Laminaria japonica Suppresses the Atopic Dermatitis-Like Responses in NC/Nga Mice and Inflamed HaCaT Keratinocytes via the Downregulation of STAT1


Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation and Chemical Profiling of LJWE
2.3. Cell Viability and Treatments
2.4. Western Blotting
2.5. Animal Study
2.6. Analysis of Dermatitis Severity, Scratching Behavior, and Trans-Epidermal Water Loss (TEWL)
2.7. Estimation of Inflammatory Chemokines
2.8. Statistical Analysis
3. Results and Discussion
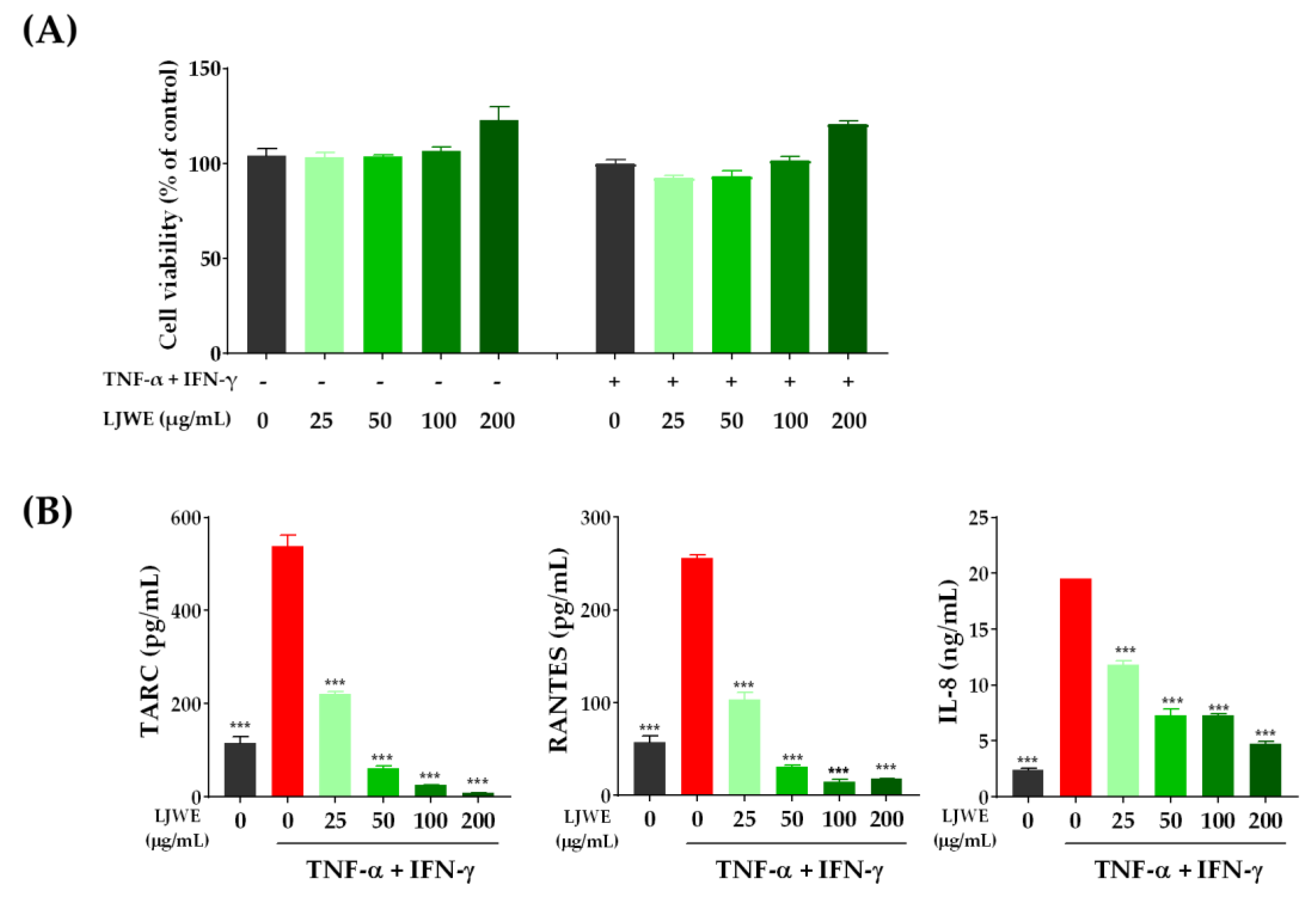
3.1. Inhibitory Effects of LJWE on the Overproduction of Inflammatory Chemokines in TNF-α/IFN-γ-Stimulated HaCaT Cells
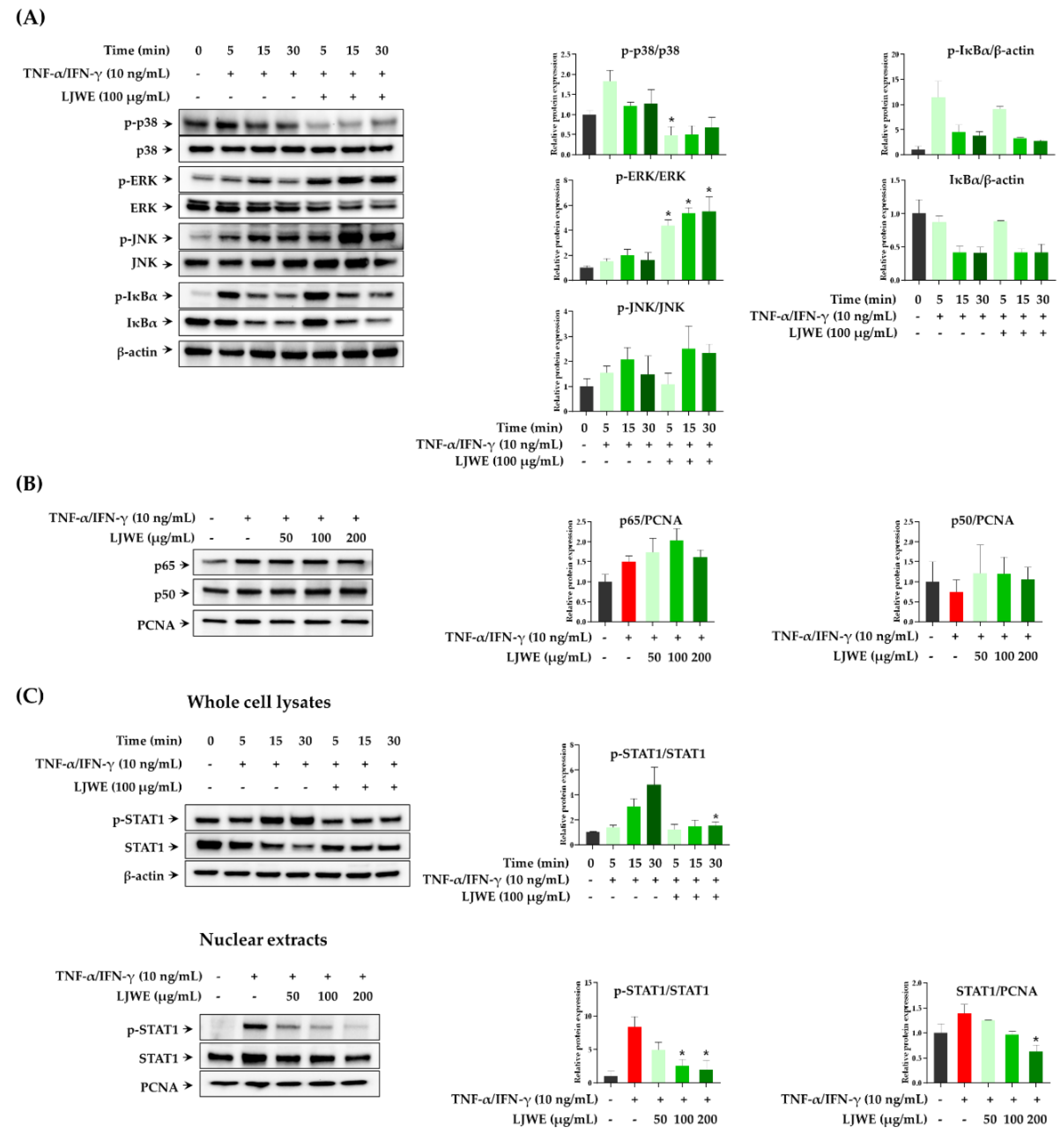
3.2. Effects of LJWE on the Expression of MAPK/NF-κB/STAT1 in TNF-α/IFN-γ-Stimulated HaCaT Cells
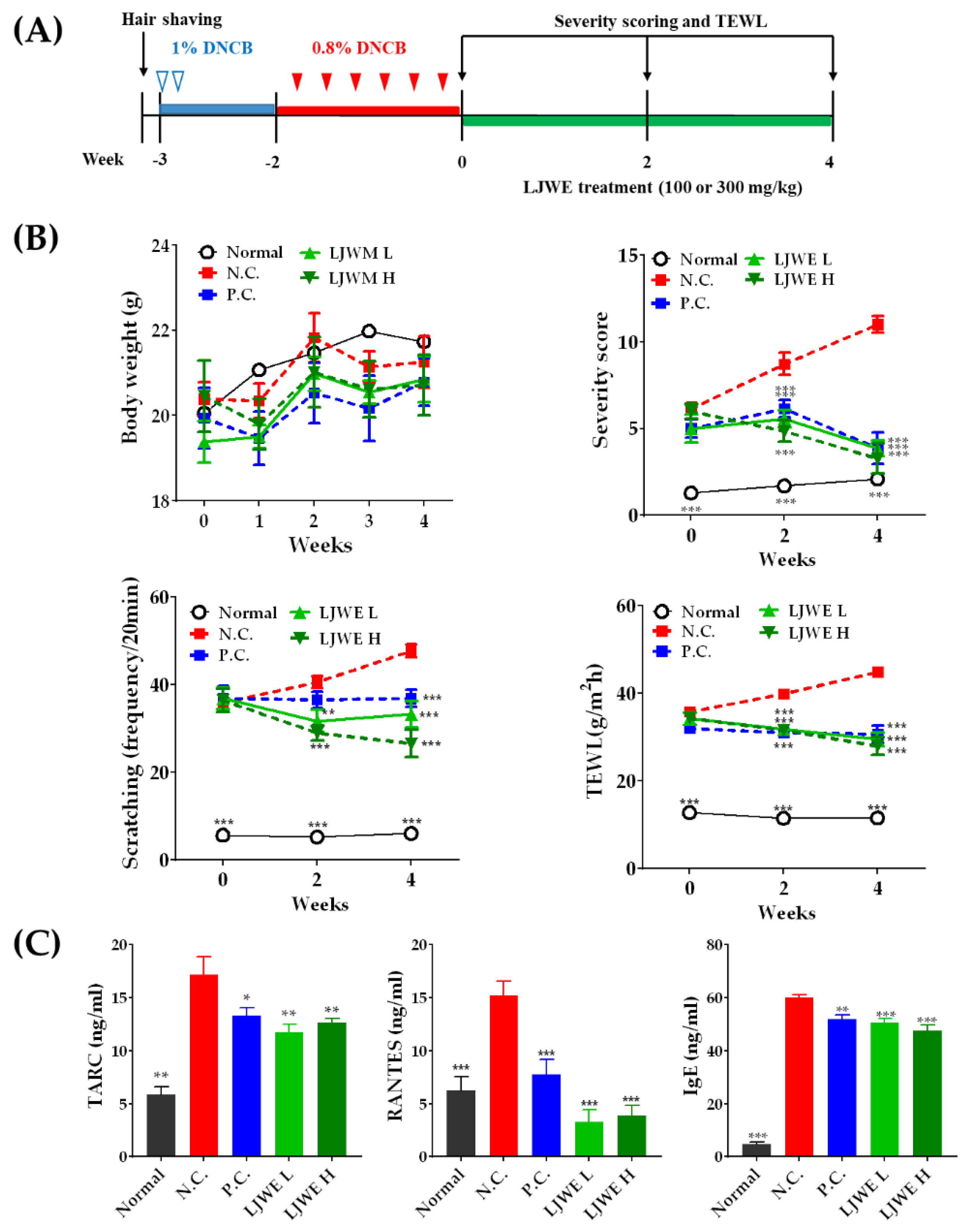
3.3. Healing of DNCB-Induced AD-Like Skin Lesions by LJWE in NC/Nga Mice
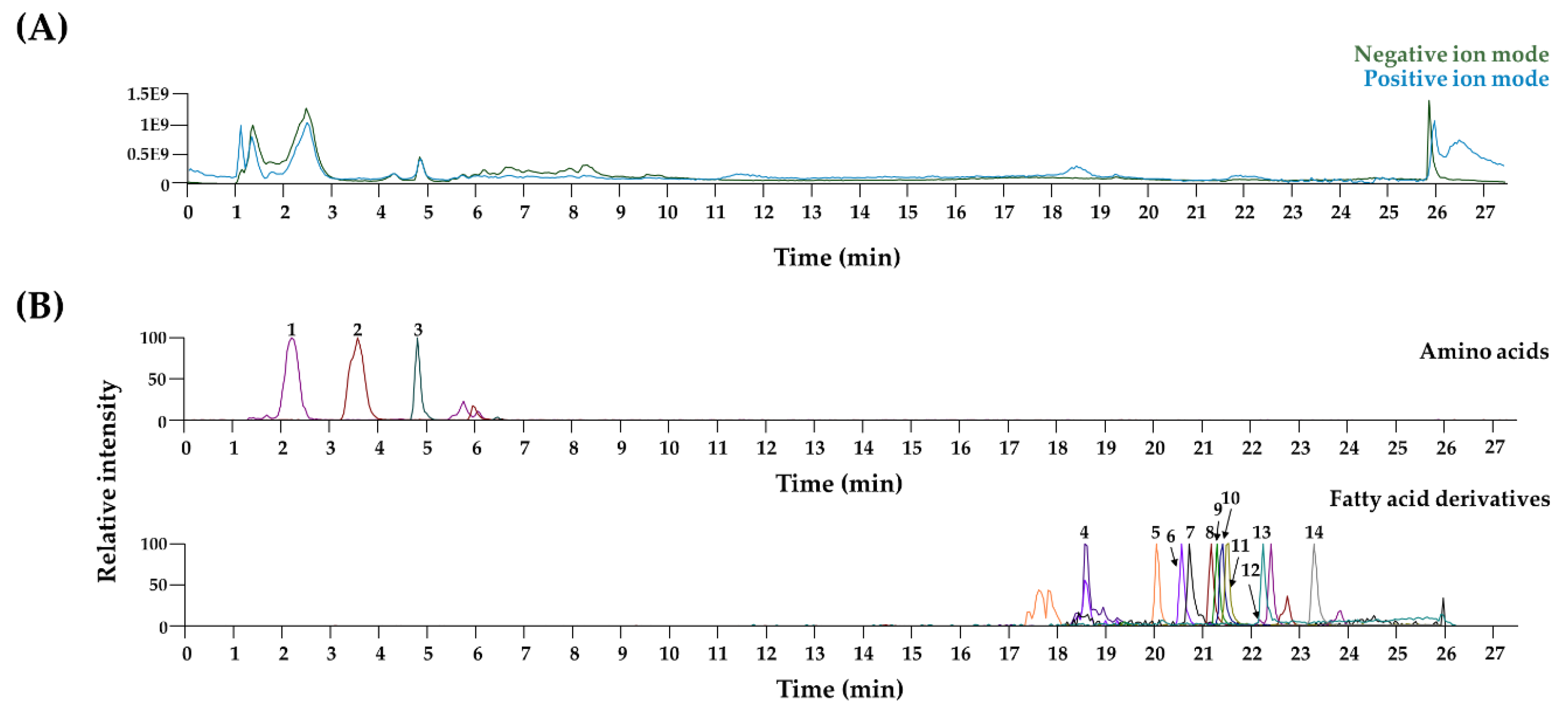
3.4. Chemical Profiling of LJWE Using UPLC–MS/MS
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Boguniewicz, M.; Leung, D.Y.M. Atopic dermatitis: A disease of altered skin barrier and immune dysregulation. Immunol. Rev. 2011, 242, 233–246. [Google Scholar] [CrossRef] [PubMed]
- Ahn, C.; Huang, W. Clinical presentation of atopic dermatitis. Adv. Exp. Med. Biol. 2017, 1027, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, R.; Woodfolk, J.A. Skin barrier defects in atopic dermatitis. Curr. Allergy Asthma Rep. 2014, 14, 433. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H.; Jin, S.W.; Park, B.H.; Kim, H.G.; Khanal, T.; Han, H.J.; Hwang, Y.P.; Choi, J.M.; Chung, Y.C.; Hwang, S.K.; et al. Cultivated ginseng inhibits 2,4-dinitrochlorobenzene-induced atopic dermatitis-like skin lesions in NC/Nga mice and TNF-α/IFN-γ-induced TARC activation in HaCaT cells. Food Chem. Toxicol. 2013, 56, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Nedoszytko, B.; Sokołowska-Wojdyło, M.; Ruckemann-Dziurdzińska, K.; Roszkiewicz, J.; Nowicki, R.J. Chemokines and cytokines network in the pathogenesis of the inflammatory skin diseases: Atopic dermatitis, psoriasis and skin mastocytosis. Postepy Dermatol. Alergol. 2014, 31, 84–91. [Google Scholar] [CrossRef]
- Chen, G.; Goeddel, D.V. TNF-R1 signaling: A beautiful pathway. Science 2002, 296, 1634–1635. [Google Scholar] [CrossRef]
- Kim, M.Y.; Lim, Y.Y.; Kim, H.M.; Park, Y.M.; Kang, H.; Kim, B.J. Synergistic inhibition of tumor necrosis factor-alpha-stimulated pro-inflammatory cytokine expression in HaCaT cells by a combination of rapamycin and mycophenolic acid. Ann. Dermatol. 2015, 27, 32–39. [Google Scholar] [CrossRef]
- Kisseleva, T.; Bhattacharya, S.; Braunstein, J.; Schindler, C.W. Signaling through the JAK/STAT pathway, recent advances and future challenges. Gene 2002, 285, 1–24. [Google Scholar] [CrossRef]
- Hajar, T.; Gontijo, J.R.V.; Hanifin, J.M. New and developing therapies for atopic dermatitis. An. Bras. Dermatol. 2018, 93, 104–107. [Google Scholar] [CrossRef]
- Man, G.; Hu, L.-Z.; Elias, P.M.; Man, M.-Q. Therapeutic benefits of natural ingredients for atopic dermatitis. Chin. J. Integr. Med. 2018, 24, 308–314. [Google Scholar] [CrossRef]
- Ming, J.X.; Wang, Z.C.; Huang, Y.; Ohishi, H.; Wu, R.J.; Shao, Y.; Wang, H.; Qin, M.Y.; Wu, Z.L.; Li, Y.Y.; et al. Fucoxanthin extracted from Laminaria Japonica inhibits metastasis and enhances the sensitivity of lung cancer to Gefitinib. J. Ethnopharmacol. 2020, 265, 113302. [Google Scholar] [CrossRef]
- Chengkui, Z.; Tseng, C.K.; Junfu, Z.; Chang, C.F. Chinese seaweeds in herbal medicine. Hydrobiologia 1984, 116, 152–154. [Google Scholar] [CrossRef]
- Lee, J.-Y.; Lee, M.-S.; Choi, H.-J.; Choi, J.-W.; Shin, T.; Woo, H.-C.; Kim, J.-I.; Kim, H.-R. Hexane fraction from Laminaria japonica exerts anti-inflammatory effects on lipopolysaccharide-stimulated RAW 264.7 macrophages via inhibiting NF-kappaB pathway. Eur. J. Nutr. 2013, 52, 409–421. [Google Scholar] [CrossRef]
- Oh, J.-H.; Kim, J.; Lee, Y. Anti-inflammatory and anti-diabetic effects of brown seaweeds in high-fat diet-induced obese mice. Nutr. Res. Pract. 2016, 10, 42–48. [Google Scholar] [CrossRef]
- Fang, Q.; Wang, J.-F.; Zha, X.-Q.; Cui, S.-H.; Cao, L.; Luo, J.-P. Immunomodulatory activity on macrophage of a purified polysaccharide extracted from Laminaria japonica. Carbohydr. Polym. 2015, 134, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-S.; Cho, E.; Weon, J.B.; Park, D.; Fréchet, M.; Chajra, H.; Jung, E. Inhibition of UVB-Induced inflammation by Laminaria japonica extract via regulation of nc886-PKR pathway. Nutrients 2020, 12, 1958. [Google Scholar] [CrossRef]
- Hwang, Y.H.; Ma, J.Y. Preventive effects of an UPLC-DAD-MS/MS fingerprinted hydroalcoholic extract of Citrus aurantium in a mouse model of ulcerative colitis. Planta Med. 2018, 84, 1101–1109. [Google Scholar] [CrossRef] [PubMed]
- Pantami, H.A.; Ahamad Bustamam, M.S.; Lee, S.Y.; Ismail, I.S.; Mohd Faudzi, S.M.; Nakakuni, M.; Shaari, K. Comprehensive GCMS and LC-MS/MS metabolite profiling of Chlorella vulgaris. Mar. Drugs 2020, 18, 367. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.C.; Cho, K.; Lee, J.H.; Subedi, L.; Yumnam, S.; Kim, S.Y. Effect of resveratrol-enriched rice on skin inflammation and pruritus in the NC/Nga mouse model of atopic dermatitis. Int. J. Mol. Sci. 2019, 20, 1428. [Google Scholar] [CrossRef]
- Leung, D.Y.M.; Boguniewicz, M.; Howell, M.D.; Nomura, I.; Hamid, Q.A. New insights into atopic dermatitis. J. Clin. Investig. 2004, 113, 651–657. [Google Scholar] [CrossRef]
- Hou, D.D.; Di, Z.H.; Qi, R.Q.; Wang, H.X.; Zheng, S.; Hong, Y.X.; Guo, H.; Chen, H.D.; Gao, X.H. Sea Buckthorn (Hippophaë rhamnoides L.) oil improves atopic dermatitis-like skin lesions via inhibition of NF-κB and STAT1 activation. Skin Pharmacol. Physiol. 2017, 30, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.-I.; Choi, B.-M.; Jang, S.I. Sulforaphane suppresses TARC/CCL17 and MDC/CCL22 expression through heme oxygenase-1 and NF-κB in human keratinocytes. Arch. Pharm. Res. 2010, 33, 1867–1876. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.-H.; Hwang, Y.-H.; Gu, M.-J.; Cho, W.-K.; Ma, J.Y. Ethanol extracts of Sanguisorba officinalis L. suppress TNF-α/IFN-γ-induced pro-inflammatory chemokine production in HaCaT cells. Phytomedicine 2015, 22, 1262–1268. [Google Scholar] [CrossRef]
- Lim, H.-S.; Seo, C.-S.; Jin, S.-E.; Yoo, S.-R.; Lee, M.-Y.; Shin, H.-K.; Jeong, S.-J. Ma Huang tang suppresses the production and expression of inflammatory chemokines via downregulating STAT1 phosphorylation in HaCaT keratinocytes. Evid. Based Complement. Alternat. Med. 2016, 2016, 7831291. [Google Scholar] [CrossRef] [PubMed]
- Barker, J.N.; Jones, M.L.; Mitra, R.S.; Crockett-Torabe, E.; Fantone, J.C.; Kunkel, S.L.; Warren, J.S.; Dixit, V.M.; Nickoloff, B.J. Modulation of keratinocyte-derived interleukin-8 which is chemotactic for neutrophils and T lymphocytes. Am. J. Pathol. 1991, 139, 869–876. [Google Scholar]
- Yang, J.-H.; Yoo, J.-M.; Lee, E.; Lee, B.; Cho, W.-K.; Park, K.-I.; Ma, J.Y. Anti-inflammatory effects of Perillae herba ethanolic extract against TNF-α/IFN-γ-stimulated human keratinocyte HaCaT cells. J. Ethnopharmacol. 2018, 211, 217–223. [Google Scholar] [CrossRef]
- Kwon, D.-J.; Bae, Y.-S.; Ju, S.M.; Goh, A.R.; Youn, G.S.; Choi, S.Y.; Park, J. Casuarinin suppresses TARC/CCL17 and MDC/CCL22 production via blockade of NF-κB and STAT1 activation in HaCaT cells. Biochem. Biophys. Res. Commun. 2012, 417, 1254–1259. [Google Scholar] [CrossRef]
- Park, J.-W.; Lee, H.-S.; Lim, Y.; Paik, J.-H.; Kwon, O.-K.; Kim, J.-H.; Paryanto, I.; Yunianto, P.; Choi, S.; Oh, S.-R.; et al. Rhododendron album blume extract inhibits TNF-α/IFN-γ-induced chemokine production via blockade of NF-κB and JAK/STAT activation in human epidermal keratinocytes. Int. J. Mol. Med. 2018, 41, 3642–3652. [Google Scholar] [CrossRef]
- Pan, Z.; Zhou, Y.; Luo, X.; Ruan, Y.; Zhou, L.; Wang, Q.; Yan, Y.J.; Liu, Q.; Chen, J. Against NF-κB/thymic stromal lymphopoietin signaling pathway, catechin alleviates the inflammation in allergic rhinitis. Int. Immunopharmacol. 2018, 61, 241–248. [Google Scholar] [CrossRef]
- Park, J.-H.; Kim, M.-S.; Jeong, G.-S.; Yoon, J. Xanthii fructus extract inhibits TNF-α/IFN-γ-induced Th2-chemokines production via blockade of NF-κB, STAT1 and p38-MAPK activation in human epidermal keratinocytes. J. Ethnopharmacol. 2015, 171, 85–93. [Google Scholar] [CrossRef]
- Goh, K.C.; Haque, S.J.; Williams, B.R. p38 MAP kinase is required for STAT1 serine phosphorylation and transcriptional activation induced by interferons. EMBO J. 1999, 18, 5601–5608. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.-F.; Kim, D.-H.; Yoon, Y.-S.; Li, J.-H.; Song, S.-B.; Jin, D.; Huang, X.-Z.; Teng, Y.-C.; Lee, K.-J. The adenylyl cyclase-cAMP system suppresses TARC/CCL17 and MDC/CCL22 production through p38 MAPK and NF-κB in HaCaT keratinocytes. Mol. Immunol. 2009, 46, 1925–1934. [Google Scholar] [CrossRef] [PubMed]
- Vestergaard, C.; Yoneyama, H.; Matsushima, K. The NC/Nga mouse: A model for atopic dermatitis. Mol. Med. Today 2000, 6, 209–210. [Google Scholar] [CrossRef]
- Jin, H.; He, R.; Oyoshi, M.; Geha, R.S. Animal models of atopic dermatitis. J. Investig. Dermatol. 2009, 129, 31–40. [Google Scholar] [CrossRef]
- Sehra, S.; Tuana, F.M.B.; Holbreich, M.; Mousdicas, N.; Kaplan, M.H.; Travers, J.B. Clinical correlations of recent developments in the pathogenesis of atopic dermatitis. An. Bras. Dermatol. 2008, 83, 57–73. [Google Scholar] [CrossRef]
- Park, J.N.; Ali-Nehari, A.; Woo, H.; Chun, B. Thermal stabilities of polyphenols and fatty acids in Laminaria japonica hydrolysates produced using subcritical water. Korean J. Chem. Eng. 2012, 29, 1604–1609. [Google Scholar] [CrossRef]
- Ziboh, V.A.; Miller, C.C.; Cho, Y. Metabolism of polyunsaturated fatty acids by skin epidermal enzymes: Generation of antiinflammatory and antiproliferative metabolites. Am. J. Clin. Nutr. 2000, 71, 361s–366s. [Google Scholar] [CrossRef]
- Kaczmarski, M.; Cudowska, B.; Sawicka-Zukowska, M.; Bobrus-Chociej, A. Supplementation with long chain polyunsaturated fatty acids in treatment of atopic dermatitis in children. Postepy Dermatol. Alergol. 2013, 30, 103–107. [Google Scholar] [CrossRef]
- Tang, L.; Li, X.; Wan, L.; Wang, H.; Mai, Q.; Deng, Z.; Ding, H. Ameliorative effect of orally administered different linoleic acid/α-linolenic acid ratios in a mouse model of DNFB-induced atopic dermatitis. J. Funct. Foods 2020, 65, 103754. [Google Scholar] [CrossRef]
- Yoshida, S.; Yasutomo, K.; Watanabe, T. Treatment with DHA/EPA ameliorates atopic dermatitis-like skin disease by blocking LTB4 production. J. Med. Investig. 2016, 63, 187–191. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Rt (min) | Calculated (m/z) | Estimated (m/z) | Adduct | Error (ppm) | Formula | MS/MS Fragments (m/z) | Identifications |
|---|---|---|---|---|---|---|---|---|
| Amino acids | ||||||||
| 1 | 2.2 | 132.102 | 132.102 | [M + H]+ | 2.683 | C6H13NO2 | 86 | L-Leucine/Isoleucine * |
| 2 | 3.6 | 166.086 | 166.087 | [M + H]+ | 2.284 | C9H11NO2 | 166, 120 | Phenylalanine * |
| 3 | 4.8 | 205.097 | 205.098 | [M + H]+ | 1.948 | C11H12N2O2 | 188, 159, 146 | L-Tryptophan * |
| Fatty acid derivatives | ||||||||
| 4 | 18.6 | 319.228 | 319.228 | [M − H]− | 1.736 | C20H32O3 | 319, 301, 257, 219, 175, 113 | 15-Hydroxy-5,8,11,13 -eicosatetraenoic acid * |
| 5 | 20.0 | 275.202 | 275.202 | [M − H]− | 2.033 | C18H28O2 | 275, 231 | Stearidonic acid * |
| 6 | 20.6 | 301.217 | 301.218 | [M − H]− | 1.673 | C20H30O2 | 301, 257, 203 | Eicosapentaenoic acid * |
| 7 | 20.7 | 277.217 | 277.218 | [M − H]− | 1.928 | C18H30O2 | 277 | Linolenic acid * |
| 8 | 21.2 | 256.263 | 256.264 | [M + H]+ | 1.305 | C16H33NO | 256, 116, 102, 88 | Hexadecanamide |
| 9 | 21.3 | 303.233 | 303.234 | [M − H]− | 2.083 | C20H32O2 | 303, 259 | Arachidonic acid * |
| 10 | 21.4 | 282.279 | 282.280 | [M + H]+ | 2.069 | C18H35NO | 282, 265, 247, 177, 149, 135 | Oleamide |
| 11 | 21.5 | 279.233 | 279.234 | [M − H]− | 1.934 | C18H32O2 | 279 | Linoleic acid * |
| 12 | 21.9 | 284.294 | 284.295 | [M + H]+ | 1.430 | C18H37NO | 284, 88 | Stearamide |
| 13 | 22.2 | 255.233 | 255.233 | [M − H]− | 1.458 | C16H32O2 | 255 | Palmitic acid * |
| 14 | 23.3 | 283.264 | 283.265 | [M + H]+ | 1.838 | C18H36O2 | 283 | Stearic acid * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hwang, Y.-H.; Song, H.-K.; Lee, A.; Ha, H.; Kim, T. Laminaria japonica Suppresses the Atopic Dermatitis-Like Responses in NC/Nga Mice and Inflamed HaCaT Keratinocytes via the Downregulation of STAT1. Nutrients 2020, 12, 3238. https://doi.org/10.3390/nu12113238
Hwang Y-H, Song H-K, Lee A, Ha H, Kim T. Laminaria japonica Suppresses the Atopic Dermatitis-Like Responses in NC/Nga Mice and Inflamed HaCaT Keratinocytes via the Downregulation of STAT1. Nutrients. 2020; 12(11):3238. https://doi.org/10.3390/nu12113238
Chicago/Turabian StyleHwang, Youn-Hwan, Hyun-Kyung Song, Ami Lee, Hyunil Ha, and Taesoo Kim. 2020. "Laminaria japonica Suppresses the Atopic Dermatitis-Like Responses in NC/Nga Mice and Inflamed HaCaT Keratinocytes via the Downregulation of STAT1" Nutrients 12, no. 11: 3238. https://doi.org/10.3390/nu12113238
APA StyleHwang, Y.-H., Song, H.-K., Lee, A., Ha, H., & Kim, T. (2020). Laminaria japonica Suppresses the Atopic Dermatitis-Like Responses in NC/Nga Mice and Inflamed HaCaT Keratinocytes via the Downregulation of STAT1. Nutrients, 12(11), 3238. https://doi.org/10.3390/nu12113238