
Role of HO-1 against Saturated Fatty Acid-Induced Oxidative Stress in Hepatocytes

 , ,
, ,
Abstract
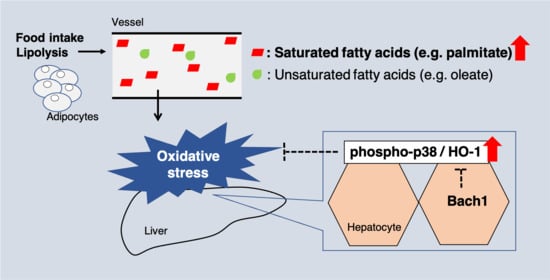
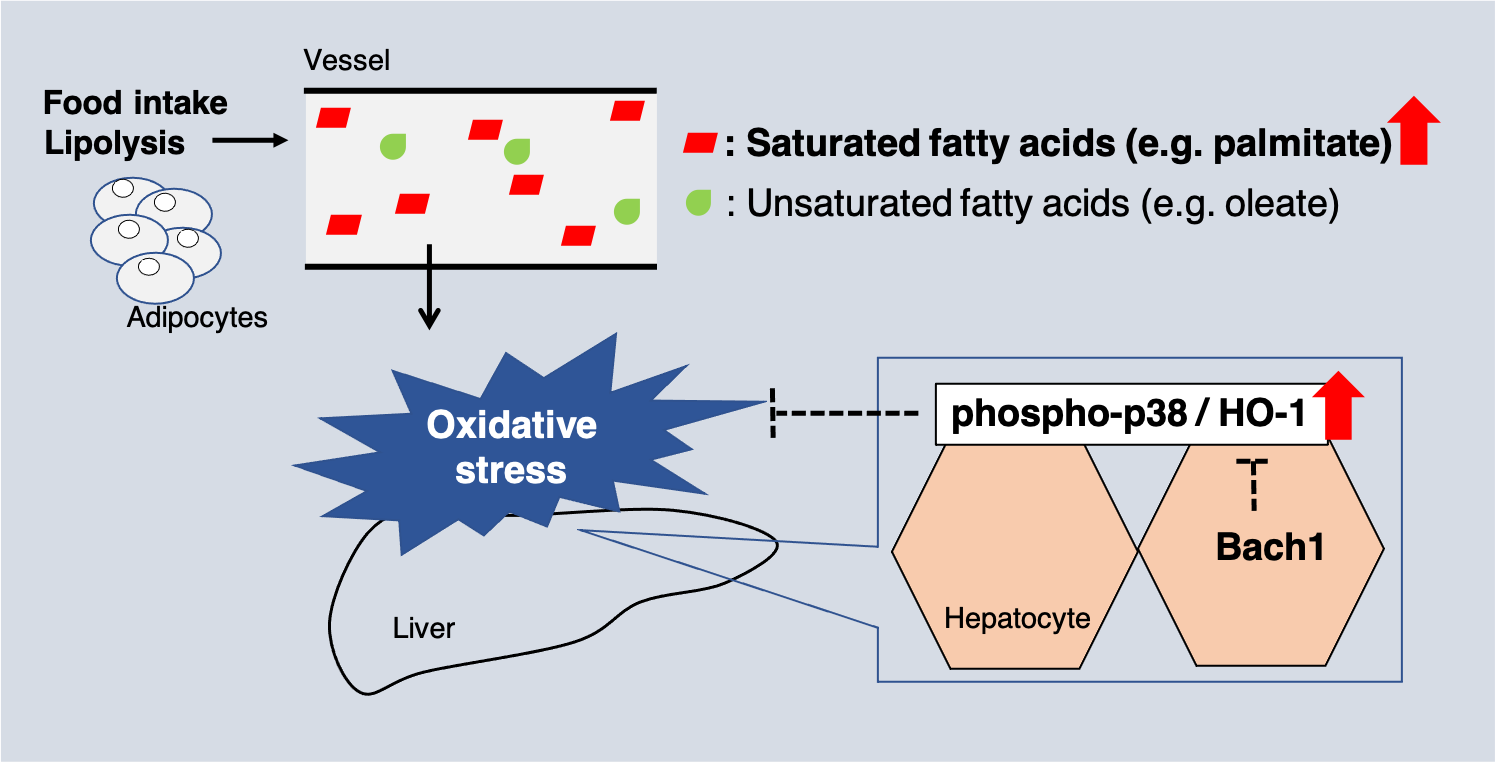
1. Introduction
2. Materials and Methods
2.1. Palmitate-Injected Mice Model
2.2. High-Fat-Diet-Induced Fatty Liver Mice Model
2.3. Cell Cultures
2.4. Preparation of Palmitate/Bovine Serum Albumin Complex Solution for Cell Culture
2.5. Antibodies, Reagents, and siRNAs
2.6. Western Blotting
2.7. Detection of Reactive Oxygen Species (ROS) Production
2.8. Lactate Dehydrogenase (LDH) Assay Using the Cell Culture Supernatant
2.9. Measurement of Malondialdehyde (MDA) in the Liver and Culture Cell Homogenate
2.10. Analysis of Gene Expression by Quantitative Reverse-Transcription Polymerase Chain Reaction (qRT-PCR)
2.11. Statistical Analysis
3. Results
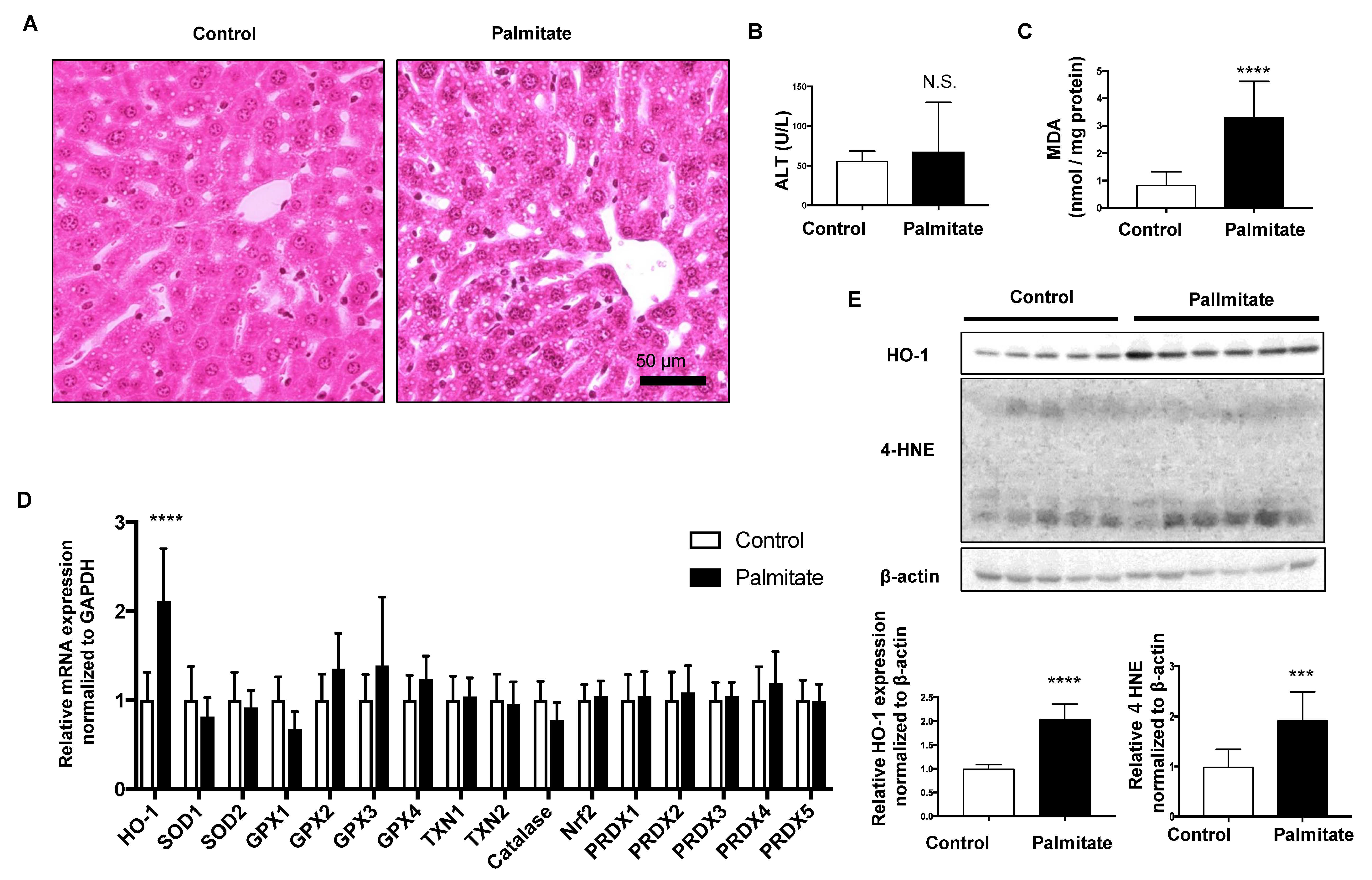
3.1. Effect of Palmitate Injection on Balb-c Mice Liver
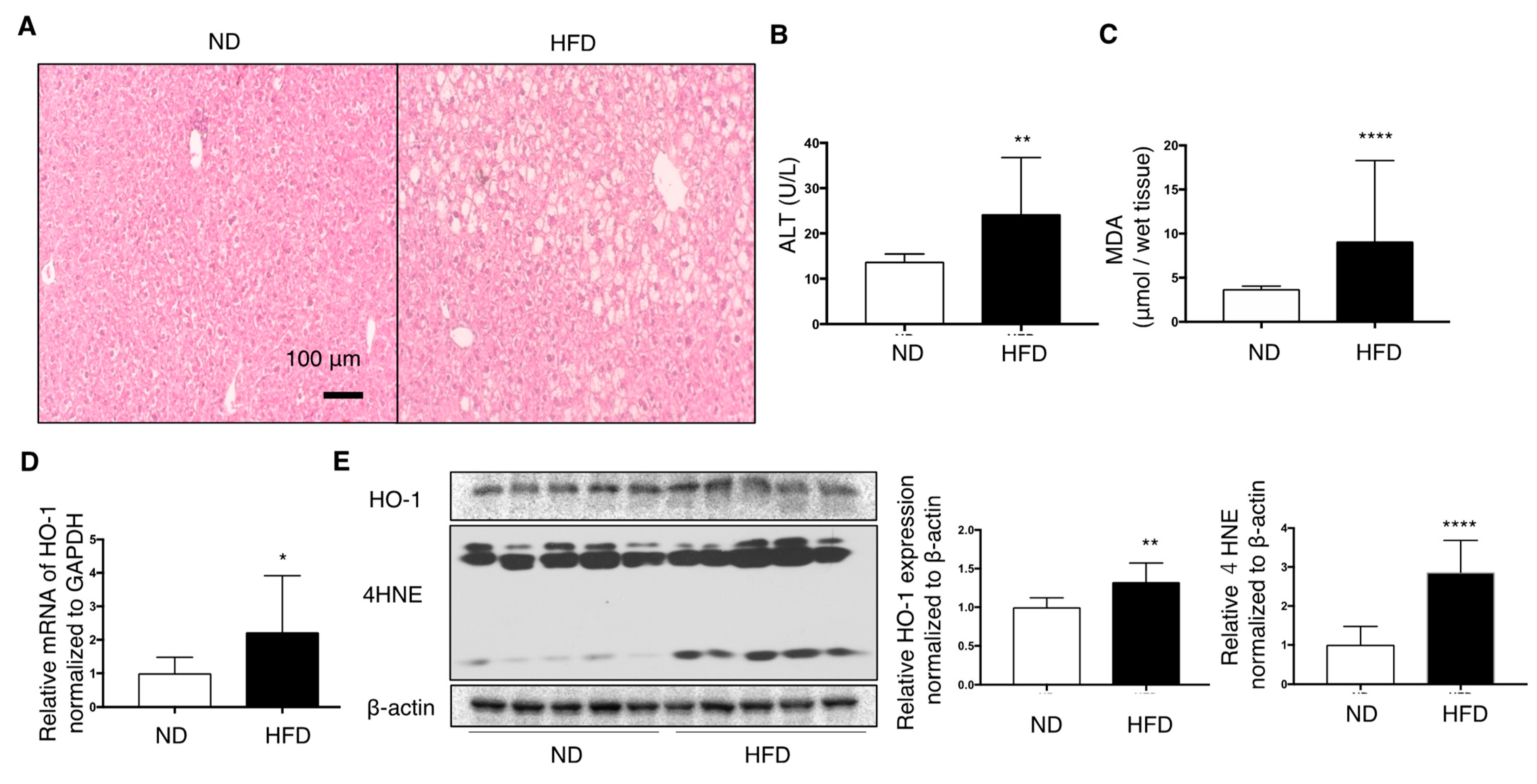
3.2. Effect of HFD on C57/BL6J Mouse Livers
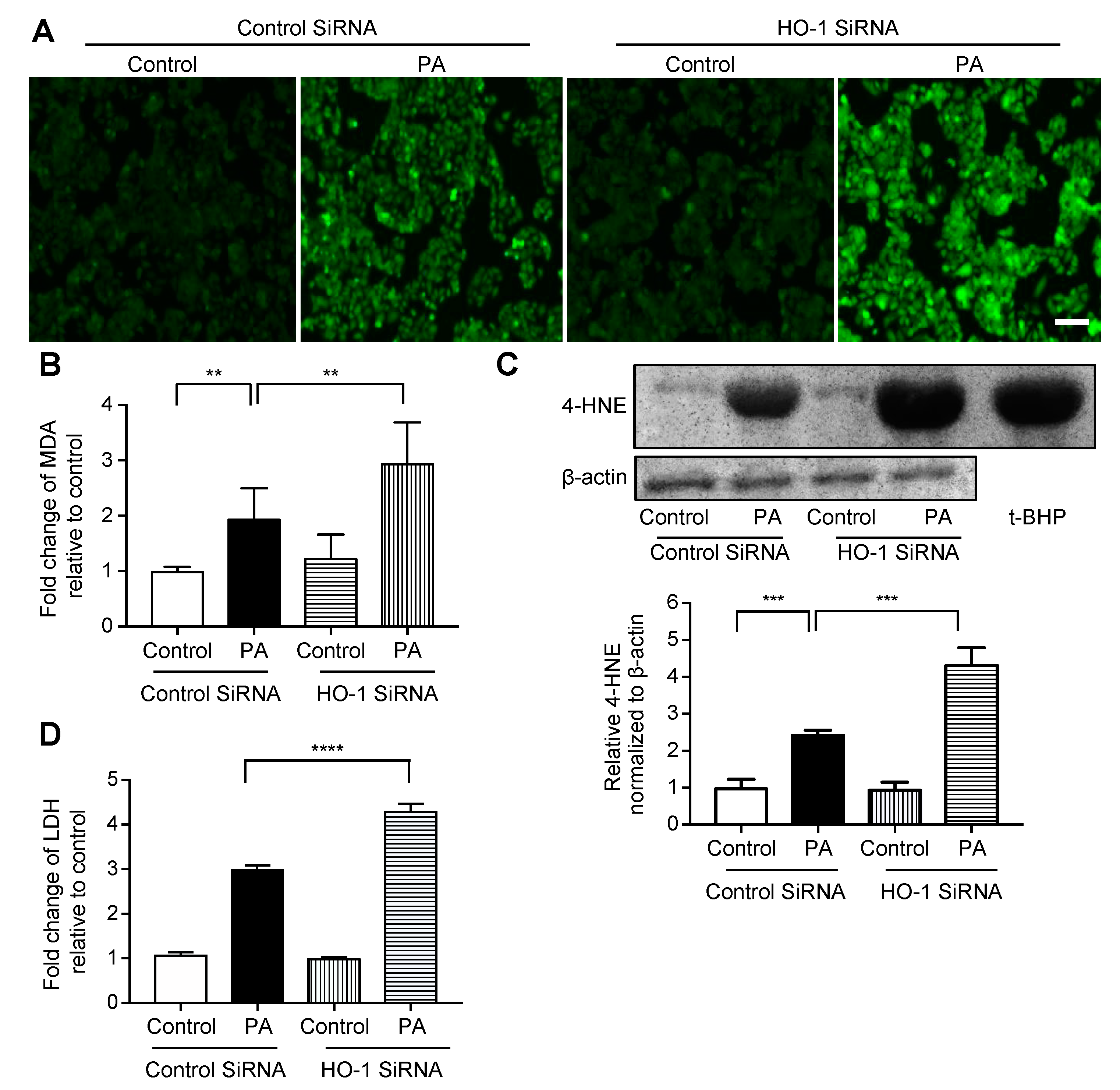
3.3. Effect of HO-1 Knockdown Against Palmitate-Induced Oxidative Stress in HepG2 Cells
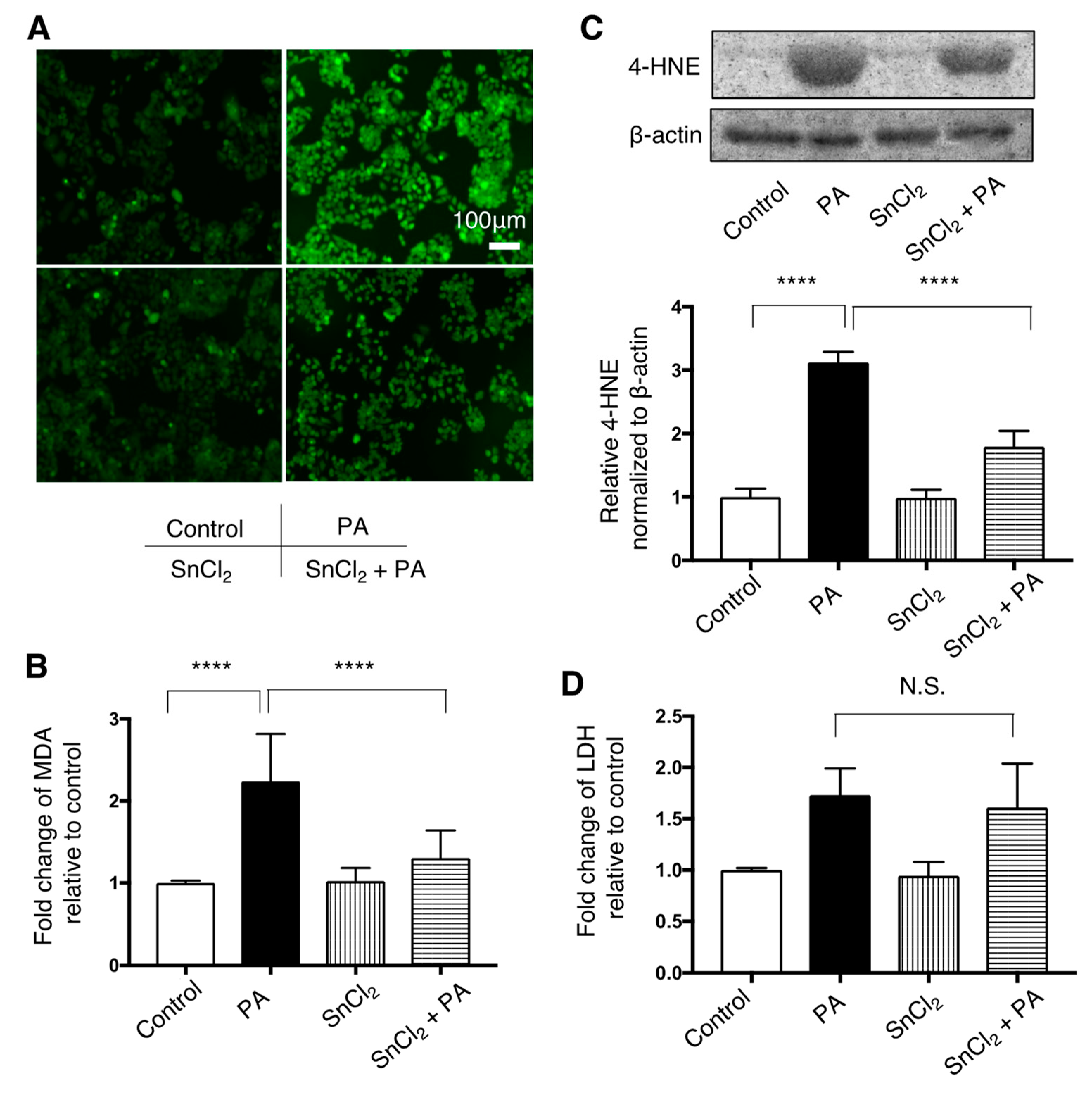
3.4. Effect of SnCl2 on Oxidative Stress in HepG2 Cells Treated with Palmitate
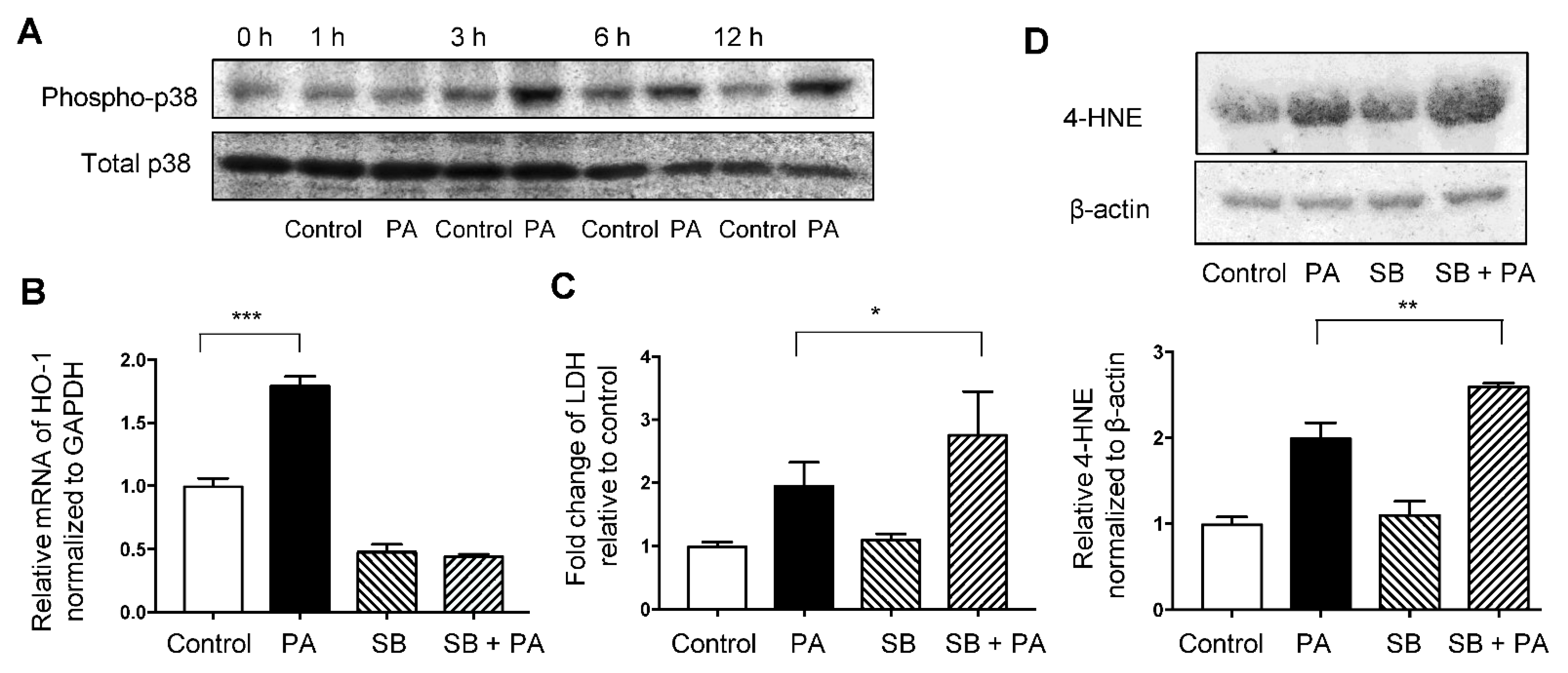
3.5. Involvement of p38/HO-1 Pathway in Palmitate-Induced Oxidative Stress in HepG2 Cells
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Younossi, Z.M.; Koenig, A.B.; Abdelatif, D.; Fazel, Y.; Henry, L.; Wymer, M. Global epidemiology of nonalcoholic fatty liver disease—Meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology 2016, 64, 73–84. [Google Scholar] [CrossRef]
- Buzzetti, E.; Pinzani, M.; Tsochatzis, E.A. The multiple-hit pathogenesis of non-alcoholic fatty liver disease (NAFLD). Metabolism 2016, 65, 1038–1048. [Google Scholar] [CrossRef]
- Postic, C.; Girard, J. Contribution of de novo fatty acid synthesis to hepatic steatosis and insulin resistance: Lessons from genetically engineered mice. J. Clin. Investig. 2008, 118, 829–838. [Google Scholar] [CrossRef]
- MacHado, M.V.; Diehl, A.M. Pathogenesis of nonalcoholic steatohepatitis. Gastroenterology 2016, 150, 1769–1777. [Google Scholar] [CrossRef]
- Trauner, M.; Arrese, M.; Wagner, M. Fatty liver and lipotoxicity. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2010, 1801, 299–310. [Google Scholar] [CrossRef]
- Marra, F.; Svegliati-Baroni, G. Lipotoxicity and the gut-liver axis in NASH pathogenesis. J. Hepatol. 2018, 68, 280–295. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Lu, Z.; Ru, J.H.; Lopes-Virella, M.F.; Lyons, T.J.; Huang, Y. Saturated fatty acid combined with lipopolysaccharide stimulates a strong inflammatory response in hepatocytes in vivo and in vitro. Am. J. Physiol. Endocrinol. Metab. 2018, 315, E745–E757. [Google Scholar] [CrossRef] [PubMed]
- Townsend, L.K.; Medak, K.D.; Peppler, W.T.; Meers, G.M.; Rector, R.S.; LeBlanc, P.J.; Wright, D.C. High-saturated-fat diet-induced obesity causes hepatic interleukin-6 resistance via endoplasmic reticulum stress. J. Lipid Res. 2019, 60, 1236–1249. [Google Scholar] [CrossRef] [PubMed]
- Ogino, N.; Miyagawa, K.; Kusanaga, M.; Hayashi, T.; Minami, S.; Oe, S.; Honma, Y.; Harada, M. Involvement of sarco/endoplasmic reticulum calcium ATPase-mediated calcium flux in the protective effect of oleic acid against lipotoxicity in hepatocytes. Exp. Cell Res. 2019, 385, 111651. [Google Scholar] [CrossRef] [PubMed]
- Miyagawa, K.; Oe, S.; Honma, Y.; Izumi, H.; Baba, R.; Harada, M. Lipid-induced endoplasmic reticulum stress impairs selective autophagy at the step of autophagosome-lysosome fusion in hepatocytes. Am. J. Pathol. 2016, 186, 1861–1873. [Google Scholar] [CrossRef] [PubMed]
- Musso, G.; Cassader, M.; Paschetta, E.; Gambino, R. Bioactive lipid species and metabolic pathways in progression and resolution of nonalcoholic steatohepatitis. Gastroenterology 2018, 155, 282–302.e8. [Google Scholar] [CrossRef] [PubMed]
- Silva Figueiredo, P.; Carla Inada, A.; Marcelino, G.; Maiara Lopes Cardozo, C.; de Cássia Freitas, K.; De Cássia Avellaneda Guimarães, R.; Pereira de Castro, A.; Aragão do Nascimento, V.; Aiko Hiane, P. Fatty Acids Consumption: The Role Metabolic Aspects Involved in Obesity and Its Associated Disorders. Nutrients 2017, 9, 1158. [Google Scholar] [CrossRef] [PubMed]
- Jensen, M.D. Role of body fat distribution and the metabolic complications of obesity. J. Clin. Endocrinol. Metab. 2008, 93, S57–S63. [Google Scholar] [CrossRef] [PubMed]
- Leamy, A.K.; Egnatchik, R.A.; Young, J.D. Molecular mechanisms and the role of saturated fatty acids in the progression of non-alcoholic fatty liver disease. Prog. Lipid Res. 2013, 52, 165–174. [Google Scholar] [CrossRef]
- Pilar Valdecantos, M.; Prieto-Hontoria, P.L.; Pardo, V.; Módol, T.; Santamaría, B.; Weber, M.; Herrero, L.; Serra, D.; Muntané, J.; Cuadrado, A.; et al. Essential role of Nrf2 in the protective effect of lipoic acid against lipoapoptosis in hepatocytes. Free Radic. Biol. Med. 2015, 84, 263–278. [Google Scholar] [CrossRef]
- Xu, D.; Xu, M.; Jeong, S.; Qian, Y.; Wu, H.; Xia, Q.; Kong, X. The role of Nrf2 in liver disease: Novel molecular mechanisms and therapeutic approaches. Front. Pharmacol. 2019, 9, 1428. [Google Scholar] [CrossRef] [PubMed]
- Loboda, A.; Damulewicz, M.; Pyza, E.; Jozkowicz, A.; Dulak, J. Role of Nrf2/HO-1 system in development, oxidative stress response and diseases: An evolutionarily conserved mechanism. Cell Mol. Life Sci. 2016, 73, 3221–3247. [Google Scholar] [CrossRef]
- Abraham, N.G.; Kappas, A. Pharmacological and clinical aspects of heme oxygenase. Pharmacol. Rev. 2008, 60, 79–127. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Kim, D.H.; Tsenovoy, P.L.; Peterson, S.J.; Rezzani, R.; Rodella, L.F.; Aronow, W.S.; Ikehara, S.; Abraham, N.G. Treatment of obese diabetic mice with a heme oxygenase inducer reduces visceral and subcutaneous adiposity, increases adiponectin levels, and improves insulin sensitivity and glucose tolerance. Diabetes 2008, 57, 1526–1535. [Google Scholar] [CrossRef]
- Tu, T.H.; Joe, Y.; Choi, H.S.; Chung, H.T.; Yu, R. Induction of heme oxygenase-1 with hemin reduces obesity-induced adipose tissue inflammation via adipose macrophage phenotype switching. Mediat. Inflamm. 2014, 2014, 290708. [Google Scholar] [CrossRef]
- Eguchi, K.; Manabe, I.; Oishi-Tanaka, Y.; Ohsugi, M.; Kono, N.; Ogata, F.; Yagi, N.; Ohto, U.; Kimoto, M.; Miyake, K.; et al. Saturated fatty acid and TLR signaling link β cell dysfunction and islet inflammation. Cell Metab. 2012, 15, 518–533. [Google Scholar] [CrossRef] [PubMed]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Ogawa, Y.; Imajo, K.; Honda, Y.; Kessoku, T.; Tomeno, W.; Kato, S.; Fujita, K.; Yoneda, M.; Saito, S.; Saigusa, Y.; et al. Palmitate-induced lipotoxicity is crucial for the pathogenesis of nonalcoholic fatty liver disease in cooperation with gut-derived endotoxin. Sci. Rep. 2018, 8, 11365. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Qian, Y.; Fang, Q.; Zhong, P.; Li, W.; Wang, L.; Fu, W.; Zhang, Y.; Xu, Z.; Li, X.; et al. Saturated palmitic acid induces myocardial inflammatory injuries through direct binding to TLR4 accessory protein MD2. Nat. Commun. 2017, 8, 13997. [Google Scholar] [CrossRef]
- Nan, Y.; Wang, R.; Zhao, S.; Han, F.; Wu, W.J.; Kong, L.; Fu, N.; Kong, L.; Yu, J. Heme oxygenase-1 prevents non-alcoholic steatohepatitis through suppressing hepatocyte apoptosis in mice. Lipids Health Dis. 2010, 9, 124. [Google Scholar] [CrossRef]
- Li, D.; Zhao, D.; Du, J.; Dong, S.; Aldhamin, Z.; Yuan, X.; Li, W.; Du, H.; Zhao, W.; Cui, L.; et al. Heme oxygenase-1 alleviated non-alcoholic fatty liver disease via suppressing ROS-dependent endoplasmic reticulum stress. Life Sci. 2020, 253, 117678. [Google Scholar] [CrossRef]
- Yang, J.; Kim, C.S.; Tu, T.H.; Kim, M.S.; Goto, T.; Kawada, T.; Choi, M.S.; Park, T.; Sung, M.K.; Yun, J.W.; et al. Quercetin protects obesity-induced hypothalamic inflammation by reducing microglia-mediated inflammatory responses via HO-1 induction. Nutrients 2017, 9, 650. [Google Scholar] [CrossRef]
- Kuo, N.C.; Huang, S.Y.; Yang, C.Y.; Shen, H.H.; Lee, Y.M. Involvement of HO-1 and Autophagy in the Protective Effect of Magnolol in Hepatic Steatosis-Induced NLRP3 Inflammasome Activation In Vivo and In Vitro. Antioxidants 2020, 9, 924. [Google Scholar] [CrossRef]
- Kwon, C.H.; Sun, J.L.; Kim, M.J.; Abd El-Aty, A.M.; Jeong, J.H.; Jung, T.W. Clinically confirmed DEL-1 as a myokine attenuates lipid-induced inflammation and insulin resistance in 3T3-L1 adipocytes via AMPK/HO-1- pathway. Adipocyte 2020, 9, 576–586. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, X.; Zhang, L.; Yan, T.; Wu, B.; Xu, F.; Jia, Y. Silychristin A activates Nrf2-HO-1/SOD2 pathway to reduce apoptosis and improve GLP-1 production through upregulation of estrogen receptor α in GLUTag cells. Eur. J. Pharmacol. 2020, 881, 173236. [Google Scholar] [CrossRef]
- Ma, W.F.; Duan, X.C.; Han, L.; Zhang, L.L.; Meng, X.M.; Li, Y.L.; Wang, M. Vanillic acid alleviates palmitic acid-induced oxidative stress in human umbilical vein endothelial cells via adenosine monophosphate-activated protein kinase signaling pathway. J. Food Biochem. 2019, 43, e12893. [Google Scholar] [CrossRef]
- Park, E.J.; Lee, A.Y.; Park, S.; Kim, J.H.; Cho, M.H. Multiple pathways are involved in palmitic acid-induced toxicity. Food Chem. Toxicol. 2014, 67, 26–34. [Google Scholar] [CrossRef]
- Kwak, H.J.; Yang, D.; Hwang, Y.; Jun, H.S.; Cheon, H.G. Baicalein protects rat insulinoma INS-1 cells from palmitate-induced lipotoxicity by inducing HO-1. PLoS ONE 2017, 12, e0176432. [Google Scholar] [CrossRef]
- Shi, Y.; Sun, Y.; Sun, X.; Zhao, H.; Yao, M.; Hou, L.; Jiang, L. Up-regulation of HO-1 by Nrf2 activation protects against palmitic acid-induced ROS increase in human neuroblastoma BE(2)-M17 cells. Nutr. Res. 2018, 52, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Brand, M.; Zenke, Y.; Tashiro, S.; Groudine, M.; Igarashi, K. Heme regulates the dynamic exchange of Bach1 and NF-E2-related factors in the Maf transcription factor network. Proc. Natl. Acad. Sci. USA 2004, 101, 1461–1466. [Google Scholar] [CrossRef] [PubMed]
- Ka, S.O.; Bang, I.H.; Bae, E.J.; Park, B.H. Hepatocyte-specific sirtuin 6 deletion predisposes to nonalcoholic steatohepatitis by up-regulation of Bach1, an Nrf2 repressor. FASEB J. 2017, 31, 3999–4010. [Google Scholar] [CrossRef] [PubMed]
- Stocker, R.; Yamamoto, Y.; McDonagh, A.F.; Glazer, A.N.; Ames, B.N. Bilirubin is an antioxidant of possible physiological importance. Science 1987, 235, 1043–1046. [Google Scholar] [CrossRef] [PubMed]
- Clark, J.E.; Foresti, R.; Green, C.J.; Motterlini, R. Dynamics of haem oxygenase-1 expression and bilirubin production in cellular protection against oxidative stress. Biochem. J. 2000, 348, 615–619. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Gene | Forward (5′-3′) | Reverse (5′-3′) | Size (bp) | Accession No. |
|---|---|---|---|---|---|
| Homo sapiens | HOMX1 | GGCCAGCAACAAAGTGCAAG | TGGCATAAAGCCCTACAGCA | 146 | NM_002133.3 |
| GAPDH | AAGGTGAAGGTCGGAGTCAA | AATGAAGGGGTCATTGATGG | 108 | NM_002046 | |
| Mus musculus | Homx1 | CCTCACAGATGGCGTCACTT | GCTGATCTGGGGTTTCCCTC | 92 | NM_010442.2 |
| Sod1 | GAACCATCCACTTCGAGCA | TACTGATGGACGTGGAACCC | 103 | NM_011434 | |
| Sod2 | ACTGAAGTTCAATGGTGGGG | GCTTGATAGCCTCCAGCAAC | 107 | NM_013671.3 | |
| Gpx1 | GTTTCCCGTGCAATCAGTTC | CAATGTAAAATTGGGCTCGAA | 110 | NM_008160.6 | |
| Gpx2 | CTGCAATGTCGCTTTCCCAG | CCCCAGGTCGGACATACTTG | 121 | NM_30677.2 | |
| Gpx3 | CATCCTGCCTTCTGTCCCT | ATGGTACCACTCATACCGCC | 100 | NM_008161.4 | |
| Gpx4 | GCCCACCCACTGTGGAAATG | TGGGACCATAGCGCTTCACC | 130 | NM_008162.4 | |
| Prdx1 | GCCGCTCTGTGGATGAGATT | ATCACTGCCAGGTTTCCAGC | 98 | NM_011034.4 | |
| Prdx2 | CTCCTCGGTATCTCCGCCTA | TAGCACTTGCATGACGAGCA | 102 | NM_007452.2 | |
| Prdx3 | ATGACCTACCTGTGGGACGC | GGCTTGATGGTGTCACTGC | 124 | NM_011563.6 | |
| Prdx4 | TGCCACTTCTACGCTGGTG | CCCAATAAGGTGCTGGCTTG | 115 | NM_016764.5 | |
| Prdx5 | CAGTTCTGTGCTCCGTGCAT | GCATCTCCCACCTTGATCGG | 132 | NM_012021.3 | |
| Nrf2 | TAGATGACCATGAGTCGCTTGC | GCCAAACTTGCTCCATGTCC | 153 | NM_010902 | |
| Txn1 | AGTGGATGTGGATGACTGCC | CCTTGTTAGCACCGGAGAACT | 116 | NM_011660.3 | |
| Txn2 | TGAGACACCAGTTGTTGTGGA | TTGGCGACCATCTTCTCTAGC | 87 | NM_019913.5 | |
| Catalase | GATGAAGCAGTGGAAGGAGC | CCCGCGGTCATGATATTAAGT | 102 | NM_009804 | |
| Bach1 | GTCTCGGCTCCGGTCGAT | TGCTATGCACAGAGGACTCG | 124 | NM_007520.2 | |
| Gapdh | AGGTCGGTGTGAACGGATTTG | TGTAGACCATGTAGTTGAGGTCA | 123 | NM_008084 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ogino, N.; Miyagawa, K.; Nagaoka, K.; Matsuura-Harada, Y.; Ogino, S.; Kusanaga, M.; Oe, S.; Honma, Y.; Harada, M.; Eitoku, M.; et al. Role of HO-1 against Saturated Fatty Acid-Induced Oxidative Stress in Hepatocytes. Nutrients 2021, 13, 993. https://doi.org/10.3390/nu13030993
Ogino N, Miyagawa K, Nagaoka K, Matsuura-Harada Y, Ogino S, Kusanaga M, Oe S, Honma Y, Harada M, Eitoku M, et al. Role of HO-1 against Saturated Fatty Acid-Induced Oxidative Stress in Hepatocytes. Nutrients. 2021; 13(3):993. https://doi.org/10.3390/nu13030993
Chicago/Turabian StyleOgino, Noriyoshi, Koichiro Miyagawa, Kenjiro Nagaoka, Yuki Matsuura-Harada, Shihona Ogino, Masashi Kusanaga, Shinji Oe, Yuichi Honma, Masaru Harada, Masamitsu Eitoku, and et al. 2021. "Role of HO-1 against Saturated Fatty Acid-Induced Oxidative Stress in Hepatocytes" Nutrients 13, no. 3: 993. https://doi.org/10.3390/nu13030993
APA StyleOgino, N., Miyagawa, K., Nagaoka, K., Matsuura-Harada, Y., Ogino, S., Kusanaga, M., Oe, S., Honma, Y., Harada, M., Eitoku, M., Suganuma, N., & Ogino, K. (2021). Role of HO-1 against Saturated Fatty Acid-Induced Oxidative Stress in Hepatocytes. Nutrients, 13(3), 993. https://doi.org/10.3390/nu13030993