



The Comparison of the Influence of Bisphenol A (BPA) and Its Analogue Bisphenol S (BPS) on the Enteric Nervous System of the Distal Colon in Mice


Abstract
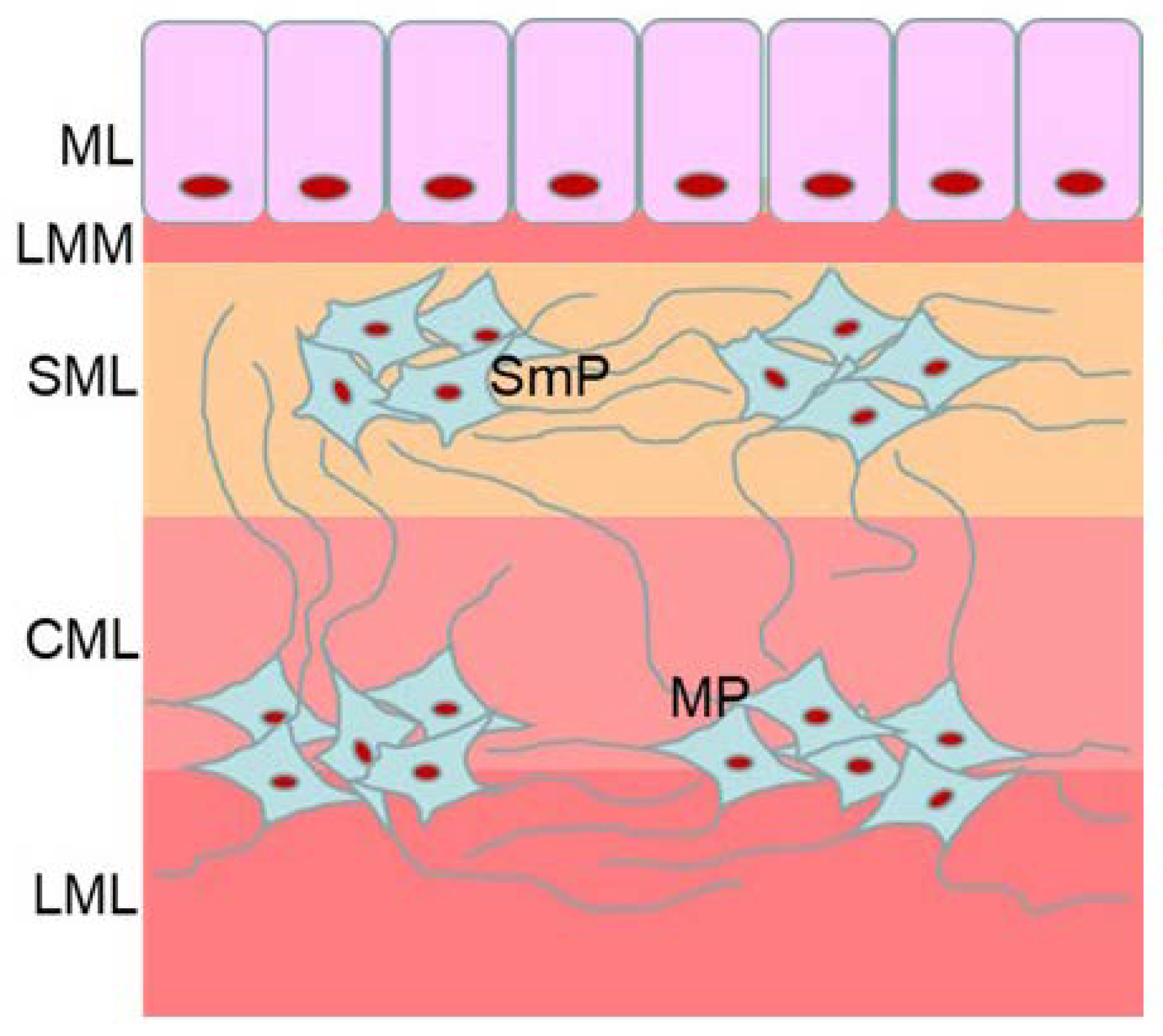
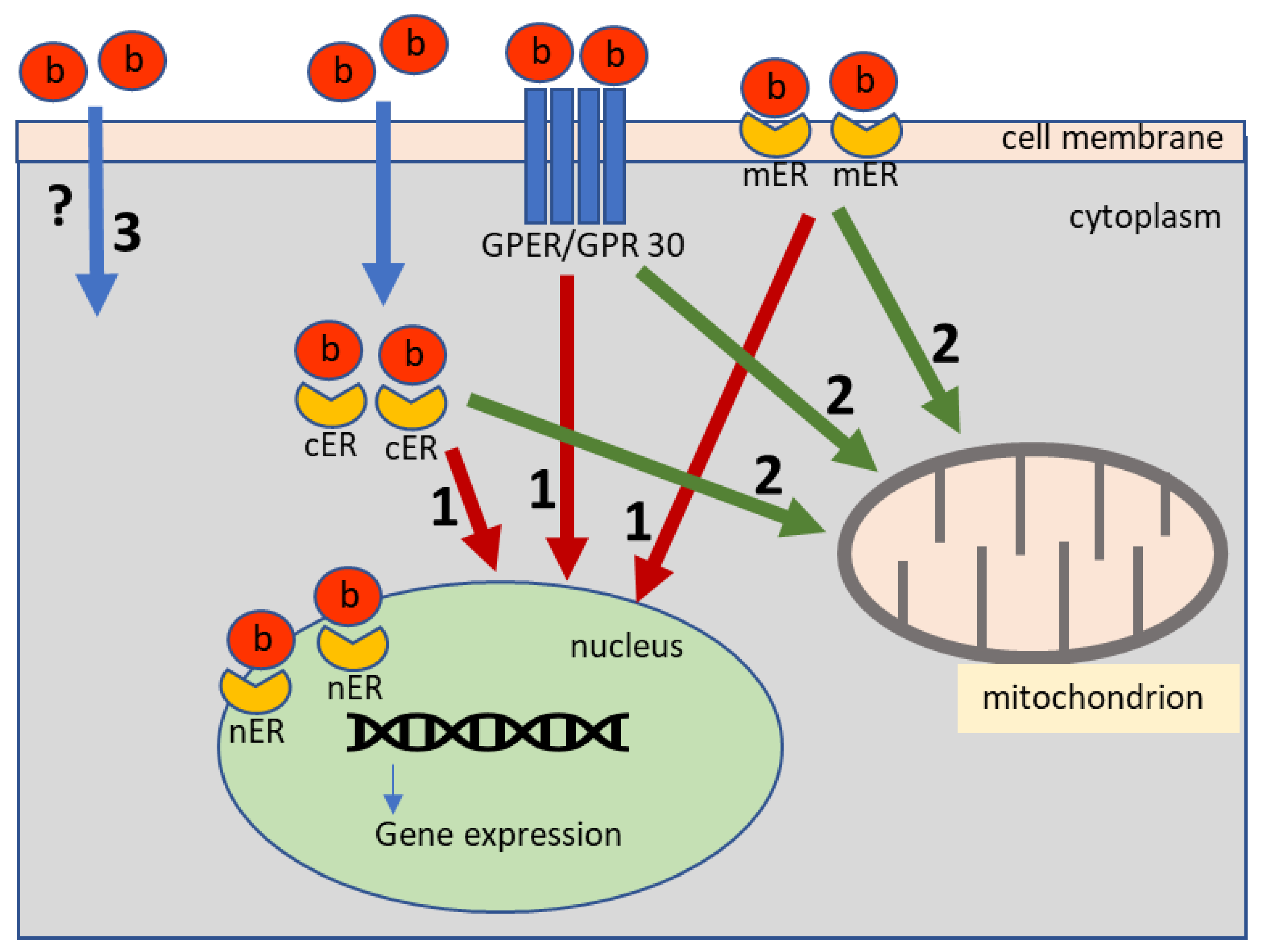
1. Introduction
2. Materials and Methods
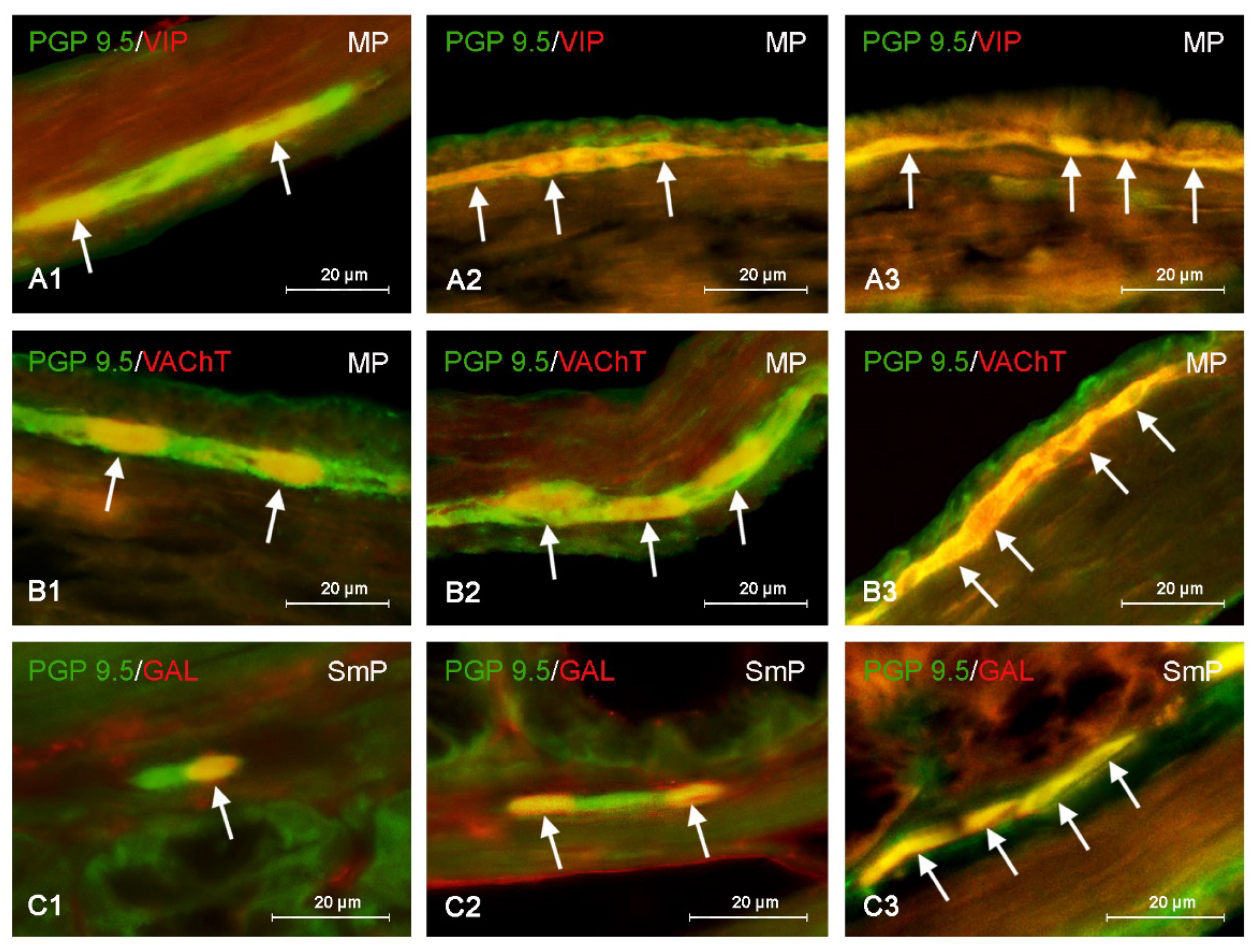
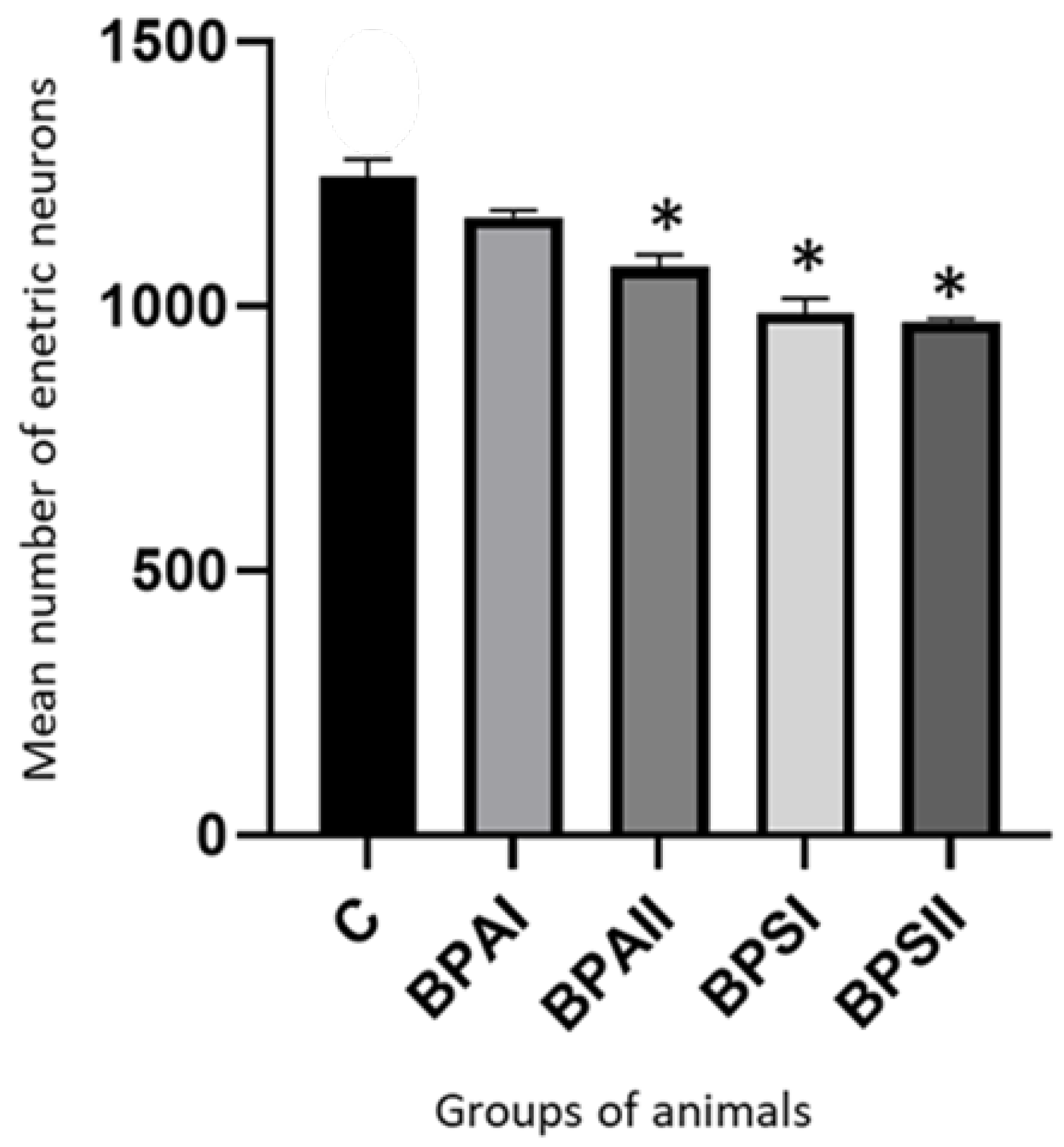
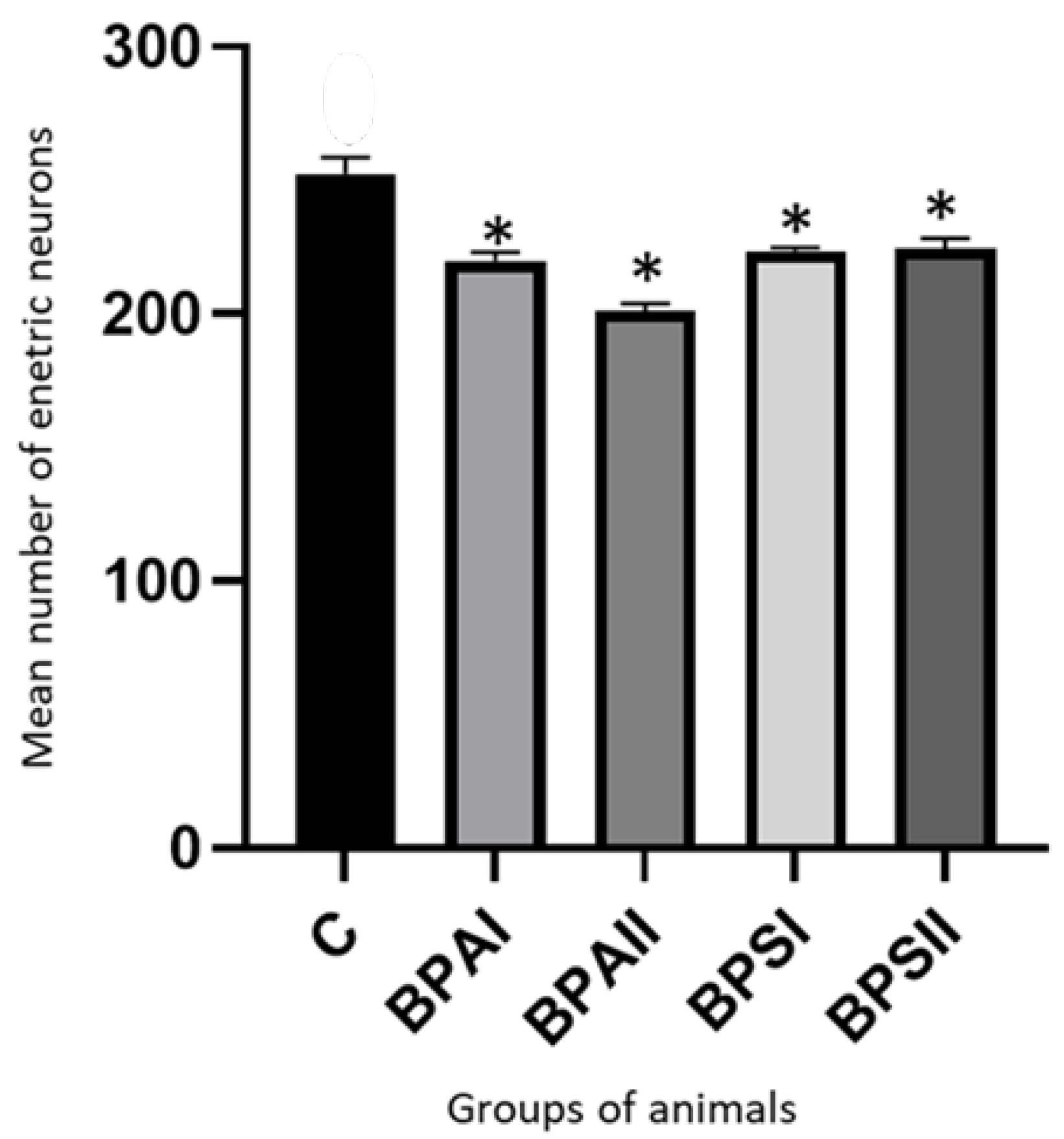
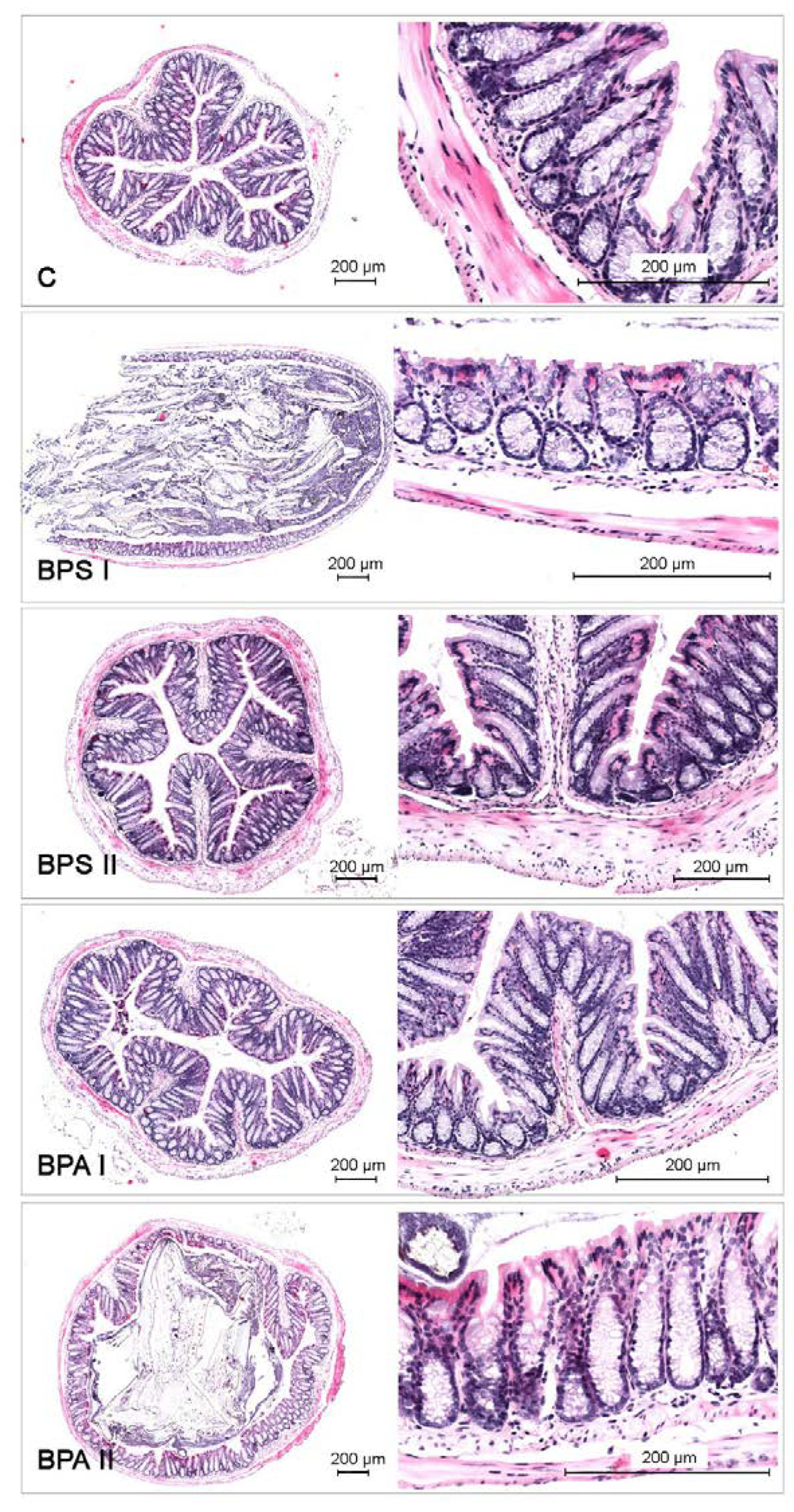
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vasiljevic, T.; Harner, T. Bisphenol A and its analogues in outdoor and indoor air: Properties, sources and global levels. Sci. Total Environ. 2021, 789, 148013. [Google Scholar] [CrossRef] [PubMed]
- Rudawska, A. Mechanical properties of epoxy compounds based on bisphenol A aged in aqueous environments. Polymers 2021, 13, 952. [Google Scholar] [CrossRef]
- Michałowicz, J. Bisphenol A—Sources, toxicity and biotransformation. Environ. Toxicol. Pharmacol. 2014, 37, 738–758. [Google Scholar] [CrossRef] [PubMed]
- Konieczna, A.; Rutkowska, A.; Rachoń, D. Health risk of exposure to Bisphenol A (BPA). Rocz. Panstw. Zakl. Hig. 2015, 66, 5–11. [Google Scholar]
- Cantoni, B.; Cappello Riguzzi, A.; Turolla, A.; Antonelli, M. Bisphenol A leaching from epoxy resins in the drinking water distribution networks as human health risk determinant. Sci. Total. Environ. 2021, 783, 146908. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, P.; Shappell, N.W.; Mukhopadhyay, M.; Onanong, S.; Rex, K.R.; Snow, D. Surveillance of plasticizers, bisphenol A, steroids and caffeine in surface water of River Ganga and Sundarban wetland along the Bay of Bengal: Occurrence, sources, estrogenicity screening and ecotoxicological risk assessment. Water Res. 2021, 190, 116668. [Google Scholar] [CrossRef] [PubMed]
- Gewurtz, S.B.; Tardif, G.; Power, M.; Backus, S.M.; Dove, A.; Dubé-Roberge, K.; Garron, C.; King, M.; Lalonde, B.; Letcher, R.J.; et al. Bisphenol A in the Canadian environment: A multimedia analysis. Sci. Total. Environ. 2021, 755, 142472. [Google Scholar] [CrossRef]
- Ahbab, M.A.; Barlas, N.; Karabulut, G. The toxicological effects of bisphenol A and octylphenol on the reproductive system of prepubertal male rats. Toxicol. Ind. Health. 2017, 33, 133–146. [Google Scholar] [CrossRef] [PubMed]
- MacLusky, N.J.; Hajszan, T.; Leranth, C. The environmental estrogen bisphenol A inhibits estradiol-induced hippocampal synaptogenesis. Environ. Health Perspect. 2005, 113, 675–679. [Google Scholar] [CrossRef]
- Engin, A.B.; Engin, A. The effect of environmental Bisphenol A exposure on breast cancer associated with obesity. Environ Toxicol Pharmacol. 2021, 81, 103544. [Google Scholar] [CrossRef]
- Pérez-Bermejo, M.; Mas-Pérez, I.; Murillo-Llorente, M.T. The role of the bisphenol A in diabetes and obesity. Biomedicines 2021, 9, 666. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhao, P.; Huang, Q.; Chi, Y.; Dong, S.; Fan, J. Bisphenol-A induces neurodegeneration through disturbance of intracellular calcium homeostasis in human embryonic stem cells-derived cortical neurons. Chemosphere 2019, 229, 618–630. [Google Scholar] [CrossRef] [PubMed]
- Salamanca-Fernández, E.; Rodríguez-Barranco, M.; Petrova, D.; Larrañaga, N.; Guevara, M.; Moreno-Iribas, C.; Chirlaque, M.D.; Colorado-Yohar, S.; Arrebola, J.P.; Vela, F.; et al. Bisphenol A exposure and risk of ischemic heart disease in the Spanish European Prospective Investigation into cancer and nutrition study. Chemosphere 2020, 261, 127697. [Google Scholar] [CrossRef] [PubMed]
- Seki, S.; Aoki, M.; Hosokawa, T.; Saito, T.; Masuma, R.; Komori, M.; Kurasaki, M. Bisphenol-A suppresses neu-rite extension due to inhibition of phosphorylation of mito-gen-activated protein kinase in PC12 cells. Chem. Biol. Interact. 2011, 194, 23–30. [Google Scholar] [CrossRef]
- Xu, X.; Xie, L.; Hong, X.; Ruan, Q.; Lu, H.; Zhang, Q.; Zhang, G.; Liu, X. Perinatal exposure to bisphenol-A inhibits synaptogenesis and affects the synaptic morphological development in offspring male mice. Chemosphere 2013, 91, 1073–1081. [Google Scholar] [CrossRef]
- Santoro, A.; Chianese, R.; Troisi, J.; Richards, S.; Nori, S.L.; Fasano, S.; Guida, M.; Plunk, E.; Viggiano, A.; Pierantoni, R.; et al. Neuro-toxic and reproductive effects of BPA. Curr. Neuropharmacol. 2019, 17, 1109–1132. [Google Scholar] [CrossRef]
- Viberg, H.; Lee, I. A single exposure to bisphenol A alters the levels of important neuroproteins in adult male and female mice. Neurotoxicology 2012, 33, 1390–1395. [Google Scholar] [CrossRef]
- Wang, W.; Wang, J.; Wang, Q.; Wu, W.; Huan, F.; Xiao, H. Bisphenol A modulates calcium currents and intracellular calcium concentration in rat dorsal root ganglion neurons. J. Membr. Biol. 2013, 246, 391–397. [Google Scholar] [CrossRef]
- Lee, S.; Suk, K.; Kim, I.K.; Jang, I.S.; Park, J.W.; Johnson, V.J.; Kwon, T.K.; Choi, B.J.; Kim, S.H. Signaling pathways of bisphenol A-induced apoptosis in hippocampal neuronal cells: Role of calcium-induced reactive oxygen species, mitogen-activated protein kinases, and nuclear factor-kappaB. J. Neurosci. Res. 2008, 86, 2932–2942. [Google Scholar] [CrossRef]
- Ni, Y.; Hu, L.; Yang, S.; Ni, L.; Ma, L.; Zhao, Y.; Zheng, A.; Jin, Y.; Fu, Z. Bisphenol A impairs cognitive function and 5-HT metabolism in adult male mice by modulating the microbiota-gut-brain axis. Chemosphere. 2021, 282, 130952. [Google Scholar] [CrossRef]
- Huang, B.; Jiang, C.; Luo, J.; Cui, Y.; Qin, L.; Liu, J. Maternal exposure to bisphenol A may increase the risks of Parkinson’s disease through down-regulation of fetal IGF-1 expression. Med. Hypotheses. 2014, 82, 245–249. [Google Scholar] [CrossRef]
- Sukjamnong, S.; Thongkorn, S.; Kanlayaprasit, S.; Saeliw, T.; Hussem, K.; Warayanon, W.; Hu, V.W.; Tencomnao, T.; Sarachana, T. Prenatal exposure to bisphenol A alters the transcriptome-interactome profiles of genes associated with Alzheimer’s disease in the offspring hippocampus. Sci. Rep. 2020, 10, 9487. [Google Scholar] [CrossRef] [PubMed]
- Frankowski, R.; Zgoła-Grześkowiak, A.; Grześkowiak, T.; Sójka, K. The presence of bisphenol A in the thermal paper in the face of changing European—A comparative global research. Environ. Pollut. 2020, 265, 114879. [Google Scholar] [CrossRef] [PubMed]
- Bousoumah, R.; Leso, V.; Iavicoli, I.; Huuskonen, P.; Viegas, S.; Porras, S.P.; Santonen, T.; Frery, N.; Robert, A.; Ndaw, S. Biomonitoring of occupational exposure to bisphenol A, bisphenol S and bisphenol F: A systematic review. Sci. Total. Environ. 2021, 783, 146905. [Google Scholar] [CrossRef] [PubMed]
- Fouyet, S.; Olivier, E.; Leproux, P.; Dutot, M.; Rat, P. Bisphenol A, bisphenol F, and bisphenol S: The bad and the ugly. Where is the good? Life 2021, 11, 314. [Google Scholar] [CrossRef]
- Marroqui, L.; Martinez-Pinna, J.; Castellano-Muñoz, M.; Dos Santos, R.S.; Medina-Gali, R.M.; Soriano, S.; Quesada, I.; Gustafsson, J.A.; Encinar, J.A.; Nadal, A. Bisphenol-S and bisphenol-F alter mouse pancreatic β-cell ion channel expression and activity and insulin release through an estrogen receptor ERβ mediated pathway. Chemosphere 2021, 265, 129051. [Google Scholar] [CrossRef]
- An, H.; Yu, H.; Wei, Y.; Liu, F.; Ye, J. Disrupted metabolic pathways and potential human diseases induced by bisphenol S. Environ. Toxicol. Pharmacol. 2021, 88, 103751. [Google Scholar] [CrossRef]
- Thoene, M.; Dzika, E.; Gonkowski, S.; Wojtkiewicz, J. Bisphenol S in food causes hormonal and obesogenic effects comparable to or worse than bisphenol A: A literature review. Nutrients 2020, 12, 532. [Google Scholar] [CrossRef]
- Parathan, P.; Wang, Y.; Leembruggen, A.J.; Bornstein, J.C.; Foong, J.P. The enteric nervous system undergoes significant chemical and synaptic maturation during adolescence in mice. Dev. Biol. 2020, 458, 75–87. [Google Scholar] [CrossRef]
- Szymanska, K.; Makowska, K.; Gonkowski, S. The influence of high and low doses of bisphenol A (BPA) on the enteric nervous system of the porcine ileum. Int. J. Mol. Sci. 2018, 19, 917. [Google Scholar] [CrossRef]
- Furness, J.B.; Callaghan, B.P.; Rivera, L.R.; Cho, H.J. The enteric nervous system and gastrointestinal innervation: Integrated local and central control. Adv. Exp. Med. Biol. 2014, 817, 39–71. [Google Scholar] [PubMed]
- Szymanska, K.; Gonkowski, S. Bisphenol A-induced changes in the enteric nervous system of the porcine duodenum. Neurotoxicology 2018, 66, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Szymanska, K.; Gonkowski, S. Neurochemical characterization of the enteric neurons within the porcine jejunum in physiological conditions and under the influence of bisphenol A (BPA). Neurogastroenterol. Motil. 2019, 31, e13580. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.Y.H.; Lin, G.; Fang, M.; Rudd, J.A. Localization of estrogen receptor Erα, Erβ and GPR30 on myenteric neurons of the gastrointestinal tract and their role in motility. Gen. Comp. Endocrinol. 2019, 272, 63–75. [Google Scholar] [CrossRef]
- D’Errico, F.; Goverse, G.; Dai, Y.; Wu, W.; Stakenborg, M.; Labeeuw, E.; De Simone, V.; Verstockt, B.; Gomez-Pinilla, P.J.; Warner, M.; et al. Estrogen receptor β controls proliferation of enteric glia and differentiation of neurons in the myenteric plexus after damage. Proc. Natl. Acad. Sci. USA 2018, 115, 5798–5803. [Google Scholar] [CrossRef]
- Bustamante-Barrientos, F.A.; Méndez-Ruette, M.; Ortloff, A.; Luz-Crawford, P.; Rivera, F.J.; Figueroa, C.D.; Molina, L.; Bátiz, L.F. The Impact of Estrogen and Estrogen-Like Molecules in Neurogenesis and Neurodegeneration: Beneficial or Harmful? Front. Cell. Neurosci. 2021, 15, 636176. [Google Scholar] [CrossRef]
- Campbell-Thompson, M.; Reyher, K.K.; Wilkinson, L.B. Immunolocalization of estrogen receptor alpha and beta in gastric epithelium and enteric neurons. J. Endocrinol. 2001, 171, 65–73. [Google Scholar] [CrossRef]
- Balasuriya, G.K.; Nugapitiya, S.S.; Hill-Yardin, E.L.; Bornstein, J.C. Nitric oxide regulates estrus cycle dependent colonic motility in mice. Front. Neurosci. 2021, 15, 647555. [Google Scholar] [CrossRef]
- Li, Y.; Xu, J.; Jiang, F.; Jiang, Z.; Liu, C.; Li, L.; Luo, Y.; Lu, R.; Mu, Y.; Liu, Y.; et al. G protein-coupled estrogen receptor is involved in modulating colonic motor function via nitric oxide release in C57BL/6 female mice. Neurogastroenterol. Motil. 2016, 28, 432–442. [Google Scholar] [CrossRef]
- Sakamoto, H.; Yokota, H.; Kibe, R.; Sayama, Y.; Yuasa, A. Excretion of bisphenol A-glucuronide into the small intestine and deconjugation in the cecum of the rat. Biochim. Biophys. Acta 2002, 1573, 171–176. [Google Scholar] [CrossRef]
- Peillex, C.; Kerever, A.; Lachhab, A.; Pelletier, M. Bisphenol A, bisphenol S and their glucuronidated metabolites modulate glycolysis and functional responses of human neutrophils. Environ. Res. 2021, 196, 110336. [Google Scholar] [CrossRef] [PubMed]
- de Silva, P.S.; Yang, X.; Korzenik, J.R.; Goldman, R.H.; Arheart, K.L.; Caban-Martinez, A.J. Association of urinary phenolic compounds, inflammatory bowel disease and chronic diarrheal symptoms: Evidence from the National Health and Nutrition Examination Survey. Environ. Pollut. 2017, 229, 621–626. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; He, H.; Wan, H.; Shen, N.; Li, J.; Zhang, S.; Zeng, Q.; Chang, J.; Lu, Q.; Zhong, R.; et al. Bisphenol A exposure, interaction with genetic variants and colorectal cancer via mediating oxidative stress biomarkers. Environ. Pollut. 2021, 287, 117630. [Google Scholar] [CrossRef]
- Godlewski, J.; Kmiec, Z. Colorectal cancer invasion and atrophy of the enteric nervous system: Potential feedback and impact on cancer progression. Int. J. Mol. Sci. 2020, 21, 3391. [Google Scholar] [CrossRef]
- Holland, A.M.; Bon-Frauches, A.C.; Keszthelyi, D.; Melotte, V.; Boesmans, W. The enteric nervous system in gastrointestinal disease etiology. Cell. Mol. Life. Sci. 2021, 78, 4713–4733. [Google Scholar] [CrossRef] [PubMed]
- Dobrzynska, M.M.; Gajowik, A.; Jankowska-Steifer, E.A.; Radzikowska, J.; Tyrkiel, E.J. Reproductive and developmental F1 toxicity following exposure of pubescent F0 male mice to bisphenol A alone and in a combination with X-rays irradiation. Toxicology 2018, 410, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Rezg, R.; Abot, A.; Mornagui, B.; Aydi, S.; Knauf, C. Effects of Bisphenol S on hypothalamic neuropeptides regulating feeding behavior and apelin/APJ system in mice. Ecotoxicol. Environ. Saf. 2018, 161, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Tyl, R.W.; Myers, C.B.; Marr, M.C.; Thomas, B.F.; Keimowitz, A.R.; Brine, D.R.; Veselica, M.M.; Fail, P.A.; Chang, T.Y.; Seely, J.C.; et al. Three-generation reproductive toxicity study of dietary bisphenol A in CD Sprague-Dawley rats. Toxicol. Sci. 2002, 68, 121–146. [Google Scholar] [CrossRef]
- Choi, C.W.; Jeong, J.; Hwang, M.S.; Jung, K.K.; Lee, K.H.; Lee, H.M. Establishment of the korean tolerable daily intake of bisphenol a based on risk assessments by an expert committee. Toxicol. Res. 2010, 26, 285–291. [Google Scholar] [CrossRef]
- Makowska, K.; Gonkowski, S. Bisphenol A (BPA) affects the enteric nervous system in the porcine stomach. Animals 2020, 10, 2445. [Google Scholar] [CrossRef]
- Makowska, K.; Szymańska, K.; Całka, J.; Gonkowski, S. The influence of bisphenol A (BPA) on the occurrence of selected active substances in neuregulin 1 (NRG1)-positive enteric neurons in the porcine large intestine. Int. J. Mol. Sci. 2021, 22, 10308. [Google Scholar] [CrossRef] [PubMed]
- Nezami, B.G.; Srinivasan, S. Enteric nervous system in the small intestine: Pathophysiology and clinical implications. Curr. Gastroenterol. Rep. 2010, 12, 358–365. [Google Scholar] [CrossRef] [PubMed]
- Velarde, E.; Alonso-Gómez, A.L.; Azpeleta, C.; Isorna, E.; Delgado, M.J. Melatonin attenuates the acetylcholine-induced contraction in isolated intestine of a teleost fish. J. Comp. Physiol. B. 2009, 179, 951–959. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.; Luo, H.; Quan, X.; Fan, H.; Tang, Q.; Yu, G.; Chen, W.; Xia, H. The role of substance P in the maintenance of colonic hypermotility induced by repeated stress in rats. Neuropeptides 2016, 56, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Norton, C.E.; Grunz-Borgmann, E.A.; Hart, M.L.; Jones, B.W.; Franklin, C.L.; Boerman, E.M. Role of perivascular nerve and sensory neurotransmitter dysfunction in inflammatory bowel disease. Am. J. Physiol. Heart. Circ. Physiol. 2021, 320, H1887–H1902. [Google Scholar] [CrossRef]
- The, F.; Cailotto, C.; van der Vliet, J.; de Jonge, W.J.; Bennink, R.J.; Buijs, R.M.; Boeckxstaens, G.E. Central activation of the cholinergic anti-inflammatory pathway reduces surgical inflammation in experimental post-operative ileus. Br. J. Pharmacol. 2011, 163, 1007–1016. [Google Scholar] [CrossRef]
- Van Geldre, L.A.; Lefebvre, R.A. Interaction of NO and VIP in gastrointestinal smooth muscle relaxation. Curr. Pharm. Des. 2004, 10, 2483–2497. [Google Scholar] [CrossRef]
- Bohlen, H.G. Rapid and slow nitric oxide responses during conducted vasodilation in the in vivo intestine and brain cortex microvasculatures. Microcirculation 2011, 18, 623–634. [Google Scholar] [CrossRef][Green Version]
- Currò, D.; Preziosi, P. Non-adrenergic non-cholinergic relaxation of the rat stomach. Gen. Pharmacol. 1998, 31, 697–703. [Google Scholar] [CrossRef]
- Brzozowska, M.; Całka, J. Review: Occurrence and distribution of galanin in the physiological and inflammatory states in the mammalian gastrointestinal tract. Front Immunol. 2021, 11, 602070. [Google Scholar] [CrossRef]
- Koller, A.; Bianchini, R.; Schlager, S.; Münz, C.; Kofler, B.; Wiesmayr, S. The neuropeptide galanin modulates natural killer cell function. Neuropeptides 2017, 64, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Brenneman, D.E.; Philips, T.M.; Hauser, J.; Hill, J.M.; Spong, C.Y.; Gozes, I. Complex array of cytokines released by vasoactive intestinal peptide. Neuropeptides 2003, 37, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Sanders, K.M.; Ward, S.M. Nitric oxide and its role as a non-adrenergic, non-cholinergic inhibitory neurotransmitter in the gastrointestinal tract. Br. J. Pharmacol. 2019, 176, 212–227. [Google Scholar] [CrossRef] [PubMed]
- Aimi, Y.; Kimura, H.; Kinoshita, T.; Minami, Y.; Fujimura, M.; Vincent, S.R. Histochemical localization of nitric oxide synthase in rat enteric nervous system. Neuroscience 1993, 53, 553–560. [Google Scholar] [CrossRef]
- Porter, A.J.; Wattchow, D.A.; Brookes, S.J.; Costa, M. Cholinergic and nitrergic interneurones in the myenteric plexus of the human colon. Gut 2002, 51, 70–75. [Google Scholar] [CrossRef]
- Furness, J.B. Types of neurons in the enteric nervous system. J. Auton. Nerv. Syst. 2000, 81, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Pimont, S.; Bruley Des Varannes, S.; Le Neel, J.C.; Aubert, P.; Galmiche, J.P.; Neunlist, M. Neurochemical coding of myenteric neurones in the human gastric fundus. Neurogastroenterol. Motil. 2003, 15, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Cserni, T.; Paran, S.; Puri, P. The effect of age on colocalization of acetylcholinesterase and nicotinamide adenine dinucleotide phosphate diaphorase staining in enteric neurons in an experimental model. J. Pediatr. Surg. 2007, 42, 300–304. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, K.; Tarafder, P.; Paul, G. Bisphenol A inhibits duodenal movement ex vivo of rat through nitric oxide-mediated soluble guanylyl cyclase and α-adrenergic signaling pathways. J. Appl. Toxicol. 2016, 36, 131–139. [Google Scholar] [CrossRef]
- Song, H.; Park, J.; Buim, P.T.C.; Choi, K.; Gye, M.C.; Hong, Y.C.; Kim, J.H.; Lee, Y.J. Bisphenol A induces COX-2 through the mitogen-activated protein kinase pathway and is associated with levels of inflammation-related markers in elderly populations. Environ. Res. 2017, 158, 490–498. [Google Scholar] [CrossRef]
- Sugita-Konishi, Y.; Shimura, S.; Nishikawa, T.; Sunaga, F.; Naito, H.; Suzuki, Y. Effect of Bisphenol A on non-specific immunodefenses against non-pathogenic Escherichia coli. Toxicol. Lett. 2003, 136, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Ge, M.; Jin, J.; Xu, H.; Mao, L.; Geng, S.; Wu, J.; Zhu, J.; Li, X.; Zhong, C. Mechanism investigation on Bisphenol S-induced oxidative stress and inflammation in murine RAW264.7 cells: The role of NLRP3 inflammasome, TLR4, Nrf2 and MAPK. J. Hazard. Mater. 2020, 394, 122549. [Google Scholar] [CrossRef] [PubMed]
- Mantyh, C.R.; Vigna, S.R.; Maggio, J.E.; Mantyh, P.W.; Bollinger, R.R.; Pappas, T.N. Substance P binding sites on intestinal lymphoid aggregates and blood vessels in inflammatory bowel disease correspond to authentic NK-1 receptors. Neurosci. Lett. 1994, 178, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Vota, D.; Aguero, M.; Grasso, E.; Hauk, V.; Gallino, L.; Soczewski, E.; Pérez Leirós, C.; Ramhorst, R. Progesterone and VIP cross-talk enhances phagocytosis and anti-inflammatory profile in trophoblast-derived cells. Mol. Cell Endocrinol. 2017, 443, 146–154. [Google Scholar] [CrossRef]
- Su, Y.; Ganea, D.; Peng, X.; Jonakait, G.M. Galanin down-regulates microglial tumor necrosis factor-alpha production by a post-transcriptional mechanism. J. Neuroimmunol. 2003, 134, 52–60. [Google Scholar] [CrossRef]
- Nakamura, K.; Itoh, K.; Sugimoto, T.; Fushiki, S. Prenatal exposure to bisphenol A affects adult murine neocortical structure. Neurosci. Lett. 2007, 420, 100–105. [Google Scholar] [CrossRef]
- Lai, K.P.; Chung, Y.T.; Li, R.; Wan, H.T.; Wong, C.K. Bisphenol A alters gut microbiome: Comparative metagenomics analysis. Environ. Pollut. 2016, 218, 923–930. [Google Scholar] [CrossRef]
- Krause, J.L.; Engelmann, B.; Nunes da Rocha, U.; Pierzchalski, A.; Chang, H.D.; Zenclussen, A.C.; von Bergen, M.; Rolle-Kampczyk, U.; Herberth, G. MAIT cell activation is reduced by direct and microbiota-mediated exposure to bisphenols. Environ. Int. 2022, 158, 106985. [Google Scholar] [CrossRef]
- Vicentini, F.A.; Keenan, C.M.; Wallace, L.E.; Woods, C.; Cavin, J.B.; Flockton, A.R.; Macklin, W.B.; Belkind-Gerson, J.; Hirota, S.A.; Sharkey, K.A. Intestinal microbiota shapes gut physiology and regulates enteric neurons and glia. Microbiome 2021, 9, 210. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary Antibodies | ||||
|---|---|---|---|---|
| Antigen | Code | Species | Working Dilution | Supplier |
| PGP 9.5 | 7863-2004 | Mouse | 1:1000 | Biogenesis Ltd., Poole, UK |
| SP | 8450-0505 | Rat | 1:1000 | Bio-Rad (AbD Serotec), Kidlington, UK |
| VIP | VA 1285 | Rabbit | 1:1000 | Enzo Life Sciences; Farmingdale, NY, USA |
| GAL | T-5036 | Guinea Pig | 1:1000 | Peninsula |
| VAChT | H-V006 | Rabbit | 1:2000 | Phoenix Pharmaceuticals |
| nNOS | AB5380 | Rabbit | 1:1000 | MercMillipore, DEU |
| Secondary antibodies | ||||
| Reagents | Working dilution | Supplier | ||
| Alexa fluor 488 donkey anti-mouse IgG | 1:1000 | Invitrogen, Carlsbad, CA, USA | ||
| Alexa fluor 546 donkey anti-rabbit IgG | 1:1000 | Invitrogen | ||
| Alexa fluor 546 donkey anti-rat IgG | 1:1000 | Invitrogen | ||
| Alexa fluor 546 donkey anti-guinea pig IgG | 1:1000 | Invitrogen | ||
| C | BPAI | BPAII | BPSI | BPSII | ||
|---|---|---|---|---|---|---|
| nNOS | MP | 38.83 ± 1.49% | 48.42 ± 3.83% * | 52.17 ± 4.05% *B | 50.05 ± 2.9% * | 68.98 ± 2.63% *B |
| SmP | 20.25 ± 0.97% | 26.09 ± 1.48% * | 36.98 ± 3.41% * | 29.63 ± 1.68% * | 34.65 ± 3.5% * | |
| GAL | MP | 17.53 ± 1.95% | 29.38 ± 2.83% * | 44.18 ± 3.09% * | 33.37 ± 1.45% * | 49.98 ± 1.89% * |
| SmP | 13.00 ± 0.73% | 21.35 ± 2.62% * | 31.53 ± 3.02% *B | 21.98 ± 1.26% * | 49.00 ± 1.95% *B | |
| VIP | MP | 37.14 ± 1.68% | 50.39 ± 1.69% *A | 64.86 ± 1.49% *B | 69.74 ± 3.36% *A | 74.01 ± 1.82% *B |
| SmP | 28.64 ± 1.13% | 42.89 ± 2.29% *A | 60.78 ± 3.12% * | 53.12 ± 5.47% *A | 55.75 ± 4.85% * | |
| SP | MP | 31.02 ± 1.49% | 43.5 ± 1.83% *A | 53.21 ± 4.14% * | 48.01 ± 2.86% *A | 53.23 ± 2.99% * |
| SmP | 13.20 ± 1.05% | 21.11 ± 2.75% * | 31.40 ± 3.41% * | 20.52 ± 1.69% * | 31.24 ± 4.39% * | |
| VAChT | MP | 48.55 ± 3.66% | 60.12 ± 1.34% * | 66.67 ± 1.07% * | 63.09 ± 2.36% * | 69.09 ± 1.31% * |
| SmP | 21.64 ± 1.38% | 41.85 ± 1.39% * | 69.44 ± 1.91% *B | 43.88 ± 4.83% * | 56.17 ± 2.9% *B |
| C | BPA I | BPA II | BPS I | BPS II | |
|---|---|---|---|---|---|
| Animal weight on 1 day of the experiment | 22.14 ± 0.74 | 22.71 ± 1.04 | 21.29 ± 1.27 | 22.43 ± 0.92 | 19.43 ± 1.46 |
| Animal weight on 91 day of the experiment | 24.14 ± 0.80 | 24.71 ± 1.11 | 23.57 ± 1.51 | 24.86 ± 0.83 | 21.86 ± 1.34 |
| Weight gain | 1.71 ± 0.29 | 2.00 ± 0.49 | 2.29 ± 0.52 | 2.43 ± 0.48 | 2.57 ± 0.48 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Makowska, K.; Lepiarczyk, E.; Gonkowski, S. The Comparison of the Influence of Bisphenol A (BPA) and Its Analogue Bisphenol S (BPS) on the Enteric Nervous System of the Distal Colon in Mice. Nutrients 2023, 15, 200. https://doi.org/10.3390/nu15010200
Makowska K, Lepiarczyk E, Gonkowski S. The Comparison of the Influence of Bisphenol A (BPA) and Its Analogue Bisphenol S (BPS) on the Enteric Nervous System of the Distal Colon in Mice. Nutrients. 2023; 15(1):200. https://doi.org/10.3390/nu15010200
Chicago/Turabian StyleMakowska, Krystyna, Ewa Lepiarczyk, and Slawomir Gonkowski. 2023. "The Comparison of the Influence of Bisphenol A (BPA) and Its Analogue Bisphenol S (BPS) on the Enteric Nervous System of the Distal Colon in Mice" Nutrients 15, no. 1: 200. https://doi.org/10.3390/nu15010200
APA StyleMakowska, K., Lepiarczyk, E., & Gonkowski, S. (2023). The Comparison of the Influence of Bisphenol A (BPA) and Its Analogue Bisphenol S (BPS) on the Enteric Nervous System of the Distal Colon in Mice. Nutrients, 15(1), 200. https://doi.org/10.3390/nu15010200