
The Protective Effect of Theaflavins on the Kidney of Mice with Type II Diabetes Mellitus

 and
and 
Abstract

1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals
2.3. Oral Glucose Tolerance Test (OGTT) and Insulin Tolerance Test (ITT)
2.4. ELISA
2.5. Biochemical Analysis
2.6. Histological Examination
2.7. Total AGEs Assay
2.8. mRNA Extraction and Quantitative Real-Time-PCR (qPCR) Analysis
2.9. Western Blot Analysis
2.10. Statistical Analysis
3. Results
3.1. Effects of TFs on Fasting Blood Glucose, Body Weight, Food Consumption, Water Consumption, and Urine Volume in T2DM Mice
3.2. Effects of TFs on Glucose and Insulin Homeostasis in T2DM Mice
3.3. TFs Intervention Improved Kidney Function and Structure in T2DM Mice
3.4. TFs Intervention Reduced the Level of AGEs in T2DM Mice
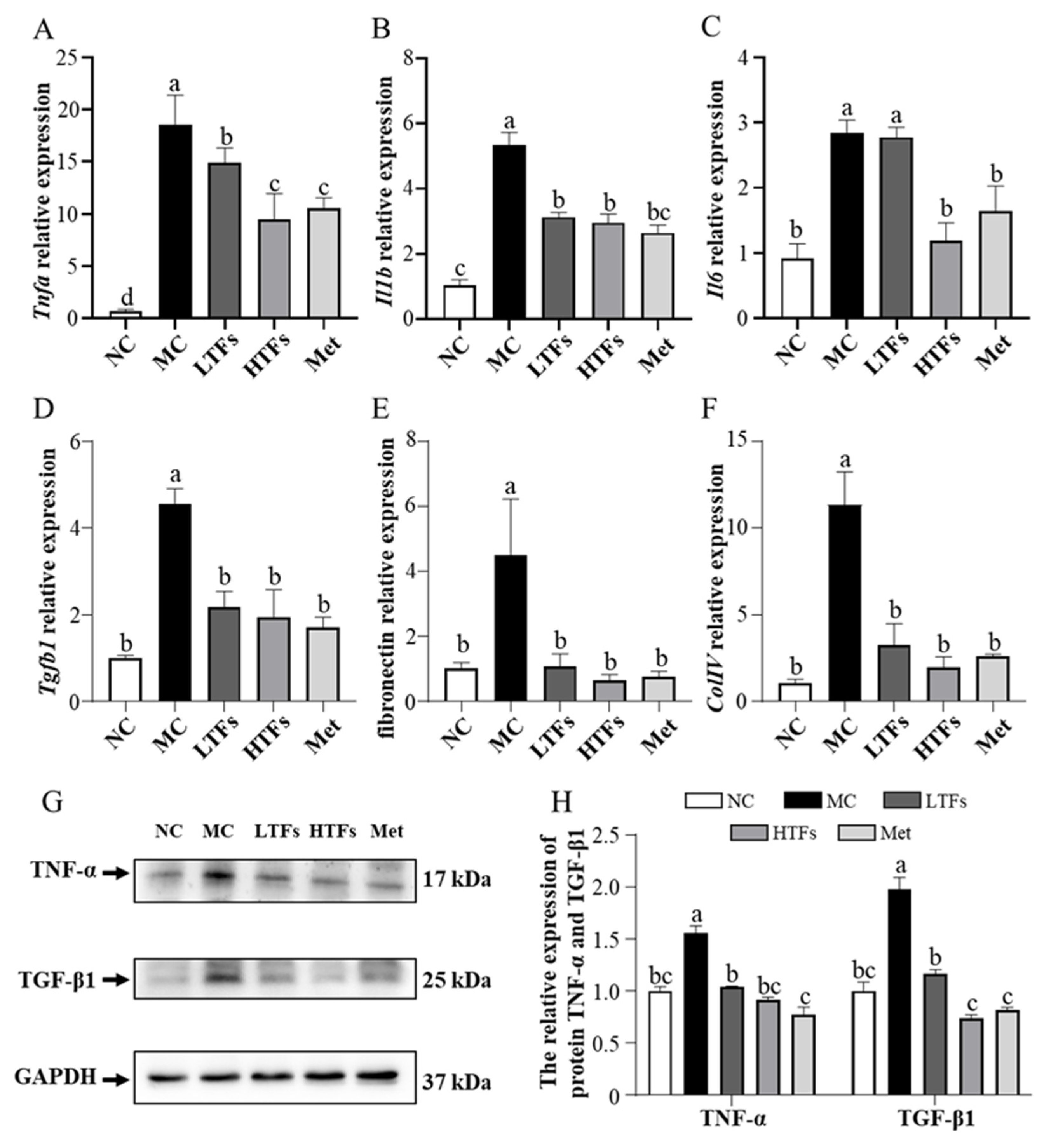
3.5. TFs Improved Inflammation and Fibrosis in the Kidney of T2DM Mice
3.6. TFs May Improve Diabetic Nephropathy through the MAPK/NF-κB Signaling Pathway
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hameed, I.; Masoodi, S.R.; Mir, S.A.; Nabi, M.; Ghazanfar, K.; Ganai, B.A. Type 2 diabetes mellitus: From a metabolic disorder to an inflammatory condition. World J. Diabetes 2015, 6, 598–612. [Google Scholar] [CrossRef]
- Singh, V.P.; Bali, A.; Singh, N.; Jaggi, A.S. Advanced glycation end products and diabetic complications. Korean J. Physiol. Pharm. 2014, 11, 1–14. [Google Scholar] [CrossRef]
- Sanajou, D.; Ghorbani Haghjo, A.; Argani, H.; Aslani, S. AGE-RAGE axis blockade in diabetic nephropathy: Current status and future directions. Eur. J. Pharmacol. 2018, 833, 158–164. [Google Scholar] [CrossRef]
- Fineberg, D.; Jandeleit-Dahm, K.A.; Cooper, M.E. Diabetic nephropathy: Diagnosis and treatment. Nat. Rev. Endocrinol. 2013, 9, 713–723. [Google Scholar] [CrossRef]
- Muskiet, M.H.; Smits, M.M.; Morsink, L.M.; Diamant, M. The gut-renal axis: Do incretin-based agents confer renoprotection in diabetes? Nat. Rev. Nephrol. 2014, 10, 88–103. [Google Scholar] [CrossRef]
- Shan, Z.; Nisar, M.F.; Li, M.; Zhang, C.; Wan, C.C. Theaflavin Chemistry and Its Health Benefits. Oxidative Med. Cell. Longev. 2021, 2021, 6256618. [Google Scholar] [CrossRef]
- Rohini, S.; Vishnupriya, P.; Rakshagan, V.; Jain, A.R. Protective effects of theaflavin. Drug Invent. Today 2018, 10, 2097–2101. [Google Scholar]
- Li, Z.; Zhu, J.; Wan, Z.; Li, G.; Chen, L.; Guo, Y. Theaflavin ameliorates renal ischemia/reperfusion injury by activating the Nrf2 signalling pathway in vivo and in vitro. Biomed. Pharmacother. Biomed. Pharmacother. 2021, 134, 111097. [Google Scholar] [CrossRef]
- Imran, A.; Arshad, M.U.; Arshad, M.S.; Imran, M.; Saeed, F.; Sohaib, M. Lipid peroxidation diminishing perspective of isolated theaflavins and thearubigins from black tea in arginine induced renal malfunctional rats. Lipids Health Dis. 2018, 17, 157. [Google Scholar] [CrossRef]
- Takemoto, M.; Takemoto, H. Synthesis of Theaflavins and Their Functions. Molecules 2018, 23, 918. [Google Scholar] [CrossRef]
- Sarkar, D.; Bose, S.K.; Chakraborty, T.; Roy, S. Theaflavin Enriched Black Tea Extract Alleviates Diabetic Nephropathy by Suppressing Hyperglycaemia-Mediated Oxidative Stress and Inflammation in Streptozotocin-Induced Rats. Nat. Prod. J. 2020, 11, 10. [Google Scholar] [CrossRef]
- Sergi, D.; Boulestin, H.; Campbell, F.M.; Williams, L.M. The Role of Dietary Advanced Glycation End Products in Metabolic Dysfunction. Mol. Nutr. Food Res. 2021, 65, e1900934. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Zhang, J.; Ho, C.-T.; Li, S. Management of Maillard reaction-derived reactive carbonyl species and advanced glycation end products by tea and tea polyphenols. Food Sci. Hum. Wellness 2022, 11, 557–567. [Google Scholar] [CrossRef]
- Lin, J.A.; Wu, C.H.; Yen, G.C. Perspective of Advanced Glycation End Products on Human Health. J. Agric. Food Chem. 2018, 66, 2065–2070. [Google Scholar] [CrossRef]
- Bettiga, A.; Fiorio, F.; Di Marco, F.; Trevisani, F.; Romani, A.; Porrini, E.; Salonia, A.; Montorsi, F.; Vago, R. The Modern Western Diet Rich in Advanced Glycation End-Products (AGEs): An Overview of Its Impact on Obesity and Early Progression of Renal Pathology. Nutrients 2019, 11, 1748. [Google Scholar] [CrossRef] [PubMed]
- Nedić, O.; Rattan, S.I.; Grune, T.; Trougakos, I.P. Molecular effects of advanced glycation end products on cell signalling pathways, ageing and pathophysiology. Free. Radic. Res. 2013, 47 (Suppl. S1), 28–38. [Google Scholar] [CrossRef]
- Wautier, M.P.; Chappey, O.; Corda, S.; Stern, D.M.; Schmidt, A.M.; Wautier, J.L. Activation of NADPH oxidase by AGE links oxidant stress to altered gene expression via RAGE. American journal of physiology. Endocrinol. Metab. 2001, 280, E685–E694. [Google Scholar]
- Zhu, D.; Wang, L.; Zhou, Q.; Yan, S.; Li, Z.; Sheng, J.; Zhang, W. (+)-Catechin ameliorates diabetic nephropathy by trapping methylglyoxal in type 2 diabetic mice. Mol. Nutr. Food Res. 2014, 58, 2249–2260. [Google Scholar] [CrossRef]
- Zhang, S.; Zhao, Y.; Ohland, C.; Jobin, C.; Sang, S. Microbiota facilitates the formation of the aminated metabolite of green tea polyphenol (-)-epigallocatechin-3-gallate which trap deleterious reactive endogenous metabolites. Free. Radic. Biol. Med. 2019, 131, 332–344. [Google Scholar] [CrossRef]
- Sampath, C.; Rashid, M.R.; Sang, S.; Ahmedna, M. Green tea epigallocatechin 3-gallate alleviates hyperglycemia and reduces advanced glycation end products via nrf2 pathway in mice with high fat diet-induced obesity. Biomed. Pharmacother. Biomed. Pharmacother. 2017, 87, 73–81. [Google Scholar] [CrossRef]
- Lo, C.Y.; Li, S.; Tan, D.; Pan, M.H.; Sang, S.; Ho, C.T. Trapping reactions of reactive carbonyl species with tea polyphenols in simulated physiological conditions. Mol. Nutr. Food Res. 2006, 50, 1118–1128. [Google Scholar] [CrossRef]
- Tan, D.; Wang, Y.; Lo, C.Y.; Ho, C.T. Methylglyoxal: Its presence and potential scavengers. Asia Pac. J. Clin. Nutr. 2008, 17 (Suppl. S1), 261–264. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.L.; Tsai, S.H.; Lin-Shiau, S.Y.; Ho, C.T.; Lin, J.K. Theaflavin-3,3′-digallate from black tea blocks the nitric oxide synthase by down-regulating the activation of NF-kappaB in macrophages. Eur. J. Pharmacol. 1999, 367, 379–388. [Google Scholar] [CrossRef]
- Guanglei, F.; Hua, W.; Youli, C.; Hui, Z.; Wenjun, F. Theaflavin alleviates inflammatory response and brain injury induced by cerebral hemorrhage via inhibiting the nuclear transcription factor kappa β-related pathway in rats. Drug Des. Dev. Ther. 2018, 12, 1609–1619. [Google Scholar]
- Gosslau, A.; En Jao, D.L.; Huang, M.T.; Ho, C.T.; Evans, D.; Rawson, N.E.; Chen, K.Y. Effects of the black tea polyphenol theaflavin-2 on apoptotic and inflammatory pathways in vitro and in vivo. Mol. Nutr. Food Res. 2011, 55, 198–208. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Li, J. Theaflavin-3, 3′-Digallate Attenuates Rheumatoid Inflammation in Mice Through the Nuclear Factor-κB and MAPK Pathways. Arch. Immunol. Ther. Exp. 2019, 67, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Jin, F.; Wang, Y.; Li, F.; Wang, L.; Wang, Q.; Ren, Z.; Wang, Y. In vitro and in vivo anti-inflammatory effects of theaflavin-3,3′-digallate on lipopolysaccharide-induced inflammation. Eur. J. Pharmacol. 2017, 794, 52–60. [Google Scholar] [CrossRef]
- Winzell, M.S.; Ahrén, B. The high-fat diet-fed mouse: A model for studying mechanisms and treatment of impaired glucose tolerance and type 2 diabetes. Diabetes 2004, 53 (Suppl. S3), S215–S219. [Google Scholar] [CrossRef]
- Deng, X.; Sun, L.; Lai, X.; Xiang, L.; Li, Q.; Zhang, W.; Zhang, L.; Sun, S. Tea Polypeptide Ameliorates Diabetic Nephropathy through RAGE and NF-κB Signaling Pathway in Type 2 Diabetes Mice. J. Agric. Food Chem. 2018, 66, 11957–11967. [Google Scholar] [CrossRef]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef]
- Mohan, T.; Velusamy, P.; Chakrapani, L.N.; Srinivasan, A.K.; Singh, A.; Johnson, T.; Periandavan, K. Impact of EGCG Supplementation on the Progression of Diabetic Nephropathy in Rats: An Insight into Fibrosis and Apoptosis. J. Agric. Food Chem. 2017, 65, 8028–8036. [Google Scholar] [CrossRef] [PubMed]
- Bansal, S.; Chawla, D.; Siddarth, M.; Banerjee, B.D.; Madhu, S.V.; Tripathi, A.K. A study on serum advanced glycation end products and its association with oxidative stress and paraoxonase activity in type 2 diabetic patients with vascular complications. Clin. Biochem. 2013, 46, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Greifenhagen, U.; Nguyen, V.D.; Moschner, J.; Giannis, A.; Frolov, A.; Hoffmann, R. Sensitive and site-specific identification of carboxymethylated and carboxyethylated peptides in tryptic digests of proteins and human plasma. J. Proteome Res. 2015, 14, 768–777. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Rai, V.; Singer, D.; Chabierski, S.; Xie, J.; Reverdatto, S.; Burz, D.S.; Schmidt, A.M.; Hoffmann, R.; Shekhtman, A. Advanced glycation end product recognition by the receptor for AGEs. Structure 2011, 19, 722–732. [Google Scholar] [CrossRef] [PubMed]
- O’Grady, K.L.; Khosla, S.; Farr, J.N.; Bondar, O.P.; Atkinson, E.J.; Achenbach, S.J.; Eckhardt, B.A.; Thicke, B.S.; Tweed, A.J.; Volkman, T.L.; et al. Development and Application of Mass Spectroscopy Assays for Nε-(1-Carboxymethyl)-L-Lysine and Pentosidine in Renal Failure and Diabetes. J. Appl. Lab. Med. 2020, 5, 558–568. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Zheng, Y.; Wang, Y.; Xiong, W.; Lin, L. Advanced glycation end products interfere with gastric smooth muscle contractile marker expression via the AGE/RAGE/NF-κB pathway. Exp. Mol. Pathol. 2017, 102, 7–14. [Google Scholar] [CrossRef]
- Ahmed, L.A.; Shiha, N.A.; Attia, A.S. Escitalopram Ameliorates Cardiomyopathy in Type 2 Diabetic Rats via Modulation of Receptor for Advanced Glycation End Products and Its Downstream Signaling Cascades. Front. Pharmacol. 2020, 11, 579206. [Google Scholar] [CrossRef]
- Gothandam, K.; Ganesan, V.S.; Ayyasamy, T.; Ramalingam, S. Antioxidant potential of theaflavin ameliorates the activities of key enzymes of glucose metabolism in high fat diet and streptozotocin—Induced diabetic rats. Redox Rep. Commun. Free. Radic. Res. 2019, 24, 41–50. [Google Scholar] [CrossRef]
- Kobayashi, M.; Ichitani, M.; Suzuki, Y.; Unno, T.; Sugawara, T.; Yamahira, T.; Kato, M.; Takihara, T.; Sagesaka, Y.; Kakuda, T.; et al. Black-tea polyphenols suppress postprandial hypertriacylglycerolemia by suppressing lymphatic transport of dietary fat in rats. J. Agric. Food Chem. 2009, 57, 7131–7136. [Google Scholar] [CrossRef]
- Qu, Z.; Liu, A.; Liu, C.; Tang, Q.; Zhan, L.; Xiao, W.; Huang, J.; Liu, Z.; Zhang, S. Theaflavin Promotes Mitochondrial Abundance and Glucose Absorption in Myotubes by Activating the CaMKK2-AMPK Signal Axis via Calcium-Ion Influx. J. Agric. Food Chem. 2021, 69, 8144–8159. [Google Scholar] [CrossRef]
- Miyata, Y.; Tamaru, S.; Tanaka, T.; Tamaya, K.; Matsui, T.; Nagata, Y.; Tanaka, K. Theflavins and theasinensin A derived from fermented tea have antihyperglycemic and hypotriacylglycerolemic effects in KK-A(y) mice and Sprague-Dawley rats. J. Agric. Food Chem. 2013, 61, 9366–9372. [Google Scholar] [CrossRef] [PubMed]
- Imran, A.; Butt, M.S.; Arshad, M.S.; Arshad, M.U.; Saeed, F.; Sohaib, M.; Munir, R. Exploring the potential of black tea based flavonoids against hyperlipidemia related disorders. Lipids Health Dis. 2018, 17, 57. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.J.; Wang, L.; Li, Z.; Zhu, D.N.; Guo, S.C.; Xin, W.F.; Yang, Y.F.; Cong, X.; Ma, T.; Shen, P.P.; et al. Inhibition of advanced glycation end product formation by Pu-erh tea ameliorates progression of experimental diabetic nephropathy. J. Agric. Food Chem. 2012, 60, 4102–4110. [Google Scholar] [CrossRef] [PubMed]
- Brosius, F.C., 3rd; Alpers, C.E.; Bottinger, E.P.; Breyer, M.D.; Coffman, T.M.; Gurley, S.B.; Harris, R.C.; Kakoki, M.; Kretzler, M.; Leiter, E.H.; et al. Mouse models of diabetic nephropathy. J. Am. Soc. Nephrol. JASN 2009, 20, 2503–2512. [Google Scholar] [CrossRef] [PubMed]
- Dawnay, A. Renal clearance of glycation adducts: Anti-glycation defence in uraemia and dialysis. Biochem. Soc. Trans. 2003, 31, 1386–1389. [Google Scholar] [CrossRef] [PubMed]
- Hartog, J.W.; Smit, A.J.; van Son, W.J.; Navis, G.; Gans, R.O.; Wolffenbuttel, B.H.; de Jong, P.E. Advanced glycation end products in kidney transplant patients: A putative role in the development of chronic renal transplant dysfunction. Am. J. Kidney Dis. Off. J. Natl. Kidney Found. 2004, 43, 966–975. [Google Scholar] [CrossRef]
- Genuth, S.; Sun, W.; Cleary, P.; Sell, D.R.; Dahms, W.; Malone, J.; Sivitz, W.; Monnier, V.M. Glycation and carboxymethyllysine levels in skin collagen predict the risk of future 10-year progression of diabetic retinopathy and nephropathy in the diabetes control and complications trial and epidemiology of diabetes interventions and complications participants with type 1 diabetes. Diabetes 2005, 54, 3103–3111. [Google Scholar]
- Rasheed, Z.; Akhtar, N.; Anbazhagan, A.N.; Ramamurthy, S.; Voss, F.R.; Haqqi, T.M. 210 green tea polyphenol epigallocatechin-3-gallate (EGCG) inhibits advanced glycation end products-induced expression of tumor necrosis factor-α and matrix metalloproteinase-13 in human osteoarthritis chondrocytes. Osteoarthr. Cartil. 2009, 17, S119–S120. [Google Scholar] [CrossRef]
- Zeng, L.; Lin, L.; Xiao, W.; Li, Y. L-theanine protects rat kidney from D-galactose-induced injury via inhibition of the AGEs/RAGE signaling pathway. Eur. J. Pharmacol. 2022, 927, 175072. [Google Scholar] [CrossRef]
- Kyriakis, J.M.; Avruch, J. Mammalian MAPK signal transduction pathways activated by stress and inflammation: A 10-year update. Physiol. Rev. 2012, 92, 689–737. [Google Scholar] [CrossRef] [PubMed]
- Cuadrado, A.; Nebreda, A.R. Mechanisms and functions of p38 MAPK signalling. Biochem. J. 2010, 429, 403–417. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | NC | MC | LTFs | HTFs | Met |
|---|---|---|---|---|---|
| Insulin (mU/L) | 20.58 ± 1.95 c | 49.68 ± 2.56 a | 39.82 ± 3.01 b | 25.87 ± 3.90 c | 21.43 ± 4.08 c |
| HOMA-IR | 6.03 ± 0.72 c | 59.64 ± 10.85 a | 32.16 ± 8.62 b | 21.36 ± 7.02 bc | 12.03 ± 2.07 c |
| HOMA-IS | 0.17 ± 0.022 a | 0.017 ± 0.0036 d | 0.033 ± 0.0082 cd | 0.051 ± 0.016 c | 0.085 ± 0.015 b |
| Parameter | NC | MC | LTFs | HTFs | Met |
|---|---|---|---|---|---|
| Plasma | |||||
| Total AGEs (AU) | 56.50 ± 1.67 b | 80.03 ± 8.56 a | 60.26 ± 1.45 b | 58.07 ± 1.29 b | 59.11 ± 1.19 b |
| CML (ng/mL) | 1252.20 ± 64.86 b | 1524.56 ± 80.30 a | 1098.20 ± 53.16 bc | 990.18 ± 46.48 c | 684.49 ± 36.21 d |
| CEL (ng/mL) | 14.00 ± 0.48 b | 17.19 ± 0.62 a | 15.08 ± 0.50 b | 11.17 ± 0.34 c | 8.12 ± 0.30 d |
| MGO (ng/mL) | 19.09 ± 0.29 b | 21.93 ± 0.43 a | 11.29 ± 0.49 d | 14.49 ± 0.39 c | 13.69 ± 0.83 c |
| MDA (nmol/mL) | 2.53 ± 0.057 d | 7.21 ± 0.11 a | 5.51 ± 0.25 b | 4.47 ± 0.20 c | 4.84 ± 0.24 bc |
| Kidney | |||||
| Total AGEs (AU/mgprot) | 0.17 ± 0.027 c | 0.56 ± 0.028 a | 0.49 ± 0.031 ab | 0.41 ± 0.042 b | 0.43 ± 0.019 ab |
| CML (ng/mgprot) | 389.41 ± 33.56 a | 460.32 ± 30.35 a | 217.28 ± 3.07 b | 359.05 ± 22.84 a | 359.60 ± 10.53 a |
| CEL (ng/mgprot) | 3.87 ± 0.22 bc | 5.42 ± 0.47 a | 5.08 ± 0.31 ab | 2.88 ± 0.21 c | 3.79 ± 0.20 c |
| MGO (ng/mgprot) | 4.46 ± 0.40 b | 7.44 ± 0.41 a | 6.98 ± 0.48 a | 4.83 ± 0.24 b | 5.03 ± 0.16 ab |
| MDA (nmol/mgprot) | 0.72 ± 0.061 c | 2.20 ± 0.010 a | 1.47 ± 0.054 b | 1.35 ± 0.015 b | 1.22 ± 0.12 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Jiang, J.; Zhao, C.; Shan, H.; Shao, Z.; Wang, C.; Guan, J.; Xie, Z.; Li, S. The Protective Effect of Theaflavins on the Kidney of Mice with Type II Diabetes Mellitus. Nutrients 2023, 15, 201. https://doi.org/10.3390/nu15010201
Wang J, Jiang J, Zhao C, Shan H, Shao Z, Wang C, Guan J, Xie Z, Li S. The Protective Effect of Theaflavins on the Kidney of Mice with Type II Diabetes Mellitus. Nutrients. 2023; 15(1):201. https://doi.org/10.3390/nu15010201
Chicago/Turabian StyleWang, Jun, Jingjing Jiang, Changyu Zhao, Hongyan Shan, Ziheng Shao, Chun Wang, Jiayun Guan, Zhongwen Xie, and Songnan Li. 2023. "The Protective Effect of Theaflavins on the Kidney of Mice with Type II Diabetes Mellitus" Nutrients 15, no. 1: 201. https://doi.org/10.3390/nu15010201
APA StyleWang, J., Jiang, J., Zhao, C., Shan, H., Shao, Z., Wang, C., Guan, J., Xie, Z., & Li, S. (2023). The Protective Effect of Theaflavins on the Kidney of Mice with Type II Diabetes Mellitus. Nutrients, 15(1), 201. https://doi.org/10.3390/nu15010201