastrointestinal Microbiota on Gut Motility and Constipation. Adv. Nutr. 2017, 8, 484–494. [Google Scholar] [CrossRef]
- Bharucha, A.E.; Lacy, B.E. Mechanisms, Evaluation, and Management of Chronic Constipation. Gastroenterology 2020, 158, 1232–1249.e3. [Google Scholar] [CrossRef]
- Ge, X.; Zhao, W.; Ding, C.; Tian, H.; Xu, L.; Wang, H.; Ni, L.; Jiang, J.; Gong, J.; Zhu, W.; et al. Potential Role of Fecal Microbiota from Patients with Slow Transit Constipation in the Regulation of Gastrointestinal Motility. Sci. Rep. 2017, 7, 441. [Google Scholar] [CrossRef]
- Xu, M.; Wang, L.; Guo, Y.; Zhang, W.; Chen, Y.; Li, Y. Corrigendum to “Positive Effect of Electro-Acupuncture Treatment on Gut Motility in Constipated Mice Is Related to Rebalancing the Gut Microbiota”. Evid. Based Complement. Alternat Med. 2021, 2021, 9835654. [Google Scholar] [CrossRef] [PubMed]
- Vriesman, M.H.; Koppen, I.J.N.; Camilleri, M.; Di Lorenzo, C.; Benninga, M.A. Management of Functional Constipation in Children and Adults. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 21–39. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Song, L.; Huang, Y.; Li, X.; Xiao, Y.; Wang, Z.; Ren, Z. Latilactobacillus Sakei Furu2019 and Stachyose as Probiotics, Prebiotics, and Synbiotics Alleviate Constipation in Mice. Front. Nutr. 2022, 9, 1039403. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhang, Y.; Ma, K.; Wang, G.; Tang, M.; Wang, R.; Xia, Z.; Xu, Z.; Sun, M.; Bao, X.; et al. Lactobacillus Plantarum Lp3a Improves Functional Constipation: Evidence from a Human Randomized Clinical Trial and Animal Model. Ann. Transl. Med. 2022, 10, 316. [Google Scholar] [CrossRef]
- Ma, T.; Yang, N.; Xie, Y.; Li, Y.; Xiao, Q.; Li, Q.; Jin, H.; Zheng, L.; Sun, Z.; Zuo, K.; et al. Effect of the Probiotic Strain, Lactiplantibacillus Plantarum P9, on Chronic Constipation: A Randomized, Double-Blind, Placebo-Controlled Study. Pharmacol. Res. 2023, 191, 106755. [Google Scholar] [CrossRef]
- Basturk, A.; Artan, R.; Atalay, A.; Yilmaz, A. Investigation of the Efficacy of Synbiotics in the Treatment of Functional Constipation in Children: A Randomized Double-Blind Placebo-Controlled Study. Turk. J. Gastroenterol. 2017, 28, 388–393. [Google Scholar] [CrossRef]
- Krammer, H.-J.; Schlieger, F.; Harder, H.; Franke, A.; Singer, M.V. Probiotika in der Therapie des Reizdarmsyndroms. Z. Gastroenterol. 2005, 43, 467–471. [Google Scholar] [CrossRef]
- He, Y.; Zhu, L.; Chen, J.; Tang, X.; Pan, M.; Yuan, W.; Wang, H. Efficacy of Probiotic Compounds in Relieving Constipation and Their Colonization in Gut Microbiota. Molecules 2022, 27, 666. [Google Scholar] [CrossRef]
- Yi, R.; Zhou, X.; Liu, T.; Xue, R.; Yang, Z. Amelioration Effect of Lactobacillus Plantarum KFY02 on Low-Fiber Diet-Induced Constipation in Mice by Regulating Gut Microbiota. Front. Nutr. 2022, 9, 938869. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Zhang, Z.; Tong, L.; Zhou, X.; Liang, X.; Yi, H.; Gong, P.; Liu, T.; Zhang, L.; Yang, L.; et al. Mechanisms Underlying the Promotion of 5-hydroxytryptamine Secretion in Enterochromaffin Cells of Constipation Mice by Bifidobacterium and Lactobacillus. Neurogastroenterol. Motil. 2021, 33, e14082. [Google Scholar] [CrossRef]
- Chen, Z.; Feng, J.; Hu, S.; Hua, Y.; Ma, S.; Fu, W.; Yang, Q.; Zhang, X. Bacillus Subtilis Promotes the Release of 5-HT to Regulate Intestinal Peristalsis in STC Mice via Bile Acid and Its Receptor TGR5 Pathway. Dig. Dis. Sci. 2022, 67, 4410–4421. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Ren, Z.; Cao, K.; Li, X.; Yang, J.; Luo, X.; Zhu, L.; Wang, X.; Ding, L.; Liang, J.; et al. Boosting Vaccine-Elicited Respiratory Mucosal and Systemic COVID-19 Immunity in Mice with the Oral Lactobacillus Plantarum. Front. Nutr. 2021, 8, 789242. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Fu, Z.; Yan, W.; Nie, K.; Ding, L.; Ma, D.; Huang, H.; Li, T.; Xie, J.; Fu, L. The Different Effects of Chinese Herb Solid Drink and Lactulose on Gut Microbiota in Rats with Slow Transit Constipation Induced by Compound Diphenoxylate. Food Res. Int. 2021, 143, 110273. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME Improves Sensitivity and Speed of Chimera Detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Search and Clustering Orders of Magnitude Faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A Versatile Open Source Tool for Metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhong, D.; Sun, R.; Zhang, Y.; Pegg, R.B.; Zhong, G. Prevention of Loperamide Induced Constipation in Mice by KGM and the Mechanisms of Different Gastrointestinal Tract Microbiota Regulation. Carbohydr. Polym. 2021, 256, 117418. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Tang, J.; Hu, Y.; Zhang, W. Role of Gut Microbiota-Derived Signals in the Regulation of Gastrointestinal Motility. Front. Med. 2022, 9, 961703. [Google Scholar] [CrossRef]
- Jani, B.; Marsicano, E. Constipation: Evaluation and Management. Mo Med. 2018, 115, 236–240. [Google Scholar] [PubMed]
- Korterink, J.J.; Rutten, J.M.T.M.; Venmans, L.; Benninga, M.A.; Tabbers, M.M. Pharmacologic Treatment in Pediatric Functional Abdominal Pain Disorders: A Systematic Review. J. Pediatr. 2015, 166, 424–431.e6. [Google Scholar] [CrossRef] [PubMed]
- Kusumo, P.D.; Maulahela, H.; Utari, A.P.; Surono, I.S.; Soebandrio, A.; Abdullah, M. Probiotic Lactobacillus Plantarum IS 10506 Supplementation Increase SCFA of Women with Functional Constipation. Iran. J. Microbiol. 2019, 11, 389–396. [Google Scholar]
- Wang, L.; Hu, L.; Xu, Q.; Jiang, T.; Fang, S.; Wang, G.; Zhao, J.; Zhang, H.; Chen, W. Bifidobacteria Exert Species-Specific Effects on Constipation in BALB/c Mice. Food Funct. 2017, 8, 3587–3600. [Google Scholar] [CrossRef] [PubMed]
- Tan, A.H.; Lim, S.-Y.; Chong, K.K.; Manap, M.A.A.A.; Hor, J.W.; Lim, J.L.; Low, S.C.; Chong, C.W.; Mahadeva, S.; Lang, A.E. Probiotics for Constipation in Parkinson Disease: A Randomized Placebo-Controlled Study. Neurology 2021, 96, e772–e782. [Google Scholar] [CrossRef]
- Wang, G.; Yang, S.; Sun, S.; Si, Q.; Wang, L.; Zhang, Q.; Wu, G.; Zhao, J.; Zhang, H.; Chen, W. Lactobacillus Rhamnosus Strains Relieve Loperamide-Induced Constipation via Different Pathways Independent of Short-Chain Fatty Acids. Front. Cell Infect. Microbiol. 2020, 10, 423. [Google Scholar] [CrossRef]
- Göthert, M.; Bönisch, H.; Malinowska, B.; Schlicker, E. Serotonin Discovery and Stepwise Disclosure of 5-HT Receptor Complexity over Four Decades. Part II. Some Contributions of Manfred Göthert. Pharmacol. Rep. 2020, 72, 271–284. [Google Scholar] [CrossRef]
- Tsukamoto, A.; Ohno, K.; Maeda, S.; Nakashima, K.; Fukushima, K.; Fujino, Y.; Tsujimoto, H. Prokinetic Effect of the 5-HT4R Agonist Mosapride on Canine Gastric Motility. J. Vet. Med. Sci. 2011, 73, 1635–1637. [Google Scholar] [CrossRef]
- Yi, R.; Peng, P.; Zhang, J.; Du, M.; Lan, L.; Qian, Y.; Zhou, J.; Zhao, X. Lactobacillus plantarum CQPC02-Fermented Soybean Milk Improves Loperamide-Induced Constipation in Mice. J. Med. Food 2019, 22, 1208–1221. [Google Scholar] [CrossRef]
- Zhao, X.; Yi, R.; Qian, Y.; Park, K.-Y. Lactobacillus plantarum YS-3 Prevents Activated Carbon-Induced Constipation in Mice. J. Med. Food 2018, 21, 575–584. [Google Scholar] [CrossRef]
- Gan, Y.; Liang, J.; Diao, W.; Zhou, X.; Mu, J.; Pang, L.; Tan, F.; Zhao, X. Lactobacillus Plantarum KSFY06 and Geniposide Counteract Montmorillonite-Induced Constipation in Kunming Mice. Food Sci. Nutr. 2020, 8, 5128–5137. [Google Scholar] [CrossRef] [PubMed]
- Obata, Y.; Pachnis, V. The Effect of Microbiota and the Immune System on the Development and Organization of the Enteric Nervous System. Gastroenterology 2016, 151, 836–844. [Google Scholar] [CrossRef] [PubMed]
- Barbara, G.; Feinle-Bisset, C.; Ghoshal, U.C.; Quigley, E.M.; Santos, J.; Vanner, S.; Vergnolle, N.; Zoetendal, E.G. The Intestinal Microenvironment and Functional Gastrointestinal Disorders. Gastroenterology 2016, 150, 1305–1318. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yu, Y.-B. Intestinal Microbiota and Chronic Constipation. Springerplus 2016, 5, 1130. [Google Scholar] [CrossRef]
- Khalif, I.L.; Quigley, E.M.M.; Konovitch, E.A.; Maximova, I.D. Alterations in the Colonic Flora and Intestinal Permeability and Evidence of Immune Activation in Chronic Constipation. Dig. Liver Dis. 2005, 37, 838–849. [Google Scholar] [CrossRef]
- Choi, C.H.; Chang, S.K. Alteration of Gut Microbiota and Efficacy of Probiotics in Functional Constipation. J. Neurogastroenterol. Motil. 2015, 21, 4–7. [Google Scholar] [CrossRef]
- Jiang, J.-G.; Luo, Q.; Li, S.-S.; Tan, T.-Y.; Xiong, K.; Yang, T.; Xiao, T.-B. Cinnamic Acid Regulates the Intestinal Microbiome and Short-Chain Fatty Acids to Treat Slow Transit Constipation. World J. Gastrointest. Pharmacol. Ther. 2023, 14, 4–21. [Google Scholar] [CrossRef]
- Chu, C.; Yu, L.; Li, Y.; Guo, H.; Zhai, Q.; Chen, W.; Tian, F. Lactobacillus plantarum CCFM405 against Rotenone-Induced Parkinson’s Disease Mice via Regulating Gut Microbiota and Branched-Chain Amino Acids Biosynthesis. Nutrients 2023, 15, 1737. [Google Scholar] [CrossRef]
- Zhan, Y.; Wen, Y.; Du, L.-J.; Wang, X.-X.; Tang, S.-Y.; Kong, P.-F.; Huang, W.-G.; Tang, X.-G. Effects of Maren Pills on the Intestinal Microflora and Short-Chain Fatty Acid Profile in Drug-Induced Slow Transit Constipation Model Rats. Front. Pharmacol. 2022, 13, 804723. [Google Scholar] [CrossRef]
- Tett, A.; Pasolli, E.; Masetti, G.; Ercolini, D.; Segata, N. Prevotella Diversity, Niches and Interactions with the Human Host. Nat. Rev. Microbiol. 2021, 19, 585–599. [Google Scholar] [CrossRef]
- Yeoh, Y.K.; Sun, Y.; Ip, L.Y.T.; Wang, L.; Chan, F.K.L.; Miao, Y.; Ng, S.C. Prevotella Species in the Human Gut Is Primarily Comprised of Prevotella Copri, Prevotella Stercorea and Related Lineages. Sci. Rep. 2022, 12, 9055. [Google Scholar] [CrossRef] [PubMed]
- Maeda, Y.; Kurakawa, T.; Umemoto, E.; Motooka, D.; Ito, Y.; Gotoh, K.; Hirota, K.; Matsushita, M.; Furuta, Y.; Narazaki, M.; et al. Dysbiosis Contributes to Arthritis Development via Activation of Autoreactive T Cells in the Intestine. Arthritis Rheumatol. 2016, 68, 2646–2661. [Google Scholar] [CrossRef]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.-Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking Long-Term Dietary Patterns with Gut Microbial Enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef]
- De Filippis, F.; Pellegrini, N.; Vannini, L.; Jeffery, I.B.; La Storia, A.; Laghi, L.; Serrazanetti, D.I.; Di Cagno, R.; Ferrocino, I.; Lazzi, C.; et al. High-Level Adherence to a Mediterranean Diet Beneficially Impacts the Gut Microbiota and Associated Metabolome. Gut 2016, 65, 1812–1821. [Google Scholar] [CrossRef] [PubMed]
- Hjorth, M.F.; Roager, H.M.; Larsen, T.M.; Poulsen, S.K.; Licht, T.R.; Bahl, M.I.; Zohar, Y.; Astrup, A. Pre-Treatment Microbial Prevotella-to-Bacteroides Ratio, Determines Body Fat Loss Success during a 6-Month Randomized Controlled Diet Intervention. Int. J. Obes. 2018, 42, 580–583. [Google Scholar] [CrossRef]
- Hjorth, M.F.; Blædel, T.; Bendtsen, L.Q.; Lorenzen, J.K.; Holm, J.B.; Kiilerich, P.; Roager, H.M.; Kristiansen, K.; Larsen, L.H.; Astrup, A. Prevotella-to-Bacteroides Ratio Predicts Body Weight and Fat Loss Success on 24-Week Diets Varying in Macronutrient Composition and Dietary Fiber: Results from a Post-Hoc Analysis. Int. J. Obes. 2019, 43, 149–157. [Google Scholar] [CrossRef] [PubMed]
- De Vadder, F.; Kovatcheva-Datchary, P.; Zitoun, C.; Duchampt, A.; Bäckhed, F.; Mithieux, G. Microbiota-Produced Succinate Improves Glucose Homeostasis via Intestinal Gluconeogenesis. Cell Metab. 2016, 24, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, H.K.; Gudmundsdottir, V.; Nielsen, H.B.; Hyotylainen, T.; Nielsen, T.; Jensen, B.A.H.; Forslund, K.; Hildebrand, F.; Prifti, E.; Falony, G.; et al. Human Gut Microbes Impact Host Serum Metabolome and Insulin Sensitivity. Nature 2016, 535, 376–381. [Google Scholar] [CrossRef]
- Peng, Y.; Zeng, Y.; Zheng, T.; Xie, X.; Wu, J.; Fu, L.; Lu, F.; Zhang, L.; Chen, Y.; Liu, X.; et al. Effects of Tiaopi Xiezhuo Decoction on Constipation and Gut Dysbiosis in Patients with Peritoneal Dialysis. Pharm. Biol. 2023, 61, 531–540. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Y.; Guo, Y.; Li, X.; Xiao, Y.; Wang, Z.; Song, L.; Ren, Z. Effects of Lactiplantibacillus plantarum GUANKE on Diphenoxylate-Induced Slow Transit Constipation and Gut Microbiota in Mice. Nutrients 2023, 15, 3741. https://doi.org/10.3390/nu15173741
Huang Y, Guo Y, Li X, Xiao Y, Wang Z, Song L, Ren Z. Effects of Lactiplantibacillus plantarum GUANKE on Diphenoxylate-Induced Slow Transit Constipation and Gut Microbiota in Mice. Nutrients. 2023; 15(17):3741. https://doi.org/10.3390/nu15173741
Chicago/Turabian StyleHuang, Yuanming, Yanan Guo, Xianping Li, Yuchun Xiao, Zhihuan Wang, Liqiong Song, and Zhihong Ren. 2023. "Effects of Lactiplantibacillus plantarum GUANKE on Diphenoxylate-Induced Slow Transit Constipation and Gut Microbiota in Mice" Nutrients 15, no. 17: 3741. https://doi.org/10.3390/nu15173741
APA StyleHuang, Y., Guo, Y., Li, X., Xiao, Y., Wang, Z., Song, L., & Ren, Z. (2023). Effects of Lactiplantibacillus plantarum GUANKE on Diphenoxylate-Induced Slow Transit Constipation and Gut Microbiota in Mice. Nutrients, 15(17), 3741. https://doi.org/10.3390/nu15173741