
Spatiotemporal Characteristics and Heterogeneity of Vegetation Phenology in the Yangtze River Delta

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
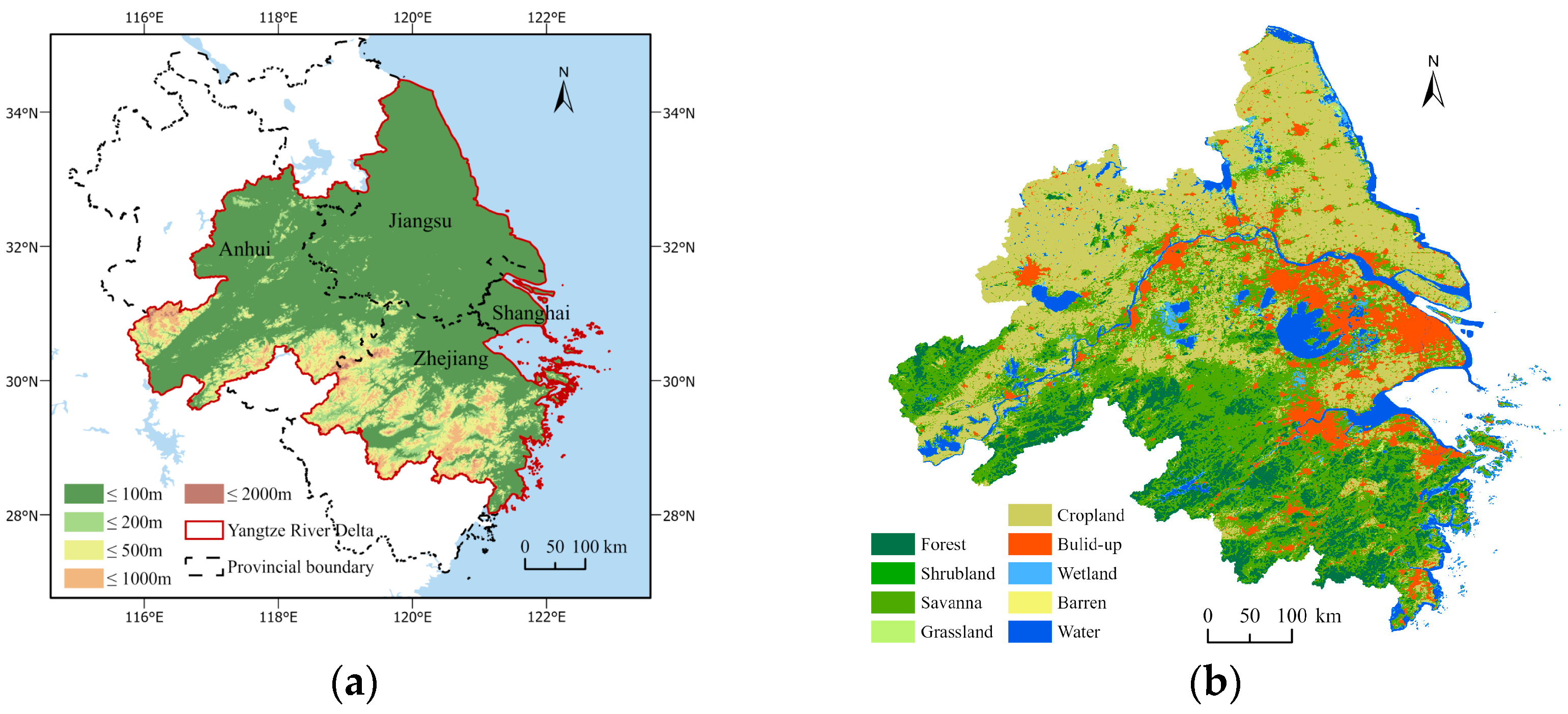
2.1. Experimental Area
2.2. Data and Preprocessing
2.3. Extraction of Vegetation Phenology Information
2.4. Analysis of Phenological Changes
2.5. Spatiotemporal Heterogeneity Analysis
3. Results
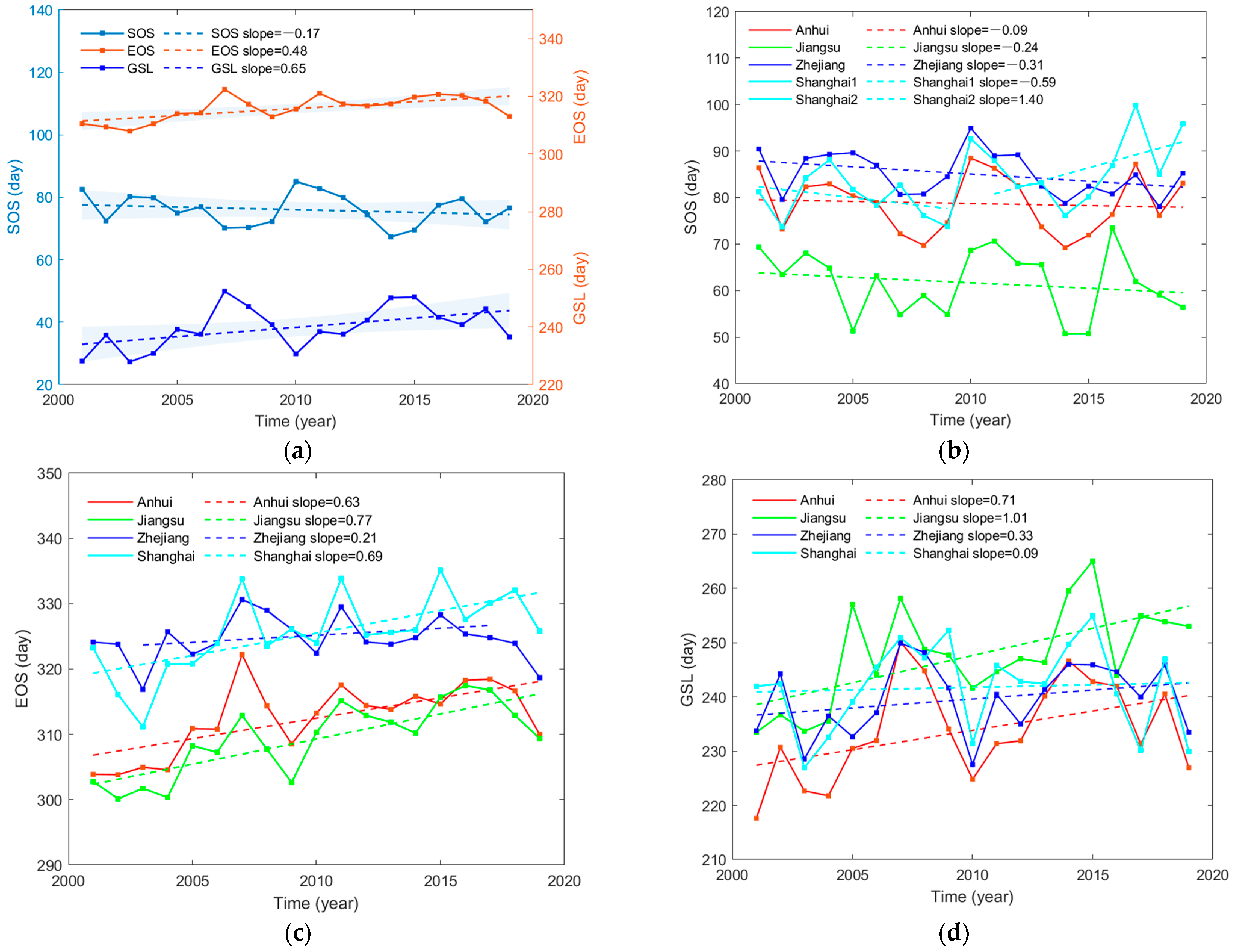
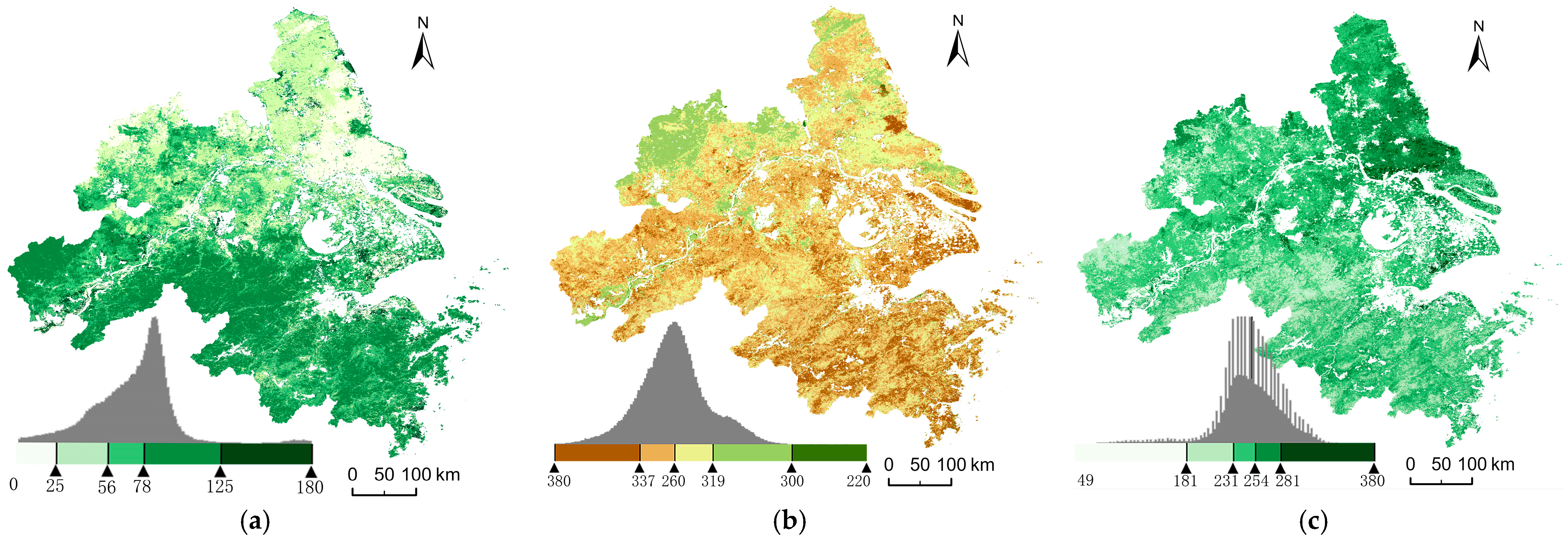
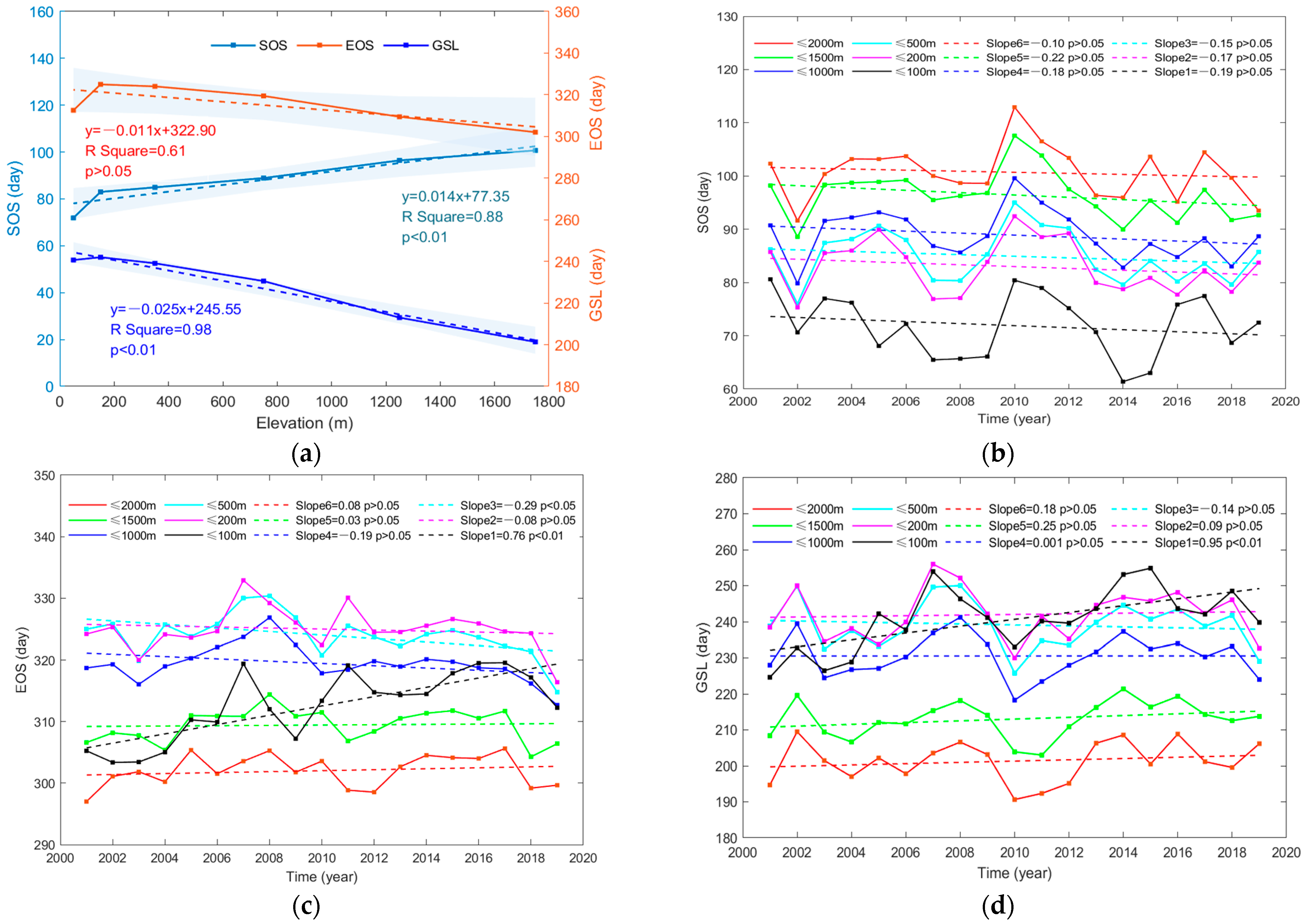
3.1. Spatiotemporal Variation Characteristics of Phenology
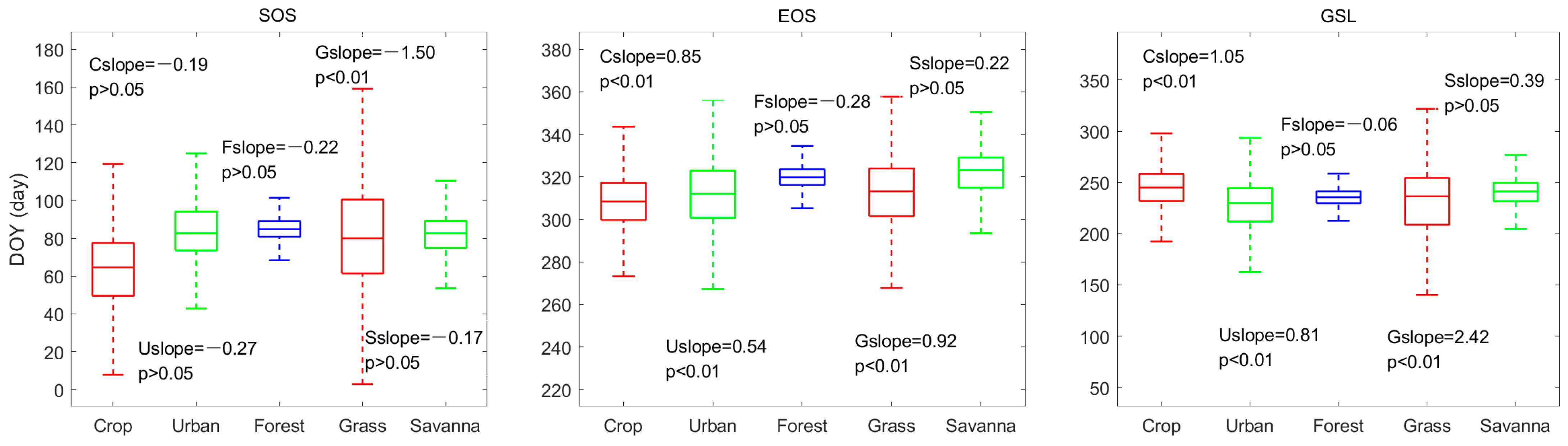
3.2. Phenological Characteristics of Different Land Cover Types
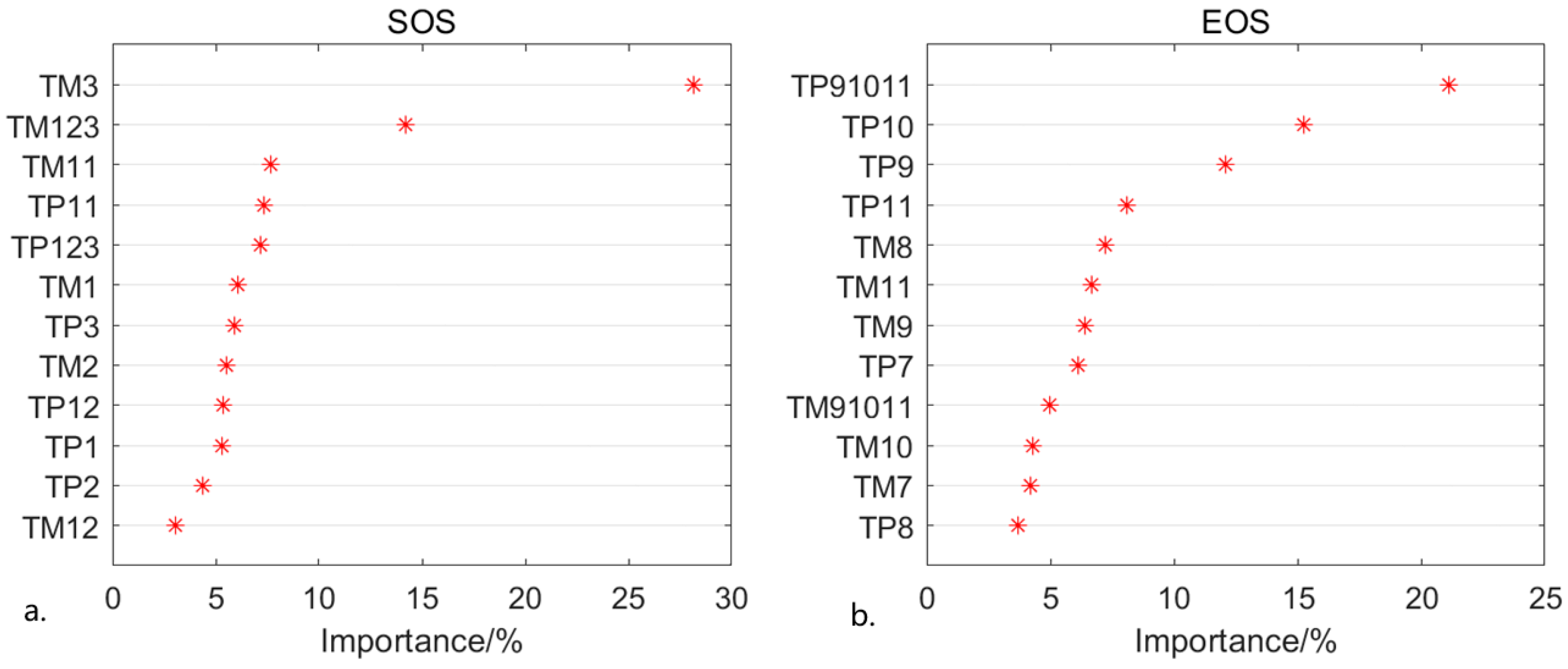
3.3. Relationship between Phenology and Climate
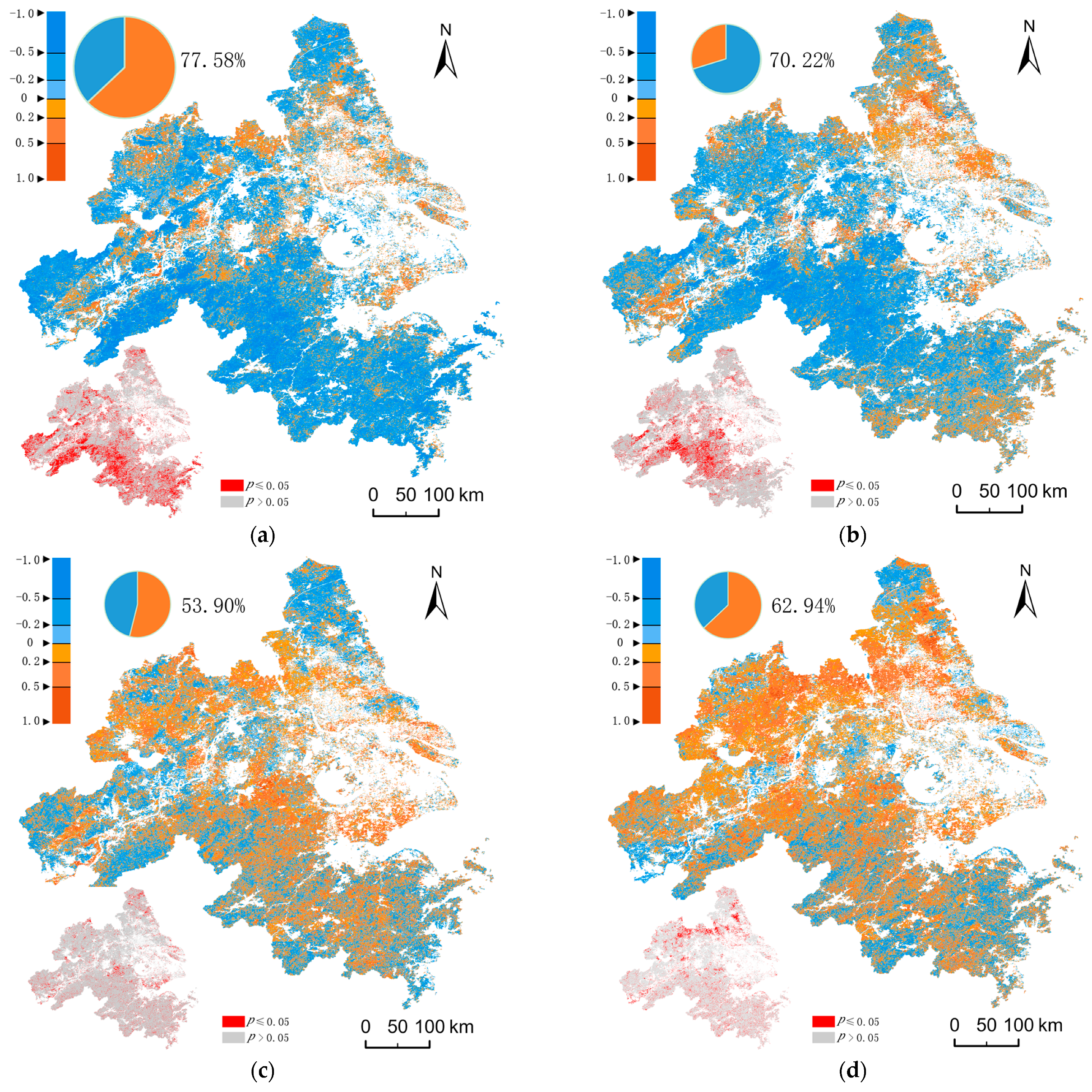
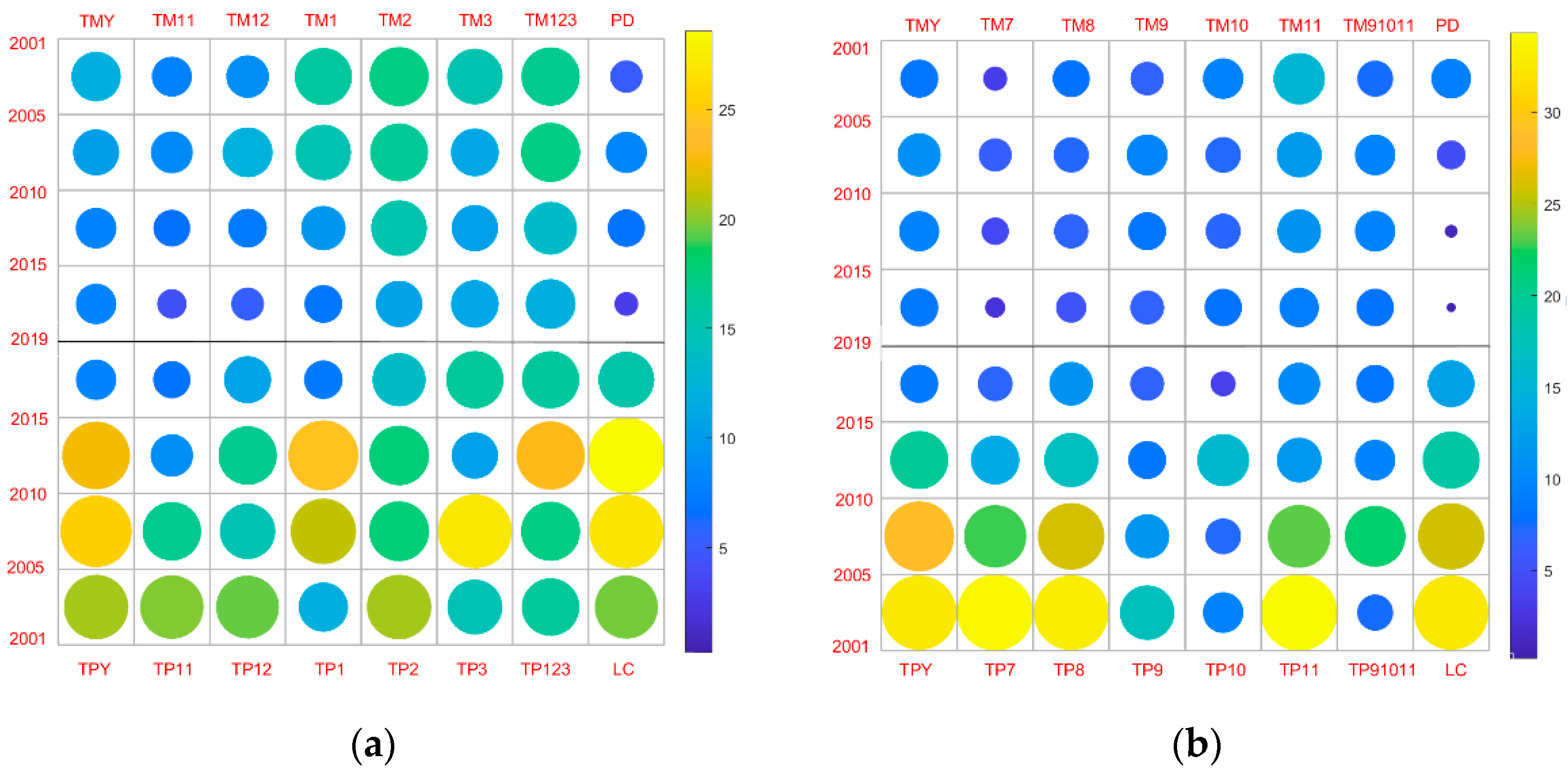
3.4. Spatial Heterogeneity and Its Driving Factors
4. Discussion
4.1. Phenological Change Characteristics
4.2. Impacts of Land Cover Type on Phenology
4.3. Effects of Population Density and Land Surface Temperature on Phenology
5. Conclusions
- (1)
- SOS in the Yangtze River Delta shows an insignificant advance (p > 0.05) of 0.17 days per year, EOS is significantly delayed (p < 0.01) by 0.48 days per year and GSL is prolonged by 0.65 days per year (p < 0.05). SOS has an obvious negative correlation with temperature and precipitation, and EOS has a positive correlation with them. Preseason temperature has a greater contribution to SOS, while preseason precipitation is a major factor of EOS. Two months before the growing season, it is jointly affected by temperature and precipitation.
- (2)
- Large divergent responses of vegetation phenology to spatial distribution and elevation grades are found. The variation characteristics of SOS and GSL in the Yangtze River Delta and its provinces do not obey the rules of latitude variation. EOS values above 200 m follow the trend of decreasing as elevation increases, while the EOS values below 200 m have the opposite characteristic. In addition, the changes in phenological indexes of vegetation below 100 m are the most obvious.
- (3)
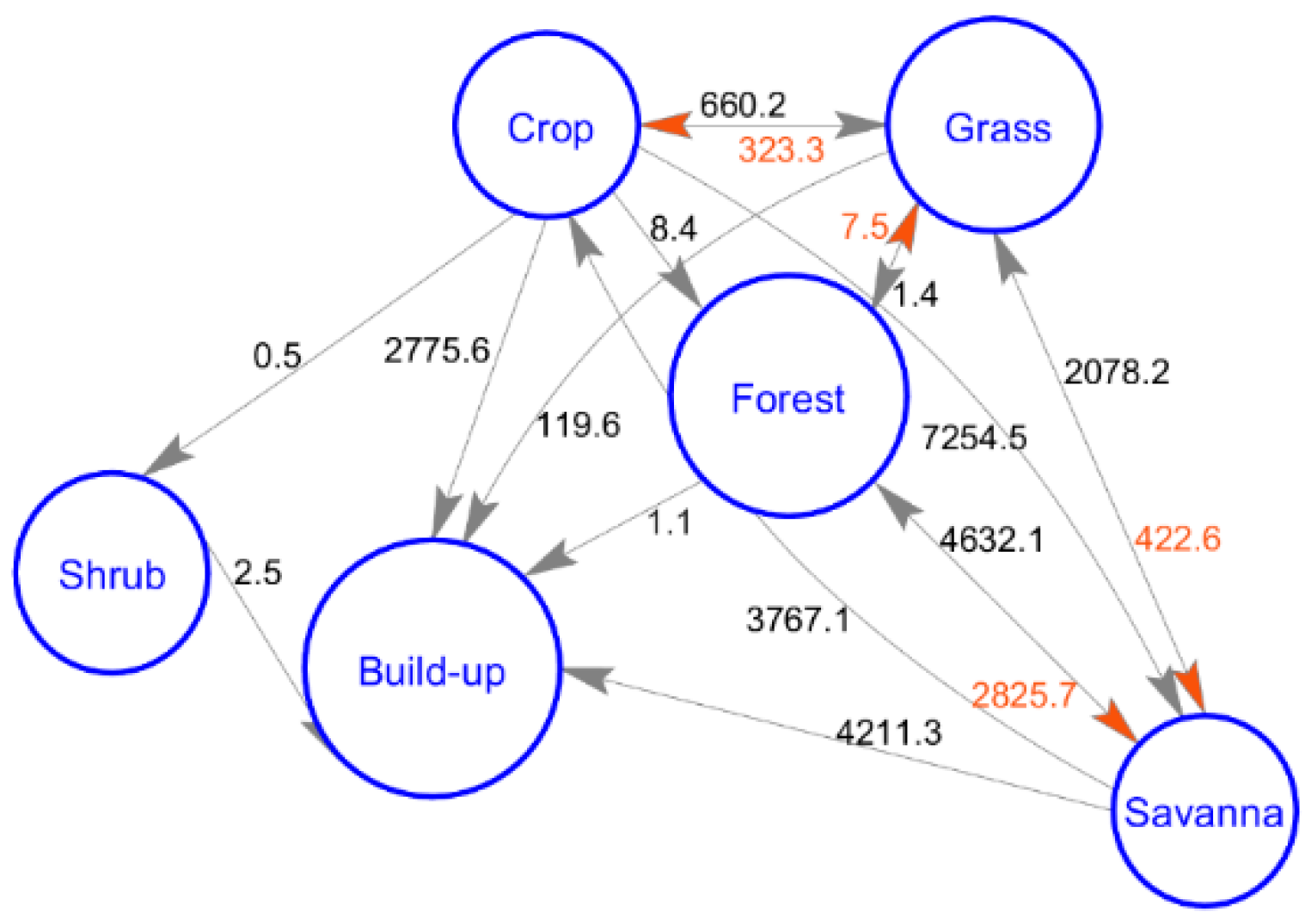
- The phenology of vegetation is different in the five LC types. In addition, the contribution of LC types to the spatial change of phenology is greater than that of temperature and precipitation, and the transfer of LC may also be one factor driving the interannual change in phenology.
- (4)
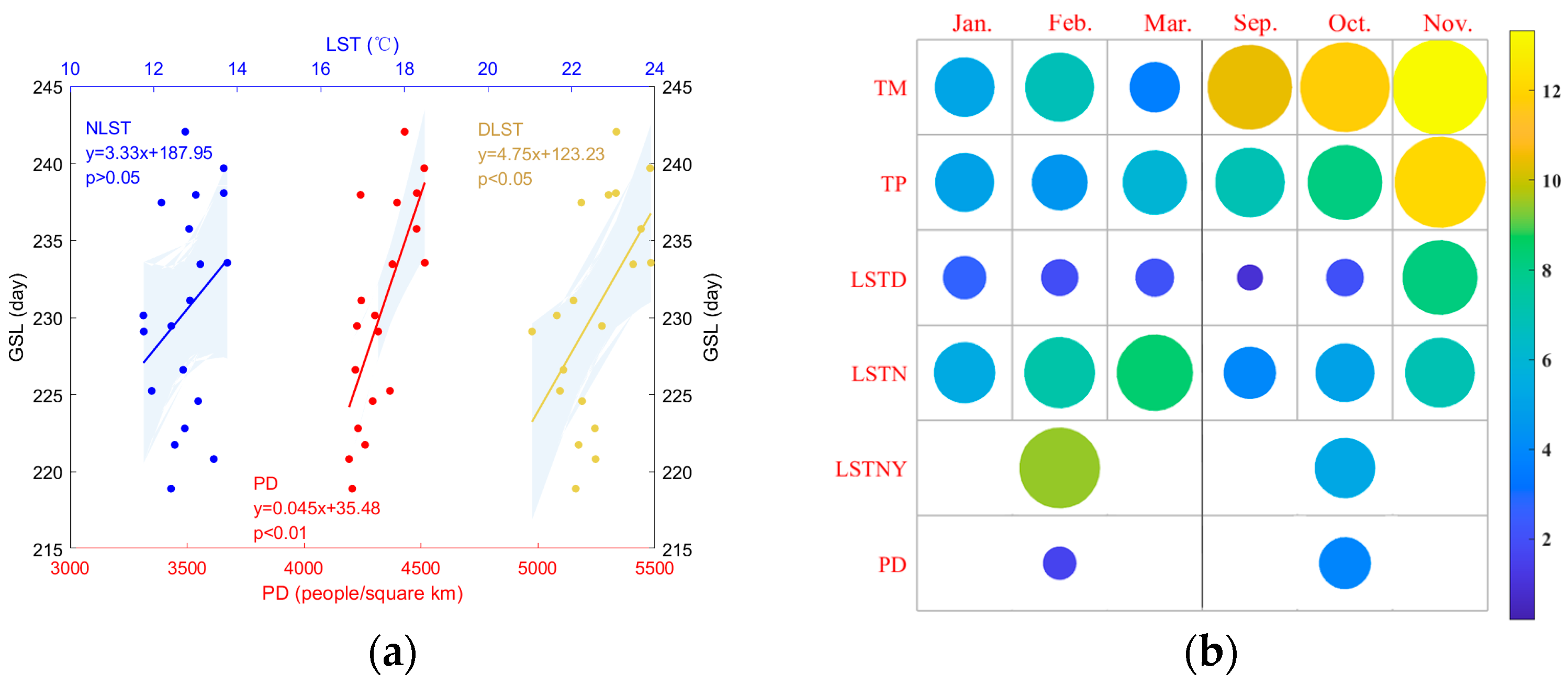
- In non-core urban areas, NLST has a relatively large contribution to the spatial heterogeneity of phenology. Although PD has a strong correlation with GSL, it contributes little to spatial heterogeneity.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- White, M.A.; Nemani, R.R.; Thornton, P.E.; Running, S.W. Satellite evidence of phenological differences between urbanized and rural areas of the Eastern United States deciduous broadleaf forest. Ecosystems 2002, 5, 260–273. [Google Scholar] [CrossRef]
- White, M.A.; Beurs, K.M.D.; Didan, K.; Inouye, D.W.; Richardson, A.D.; Jensen, O.P.; O’keefe, J.; Zhang, G.; Nemani, R.R.; van Leeuwen, W.J.D.; et al. Intercomparison, interpretation, and assessment of spring phenology in North America estimated from remote sensing for 1982–2006. Glob. Change Biol. 2009, 15, 2335–2359. [Google Scholar] [CrossRef]
- Richardson, A.D.; Keenan, T.F.; Migliavacca, M.; Ryu, Y.; Sonnentag, O.; Toomey, M. Climate change, phenology, and phenological control of vegetation feedbacks to the climate system. Agric. For. Meteorol. 2013, 169, 156–173. [Google Scholar] [CrossRef]
- Li, X.; Zhou, Y.; Asrar, G.R.; Mao, J.; Li, X.; Li, W. Response of vegetation phenology to urbanization in the conterminous United States. Glob. Change Biol. 2017, 23, 2818–2830. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, W.; Yuan, W.; Chen, X.; Zhang, B.; Fan, L.; He, B.; Hu, Z.; Liu, S.; Liu, W. Higher plant photosynthetic capability in autumn responding to low atmospheric vapor pressure deficit. Innovation 2021, 2, 100163. [Google Scholar] [CrossRef]
- Chang, Q.; Xiao, X.; Jiao, W.; Ling, D.; Zeng, Z.; Qin, Y. Assessing consistency of spring phenology of snow-covered forests as estimated by vegetation indices, gross primary production, and solar induced chlorophyll fluorescence. Agric. For. Meteorol. 2019, 275, 305–316. [Google Scholar] [CrossRef]
- Qiu, T.; Song, C.; Li, J. Deriving annual double-season cropland phenology using Landsat imagery. Remote Sens. 2020, 12, 3275. [Google Scholar] [CrossRef]
- Cui, L.; Shi, J.; Du, H. Advances in remote sensing extraction of vegetation phenology and its driving factors. Adv. Earth Sci. 2021, 36, 9–16. [Google Scholar] [CrossRef]
- Piao, S.; Fang, J.; Zhou, L.; Ciais, P.; Zhu, B. Variations in satellite-derived phenology in China’s temperate vegetation. Glob. Change Biol. 2006, 12, 672–685. [Google Scholar] [CrossRef]
- Ge, Q.; Wang, H.; Rutishauser, T.; Dai, J. Phenological response to climate change in China: A meta-analysis. Glob. Change Biol. 2015, 21, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chen, Y.; Li, Z.; Li, Y.; Zhang, Q.; Zan, M. Driving forces of the changes in vegetation phenology in the Qinghai-Tibet Plateau. Remote Sens. 2021, 13, 4952. [Google Scholar] [CrossRef]
- Jiao, F.; Liu, H.; Xu, X.; Gong, H.; Lin, Z. Trend evolution of vegetation phenology in china during the period of 1981–2016. Remote Sens. 2020, 12, 572. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Du, P.; Chen, D.; Lin, C.; Zheng, H.; Guo, S. Characterizing urbanization induced land surface phenology change from time-series remotely sensed images at fine spatio-temporal scale: A case study in Nanjing, China (2001–2018). J. Clean. Prod. 2020, 274, 122487. [Google Scholar] [CrossRef]
- Shen, M.; Zhang, G.; Cong, N.; Wang, S.; Kong, W.; Piao, S. Increasing altitudinal gradient of spring vegetation phenology during the last decade on the Qinghai-Tibetan Plateau. Agric. For. Meteorol. 2014, 189, 71–80. [Google Scholar] [CrossRef]
- Zhang, Y.; Yin, P.; Li, X.; Niu, Q.; Wang, Y.; Cao, W.; Huang, J.; Chen, H.; Yao, X.; Yu, L.; et al. The divergent response of vegetation phenology to urbanization: A case study of Beijing city, China. Sci. Total Environ. 2022, 803, 150079. [Google Scholar] [CrossRef]
- Ding, H.; Xu, L.; Elmore, A.J.; Shi, Y. Vegetation Phenology Influenced by Rapid Urbanization of The Yangtze Delta Region. Remote Sens. 2020, 12, 1783. [Google Scholar] [CrossRef]
- Guo, J.; Liu, X.; Ge, W.; Ni, X.; Ma, W.; Lu, Q.; Xing, X. Specific Drivers and Responses to Land Surface Phenology of Different Vegetation Types in the Qinling Mountains, Central China. Remote Sens. 2021, 13, 4538. [Google Scholar] [CrossRef]
- Kelsey, K.C.; Pedersen, S.H.; Leffler, A.J.; Sexton, J.O.; Feng, M.; Welker, J.M. Winter snow and spring temperature have differential effects on vegetation phenology and productivity across plant communities. Glob. Change Biol. 2021, 27, 1572–1586. [Google Scholar] [CrossRef]
- Jia, W.; Zhao, S.; Zhang, X.; Liu, S.; Henebry, G.M.; Liu, L.L. Urbanization imprint on land surface phenology: The urban-rural gradient analysis for Chinese cities. Glob. Change Biol. 2021, 27, 2895–2904. [Google Scholar] [CrossRef]
- Tang, H.; Li, Z.; Zhu, Z.; Chen, B.; Zhang, B.; Xin, X. Variability and climate change trend in vegetation phenology of recent decades in the Greater Khingan Mountain area, Northeastern China. Remote Sens. 2015, 7, 11914–11932. [Google Scholar] [CrossRef] [Green Version]
- Qiu, T.; Song, C.; Li, J. Impacts of urbanization on vegetation phenology over the past three decades in Shanghai, China. Remote Sens. 2017, 9, 970. [Google Scholar] [CrossRef] [Green Version]
- Yang, K.; Sun, W.; Luo, Y.; Zhao, L. Impact of urban expansion on vegetation: The case of China (2000–2018). J. Environ. Manag. 2021, 291, 112598. [Google Scholar] [CrossRef] [PubMed]
- Ren, S.; Yi, S.; Peich, M.; Wang, X. Diverse Responses of Vegetation Phenology to Climate Change in Different Grasslands in Inner Mongolia during 2000–2016. Remote Sens. 2017, 10, 17. [Google Scholar] [CrossRef] [Green Version]
- Ren, S.; Qin, Q.; Ren, H.; Sui, J.; Zhang, Y. New model for simulating autumn phenology of herbaceous plants in the Inner Mongolian Grassland. Agric. For. Meteorol. 2019, 275, 136–145. [Google Scholar] [CrossRef]
- Cong, N.; Wang, T.; Nan, H.; Ma, Y.; Wang, X.; Myneni, R.B.; Piao, S. Changes in satellite-derived spring vegetation green-up date and its linkage to climate in China from 1982 to 2010: A multimethod analysis. Glob. Change Biol. 2013, 19, 881–891. [Google Scholar] [CrossRef]
- Li, P.; Zhu, Q.; Peng, C.; Zhang, J.; Wang, M.; Zhang, J.; Ding, J.; Zhou, X. Change in Autumn Vegetation Phenology and the Climate Controls From 1982 to 2012 on the Qinghai-Tibet Plateau. Front. Plant. Sci. 2020, 10, 1677. [Google Scholar] [CrossRef] [Green Version]
- Shen, M.; Piao, S.; Cong, N.; Zhang, G.; Jassens, I.A. Precipitation impacts on vegetation spring phenology on the Tibetan Plateau. Glob. Change Biol. 2015, 21, 3647–3656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, F.; Yokozawa, M.; Zhang, Z.; Hayashi, Y.; Ishigooka, Y. Land surface phenology dynamics and climate variations in the North East China Transect(NECT), 1982–2000. Int. J. Remote Sens. 2008, 29, 5461–5478. [Google Scholar] [CrossRef]
- Peng, S.; Piao, S.; Ciais, P.; Myneni, R.B.; Chen, A.; Chevallier, F.; Dolman, A.J.; Janssens, I.A.; Peñuelas, J.; Zhang, G.; et al. Asymmetric effects of daytime and night-time warming on Northern Hemisphere vegetation. Nature 2013, 501, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Wan, S.; Xia, J.; Liu, W.; Niu, S. Photosynthetic overcompensation under nocturnal warming enhances grassland carbon sequestration. Ecology 2009, 90, 2700–2710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, F.; Zhang, L.; Zhang, Z.; Jiang, L.; Wang, Y.; Duan, J.; Wang, Q.; Li, B.; Liu, P.; Hong, H. Opposite effects of winter day and night temperature changes on early phenophases. Ecology 2019, 100, e02775. [Google Scholar] [CrossRef] [PubMed]
- Piao, S.; Tan, J.; Chen, A.; Fu, Y.; Ciais, P.; Qiang, L.; Janssens, I.A.; Vicca, S.; Zeng, Z.; Jeong, S.J. Leaf onset in the northern hemisphere triggered by daytime temperature. Nat. Commun. 2015, 6, 6911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.H.; Piao, S.; Zhou, X.; Geng, X.; Hao, F.; Vitasse, Y.; Janssens, I.A. Short photoperiod reduces the temperature sensitivity of leaf-out in saplings of Fagus sylvatica but not in Horse chestnut. Glob. Change Biol. 2019, 25, 1696–1703. [Google Scholar] [CrossRef]
- Fu, Y.H.; Zhao, H.; Piao, S.; Peaucelle, M.; Peng, S.; Zhou, G.; Ciais, P.; Huang, M.; Menzel, A.; Peñuelas, J.; et al. Declining global warming effects on the phenology of spring leaf unfolding. Nature 2015, 526, 104–107. [Google Scholar] [CrossRef] [Green Version]
- Korner, C.; Basler, D. Phenology Under Global Warming. Science 2010, 327, 1461–1462. [Google Scholar] [CrossRef]
- David, B.; Christian, K. Photoperiod and temperature responses of bud swelling and bud burst in four temperate forest tree species. Tree Physiol. 2014, 34, 377–388. [Google Scholar] [CrossRef] [Green Version]
- Caffarra, A.; Donnelly, A.; Chuine, I. Modelling the timing of Betula pubescens budburst. II. Integrating complex effects of photoperiod into process-based models. Clim. Res. 2011, 46, 159–170. [Google Scholar] [CrossRef]
- Moon, M.; Richardson, A.D.; O’Keefe, J.; Friedl, M.A. Senescence in temperate broadleaf trees exhibits species-specific dependence on photoperiod versus thermal forcing. Agric. For. Meteorol. 2022, 322, 109026. [Google Scholar] [CrossRef]
- Meng, L.; Zhou, Y.; Gu, L.; Richardson, A.D.; Peñuelas, J.; Fu, Y.; Wang, Y.; Asrar, G.R.; de Boeck, H.J.; Mao, J.; et al. Photoperiod decelerates the advance of spring phenology of six deciduous tree species under climate warming. Glob. Change Biol. 2021, 27, 2914–2927. [Google Scholar] [CrossRef]
- Zohner, C.M.; Benito, B.M.; Svenning, J.C.; Renner, S.S. Day length unlikely to constrain climate-driven shifts in leaf-out times of northern woody plants. Nat. Clim. Change 2016, 6, 1120–1123. [Google Scholar] [CrossRef]
- Lv, A.L.; Huo, Z.G.; Yang, J.Y. Phenological Characteristics of Representative Woody Plants at Different Altitude Sites in Jinnan Region and Their Response to Climate Change. Chin. J. Agrometeorol. 2020, 41, 65–75. [Google Scholar] [CrossRef]
- Yuan, Q.L.; Yang, J. Phenological Changes of Grassland Vegetation and Its Response to Climate Change in Qinhai-Tibet Plateau. Chin. J. Grassl. 2021, 43, 32–43. [Google Scholar] [CrossRef]
- Thompson, J.A.; Paul, D.J. Assessing spatial and temporal patterns in land surface phenology for the Australian Alps (2000–2014). Remote Sens. Environ. 2017, 199, 1–13. [Google Scholar] [CrossRef]
- Jochner, S.; Menzel, A. Urban phenological studies- Past, present, future. Environ. Pollut. 2015, 203, 250–261. [Google Scholar] [CrossRef]
- Meng, L.; Mao, J.; Zhou, Y.; Richardson, A.D.; Jia, G. Urban warming advances spring phenology but reduces the response of phenology to temperature in the conterminous United States. Proc. Natl. Acad. Sci. USA 2020, 117, 201911117. [Google Scholar] [CrossRef]
- Zipper, S.C.; Schatz, J.; Singh, A.; Kucharik, C.J.; Townsend, P.A.; Loheide, S.P. Urban heat island impacts on plant phenology: Intra-urban variability and response to land cover. Environ. Res. Lett. 2016, 11, 054023. [Google Scholar] [CrossRef]
- Zheng, J.; Ge, Q.; Hao, Z.; Zhong, S.; Ma, X.; Zhang, X. Changes of spring phenodate in Yangtze River Delta region in the past 150 years. Acta Geogr. Sin. 2012, 67, 45–52. [Google Scholar] [CrossRef]
- Han, G.; Xu, J. Land Surface phenology and land surface temperature changes along an urban-rural gradient in Yangtze River Delta, China. Environ. Manag. 2013, 52, 234–249. [Google Scholar] [CrossRef]
- Wang, Y.; Xue, Z.; Chen, J.; Chen, G. Spatio-temporal analysis of phenology in Yangtze River Delta based on MODIS NDVI time series from 2001 to 2015. Front. Earth Sci. 2018, 13, 92–110. [Google Scholar] [CrossRef]
- Xiao, R.; Cao, W.; Liu, Y.; Lu, B. The impacts of landscape patterns spatio-temporal changes on land surface temperature from a multi-scale perspective: A case study of the Yangtze River Delta. Sci. Total Environ. 2022, 821, 153381. [Google Scholar] [CrossRef]
- Farr, T.G.; Rosen, P.A.; Caro, E.; Crippen, R.; Duren, R.; Hensley, S.; Kobrick, M.; Paller, M.; Rodriguez, E.; Roth, L.; et al. The shuttle radar topography mission. Rev. Geophys. 2007, 45, 361. [Google Scholar] [CrossRef] [Green Version]
- Sulla-Menashe, D.; Friedl, M.A. User Guide to Collection 6 MODIS Land Cover (MCD12Q1 and MCD12C1) Product; USGS: Reston, VA, USA, 2018; 18p. Available online: http://girps.net/wp-content/uploads/2019/03/MCD12_User_Guide_V6.pdf (accessed on 22 February 2022).
- Ji, Y.; Jin, J.; Zhan, W.; Guo, F.; Yan, T. Quantification of urban heat island-induced contribution to advance in spring phenology: A case study in Hangzhou, China. Remote Sens. 2021, 13, 3684. [Google Scholar] [CrossRef]
- Li, B.Y.; Pan, B.T.; Han, J.F. Basic terrestrial geomorphological types in China and their circum scriptions. Quat. Sci. 2008, 28, 535–543. [Google Scholar] [CrossRef]
- Liang, L.; Schwartz, M.D.; Fei, S. Validating satellite phenology through intensive ground observation and landscape scaling in a mixed seasonal forest. Remote Sens. Environ. 2011, 115, 143–157. [Google Scholar] [CrossRef]
- Zhang, X.; Friedl, M.A.; Schaaf, C.B. Global vegetation phenology from Moderate Resolution Imaging Spectroradiometer (MODIS): Evaluation of global patterns and comparison with in situ measurements. J. Geophys. Res. Biogeosci. 2015, 111, 367–375. [Google Scholar] [CrossRef]
- Wang, L.; De Boeck, H.J.; Chen, L.; Song, C.; Chen, Z.; McNulty, S.; Zhang, Z. Urban warming increases the temperature sensitivity of spring vegetation phenology at 292 cities across China. Sci. Total Environ. 2022, 834, 155154. [Google Scholar] [CrossRef]
- Mann, H.B. Nonparametric Tests Against Trend. Econometrica 1945, 13, 245–259. [Google Scholar] [CrossRef]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Xu, C. Geodetector: Principle and prospective. Acta Geogr. Sin. 2017, 72, 116–134. [Google Scholar] [CrossRef]
- He, B.; Cai, Y.; Ran, W.; Jiang, H. Spatial and seasonal variations of soil salinity following vegetation restoration in coastal saline land in eastern China. CATENA 2014, 118, 147–153. [Google Scholar] [CrossRef]
- Tian, J.; Cai, Y.; Zhao, X.; Xie, C. Phenological analysis and comparison of six greening trees between coastal island and inland, choosing Chongming island and Minghang area of Shanghai for an example. J. Shanghai Jiaotong Univ. 2015, 33, 71–79. [Google Scholar] [CrossRef]
- Zhao, X.; Shi, C.; He, B.; Ran, W.; Cai, Y. Spring phenological characteristics and phenophase classification of landscape greening tree species in Chongming Island of Shanghai. Chin. J. Ecol. 2013, 32, 2275–2280. [Google Scholar] [CrossRef]
- He, Z.; Du, J.; Chen, L.; Xi, Z.; Lin, P.; Zhao, M.; Fang, S. Impacts of recent climate extremes on spring phenology in arid-mountain ecosystems in China. Agric. For. Meteorol. 2018, 260–261, 31–40. [Google Scholar] [CrossRef]
- Yuan, M.; Zhao, L.; Lin, A.; Wang, L.; Li, Q.; She, D.; Qu, S. Impacts of preseason drought on vegetation spring phenology across the Northeast China Transect. Sci. Total Environ. 2020, 738, 140297. [Google Scholar] [CrossRef] [PubMed]
- Stöckli, R.; Vidale, P.L. European plant phenology and climate as seen in a 20-year AVHRR land-surface parameter dataset. Int. J. Remote Sens. 2004, 25, 3303–3330. [Google Scholar] [CrossRef]
- Yu, Z.; Chen, L.; Li, L.; Zhang, T.; Yuan, L.; Liu, R.; Wang, Z.; Zang, J.; Shi, S. Spatiotemporal Characterization of the Urban Expansion Patterns in the Yangtze River Delta Region. Remote Sens. 2021, 13, 4484. [Google Scholar] [CrossRef]
- Ding, H.; Bu, Y.; Xu, L. Spatial and temporal variation of the plant phenology and its response to the urbanization trend in the Yangtze River delta. J. Saf. Environ. 2021, 21, 1352–1360. [Google Scholar] [CrossRef]
- Wohlfahrt, G.; Tomelleri, E.; Hammerle, A. The urban imprint on plant phenology. Nat. Ecol. Evol. 2019, 3, 1668–1674. [Google Scholar] [CrossRef]
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, C.; Deng, K.; Peng, D.; Jiang, L.; Zhao, M.; Liu, J.; Qiu, X. Spatiotemporal Characteristics and Heterogeneity of Vegetation Phenology in the Yangtze River Delta. Remote Sens. 2022, 14, 2984. https://doi.org/10.3390/rs14132984
Yang C, Deng K, Peng D, Jiang L, Zhao M, Liu J, Qiu X. Spatiotemporal Characteristics and Heterogeneity of Vegetation Phenology in the Yangtze River Delta. Remote Sensing. 2022; 14(13):2984. https://doi.org/10.3390/rs14132984
Chicago/Turabian StyleYang, Cancan, Kai Deng, Daoli Peng, Ling Jiang, Mingwei Zhao, Jinbao Liu, and Xincai Qiu. 2022. "Spatiotemporal Characteristics and Heterogeneity of Vegetation Phenology in the Yangtze River Delta" Remote Sensing 14, no. 13: 2984. https://doi.org/10.3390/rs14132984
APA StyleYang, C., Deng, K., Peng, D., Jiang, L., Zhao, M., Liu, J., & Qiu, X. (2022). Spatiotemporal Characteristics and Heterogeneity of Vegetation Phenology in the Yangtze River Delta. Remote Sensing, 14(13), 2984. https://doi.org/10.3390/rs14132984