
Simultaneous Tests of Theaflavin-3,3′-digallate as an Anti-Diabetic Drug in Human Hepatoma G2 Cells and Zebrafish (Danio rerio)

 , ,
, ,  ,
,  and
and
Abstract
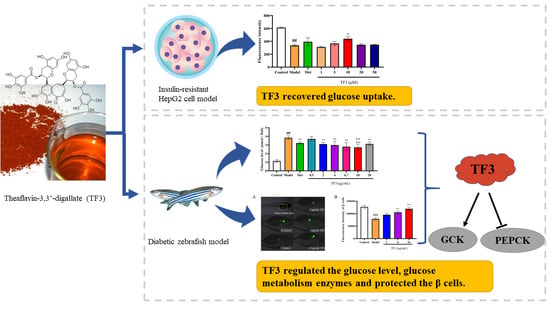
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture and Insulin Resistance Model Establishment
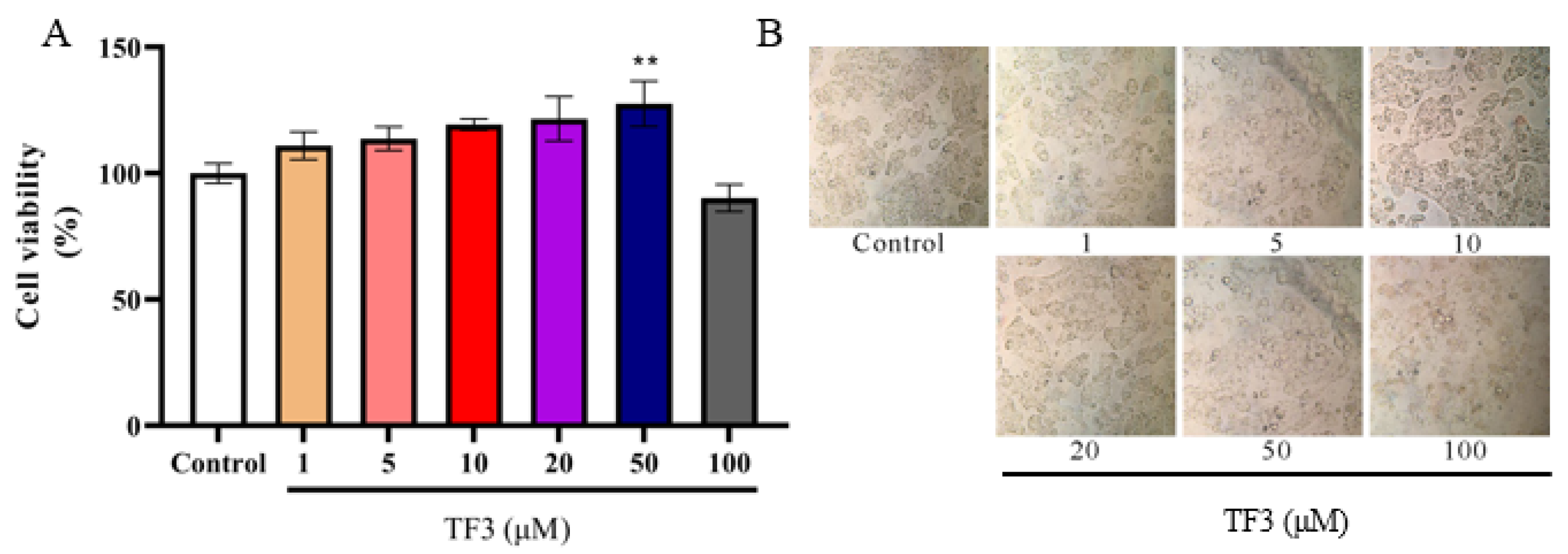
2.3. Cell Viability Measurement after TF3 Treatment
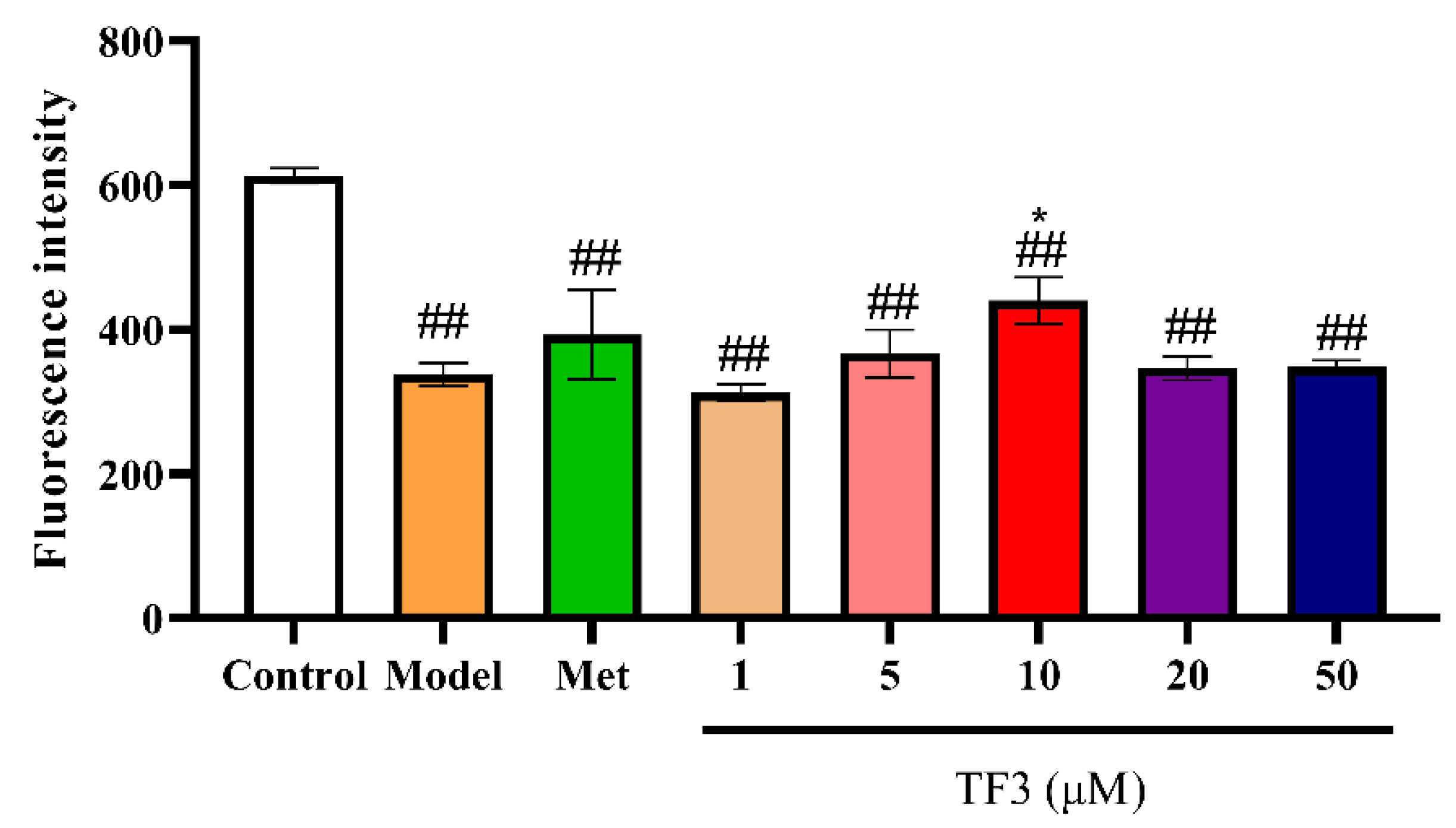
2.4. Glucose Absorption Determination in Insulin Resistance Model Cells
2.5. Zebrafish Culture and Diabetic Model Establishment
2.6. TF3 Toxicity Determination on Zebrafish
2.7. TF3 Hypoglycemic Effect Determination on Zebrafish Diabetic Model
2.8. Western Blot Analysis for Zebrafish Diabetic Model
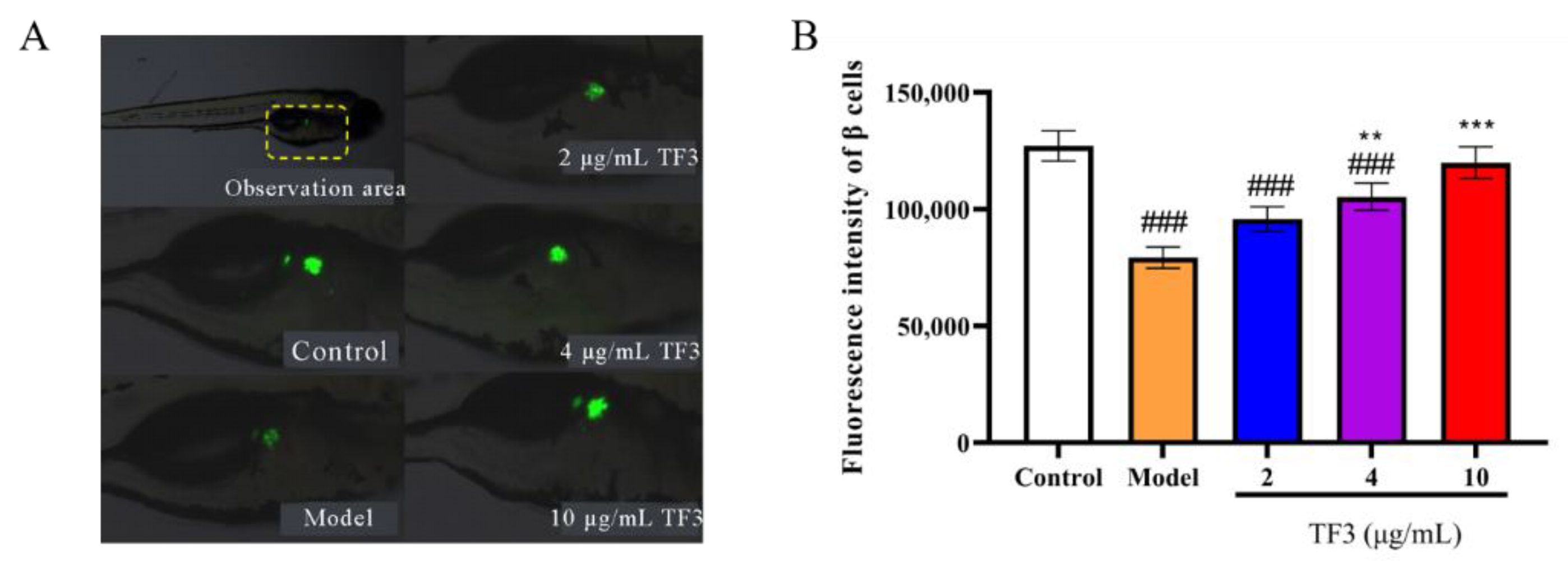
2.9. Zebrafish β Cell Fluorescence Intensity Determination
2.10. Statistical Analysis
3. Results
3.1. The Effect of TF3 on HepG2 Cells Viability and Morphology
3.2. TF3 Promotes Glucose Absorption Capacity in Glucose-Induced Insulin Resistance HepG2 Cells
3.3. The Toxicity of TF3 on Zebrafish
3.4. TF3 Reduces Glucose Level in Alloxan-Induced Diabetic Zebrafish
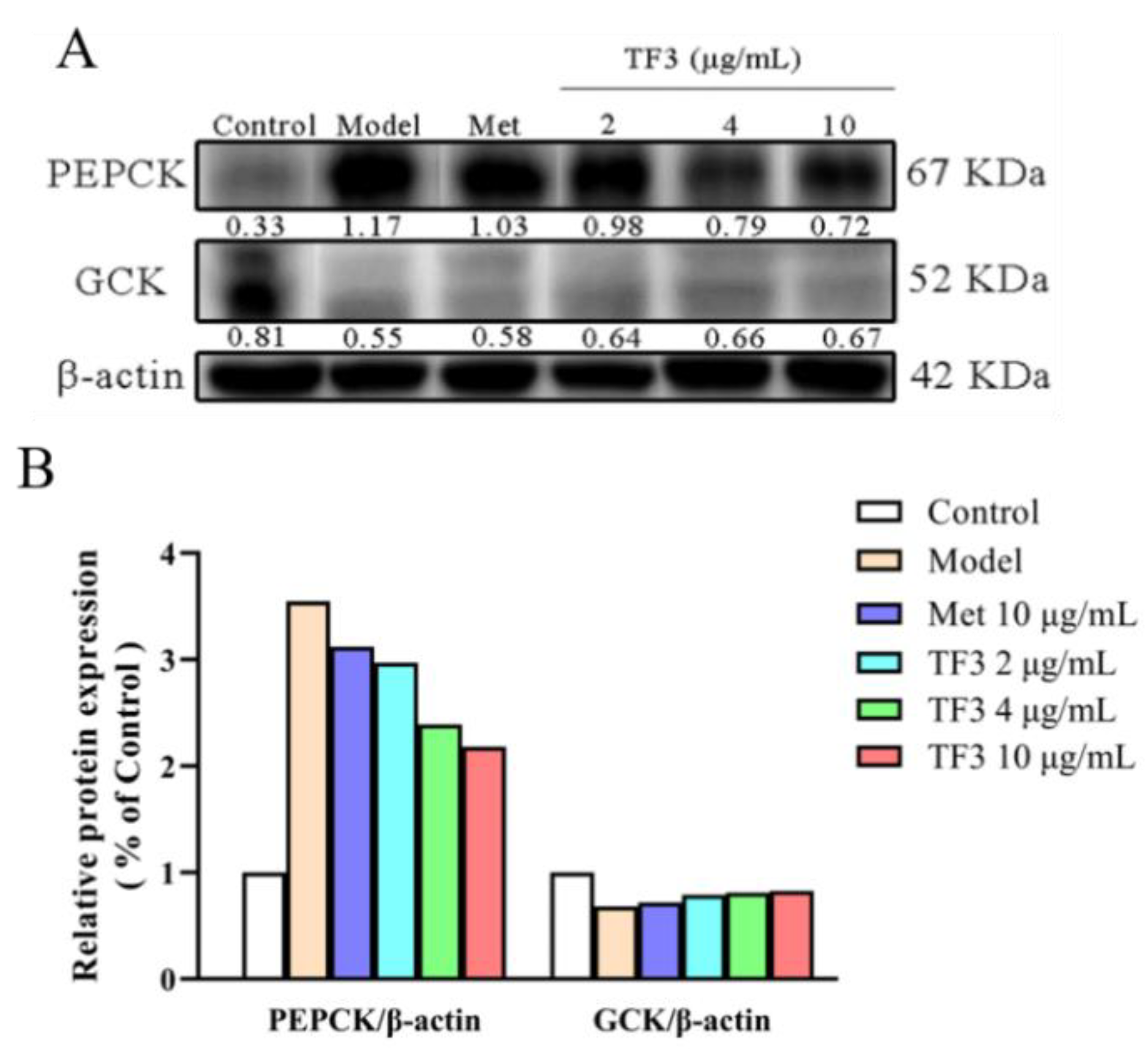
3.5. TF3 Inhibits Alloxan-Induced Diabetes via Regulating the Expression of PEPCK and GCK
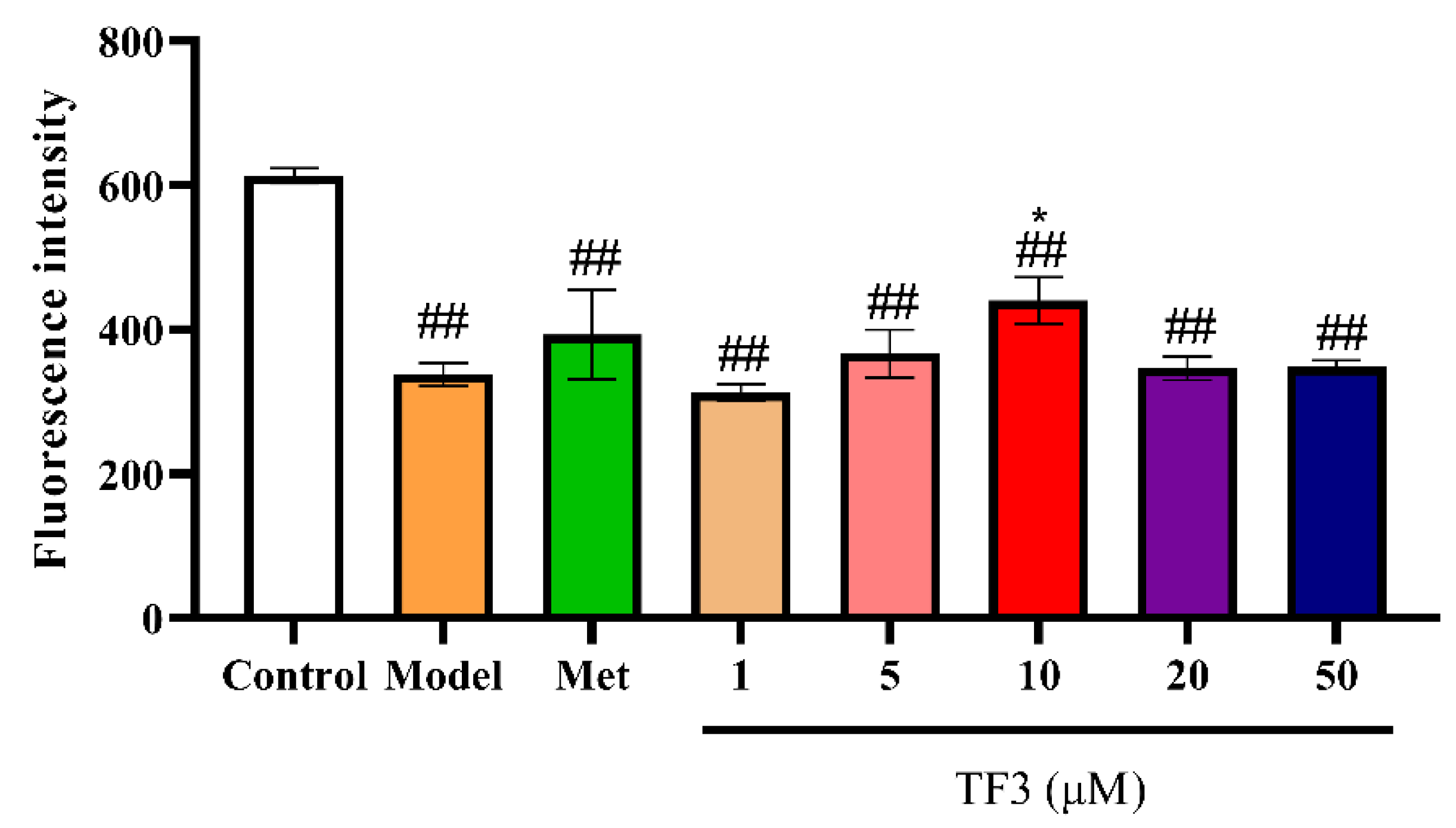
3.6. TF3 Promotes Zebrafish Islet β Cells Regeneration
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Vinik, A.I.; Vinik, E. Prevention of the complications of diabetes. Am. J. Manag. Care 2003, 9, S63–S80. [Google Scholar]
- Henagan, T.M.; Lenard, N.R.; Gettys, T.W.; Stewart, L.K. Dietary quercetin supplementation in mice increases skeletal muscle PGC1 alpha expression, improves mitochondrial function and attenuates insulin resistance in a time-specific manner. PLoS ONE 2014, 9, e89365. [Google Scholar] [CrossRef]
- Dujic, T.; Causevic, A.; Bego, T.; Malenica, M.; Velija-Asimi, Z.; Pearson, E.R.; Semiz, S. Organic cation transporter 1 variants and gastrointestinal side effects of metformin in patients with Type 2 diabetes. Diabet. Med. 2016, 33, 511–514. [Google Scholar] [CrossRef] [Green Version]
- Stull, A.J.; Cash, K.C.; Johnson, W.D.; Champagne, C.M.; Cefalu, W.T. Bioactives in blueberries improve insulin sensitivity in obese, insulin-resistant men and women. J. Nutr. 2010, 140, 1764–1768. [Google Scholar] [CrossRef]
- Jennings, A.; Welch, A.A.; Spector, T.; Macgregor, A.; Cassidy, A. Intake of anthocyanins and flavones are associated with biomarkers of insulin resistance and inflammation in women. J. Nutr. 2014, 144, 202–208. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.Y.; Li, W.; Xu, Y.; Jin, E.H.; Tu, Y.Y. Evaluation of the antioxidant effects of four main theaflavin derivatives through chemiluminescence and DNA damage analyses. J. Zhejiang Univ.-Sci. B 2011, 12, 744–751. [Google Scholar] [CrossRef] [Green Version]
- Satoh, T.; Igarashi, M.; Yamada, S.; Takahashi, N.; Watanabe, K. Inhibitory effect of black tea and its combination with acarbose on smallintestinal alpha-glucosidase activity. J. Ethnopharmacol. 2015, 161, 147–155. [Google Scholar] [CrossRef]
- Peluso, I.; Serafini, M. Antioxidants from black and green tea: From dietary modulation of oxidative stress to pharmacological mechanisms. Br. J. Pharmacol. 2017, 174, 1195–1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imran, A.; Butt, M.S.; Arshad, M.S.; Arshad, M.U.; Saeed, F.; Sohaib, M.; Munir, R. Exploring the potential of black tea based flavonoids against hyperlipidemia related disorders. Lipids Health Dis. 2018, 17, e57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gothandam, K.; Ganesan, V.S.; Ayyasamy, T.; Ramalingam, S. Antioxidant potential of theaflavin ameliorates the activities of key enzymes of glucose metabolism in high fat diet and streptozotocin-induced diabetic rats. Redox Rep. 2019, 24, 41–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, D.Y.; Xu, Y.; Mei, X.; Meng, Q.; Gao, Y.; Li, B.; Tu, Y.Y. Antiobesity and lipid lowering effects of theaflavins on high-fat diet induced obese rats. J. Funct. Foods 2013, 5, 1142–1150. [Google Scholar] [CrossRef]
- Tong, T.T.; Ren, N.; Soomi, P.; Wu, J.F.; Guo, N.; Kang, H.; Kim, E.; Wu, Y.Y.; He, P.M.; Tu, Y.Y.; et al. Theaflavins improve insulin sensitivity through regulating mitochondrial biosynthesis in palmitic acid-induced HepG2 cells. Molecules 2018, 23, 3382. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.Y.; Hou, Z.X.; Liu, D.Y.; Yang, X.B. Tartary buckwheat flavonoids protect hepatic cells against high glucose-induced oxidative stress and insulin resistance via MAPK signaling pathways. Food Funct. 2016, 7, 1523–1536. [Google Scholar] [CrossRef] [PubMed]
- Rubinstein, A.L. Zebrafish: From disease modeling to drug discovery. Curr. Opin. Drug Discov. Dev. 2003, 6, 218–223. [Google Scholar]
- Yan, H.; Su, R.B.; Gong, Z.H. Adult zebrafish as a model organism for assessing the effects of hallucinogenic drugs on behaviors. Chin. Pharmacol. Bull. 2014, 30, 1464–1468. [Google Scholar]
- Zang, L.Q.; Maddison, L.A.; Chen, W.B. Zebrafish as a model for obesity and diabetes. Front. Cell Dev. Biol. 2018, 6, e91. [Google Scholar] [CrossRef] [Green Version]
- Tu, Y.Y.; Tang, A.B.; Watanabe, N. The theaflavin monomers inhibit the cancer cells growth in vitro. Acta Biochim. Biolphys. Sin. 2004, 36, 508–512. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Rankin, G.O.; Tu, Y.Y.; Chen, Y.C. Inhibitory effects of the four main theaflavin derivatives found in black tea on ovarian cancer cells. Anticancer Res. 2016, 36, 643–651. [Google Scholar]
- Jiang, H.Y.; Cheng, Q.K.; Du, Q.Z.; Wang, H.F. Theaflavins in black tea. China Tea 1998, 20, 18–20. [Google Scholar]
- Yang, Z.Y.; Tu, Y.Y.; Xia, H.L.; Jie, G.L.; Chen, X.M.; He, P.M. Suppression of free-radicals and protection against H2O2-induced oxidative damage in HPF-1 cell by oxidized phenolic compounds present in black tea. Food Chem. 2007, 105, 1349–1356. [Google Scholar] [CrossRef]
- Chen, N.; Bezzina, R.; Hinch, E.; Lewandowski, P.A.; Cameron-smith, D.; Mathai, M.L.; Jois, M.; Sinclair, A.J.; Begg, D.P.; Wark, J.D.; et al. Green tea, black tea, and epigallocatechin modify body composition, improve glucose tolerance, and differentially alter metabolic gene expression in rats fed a high-fat diet. Nutr. Res. 2009, 29, 784–793. [Google Scholar] [CrossRef]
- Wang, K.B.; Liu, Z.H.; Huang, J.A.; Bekhit, A.E.; Liu, F.; Dong, X.R.; Gong, Y.S.; Fu, D.H. The inhibitory effects of pure black tea theaflavins on the growth of four selected human cancer cells. J. Food Biochem. 2011, 35, 1561–1567. [Google Scholar] [CrossRef]
- Ho, G.T.T.; Kase, E.T.; Wangensteen, H.; Barsett, H. Phenolic elderberry extracts, anthocyanins, procyanidins, and metabolites influence glucose and fatty acid uptake in human skeletal muscle cells. J. Agric. Food Chem. 2017, 65, 2677–2685. [Google Scholar] [CrossRef] [PubMed]
- Waltner-Law, M.E.; Wang, X.H.L.; Law, B.K.; Hall, R.K.; Nawano, M.; Granner, D.K. Epigallocatechin gallate, a constituent of green tea, represses hepatic glucose production. J. Biol. Chem. 2002, 277, 34933–34940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, F.J.; Zheng, X.D. Anthocyanin-rich mulberry fruit improves insulin resistance and protects hepatocytes against oxidative stress during hyperglycemia by regulating AMPK/ACC/mTOR pathway. J. Funct. Foods 2017, 30, 270–281. [Google Scholar] [CrossRef]
- Field, H.A.; Dong, P.D.S.; Beis, D.; Stainier, D.Y.R. Formation of the digestive system in zebrafish. II. pancreas morphogenesis. Dev. Biol. 2003, 261, 197–208. [Google Scholar] [CrossRef] [Green Version]
- Wan, H.Y.; Korzh, S.; Li, Z.; Mudumana, S.P.; Korzh, V.; Jiang, Y.J.; Lin, S.; Gong, Z.Y. Analyses of pancreas development by generation of gfp transgenic zebrafish using an exocrine pancreas-specific elastaseA gene promoter. Exp. Cell Res. 2006, 312, 1526–1539. [Google Scholar] [CrossRef]
- Benchoula, K.; Khatib, A.; Quzwain, F.M.C.; Mohamad, C.A.C.; Sulaiman, W.M.A.W.; Wahab, R.A.; Ahmed, Q.U.; Ghaffar, M.A.; Saiman, M.Z.; Alajmi, M.F.; et al. Optimization of hyperglycemic induction in zebrafish and evaluation of its blood glucose level and metabolite fingerprint treated with Psychotria malayana Jack leaf extract. Molecules 2019, 24, 1506. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.A.; Lee, S.H.; Lee, J.H.; Kang, N.; Oh, J.Y.; Seun-heui, C.; Ahn, G.; Ko, S.C.; Fernando, S.P.; Kim, S.J.; et al. A marine algal polyphenol, dieckol, attenuates blood glucose levels by Akt pathway in alloxan induced hyperglycemia zebrafish model. RSC Adv. 2016, 6, 78570–78575. [Google Scholar] [CrossRef]
- Lenzen, S.; Mirzaie-petri, M. Inhibition of glucokinase and hexokinase from pancreatic b-cells and liver by alloxan, alloxantin, dialuric acid, and t-butyl hydroperoxide. Biomed. Res. 1991, 12, 297–307. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.F.; Zhang, Y.B.; Wang, C.H.; Sun, Z.Y.; Li, L.F.; Cheng, S.Q.; Zhou, W.P. Overexpression of PCK1 gene antagonizes hepatocellular carcinoma through the activation of gluconeogenesis and suppression of glycolysis pathways. Cell. Physiol. Biochem. 2018, 47, 344–355. [Google Scholar] [CrossRef]
- Judy, W.V.; Hari, S.P.; Stogsdill, W.W.; Judy, J.S.; Naguib, Y.M.A.; Passwater, R. Antidiabetic activity of a standardized extract (Glucosol) from Lagerstroemia speciosa leaves in type II diabetics—A dose-dependence study. J. Ethnopharmacol. 2003, 87, 115–117. [Google Scholar] [CrossRef]
- Li, X.G.; Schmohl, F.; Qi, H.Z.; Bennewitz, K.; Tabler, C.T.; Poschet, G.; Hell, R.; Volk, N.; Poth, T.; Hausser, I.; et al. Regulation of gluconeogenesis by Aldo-keto-reductase 1a1b in zebrafish. Iscience 2020, 23, e101763. [Google Scholar] [CrossRef]
- Nam, Y.H.; Hong, B.N.; Rodriguez, I.; Park, M.S.; Jeong, S.Y.; Lee, Y.G.; Shim, J.H.; Yasmin, T.; Kim, N.W.; Koo, Y.T.; et al. Steamed ginger may enhance insulin secretion through KATP channel closure in pancreatic β-Cells potentially by increasing 1-Dehydro-6-Gingerdione content. Nutrients 2020, 12, 324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cha, S.H.; Zhang, C.Y.; Heo, S.J.; Jun, H.S. 5-Bromoprotocatechualdehyde combats against palmitate toxicity by inhibiting parkin degradation and reducing ROS-induced mitochondrial damage in pancreatic beta-cells. Antioxidants 2021, 10, 264. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.H.; Nuankaew, W.; Joo, S.W.; Kim, H.G.; Thi, N.N.; Lee, Y.G.; Kang, T.H.; Baek, N.I. Phenolic compounds from the aerial parts of Malva verticillata and their anti-diabetic effect. Nat. Prod. Commun. 2018, 13, 861–864. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Objects | Model | Treatment | Recovery Rate | Positive Control | Positive Control Group Recovery Rate |
|---|---|---|---|---|---|
| HepG2 cells | Insulin resistance model | 10 μM TF3 | 29.31% | Metformin | 17.24% |
| SD rats | Insulin resistance model | 100 mg/(Kg·bw) TFs (mainly TF3) | 54.93% | / | / |
| Zebrafish | Diabetic model | 2–20 μg/mL TF3 | 27.41–41.48% | Metformin | 18.05% |
| Transgenic β cell fluorescent zebrafish | β cell injury model | 4–10 μg/mL TF3 | 20.80–51.29% | / | / |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, H.; Wu, Y.; Kim, E.; Pan, H.; He, P.; Li, B.; Chen, Y.C.; Tu, Y. Simultaneous Tests of Theaflavin-3,3′-digallate as an Anti-Diabetic Drug in Human Hepatoma G2 Cells and Zebrafish (Danio rerio). Nutrients 2021, 13, 4379. https://doi.org/10.3390/nu13124379
Zhou H, Wu Y, Kim E, Pan H, He P, Li B, Chen YC, Tu Y. Simultaneous Tests of Theaflavin-3,3′-digallate as an Anti-Diabetic Drug in Human Hepatoma G2 Cells and Zebrafish (Danio rerio). Nutrients. 2021; 13(12):4379. https://doi.org/10.3390/nu13124379
Chicago/Turabian StyleZhou, Hui, Yuanyuan Wu, Eunhye Kim, Haibo Pan, Puming He, Bo Li, Yi Charlie Chen, and Youying Tu. 2021. "Simultaneous Tests of Theaflavin-3,3′-digallate as an Anti-Diabetic Drug in Human Hepatoma G2 Cells and Zebrafish (Danio rerio)" Nutrients 13, no. 12: 4379. https://doi.org/10.3390/nu13124379
APA StyleZhou, H., Wu, Y., Kim, E., Pan, H., He, P., Li, B., Chen, Y. C., & Tu, Y. (2021). Simultaneous Tests of Theaflavin-3,3′-digallate as an Anti-Diabetic Drug in Human Hepatoma G2 Cells and Zebrafish (Danio rerio). Nutrients, 13(12), 4379. https://doi.org/10.3390/nu13124379