1. Introduction
Polyphenols are found mainly in plant-derived foods and beverages and provide the taste and color of plant foods. Moreover, epidemiological studies have reported a greater reduction in cardiovascular risk and cancer associated with diets rich in polyphenols [
1,
2,
3].
Delphinidin (2-(3,4,5-trihydroxyphenyl)chromenylium-3,5,7-triol) is an anthocyanin abundantly identified in pigmented vegetables and fruits, particularly berries and red grapes. We previously reported that delphinidin possesses the same pharmacological profile as a total extract of red wine polyphenolic compounds to promote the increase of intracellular calcium concentration and activation of tyrosine kinases [
3], leading to endothelial nitric oxide (NO) production subsequent to estrogen receptor alpha (ERα) stimulation [
4]. In addition, we reported that delphinidin via ERα acts as an immuno-modulatory and anti-inflammatory molecule that can alter T lymphocyte proliferation and differentiation in patients with cardiovascular risk factors [
5].
Finally, we demonstrated that delphinidin displays anti-angiogenic properties, both on in vitro and in vivo angiogenesis models, and reduces in vivo tumor growth of melanoma [
6,
7,
8,
9,
10]. Indeed, delphinidin inhibits endothelial cell proliferation through the involvement of cyclin D1- and A-dependent pathways [
6,
7]. We also reported a possible association between inhibition of VEGF-induced mitochondrial biogenesis through the Akt pathway by delphinidin and its anti-angiogenic effect [
8]. Moreover, delphinidin reduces tumor growth of melanoma tumor cell in vivo by acting specifically on endothelial cell proliferation. The mechanism implies an association between inhibition of VEGF-induced proliferation via VEGFR2 signaling, MAPK, PI3K and at transcription level on CREB/ATF1 factors, and the inhibition of phopsphodiesterase2 [
9]. Most interestingly, high doses of delphinidin decreased neovascularization in an in vivo model of angiogenesis triggered by ischemia using a rat model of femoral artery ligature [
10]. Together, these data show that delphinidin is a promising compound to prevent pathologies associated with cardiovascular disorders and tumorigenesis.
However, delphinidin is less potent to induce these beneficial effects compared to total red wine polyphenol extracts, especially in inducing endothelium-dependent NO-mediated vasodilatation [
11]. Indeed, delphinidin is light-sensitive and stable only at pH < 3; therefore, it degrades rapidly under physiological conditions. Moreover, delphinidin is poorly absorbed, and thus its modest bioavailability and stability reduce its effects both in vitro and in vivo. The measurement of delphinidin and its conjugated metabolites in plasma indicates its low bioavailability [
12]. Hence, it is important to find new strategies to enhance delphinidin bioavailability and efficacy.
One strategy to overcome such problems is the use of extracellular vesicles (EVs) as a drug delivery system. We recently found that EVs, including large and small EVs (sEVs), are nanostructures originating from different subcellular compartment properties, overcoming the limitations of classical nano-formulations. sEVs decrease instability and immunogenicity, improve bioavailability and target selectivity [
13]. Some reports underscore the protective effects of EVs released by cells treated with polyphenols. Indeed, sEVs enriched with miR-21 from cells treated with curcumin decreased tumor cell growth and angiogenesis, corrected endothelial permeability and decreased the cell viability of different cancer cell lines [
14]. In addition, miR-16-enriched sEVs from cells treated with epigallocatechin gallate suppressed tumor growth [
15].
In the present study, we took advantage of sEV properties to enhance both the stability and efficacy of delphinidin. sEV-loaded delphinidin induced angiogenesis inhibition using human aortic endothelial cells (HAoECs). Delphinidin content in terms of metabolites within these EVs was also determined.
2. Materials and Methods
2.1. Cell Culture
HAoECs (Promocell, Heidelberg, Germany) were cultured at 37 °C and 5% CO2 in endothelial cell growth medium MV2 (Promocell) supplemented with 1% penicillin/streptomycin (Sigma-Aldrich, St. Quentin Fallavier, France). Cells were trypsinized at 70/80% confluence and were used between passage 3 and 6 for all experiments.
The JAWS II dendritic cell line was purchased from the American Type Culture Collection (CRL-1194; ATCC; Manassas, VA, USA). JAWS II cells were grown at 37 °C and 5% CO2 in a complete culture medium composed of alpha minimum essential medium (Lonza; Basel, Switzerland) containing ribonucleosides and desoxyribonucleosides and supplemented with 20% fetal bovine serum (FBS) (Gibco, Life Technologies; Grand Island, NY, USA), 4 mM L-glutamine (Lonza), 1 mM sodium pyruvate (Lonza), 1% penicillin/streptomycin (penicillin/-streptomycin, Sigma-Aldrich) and 5 ng/mL murine GM-CSF (Miltenyi Biotec; San Diego, CA, USA). Cells were trypsinized at 70/80% confluence and were used between passage 8 and 16 for all experiments.
2.2. sEV Isolation
JAWS II cells were seeded at a density of 5 × 106 cells in a T175 cell culture flask in complete growth medium, and they were starved in FBS before any isolation. Cell medium was centrifuged at 300× g and 2000× g for 10 min to remove cells and cell debris, respectively. The resultant supernatant was centrifuged at 20,000× g for 30 min to exclude large EVs. The supernatant was centrifuged at 200,000× g (Optima MAX-XP ultracentrifuge and MLA-50 rotor, Beckman Coulter, Villepinte, France) for 2 h to pelletize sEVs. Then, sEVs were washed in phosphate-buffered saline (PBS) (NaCl 137 mM, KCl 2.7 mM, Na2HPO4 10 mM, KH2PO4 1.8 mM, pH = 7.4) and recentrifuged at 200,000× g for 2 h. Finally, sEV pellets were resuspended in 1 mL of PBS and stored at 4 °C until subsequent use. The amount of sEVs was determined using the method of Lowry, with bovine serum albumin (Sigma-Aldrich) as the standard. sEVs were used at 10 µg/mL.
2.3. Delphinidin Loading
Delphinidin was prepared in water at pH = 2 with 0.1% DMSO in order to reach the concentration of 10 µg/mL. The sEVs were added (2 mg), and the solution was stirred and then vortexed for 10 min. After 2 h of ultracentrifugation at 200,000× g, the obtained pellet was reconstituted in 1 mL of 0.1% DMSO or PBS. Delphinidin absorbance was measured at 530 nm, and a standard curve with different concentrations (0.1 to 10 µg/mL) of free delphinidin was performed. The percentage of the efficacy of the loading of sEVs was 9%, independently of the concentration of delphinidin used (data not shown). Thus, the amount of delphinidin was adjusted to obtain the desired concentration (0.1 to 5 µg/mL) within 10 µg/mL sEVs. To remove free delphindin, these vesicles were washed twice.
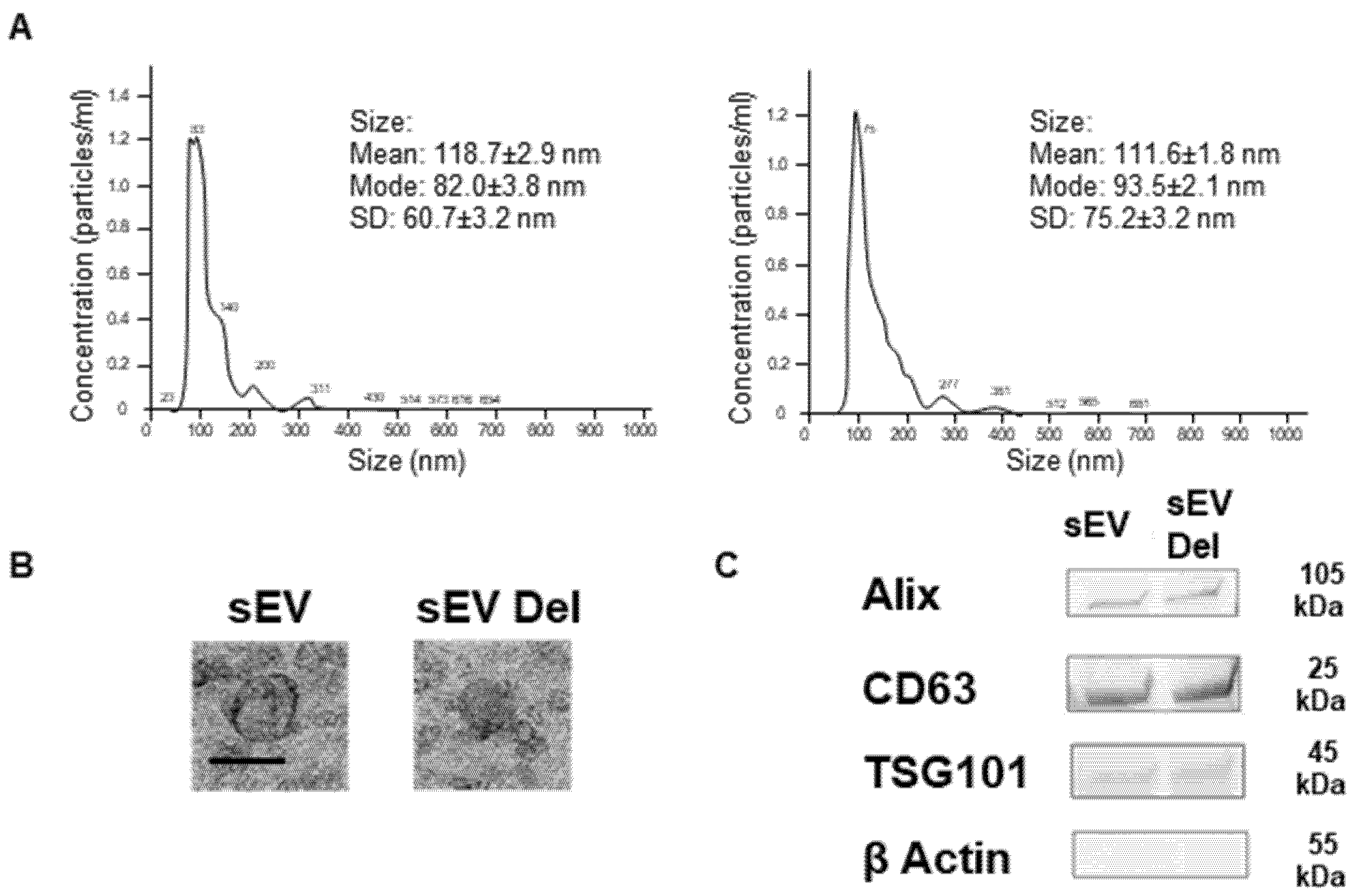
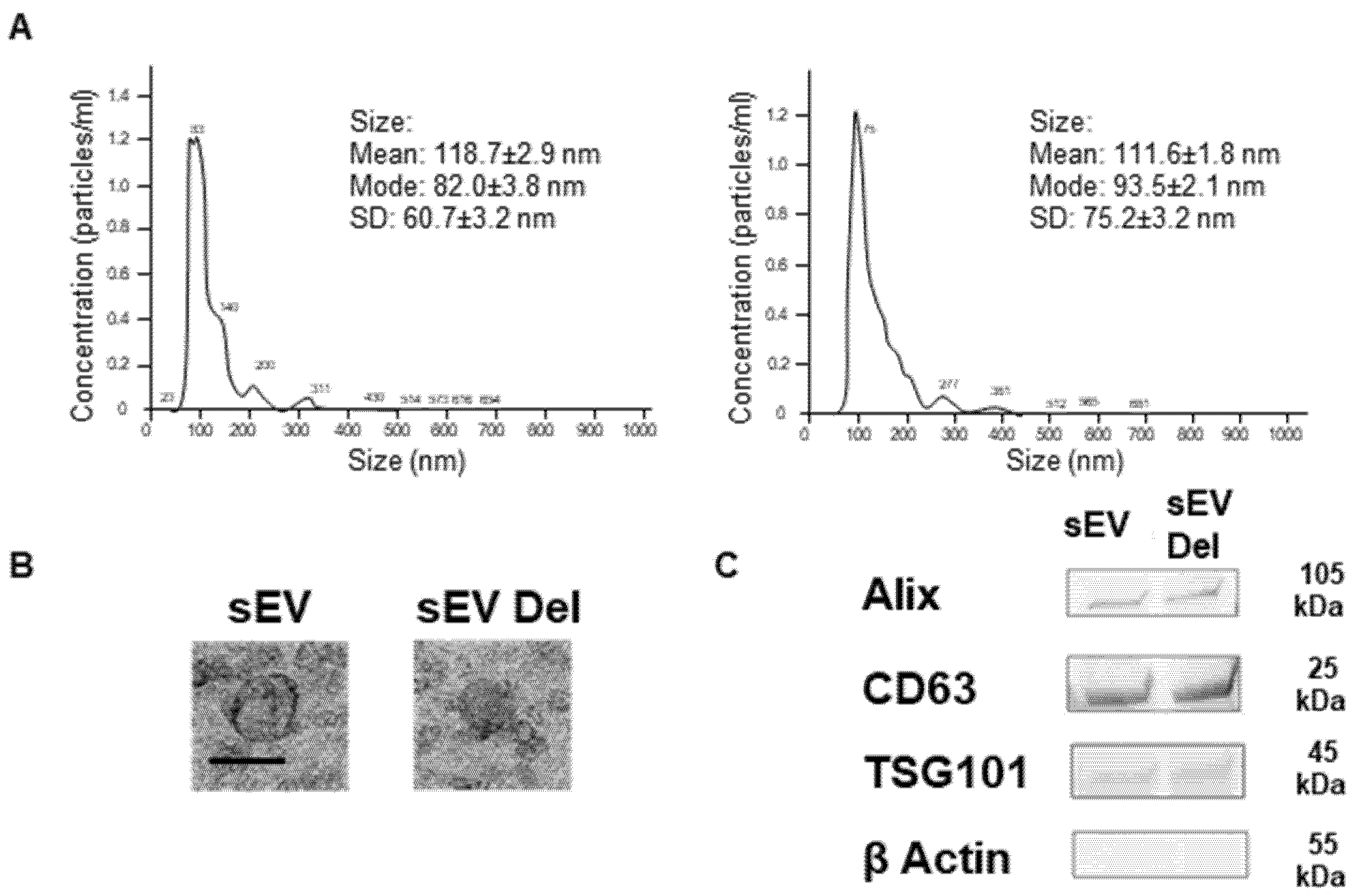
2.4. Nanoparticle Tracking Analysis (NTA)
sEV samples were diluted in sterile NaCl 0.9%, and size distribution was analyzed using the NanoSight NS300 (Malvern Instruments Ltd., Malvern, UK). Videos were recorded. NTA software determined the size distribution using the Stokes-Enstein equation.
2.5. Transmission Electronic Microscopy
sEVs were first fixed overnight at 4 °C with 2.5% glutaraldehyde (LFG Distribution, Lyon, France) in 0.1 M PBS. Then, sEVs were washed two times in PBS by 100,000× g centrifugation for 70 min. sEVs were deposited on copper grids for 2 min and negatively stained with 20 μL of uranyl acetate 5% (diluted in ethanol 50%) for 30 s. Grids were then observed with a Jeol JEM 1400 microscope (Jeol, Croissy sur Seine, France) operated at 120 keV.
2.6. Determination of Delphinidin Metabolites within sEVs
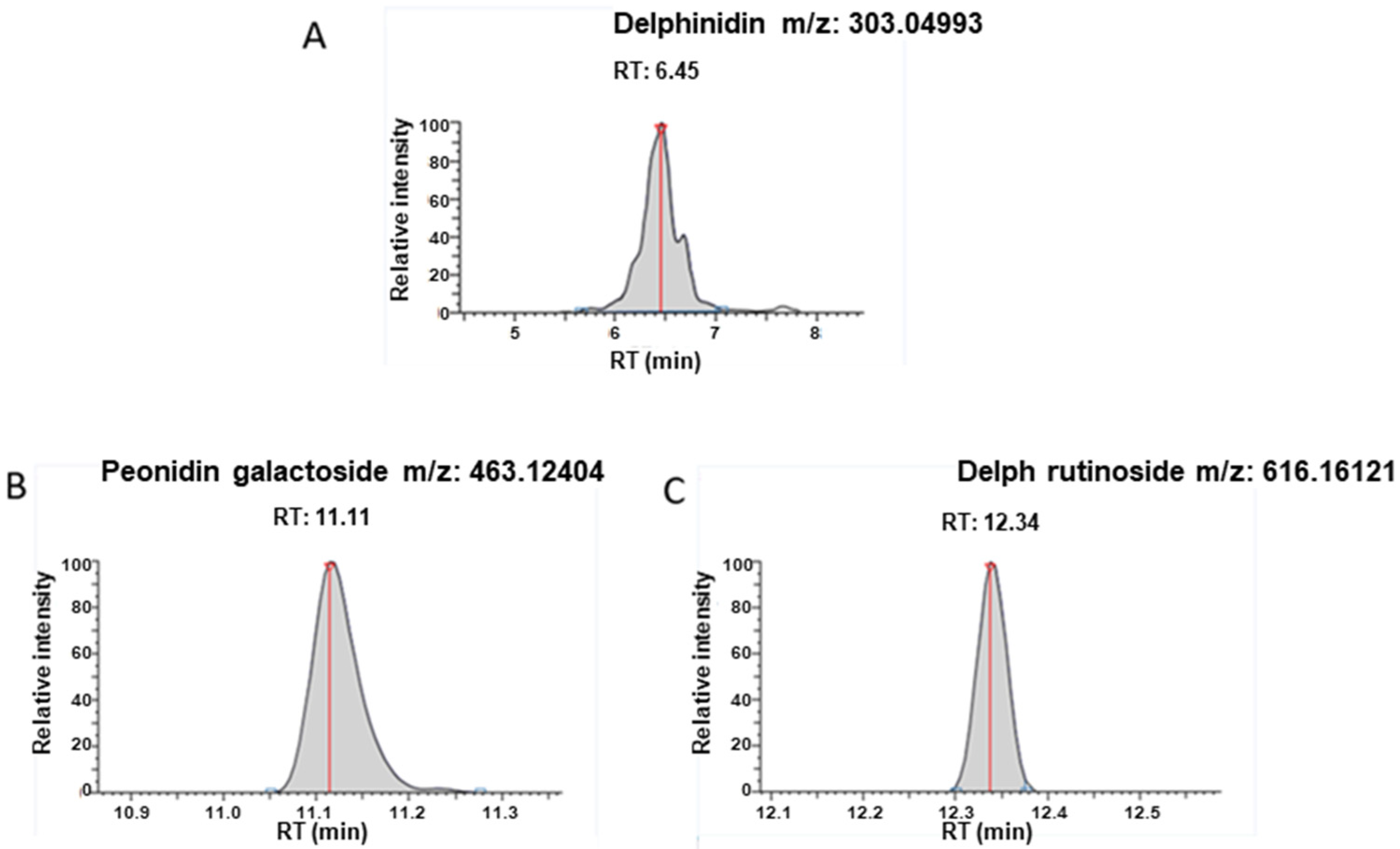
Sample preparation was as follows: 250 µL methanol (MeOH) was added to 10 µg sEVs reconstituted in PBS, and samples were subjected to a 20 min ultrasonication. Two hundred µL of MeOH was further added, and samples were centrifuged (10,000× g, 10 min, 4 °C) and evaporated in a miVac duo concentrator (Genevac Ltd., Ipswich, UK). The dry extract was reconstituted with 200 µL LC-MS grade water containing 1% formic acid. The mixture was subjected to a second centrifugation (10,000× g, 5 min, 4 °C) prior to ultra-high-performance liquid chromatography coupled to high-resolution mass spectrometry (UHPLC-HRMS) analysis in order to analyze delphinidin metabolites with accurate mass measurements.
The chromatographic separation was achieved with a Kinetex® 1.7 µm XB C18, 150 × 2.1 mm column together with the corresponding SecurityGard C18 column (Phenomenex®). Mobile phases consisted of H2O in channel A and acetonitrile in channel B, both containing 0.1% formic acid. The elution gradient (A:B, v/v) was as follows: hold initial conditions 95:5 for 2 min, followed by a linear gradient from 95:5 to 0:100 over a 6 min period, hold at 0:100 for 3 min, return to initial conditions 95:5 and hold these conditions for 3.5 min. A constant flow rate of 0.300 mL/min was used; the injection volume was 10 μL.
Full scan and targeted SIM mass spectra were acquired in positive ionization mode, using resolution 70,000 Full Width at Half Maximum (FWHM) with automatic gain control (AGC) target of 3 × 106 ions and a maximum ion injection time (IT) of 200 ms. Data-dependent MS/MS experiments were acquired in ‘Top5′ data-dependent mode.
Metabolites reported in the literature [
16,
17,
18] were monitored: Delphinidin, aldehyde, phloroglucinol aldehyde, gallic acid, chalcone, petunidin-3-galactoside, petunidin-3-arabinoside, petunidin 3-O-rutinoside, delphinidin-3-arabinoside, delphinidin-3-galactoside, delphinidin 3-O-(6-coumaroylglucoside), delphinidin 3-O-β-rutinoside, cyanidin-3-galactoside, cyanidin 3-O-β-rutinoside, Peonidin-3-galactoside and malvidin-3-galactoside.
Daily instrument calibration was performed by infusion of Pierce LTQ Velos ESI positive/negative calibration kits as recommended by the manufacturer. Xcalibur 2.2 software (Thermo Fisher Scientific, San Jose, CA, USA) was used for data acquisition, and TraceFinder 3.0 software (Thermo Fisher Scientific) was employed for data processing.
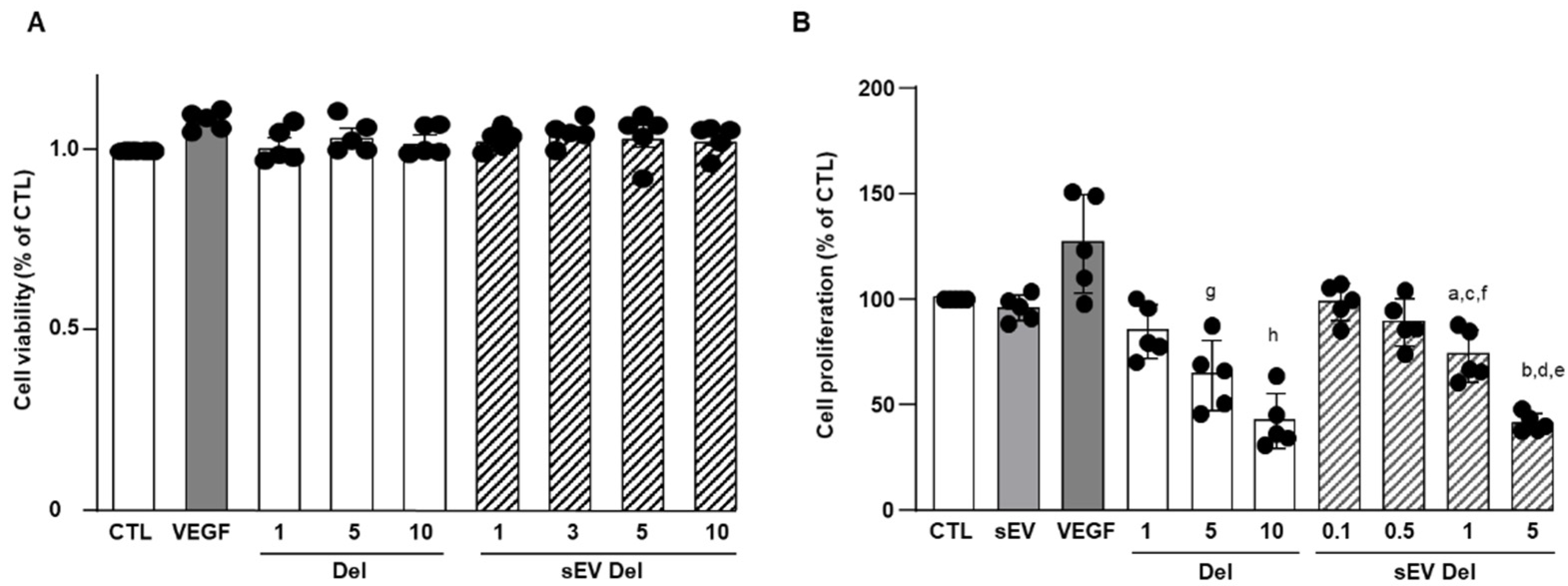
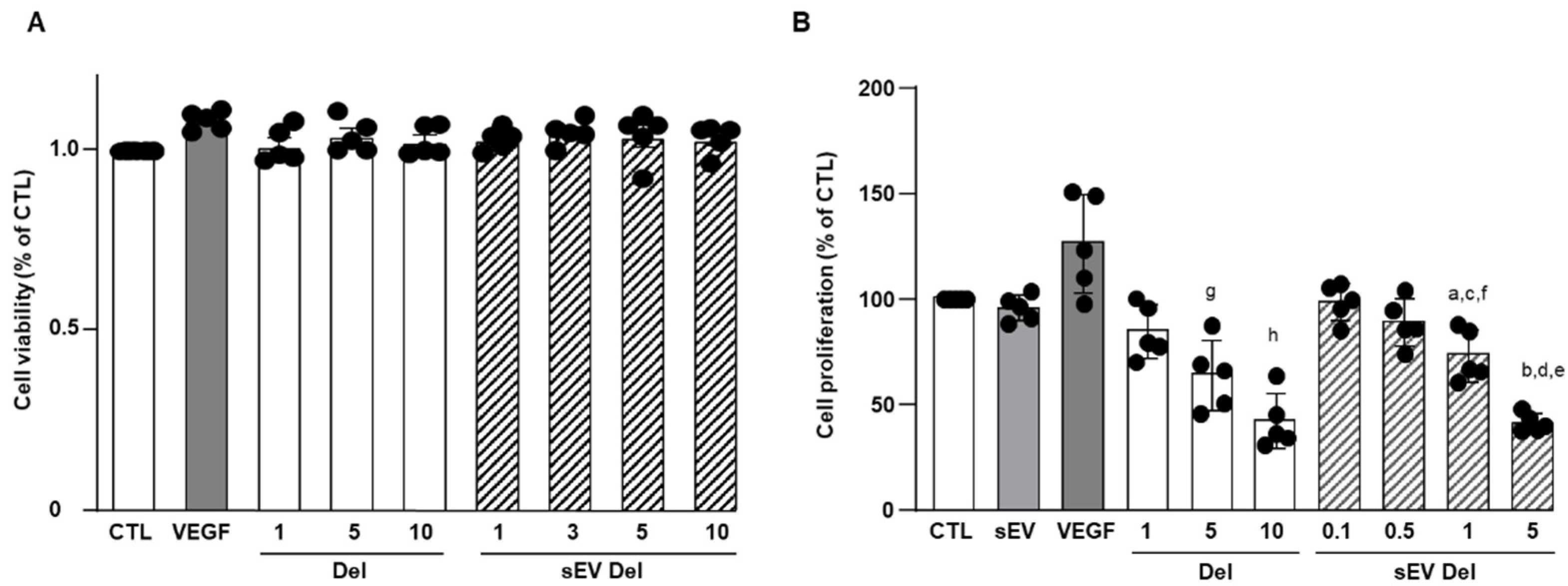
2.7. Cell Viability Assay
1 × 104 HAoECs were seeded onto a 96-well plate and cultured for 24 h and treated with delphinidin (1 to 10 µg/mL). Then, 5 μg/mL of 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)- 2-(4-sulfophenyl)-2H- tetrazolium (MTS reagent, Promega, WI, USA) was added into each well and incubated at 37 °C for 120 min. The absorbance was measured on a CLARIOstar® (BMG LABTECH, Ortenberg, Germany) spectrophotometer at 490 nm.
2.8. Proliferation Assay
Proliferation assays were conducted using CyQUANT Cell proliferation Assay kit (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s recommendations. Briefly, 1.5 × 104 cells were seeded in a 96-well plate. Cells were serum-starved for 2 h and then treated with delphinidin, native sEVs or sEVs loaded with delphinidin at different concentrations. After 24 h of incubation, cells were washed with PBS, and dye-binding solution was added. Cells were incubated at 37 °C for 30 min. A fluorescent microplate reader (CLARIOstar®, BMG LABTECH, Ortenberg, Germany) with filters for 485 nm excitation and 530 nm emission was used for fluorescence measurement.
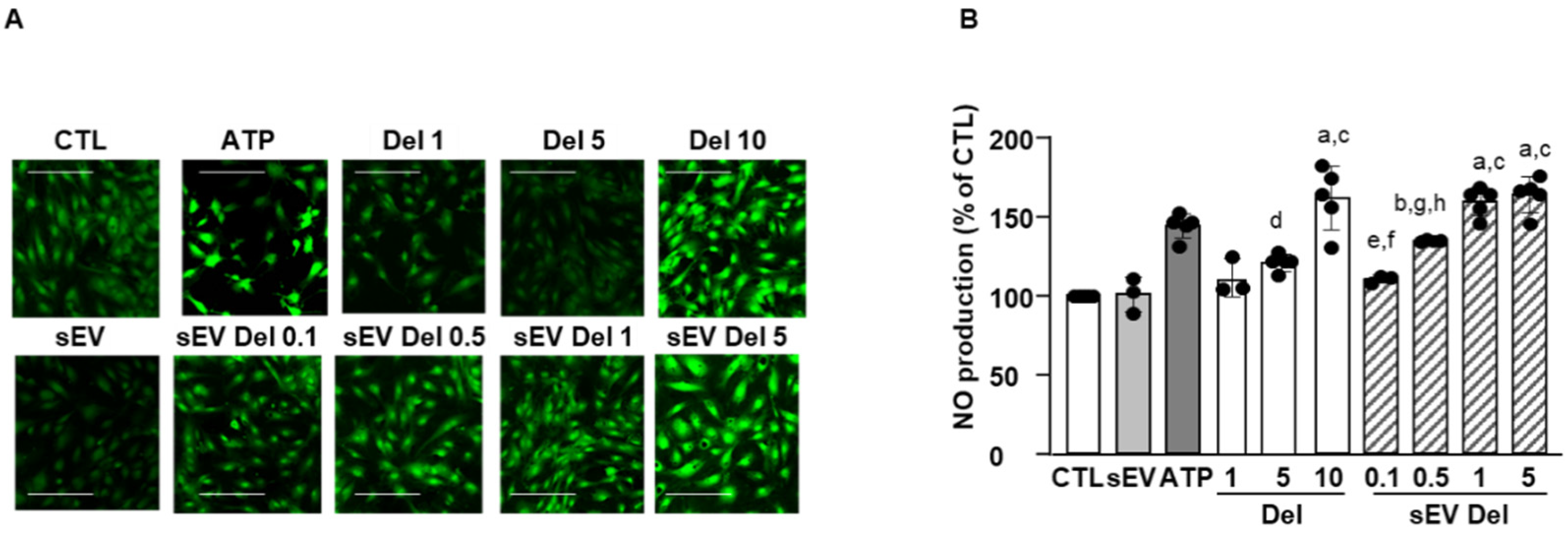
2.9. NO Production Assay
HAoECs were seeded on a 8-well slide (Ibidi, Gräfelfing, Germany) at a rate of 3 × 104 cells per well (i.e., 3 × 104 cells/cm2) in 300 μL of medium. At 70–80% confluence, cells were stimulated for 24 h with delphinidin, native sEVs or sEV-loaded delphinidin. Adenosine triphosphate (ATP) was used as a positive control (10 μM, Sigma-Aldrich) to stimulate the production of NO. After 24 h, medium of each well was removed, and the diaminofluoroscein diacetate (DAF-2 DA) probe was added (5 μM for 30 min, Santa Cruz Biotechnology, Santa Cruz, CA, USA). Then, the wells were washed with PBS. Cells were fixed with paraformaldehyde (4%, 20 min). Fluorescence was read by confocal microscopy (Zeiss, Jena, Germany, LSM700). Four pictures were acquired, and ImageJ software was used for quantification.
2.10. Matrigel Assay
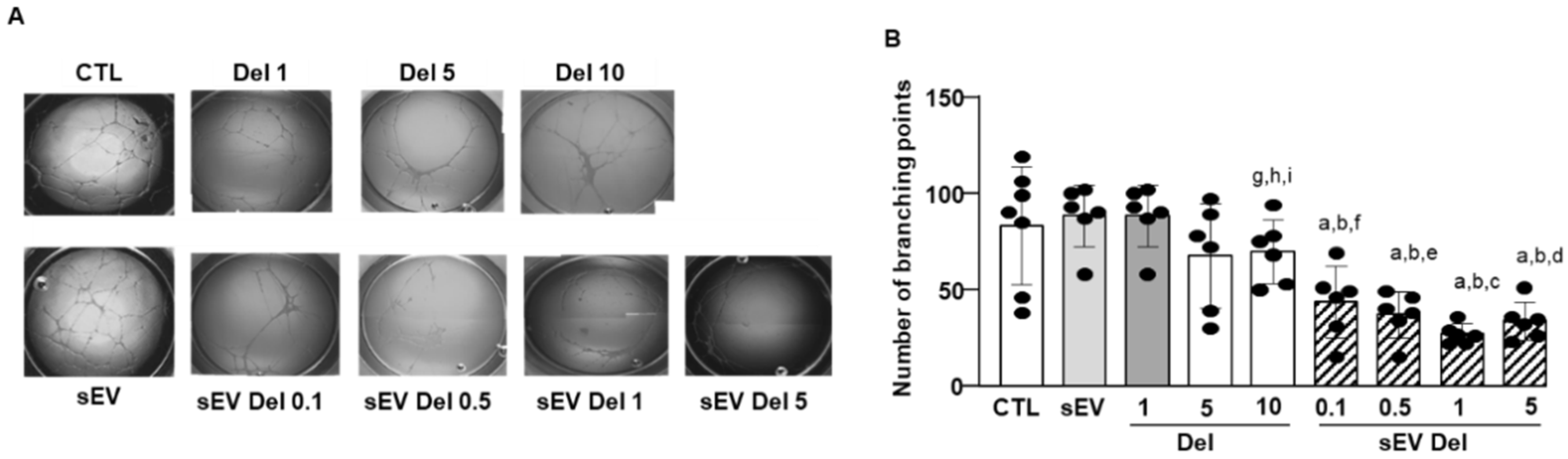
HAoECs were seeded in wells coated with Matrigel® (gel of extracellular matrix of murine sarcoma of Engelbreth-Holm-Swarm, Sigma-Aldrich). Briefly, 10 µL of liquid Matrigel® was placed in each well of a 15-well Ibidi µ-slide Angiogenesis plate (Ibidi) and then incubated for 45 min at 37 °C to form a gel. HAoECs were then seeded and incubated at 37 °C and 5% CO2 for 45 min before treatments with either delphinidin, native sEVs or sEV-loaded delphinidin, followed by an incubation of 12 to 14 h at 37 °C and 5% CO2. The formation of “capillary-like structures” was observed with an optical microscope (Olympus CK40). Quantification was performed by measuring the number of capillary-like structures using Image J software.
2.11. Statistical Analysis
Results are expressed as mean ± SEM. Significance of the differences between groups was determined by analysis of variance (ANOVA), followed by Tukey’s multiple comparisons test. p-values of < 0.05 were considered significant.
4. Discussion
The current study shows that the delphinidin loaded within sEVs was obviously more potent than free delphinidin regarding its ability to release endothelial NO, to inhibit endothelial proliferation and to reduce capillary-like structures. In addition to delphinidin found into sEVs, the analysis of delphinidin metabolites within the sEVs showed the presence of two metabolites (i.e., delphinidin 3-O-ß-rutinoside and peonidin-3-galactoside) present in delphinidin samples. Thus, delphinidin degraded into the same metabolites in its free form or when loaded into sEVs; however, these were more potent in acting on endothelial cells. Of importance, when encapsulated within sEVs, delphinidin (used as a generic term and encompassing natural metabolites delphinidin 3-O-ß-rutinoside and peonidin-3-galactoside) was 2-fold, 10-fold and 100-fold more potent than free delphinidin regarding endothelial proliferation, endothelial NO production and capillary-like formation. Thus, sEV-loaded delphinidin exerts effects on different steps leading to angiogenesis. These results indicate that sEVs may be considered as a promising delivery of delphinidin as an innovative approach to target diseases associated with increased angiogenesis, including cancer, atherosclerosis and diabetic retinopathies.
The encapsulation of polyphenols to protect them from degradation is a natural phenomenon. Indeed, it has been shown that plants rich in polyphenols produce EVs carrying these molecules. For instance, the flavonoid glycoside naringin and its metabolite, naringenin, are found in grapefruit-derived EVs [
12]. Moreover, nanoparticles derived from plants can be used as vectors for other molecules of interest. Indeed, it has been reported that grapefruit-derived nanoparticles loaded with a STAT3 inhibitor inactivate STAT3 in GL26 tumor cells and improve survival rates of mice [
19]. Another strategy for the encapsulation of polyphenols is to use vesicles derived from mammalian cells. A recent report evaluated the exosomal formulation of anthocyanidins against different types of cancer [
19]. sEVs harvested from raw bovine milk loaded with a mixture of cyanidin, delphinidin, petunidin, peonidin and malvidin increase the anti-proliferative activity of anthocyanidins against six different types of cancer cells via the inhibition of TNFα-induced activation of NF-κB [
20]. Indeed, the effects of sEVs loaded with anthocyanidins are more effective than those obtained by free anthocyanidins. This method has advantages; however, it remains risky. Indeed, the use of EVs from mammalian cells can cause immune reactions. sEVs derived from immature human dendritic cells did not induce any toxicity, and the immature nature of dendritic cells induced low immunogenicity [
21,
22]. To our knowledge, this is the first time that a loading efficiency for delphinidin within JAWS II sEVs has been described. Loading of delphinidin into the sEVs (9%) protects and probably limits its degradation into metabolites under the experimental conditions used. The mechanisms involved require further study. The metabolites found in the sEVs, such as delphinidin 3-O-β-rutinoside or peonidin-3-galactoside, are also found in in vivo experiments with delphinidin [
14,
15]. Previous works have reported that degradation products of delphinidin have potent biological activities, including anti-cancer and anti-inflammatory activities [
20]. Among phenolic acids, gallic acid is mostly formed by the degradation of delphinidin in culture media [
23]. In the present study, we found that metabolites detected in sEV-loaded delphinidin were identical to those detected from free delphinidin [
18]. Although the exact proportion of metabolites encapsulated in these sEVs was not determined, they were more effective on target cells than metabolites alone. Thus, delphinidin and its metabolites were probably more stable and protected from degradation.
We previously reported that, in bovine aortic endothelial cells, delphinidin stimulates NO release by increasing intracellular Ca
2+ concentrations via the increase of superoxide anion formation. This was associated with increased tyrosine phosphorylation of several intracellular proteins, resulting in endothelium-dependent vasodilatation [
24,
25]. Delphinidin interacts directly with the activator site of ERα, leading to the activation of endothelial NO-synthase, NO production and endothelium-dependent vasorelaxation [
4]. In the present study, sEV-loaded delphinidin was 10 times more potent than free delphinidin; thus, it would probably be more effective in correcting the NO-endothelial dysfunction associated with cardiovascular diseases, including hypertension, stroke or metabolic diseases [
3].
We previously reported that upregulation of the NO pathway is not responsible for the antiproliferative effect of delphinidin. Indeed, delphinidin inhibits endothelial cell proliferation by the activation of ERK-1/-2 pathway, leading to cell cycle arrest and accumulation of cells in the G0/G1 phase via down-regulation of cyclin A and D1 expression and an upregulation of p27kip1 [
6,
7]. We also found that delphinidin reduces tumor growth of melanoma cells in vivo by acting specifically on endothelial cell proliferation via the inhibition of VEGFR2 signaling, MAPK, PI3K and at transcription level on CREB/ATF1 factors, and the inhibition of phosphodiesterase 2 [
9]. In the present study, delphinidin-loaded sEVs were two-fold more potent than free delphinidin in inhibiting endothelial proliferation. Thus, these results indicate that delphinidin-loaded sEVs are a promising approach to prevent pathologies associated with excess endothelial proliferation and, therefore, generation of the vascular network such as plaque development and stability in atherosclerosis and tumor development in cancer.
In concordance with these findings, we show that delphinidin decreases capillary-like formation in an experimental model of angiogenesis. Interestingly, when encapsulated within sEVs (even at a loading as low as 9%), delphinidin was 100-fold more potent than free delphinidin in decreasing capillary-like formation.
Limitation of the study: The anti-angiogenic potential exhibited by many natural compounds contained in many Mediterranean diet constituents, including delphinidin, makes this dietary pattern especially interesting as a source of chemopreventive agents, defined within the angioprevention strategy. This has been recently reviewed by Martinez-Podeva et al. [
26]. Delphinidin appears to be as potent as other flavonoids in inducing anti-angiogenic properties. Although abundant in the diet, anthocyanins in general, and delphinidin in particular, are poorly absorbed. One consequence of the poor bioavailability of anthocyanins is that many effects observed in vitro (e.g., inhibition of COX-2) are unlikely to occur in vivo, which is not the case for delphinidin, based on our former studies [
6,
7,
8,
9,
10]. However, additional studies using delphinidin encapsulated in sEV are needed to confirm the increase in the anti-angiogenic properties of this approach in vivo.
In summary, sEV-loaded delphinidin increased the efficacy of delphinidin 100-fold for proliferation, 10-fold for NO and 2-fold for capillary-like formation. Thus, sEVs either protected delphinidin and its metabolites from degradation or some unidentified delphinidin metabolites contained in the sEVs were more potent. The differential potency obtained for proliferation, NO production and angiogenesis supports the hypothesis that delphinidin-loaded sEVs exert effects on different steps leading to angiogenesis. Nevertheless, we provide evidence that we optimized delphinidin efficacy, probably by reducing its degradation and increasing its delivery when encapsulated in EVs. Thus, delphinidin-loaded sEVs represent a powerful delivery system to decrease angiogenesis in endothelial cells, with no unwanted side effects, knowing the low bioavailability of this compound. We underscore an innovative therapeutic strategy based on bio-engineered EVs as vectors of delphinidin in helping to increase its potential health benefit to target angiogenesis-related diseases, including cancer, which could eventually be extended to further diseases with excess vascularization.


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}