Abstract
Lemongrass (Cymbopogon citratus) essential oil (EO) is a major source of bioactive compounds (BC) with anticancer activity such as α-citral, limonene, geraniol, geranyl acetate, and β-caryophyllene. Comparative studies about cytokinin effects on BC profiles in lemongrass are missing. Here, we evaluated four cytokinins (2iP, tZ, BAP, and KIN) in two different osmotic media, MS-N (3% sucrose, 3 g L−1 Gelrite™) and MS-S (5% sucrose, 5 g L−1 Gelrite™). It results in a higher multiplication rate in BAP containing medium compared to tZ, KIN, and 2iP (p ≤ 0.05). While shoots grown on MS-N/BAP, tZ, and KIN exhibited a highly branching morphology, MS-N/2iP produced a less branching architecture. BC profile analysis of established plants in pots revealed that their maxima production depends on the in vitro shoot growth conditions: i.e., highest content (80%) of α-citral in plants that were cultured in MS-S/BAP (p ≤ 0.05), limonene (41%) in MS-N/2iP, or geranyl acetate (25.79%) in MS-S/2iP. These results indicate that it is possible to increase or address the production of BC in lemongrass by manipulating the cytokinin type and osmotic pressure in culture media. The culture protocol described here is currently successfully applied for somatic embryogenesis induction and genetic transformation in lemongrass.
1. Introduction
Plants from the genus Cymbopogon (Panicoideae subfamily of the Poaceae, or grass family) are perennial medicinal herbs and aromatic plants with more than 55 species distributed in tropical and subtropical countries. They are native to India, Sri Lanka, and Malaysia. In México, the best-known and most widespread species is Cymbopogon citratus, also known as lemongrass. Cymbopogon species produce essential oil (EO), conferring these plants their characteristic fragrance. EO mainly contains monoterpenes and sesquiterpenes as bioactive compounds (BC). Citral, nerol, geraniol, citronellal, linalool, limonene, and geranyl acetate, correspond to the first class. In comparison, β-caryophyllene and humulene correspond to sesquiterpenes [1]. Because of its aroma, which is similar to the lime fruit, the EO is of great interest in the flavor, fragrances, perfumery, cosmetics, soaps, and pharmaceutical industries [1]. The EO constituents of the genus Cymbopogon are bioactive compounds with several tested biological activities such as antioxidant, anti-inflammatory, antifungal, antibacterial, and anticancer properties with potential use in medicine [2]. Different types of cancer are among the most common leading causes of death worldwide [3,4]. Citral has shown anticancer activity in various human cancer cell lines, including cervical [4] and stomach [5] cancer cell lines. At the same time, different studies have shown the activity of geraniol against lung, prostate, bowel, liver, colorectal, kidney, and skin cancer [6]. Other studies also have reported that limonene prevents the progression of mammary and liver cancers [7]. Likewise, geranyl acetate has potent antifungal and anti-inflammatory activities [8]. Teixeira et al. [9] also evaluated nerol derivatives activity against cancer cell lines. In addition, β-caryophyllene also exhibited antiproliferative effects against colorectal [10] breast, pancreas, lymphoma, and melanoma cancer cells [11].
Today, medicinal plants are of great importance in the health industry, and almost 25% of modern medicines are obtained or derived from natural sources [12]. The use of biotechnologies based on plant cell cultures is more promising, sustainable, and environmentally friendly than traditional cultivation of medicinal plants, allowing better control of the quality and higher yields of the desired secondary metabolites [12].
Different studies deal with diverse ways to improve EO productivity and yield, as well as selection of high-yielding genotypes of Cymbopogon species [13,14,15,16,17]. It is also well known that water stress significantly affects plant growth and development and might induce a high accumulation of secondary metabolites. As part of the water stress response, plants make morphologic changes, intracellular osmotic adjustments increasing the sugar content in roots and leaves and solute accumulation. Plants also express antioxidant defense mechanisms and enhance the content of monoterpenes and sesquiterpenes [18]. The EO of some herbs and medicinal plants is reported to be increased by water stress. For instance, the content of monoterpenes in Salvia officinalis is increased by drought stress [19]. In Artemisia annua, various volatiles are also increased by water deficit treatment [20].
In addition to environmental conditions, exogenous application of plant growth regulators influences terpene biosynthesis and the EO composition and production in aromatic plants [15]. Previous studies reported using plant growth regulators to enhance the productivity and yield of lemongrass EO, such as triacontanol [14], and more specifically, the use of exogenous cytokinins (CKs) to enhance monoterpene biosynthesis. CKs are a group of plant growth regulators that play an essential role during the cell cycle, influencing plant growth and development [21]. Natural plant (isoprenoid) CKs are synthesized through the mevalonic acid and MEP pathways. Terpenes are the most widely secondary metabolites distributed in plants; these and terpenoids are made up of isoprenoid precursors. The enzyme HMGR (3-hydroxy-3-methyl glutaryl-CoA reductase) is crucial in the mevalonic acid pathway that zeatin upregulates [22]. This regulatory interaction reveals the vital role and connection of CKs with secondary metabolism.
Therefore, in several species of aromatic plants such as spearmint [17], peppermint [23], and C. citratus [15,24], CKs have been used to increase monoterpenes biosynthesis. Phenylurea and BAP are the CKs that have been preferentially used in those works, and both of them are non-naturally produced CKs. Nevertheless, no research has shown a comparative study on the effect of four structurally different CKs on in vitro shoot multiplication of lemongrass associated with their terpenoid profiles. In this study, we asked about the impact of natural isoprenoid and aromatic CKs on the EO content and composition of lemongrass. Thus, the isoprenoid CKs 2-isopentenyl adenine (2iP) and trans-zeatin (tZ), as well as the aromatic compounds 6-benzylaminopurine (BAP) and kinetin (KIN) [25,26,27], were assayed. First, we aimed to know how the in vitro growth and development of C. citratus are affected by different types and concentrations of CKs. To determine which CK and quantity are the best to induce the highest shoot multiplication and to know which CK is more associated with the highest content of terpenoids. To achieve these goals, we developed an efficient method for in vitro C. citratus propagation starting with the induction of new shoots from seedling meristems. We also used different osmotic media to understand their effect on in vitro shoot multiplication, EO contents, and composition. We analyzed SPME/GC-MS EO profiles of acclimatized plants in pots on soil derived from in vitro treated shoots. Our results showed that different CK treatments affected shoot number, shoot multiplication index, and shoot branching morphology. In addition, SPME/GC-MS analysis of EO profiles from leaves of established plants showed that EO content and composition varied among treatments. These variations were also related to the CK type and in vitro shoot branching morphology. The details and results of these experiments are presented and discussed below.
2. Materials and Methods
2.1. Seed Germination
C. citratus seeds were obtained from a Poland commercial vendor (W. legutko przedsiębiorstwo hodowlano-nasienne sp. z o.o., Jutrosin, Poland; www.legutko.com.pl, accessed on 22 June 2022) and were sterilized through gasification with chlorinated gas (Cl2) in a vacuum chamber for 15 min. Chlorine gas was formed by mixing 50 mL of a commercial bleaching agent (1.2% sodium hypochlorite) with 50 mL of 6N HCl. Seeds were washed with 20% NaCl + TWEEN® 20 for 10 min and finally rinsed six times with sterile distilled water. For germination, seeds were sown in Petri dishes on MS medium [28] with or without activated charcoal (Supplementary Figure S1A–C) and incubated in the dark at 26 °C.
2.2. In Vitro Shoot Culture Establishment and Propagation Protocol
Germinated 3-day-old seedlings were subcultured to fresh MS medium for shoot proliferation (Supplementary Figures S1 and S2). This medium contained 3% sucrose, 0.3% Gelrite™, and 2.0 mg L−1 BAP (pH 5.8) and autoclaved at 121 °C, 1.14 kg cm−2 for 15 min. After two weeks of incubation in this medium, several new shoots were regenerated. These newly formed shoots were allowed to progress their development for two additional weeks in the Petri dish (Supplementary Figures S2(Aa,Ab)) at 16/8-light/dark photoperiod with 54 µmol m−2 s−1provided by Sylvania fluorescent white lamp lights. The incubation temperature was 26 °C. Developed plantlets were individualized and subcultured into the same fresh MS medium every 4 weeks.
To clarify the terminology used in this work, in vitro shoot propagation of lemongrass is initiated by lateral meristems (axillary meristems) already present in the initial shoots, which develop their leaf primordia, becoming an axillary bud. New axillary buds produce lateral shoots also called tillers. A tiller is a specialized shoot branch, which consists of a stem segment, leaves, and emerging leaf meristems. In this work, we will refer to shoots as the whole lateral shoot or tiller (Supplementary Figure S6). Shoot branching refers to the patterning, which means the entire aerial morphology (shoot branching architecture) in a grass plant, composed of the number of shoots, and the number of leaves and their spatial disposition [29,30,31].
2.3. Effect of Sucrose and Gelrite™ Concentrations on Shoot Propagation
To understand the influence of sucrose and Gelrite™ concentration on in vitro shoot propagation of lemongrass, we compared the effect of two different semisolid media prepared with distinct concentrations of sucrose and Gelrite™. Here, we introduce the MS-S (S = “stress”) that was the same as MS-N but with 5% sucrose and 0.5% Gelrite™. The higher concentrations of sucrose and gelling agent conferred a water stress-like condition to the explants. Two-week-old shoots established after three rounds of subculture on MS-N were individualized. Single shoots were placed in 100 mL flasks with 30 mL of MS-N or MS-S medium containing 2.0 mg L−1 BAP and incubated for four weeks in the same environmental conditions described for shoot culture establishment.
2.4. Effect of Culture Media and Cytokinin Type and Concentration on Shoot Propagation
2-week-old shoots previously grown on MS-N with 2.0 mg L−1 BAP were subcultured onto MS-N or MS-S media complemented with different concentrations (0.5, 1.0, 2.0 mg L−1) of 2iP, KIN, BAP, and tZ (Sigma-Aldrich, St. Louis, MO, USA) (Supplementary Figure S3 and Table S1). Three individualized shoots about 3–4 cm high were sown in each 200 mL flask for each treatment. In this case, each CK treatment was considered the control for the other treatments. Although the strict control treatment should be in vitro shoots grown on MS medium without any CK or plant growth regulator. However, in these plants, the shoot morphology and development consisted of just one tiller and an inferior root system that did not show the typical phenotype or natural morphology of C. citratus (Supplementary Figure S4). Then we considered these plants inappropriate to use as a control treatment.
2.5. Experimental Design
We used a multifactorial (2 × 4 × 3) experimental design with different levels of each tested factor: (1) MS culture media (MS-N and MS-S); (2) Four different CKs (2iP, KIN, BAP, and tZ); and (3) Different CK concentrations (0.5, 1.0, and 2.0 mg L−1) containing three replicas. Thus, 24 treatments were evaluated (Table S1) and repeated three consecutive times. The results were analyzed through a multifactorial ANOVA and Tukey’s test (p ≤ 0.05) [32] (Tables S2 and S3). The statistical procedure was done with GraphPad Prism 8 software for Windows (www.graphpad.com, accessed on 22 June 2022).
2.6. Root Induction
When required, four-week-old shoots micropropagated on MS-N supplemented with 2.0 mg L−1 BAP were transferred to MS basal medium without growth regulators for rooting. Shoots transferred to this medium were incubated in a growth chamber with fluorescent lamps at 54 μmol m−2 s−1light intensity, 26 °C, and a 16/8 h light/dark photoperiod.
All the treatments with CKs gave place to root formation except for 1.0 and 2.0 mg L−1 BAP. Therefore four-week-old shoots micropropagated on MS-N or MS-S supplemented with 1.0 or 2.0 mg L−1 BAP were transferred to MS basal medium without growth regulators for two weeks to permit the rooting before passing them to pots with soil.
2.7. Acclimatization of In Vitro Rooted Shoots
After four weeks of in vitro culture, rooted shoots were removed from flasks, washed several times with distilled water, then transferred to 500 mL pots with 350 mL of a soil combination of organic humus with Agrolita® and Perlite® (3:1:1 v/v) and incubated in a growth room (28 °C, RH 35%) for acclimatization. Pots with plantlets were covered with a plastic bag during the first week to maintain high humidity and avoid desiccation. Plastic bags were pored to decrease the humidity and acclimatize the plants to the external environment. After one month of hardening, plants were transferred to 2.5 L pots with the same proportion of soil mixture under greenhouse conditions (Supplementary Figure S5).
2.8. SPME/Gas Chromatography-Mass Spectrometry Analysis
We took leaves from 4-week-old acclimatized plants for EO profile analyses, cut them in about 5 cm, and freeze-dried them. Afterward, we ground the lyophilized pieces of leaves in a mortar with a pestle, and 50.0 mg of the powder was rapidly transferred to a clean 50 mL Erlenmeyer flask covered with aluminum foil and subjected to SPME (solid phase microextraction). For SPME, we used a 50/30 µm DVB/CAR/PDMS fiber assembly from Supelco (Sigma-Aldrich, Burlington, MA, USA). The needle with the fiber was passed through the aluminum foil that covered the flasks, and after 5 min exposure to volatiles, the fiber was introduced into an Agilent 5973 inert gas chromatograph/mass spectrometer (GC/MS) coupled to an Agilent 5973N mass selective detector. An Agilent capillary column HP-5of 30 m × 250 μm in diameter × 0.25 μm in thickness (19091J-433: USC353412K −60–325 °C (325 °C) was used. Helium, at a flow rate of 0.6 mL/min was used as the carrier gas. The initial temperature in the column was 60 °C and then increased to a maximum temperature of 260 °C. The column pressure corresponded to 3.0638 psi. Run time was 26.25 min with a post-run time of 2 min. The EO constituents were identified by their fragmentation patterns of mass spectra compared with data stored in NIST-MS-2010 through the Acquisition mode Mass Hunter\GCMS\2\5973N\attune. The quantitative analysis was carried out in triplicate.
3. Results
3.1. Effect of Sucrose and Gelrite™ Concentrantions on Shoot Development
After two weeks, plants grown on MS-S (“stress” medium) began to show qualitative phenotypic differences compared to shoots grown on MS-N (“normal” medium; Supplementary Figure S2B). Shoots generated on MS-S had leaves that were darker green and more coriaceous than leaves on MS-N. Some senescent leaves were also present on MS-S but absent in MS-N (Supplementary Figure S2(Bc,Be,Bd,Bf)). The above observations were evident in 2iP, tZ, BAP, and tZ during shoot propagation on MS-S.
3.2. Effect of Cytokinin Type and Concentration on Shoot Propagation and Shoot Morphology
Table 1 shows the average number of shoots per flask, and the multiplication index per individual shoot is given for each CK treatment. The initial shoot cultures from germinated seeds were established using 2 mg L−1 BAP. The highest multiplication rates per flask were obtained at 2 mg L−1 BAP in MS-S (23.3), although near higher levels are observed in all BAP concentrations in MS-N. In contrast, we recorded the lowest multiplication rates per flask with 2iP in MS-S (4.4) (Table 1).

Table 1.
Effect of the type of CK and culture media on shoot multiplication.
A statistical analysis (ANOVA) of the data shown in Table 1 demonstrated that CK concentrations tested in the range of 0.5–2.0 mg L−1 were not significantly different (p ≤ 0.05) for shoot multiplication. Therefore, Figure 1 shows a representative in vitro shoot culture experiment with the four CK treatments. Photos in this figure show that micropropagated shoots induced with different types of CKs produced different shoot branching morphologies depending on which CK was supplied to MS-N or MS-S media. Shoots grown on 2iP, tZ, BAP, and KIN showed variations in shoot number when they were propagated on MS-N and MS-S. Propagated shoots on MS-N with KIN, BAP, and tZ showed a bushier pattern in the aerial part compared to shoots reproduced on 2iP. Shoots grown on MS-S with KIN, BAP, and tZ showed variation in shoots, with less quality.

Figure 1.
The effects of different types of CKs on plant morphology during micropropagation of C. citratus shoots cultured on MS-N and M-S media.
Figure 2A shows the average shoot number produced by in vitro shoots cultured on MS-N and MS-S. Figure 2B,C show a graphical representation of statistical analysis based on the shoot number per flask obtained from the average of the three concentrations. The number of the newly formed shoots significantly differed according to the type of CK (Figure 2B,C). tZ, BAP, and KIN significantly (p ≤ 0.05) induced a higher number of shoots than 2iP (Table 1, Figure 2A). BAP produced the highest number of shoots in MS-S (Table 1, Figure 2B), whereas 2iP and tZ had the lowest (Figure 2A,B). The blue and red columns on the graph show significant (p ≤ 0.05) differences in the effect of MS-N and MS-S on the number of shoots produced by the different CKs. The height of the blue and red columns represents the average number of shoots obtained from the three concentrations (0.5, 1.0, and 2.0 mg L−1) for each CK. (Tables S2 and S3).
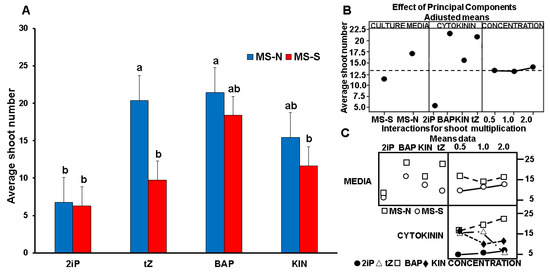
Figure 2.
(A) Average shoot number of in vitro shoots produced after four weeks of culture on MS-N and MS-N with different CK types and concentrations. Results represent the average values of three independent replicates. The a, b, and c lowercase letters over the columns indicate the result of Tukey’s test (p ≤ 0.05) after comparing the various treatments. (B) Representation of the multifactorial statistical analysis of variance on the main factors’ different effects in our experiment. (C) Result of the relationships of variables on plant shoot production.
3.3. Direct Root Formation Was Induced in Lemongrass Shoots during Clonal Propagation
After two weeks of culture, all plant shoots growing on 0.5, 1.0, and 2.0 mg L−1 2iP, KIN, and tZ began to produce roots on both MS-N and MS-S media (Supplementary Figure S7). On MS-N, roots grew faster for 2iP and tZ treatments than for KIN and BAP. On MS-S, root formation was more evident on 2iP treatments than on KIN, BAP, and tZ. After four weeks of culture on MS-N, vigorous roots continued developing mainly in the three concentrations of 2iP and tZ on MS-N, exhibiting profuse growth as shown in Supplementary Figure S8. After four weeks, shoots growing on MS-N at 0.5 and 1.0 mg L−1 KIN showed more developed roots than in the first two weeks, while roots from shoots grown on BAP did not show similar progress except at the lowest concentration of 0.5 mg L−1 (Supplementary Figure S8). On MS-S, shoots grown on 2iP and KIN produced more profuse roots than tZ for all three tested concentrations.
Interestingly, although the concentration of 0.5 mg L−1 BAP produced rooted shoots on MS-N and MS-S, the shoots had more profuse roots in MS-S (Supplementary Figure S8). As mentioned, shoots treated with 1.0 and 2.0 mg L−1 BAP must be transferred to a medium free of CKs to permit rooting. These exhibited a different root phenotype than those treated with the other CKs. These roots were shorter, white, and thinner than the thick and longer purple roots produced by in vitro shoots induced with 2iP, tZ, and KIN (Supplementary Figure S9A–D).
3.4. Main Constituents of BC Profiles of Plants in Pots
We start validating our GC-MS method for solid-phase microextraction (SPME).
Firstly, we compared the profile of a citral analytical standard sample (a mixture of neral and geranial from AMCO International SA de C.V. México) against the GC-MS profile obtained from fresh leaves of an adult plant of lemongrass grown in the field around (Supplementary Figure S10). Neral (β-citral) and geranial (α-citral) mass spectra and retention times (RT) obtained from SPME/GC-MS analysis of citral analytical standard were identical to those of C. citratus leaves (Table 2). The SPME/GC-MS analysis of C. citratus essential oil from the different CK treatments identified many constituents, though most were present in small quantities (less than 1%). About fourteen typical components of C. citratus EO were identified: hexenol, citronellal-like monoterpenes, terpinene, cadinene, limonene, ocimene, cymene, cis-geraniol (nerol), trans-geraniol (geraniol), neral (β-citral), geranial (α-citral), geranyl acetate, and sesquiterpenes such as β-caryophyllene and α-humulene [33]. In Table 2, we summarize the most constant marker components present in the essential oil of established plants of C. citratus.
Table 2.
Chemical composition of the main volatile oils obtained from lyophilized (freeze-dried) leaves of established plants in pots. RT = retention time; (%) = maximum content.
Values represent the average area % of the 0.5 mg L−1concentration obtained for each terpenoid among the different treatments. Area % represents the total sum of the abundance of all terpenoids in the EO profile divided by the average quantity of each constituent × 100.
In all treatments, the monoterpene fraction was dominated by β- and α-citral, ranging from 15% to 80% of the total terpenes’ EO profiles. Limonene ranged from 1 to 41%, and the sesquiterpene β-caryophyllene ranged between 1 and 5% of the full content. Other monoterpenes were less present than 5% considered altogether.
3.5. BC Profiles of Lyophilized Leaves of Established Plants in Pots
Figure 3A,B shows a representative example of the comparison of SPME/GC-MS EO profiles obtained from lyophilized leaves. These EO profiles correspond to established plants from shoots grown on MS-N or MS-S and with 0.5 mg L−1 2iP, tZ, BAP, or KIN. We decided to compare the 0.5 mg L−1 concentration of the different CKs because it permits the direct development of roots in the treatments (Supplementary Figures S7 and S8).
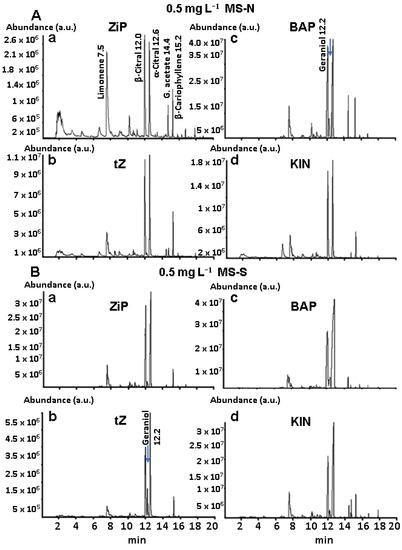
Figure 3.
Terpenoid profile of essential oils from leaf samples of established plants in pots. Plants came from shoots grown on MS-N and MS-S media (A,B) with 0.5 mg L−1 of the different CKs (a–d). Terpenoid names and retention times are shown. The blue arrow indicates the presence of geraniol.
Plants obtained from MS-N added with 2iP show a significant production of limonene regarding the other conditions. Additionally, maybe the most crucial qualitative change observed is the emergence of geraniol (RT = 12.2 min) in between the peaks for β- and α-citral in the EO profile of MS-N containing BAP (Figure 3A) and in the EO profiles of MS-S containing 2iP, tZ, BAP, and KIN (Figure 3B). Several peaks in MS-S diminished, except those of limonene, geraniol, β-citral, α-citral, and β-caryophyllene.
3.6. Quantitative Analysis of the Main Constituents of BC Profiles of Established Plants in Pots
Figure 4 presents a comparative analysis of the relative peak areas (%) of the EO constituents shown in Figure 3. Quantitative analyses were carried out in triplicate by obtaining the abundance (peak area) of each of the constituents in the EO profile in each sample. Notably, the highest fraction corresponding to α-citral in EO profiles was significantly (p ≤ 0.05) induced with BAP (80%) in MS-S (Figure 4A,D). β-citral followed in abundance in derived plants grown on MS-N or MS-S (Figure 4A,D). While KIN and BAP significantly induced the highest quantity of α-citral, there were no significant differences among CKs to yield β-citral in both MS-N and MS-S media (Figure 4A,D). In the case of the individual contents of the less present terpenes, limonene and β-caryophyllene are produced in both media. 2iP significantly (p ≤ 0.05) induced the highest amount (41.61%) of limonene in plants that grew on MS-N, while KIN did the same in plants grown on MS-S (Figure 4B, E). BAP induced the highest abundance of β-caryophyllene (2.89%) in micropropagated plants from shoots grown on MS-S (Figure 4E). Additionally, as we can observe, except for limonene (41%) (Figure 4B), the relative individual contents (<5%) of the other terpenes were similar in plants grown on MS-N and MS-S (Figure 4B,E). While the sum of less abundant volatiles was higher in plants induced with 2iP (MS-N) versus the other CKs (Figure 4C), the contrary happened in MS-S, although any CK produced above the values on MS-N (Figure 4F).
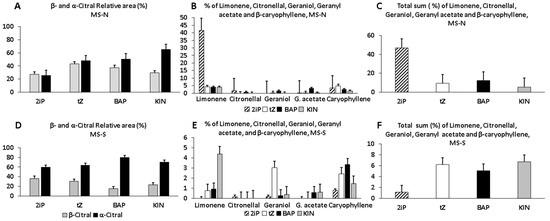
Figure 4.
Quantitative analysis of relative abundance (%) of essential oil detected from lyophilized leaf samples of C. citratus grown in pots derived from in vitro shoots treated with 0.5 mg L−1 each CK. (A,D) Effect of each CK on the content (%) of the major EO bioactive compounds, β- and α-Citral, in leaves of established plants derived from in vitro shoots grown in MS-N or MS-S. (B,E) Effect of the different CKs on the individual content (%) of each minor EO bioactive compound quantified from established plants derived from in vitro shoots grown on MS-N or MS-S. (C,F) The sum of the contents (%) of the five minor EO bioactive compounds produced by each CK treatment showed in (B) and (E), respectively.
4. Discussion
This research describes a simple protocol for in vitro clonal micropropagation of C. citratus, starting from seeds, and multiplied shoots until established plants in pots. During the process, regenerated shoots were subject to two media with different osmotic conditions and three concentrations of the CKs 2iP, tZ, BAP, and KIN. Finally, lyophilized pieces of leaves from plants in pots were analyzed from the EO profile by GC-MS. The critical points of the research are discussed below.
4.1. CK Treatments Affected Shoot Number and Shoot Branching Morphology
Among the different CK treatments (Figure 1 and Figure 2), tZ, BAP, and KIN did not show significant (p ≤ 0.05) differences in multiplication rates and multiplication index. However, 2.0 mg L−1 of BAP induced the highest number of shoots per flask grown on both MS-N (21.9) and MS-S medium (23.3) (Table 1, Figure 2). This result is an impressive effect caused by BAP. However, looking at the literature, we find that among different concentrations of BAP, tZ, or KIN, BAP also induced the highest biomass shoot growth of in vitro shoots of Thymus vulgaris L. [34]. In addition, our work shows that 2iP generated the lowest number of in vitro shoots. 2iP also resulted in a different morphology displaying a reduced shoot branching with elongated tillers compared to the bushy morphology caused on MS-N by KIN, BAP, and tZ (Figure 1). The less branching shoot morphology induced by 2iP in MS-N and by 2iP and KIN in MS-S (Figure 1) resembled the activity of strigolactones during tiller outgrowth in rice [35]. The role of CKs is complex to understand since these have also been implicated in shoot branching regulation in Arabidopsis and monocots, including maize and rice [36].
After four weeks of culture, we observed the highest number of newly formed shoots per flask in MS-N with 0.5–2 mg L−1 BAP, which gave a multiplication index of 7.1–7.3, and in MS-S at 2 mg L−1 BAP with a multiplication index of 7.8 (Table 1). This multiplication index was higher than previously reported for lemongrass shoot multiplication in semisolid medium and temporary immersion systems [37,38,39]. Accordingly, the addition of BAP to MS medium also resulted in the highest number of shoots per explant of in vitro shoot cultures of Lantana camara L. [40].
4.2. Direct Root Formation and Development Are not Related to Essential Oil Production
Root formation happens in all 2iP, tZ, and KIN concentrations during in vitro shoot propagation ( Figures S7 and S8). This rooting was unexpected because CKs are considered negative regulators of root growth and lateral root formation [41] and have not been previously reported for in vitro propagation of C. citratus. However, since 1998, this phenomenon had already been registered by Kamenicka and colleagues [42]. They found that the CKs BAP and KIN riboside (KR) produced roots from in vitro shoots of Rhododendron forrestii. However, in this work, BAP did not induce a root system, except in vitro shoots propagated on MS-N and MS-S at the lowest quantity of 0.5 mg L−1 (Supplementary Figures S7 and S8). This different effect of BAP in two plants implies a differential response of plants to the same CKs. The roots developed in the presence of 2iP, tZ, and KIN were generally thick, with intense purple pigmentation, and longer compared to the white and thinner roots grown on a cytokinin-free basal medium (Supplementary Figure S11). Considering the EO profile production, we cannot correlate root development to essential oil production. For instance, in vitro shoots grown on 0.5, 1.0, or 2.0 mg L−1 KIN produced roots, and at the same time, 1.0 or 2.0 mg L−1 BAP did not, and there were no significant differences in the production of β- and α-Citral between tZ and BAP at 0.5 mg L−1.
4.3. Water Stress Medium (MS-S) Altered the Number and Morphology of In Vitro Shoots
Our results showed that increasing sucrose and Gelrite™ concentrations to 5% and 0.5%, respectively, created a simulated water stress-like that altered the number and morphology of shoots grown on MS-S medium (Figure 1) and stimulated direct root formation in C. citratus shoot cultures (Supplementary Figures S7 and S8). Different concentrations of sucrose and other monosaccharides in the media also promoted shoot multiplication, biomass accumulation, and higher regeneration rates in C. pulegium [43]. In the explant medium, osmotic agents such as sucrose, mannitol, or sorbitol can simulate a water-stress response [44]. Increasing the gelling agent concentration can produce the same effect [45]. Different concentrations of distinct gelling agents in MS medium affected in vitro shoot phenotypes of various plant species [46,47]. Besides the effect of sucrose and Gelrite™ concentration, adding 2.0 mg L−1 BAP to the water stress MS-S medium exhibited shorter shoots, with the presence of senescent leaves as the most noticeable difference (Figure 1, Supplementary Figure S2(Ba,Bb)). A possible explanation is that exogenous BAP could be metabolized differently in the water stress MS-S medium compared to the MS-N medium. This difference could be feasible since differences in BAP glycosylation, stability, permeability, and transport have been reported under water stress conditions [48]. Endogenous biosynthesis of CK under these conditions may help relieve the water stress imposed by MS-S. CK and cell wall invertases have been reported to confer stress tolerance in tobacco and tomato [49,50]. We can assume that growing shoots could assimilate the sucrose from the media. Hence, we might suggest that CK and sucrose metabolism might be involved in the stress tolerance response of C. citratus shoots to water stress impaired by MS-S. More study requires on how CK and sucrose metabolism is involved in the regulatory networks controlling bud outgrowth [51,52].
4.4. CK Treatments Caused Qualitative and Quantitative Changes in BC Profiles
BC profiles showed qualitative and quantitative (Figure 3) variations depending on the type and concentration of CK used. For example, Figure 3 shows that trans-geraniol (geraniol, RT = 12.20) appears only in some BC profiles of acclimatized plants (Figure S3(Ab,Bc)).
The pattern of EO terpenes also varied quantitatively between MS-N and MS-S culture media (Figure 3). The content of these terpenes was similar to or higher than the terpene contents reported by Trang et al. [53] and Kim et al. [54] extracted by hydrodistillation and by Ribeiro et al. [40] adsorbed by SPME. According to quantitative variations, different concentrations of BAP and tZ induced quantitative changes in the production of the main volatile compounds of in vitro shoots of Thymus vulgaris L. [34]. In general, terpene content (%) differed between EO profiles of MS-N (3% sucrose) and MS-S (5% sucrose) (Figure 4). For instance, in MS-N, the highest content of α-citral (64.96%) was induced by KIN, while in MS-S, the highest content of α-citral (80%) was induced by BAP (Figure 4). In addition, the content of β-citral ranged from 27.52% (2iP) to −42.99% (tZ) in MS-S, while the content of α-citral ranged higher, from 25.62 (2iP) to −64.96% (KIN). Likewise, the content of β-citral ranged lower (14.89–36.28%) in MS-S than in MS-N, but the content of α-citral ranged higher (59.48–80.04%) in MS-S than in MS-N. Overall, the content of β-citral decreased by 31.1% from MS-N to MS-S, whereas α-citral increased by 30.82%. Furthermore, when the effect of each CK was analyzed concerning α-citral, it was found that the content of α-citral was relatively increased in established plants derived from MS-S medium. The increments were 56.8% (2iP), 24.5%(tZ), 38% (BAP), and 7.54% (KIN) in relation to MS-N. This increase was also consistent with the result that different concentrations of sucrose in culture media significantly affected the composition of the volatile profile in C. pulegium [43] (Figure 4). Besides the contents of β- and α-citral, high limonene content (41%) was reached in plants derived from MS-N. Additionally, the content of β-caryophyllene (12.3%) was higher in established plants derived from MS-S with BAP (Table 2, Figure 4). Moreover, it has been reported that terpenoids accumulate in Artemisia annua concomitantly with a high expression of HMGR. The rate-limiting enzyme of MVA (mevalonic acid pathway/terpenoid biosynthesis pathways) increases under water stress as an adaptive response [20].
4.5. Variations in BC Profiles Related to Shoot Number and Shoot Branching Morphology Could Be Explained Because Plants Sense Distinct CK Types Differently
Structurally distinct CKs are sensed differentially in Arabidopsis by different histidine kinase (AHK) receptors [55,56]. Also, CK catabolism differs between isoprenoid and aromatic CKs [57,58,59,60,61,62]. These differences may explain why in various reports, including this work, the use of a different type of CK has influenced the plant EO content and composition, such as in Catharanthus roseus [63], spearmint [17], Mentha arvensis [64], Mentha piperita [23], Thymus vulgaris [34], and Artemisia alba [65,66]. In addition, as we mentioned before, the highest content of α-citral (80%) was detected in micropropagated plants grown on BAP (Figure 3 and Figure 4). Therefore, we suggest that BAP could also be involved in regulating HMGR activity during citral biosynthesis. In support of this trial, the activity of HMGR was upregulated by the effect of the CK zeatin and other plant growth regulators [22]. Because BAP also produced the highest number of shoots per flask in MS-S (Table 1, Figure 2), it also might be involved in controlling axillary meristem growth and development. In this regard, it has been found that exogenous CK supplied to the in vitro growth media influences the morphogenesis and terpene profile of EO [67] and that terpene profiles are tightly associated with the morphologic development of in vitro shoot cultures [65]. Notably, a direct correlation was reported between BAP concentration and citral accumulation in C. citratus plants [15].
On the contrary, in vitro shoots grown on MS-N with 2iP displayed a reduced branching morphology with direct root formation (Figure 1), which was related to the lowest content (p ≤ 0.05) of α-citral but to the highest range (p ≤ 0.05) of limonene in the EO analysis. Geraniol, which was another monoterpene detected in the EO of lemongrass (Figure 3), is the precursor of citral biosynthesis (Figure 5) [29] and may play a role in regulating citral abundance and activity in C. citratus plants.
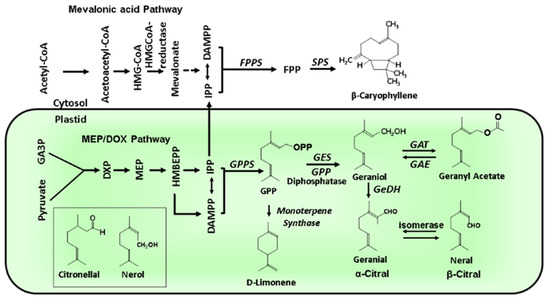
Figure 5.
MEP/DOX and mevalonate pathways in plants. The two main biosynthetic routes involved in terpenoid biosynthesis are shown. The figure shows the chemical structures of the main monoterpenes and β-caryophyllene. GPPS geranyl diphosphate synthase, GES geraniol synthase, GPPD geranyl diphosphate diphosphatase, GAT geraniol acetyl-CoA acetyltransferase, GAE geranyl acetate esterase, GeDH geraniol dehydrogenase, FPPS farnesyl diphosphate synthase, SPS sesquiterpene synthase, MEP methyl erythritol phosphate, DOX deoxy-xylulose 5-phosphate [68,69].
4.6. BC of Lemongrass Might Regulate Axillary Meristem Growth and Development
Many studies evidence the anticancer activity of citral, limonene, citronellal, geraniol, geranyl acetate, and β-caryophyllene. All these were detected in the EO of lemongrass (Table 2). Still, scarce data are available on their specific mode of action to induce apoptosis and block the proliferation of carcinogenic human cell lines [3,53]. In addition, each BC induces apoptosis using different mechanisms of action even to stop cell proliferation and cell cycle arrest in various types of cancer. To be updated, we recommend the reader to consult the following reviews [2,70]. It has been reported that limonene and geraniol inhibit the activity of HMGR in various human cancer cell lines. A reduction of isoprenoid and terpenoid biosynthesis is the consequence, arresting the cell cycle and stopping the growth of culture tumor cells [71,72]. Izumi and colleagues [73] also reported that geraniol induces apoptosis, including DNA and nuclei fragmentation in cultured shoot primordia of Matricaria chamomilla, and Ghosh et al. [74] said that geraniol induces senescence in tomatoes. In addition, Chaimovitsh et al. found that microtubules are the intracellular targets of plant terpene citral [75].
Furthermore, β-cyclocitral has recently been reported to induce root apical meristematic cell division and promote root growth in Arabidopsis [76]. Based on these reports, we also suggest exploring the possibility that these lemongrass BC could regulate the cell cycle in plants by inhibiting or increasing HMGR activity and, by the way, be involved in the control of axillary meristem growth and development. Therefore, a better understanding of the effects of CKs on the regulation of plant terpenes biosynthesis, and the mode of action of these terpenes on regulating plant cell cycle and axillary meristem growth, should let us establish and improve lemongrass in vitro propagation protocols. These protocols can permit the manipulation of in vitro shoot outgrowth and increase plant biomass and the content of BC in the EO. Finally, we are addressing a gene network study involving plant mevalonic acid pathway, CK biosynthesis, CK activity and signaling, plant BC and cell cycle checkpoints to help us elucidate the regulation of axillary meristem growth and development by BC to increase lemongrass biomass and BC production.
5. Conclusions
Significant differences were found in the activity of different sources of CKs in shoot multiplication rates, direct root formation, and shoot branching morphology. Sucrose and Gelrite™ concentrations in the basal medium also contributed to these parameters. From the perspective of this work, one of the best ways to obtain higher yields of the EO of lemongrass would be to induce in vitro shoot multiplication with the combination of 2iP and BAP in the culture medium, or 2iP first followed by rooted shoot exposed to BAP. 2iP induces a less branched phenotype than tZ, BAP, and KIN, although with stronger tillers and roots. We also conclude that established plants contained a significant amount of EO components by SPME/GC-MS analysis. Our results indicate that a BC of interest might be manipulated by different concentrations and sources of CK and the type of basal medium. For instance, 2iP shoot morphology was associated with the lowest and BAP and KIN bushier shoot morphology with the highest content of α-citral, the main constituent of the EO. Another conclusion from our results was that we could not state that direct root formation and development are related to EO production. Finally, many EO content and composition examples are influenced by the different growth and developmental stages of young to mature plants. Nevertheless, our results showed that plant morphology might also be related to the EO content and composition. This relationship is a new and compelling result not in the literature yet.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijpb13030025/s1, Figure S1: Germination of Cymbopogon citratus seeds; Figure S2: Shoot in vitro propagation; Figure S3: Chemical structure of CKs used in this work; Figure S4: Control in vitro plant grown on MS (3% Sucrose, 0.3% Gelrite™) without any plant growth regulator. Figure S5: Micropropagated plants of C. citratus; Figure S6: Plant morphology of in vitro shoot cultures of C. citratus; Figure S7: Direct roots formed after two weeks of propagation; Figure S8: Direct roots formed after four weeks of propagation; Figure S9: Root induction in MS basal medium free of hormones; Figure S10: SPMG/GC-MS control profiles; Figure S9: In vitro Roots Phenotype; Table S1: Experimental Design. CK treatments statistical analysis was carried out under a multifactorial (2 × 4 × 3) ANOVA design; Table S2: Two-way analysis of variance for shoot number multiplication data (ANOVA); Table S3: Two-way analysis of variance for multiplication index data (ANOVA).
Author Contributions
Conceptualization, A.C.-R., J.L.C.-P. and A.M.-A.; methodology, A.C.-R., J.L.C.-P. and R.V.-N.; formal analysis, R.V.-N. and A.C.-R.; investigation R.V.-N., A.C.-R., J.L.C.-P. and J.M.-T.; writing—original draft preparation A.C.-R., writing—review and editing, A.M.-A. and A.C.-R.; review, B.P.-A.; project administration A.M.-A.; funding acquisition A.M.-A., A.C.-R. and B.P.-A. All authors have read and agreed to the published version of the manuscript.
Funding
This work was financed through the framework agreement CEIA/CEC/IRA/20210408/006 for collaboration in a research project between Ingredientes Especializados S.A. de C.V. and CINVESTAV-Irapuato.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data generated in this research are presented within the article.
Acknowledgments
We would like to thank Ingredientes Especializados S.A. de C.V. especially to Francisco Martínez and Julio Limón for financing the research project. We want to thank Eliana Valencia for her technical assistance in preparing culture media. We also would like to thank Enrique Ramírez for his technical assistance in the GC-MS study and Ana Lilia Hernández-Orihuela for her technical assistance in the laboratory; we are also grateful to José Antonio Cisneros for taking the professional photos included in the manuscript and also grateful to Daniel Camas-López for the English language edition of this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ganjewala, D.; Luthra, R. Essential Oil Biosynthesis and Regulation in the Genus Cymbopogon. Nat. Prod. Commun. 2010, 5, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Ganjewala, D. Cymbopogon essential oils: Chemical compositions and bioactivities. Int. J. Essent. Oil Ther. 2009, 3, 56–65. [Google Scholar]
- Mukarram, M.; Choudhary, S.; Khan, M.A.; Poltronieri, P.; Khan, M.M.A.; Ali, J.; Kurjak, D.; Shahid, M. Lemongrass essential oil components with antimicrobial and anticancer activities. Antioxidants 2022, 11, 20. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, K. Anticancer effect of lemongrass oil and citral on cervical cancer cell lines. Pharmacogn. Commun. 2013, 3, 41–48. [Google Scholar] [CrossRef]
- Balusamy, S.R.; Ramani, S.; Natarajan, S.; Kim, Y.J.; Perumalsamy, H. Integrated transcriptome and in vitro analysis revealed anti-proliferative effect of citral in human stomach cancer through apoptosis. Sci. Rep. 2019, 9, 4883. [Google Scholar] [CrossRef] [PubMed]
- Maczka, W.; Winska, K.; Grabarczyk, M. One hundred faces of geraniol. Molecules 2020, 25, 3303. [Google Scholar] [CrossRef]
- Singh, B.; Sharma, R.A. Plant terpenes: Defense responses, phylogenetic analysis, regulation and clinical applications. 3 Biotech 2015, 5, 129–151. [Google Scholar] [CrossRef]
- Gonçalves, M.J.; Cruz, M.T.; Tavares, A.C.; Cavaleiro, C.; Lopes, M.C.; Canhoto, J. LígiaSalgueiro Composition and biological activity of the essential oil from Thapsia minor, a new source of geranyl acetate. Ind. Crops Prod. 2012, 35, 166–171. [Google Scholar] [CrossRef]
- Teixeira, R.R.; Da Silva, A.M.; Siqueira, R.P.; Gonçalves, V.H.S.; Pereira, H.S.; Ferreira, R.S.; Costa, A.V.; de Melo, E.B.; Paula, F.R.; Ferreira, M.M.C. Synthesis of nerol derivatives containing a 1, 2, 3-triazole moiety and evaluation of their activities against cancer cell lines. J. Braz. Chem. Soc. 2019, 30, 541–561. [Google Scholar] [CrossRef]
- Dahham, S.S.; Tabana, Y.M.; Iqbal, M.A.; Ahamed, M.B.; Ezzat, M.O.; Majid, A.S.; Majid, A.M. The anticancer, antioxidant and antimicrobial properties of the sesquiterpene β-caryophyllene from the essential oil of Aquilaria crassna. Molecules 2015, 20, 11808–11829. [Google Scholar] [CrossRef]
- Francomano, F.; Caruso, A.; Barbarossa, A.; Fazio, A.; Torre, C.L.; Ceramella, J.; Mallamaci, R.; Saturnino, C.; Iacopetta, D.; Sinicropi, M.S. β-caryophyllene: A sesquiterpene with countless biological properties. Appl. Sci. 2019, 9, 5420. [Google Scholar] [CrossRef]
- Motolinía-Alcántara, E.A.; Castillo-Araiza, C.O.; Rodríguez-Monroy, M.; Román-Guerrero, A.; Cruz-sosa, F. Engineering considerations to produce bioactive compounds from plant cell suspension culture in bioreactors. Plants 2021, 10, 2762. [Google Scholar] [CrossRef] [PubMed]
- Koshima, F.A.T.; Ming, L.C.; Marques, M.O.M. Cymbopogon citratus (DC.) Stapf, com cobertura morta nas estações do ano. Rev. Bras. PI Med. 2006, 8, 112–116. [Google Scholar]
- Khan, Z.H.; Mohammad, F.; Khan, M.M.A. Enhancing the growth, yield and production of essential oil and citral in lemongrass by the application of Triacontanol. Int. J. Agric. Sci. Res. 2014, 4, 113–122. [Google Scholar]
- Prins, C.L.; Freitas, S.P.; Gomes, M.M.A.; Vieira, I.J.C.; Gravina, G.A. Citral accumulation in Cymbopogon citratus plant as influenced by N6-benzylaminopurine and light intensity. Theor. Exp. Plant Physiol. 2013, 25, 159–165. [Google Scholar] [CrossRef]
- Dutta, S.; Munda, S.; Chikkaputtaiah, C.; Lal, M. Assessment of selection criteria for development of high yielding genoptypes using variability parameters in Lemongrass Cymbopogon flexuosus L. J. Essent. Oil-Bearing Plants 2017, 20, 1450–1460. [Google Scholar] [CrossRef]
- Stoeva, T.; Iliev, L. Influence of some phenylurea cytokinins. Bulg. J. Plant Pysiol. 1997, 23, 66–71. [Google Scholar]
- Bhargava, S.; Sawant, K. Drought stress adaptation: Metabolic adjustment and regulation of gene expression. Plant Breed. 2013, 132, 21–32. [Google Scholar] [CrossRef]
- Radwan, A.; Kleinwacter, M.; Selmar, D. Impact of drought stress on specialised meristems: Biosynthesis and the expression of monoterpene synthases in sage (Salvia officinalis). Phytochemistry 2017, 141, 20–26. [Google Scholar] [CrossRef]
- Yadav, R.K.; Sangwan, R.S.; Sabir, F.; Srivastava, A.K.; Sangwan, N.S. Effect of prolonged water stress on specialized secondary metabolites, peltate glandular trichomes, and pathway gene expression in Artemisia annua L. Plant Physiol Biochem. 2014, 74, 70–83. [Google Scholar] [CrossRef]
- Camas-Reyes, A.; Ramírez-Laguna, R.; Jofre-Garfias, A.E.; Cardozo-Martínez, F.; Hernández-Orihuela, A.L.; Molina-Torres, J.; Martínez-Antonio, A. E. coli cultures expressing a synthetic sequence of ptz gene (stz) promoted in vitro direct organogenesis in Nicotiana tabacum L. Pant Cell Tissue Organ. Cult. 2019, 137, 87–100. [Google Scholar] [CrossRef]
- Stermer, B.A.; Bianchini, G.M.; Korth, K.L. Regulation of HMG-CoA reductase activity in plants. J. Lipid Res. 1994, 35, 1133–1140. [Google Scholar] [CrossRef]
- Scavroni, J.; Vasconcellos, M.C.; Valmorbida, J.; Ferri, A.F.; Marques, M.O.M.; Ono, E.O.; Rodrigues, J.D. Rendimento e composição química do óleo essencial de Mentha piperita L. submetida a aplicações de giberelina e citocinina. Rev. Bras. Pl Med. 2006, 510, 40–43. [Google Scholar]
- Craveiro, A.; Barreira, E.S.; Rabi, J.D.D. Estudo sobre o efeito de citocininas na biossintese de monoterpenos. In Proceedings of the 41st Annual SBPC Meeting, Fortaleza, Brazil, 9–15 July 1989; p. 531. [Google Scholar]
- Kamínek, M.; Březinov, A.; Gaudinová, A.; Motyka, V.; Vaňková, R.; Zaăímalová, E. Purine cytokinins: A proposal of abbreviations. Plant Growth Regul. 2000, 32, 253–256. [Google Scholar] [CrossRef]
- da Silva, J. Is BA (6-benzyladenine) BAP (6-benzylaminopurine)? J. Plant Sci Biotechnol. 2012, 6(SI1), 121–124. [Google Scholar]
- Kieber, J.J.; Schaller, G.E. Cytokinins. Arab. B 2014, 14, e0168. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with Tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Mcsteen, P.; Leyser, O. Shoot Branching. Annu. Rev. Plant Biol. 2005, 56, 353–374. [Google Scholar] [CrossRef]
- Komatsu, K.; Maekawa, M.; Ujiie, S.; Satake, Y.; Furutani, I.; Okamoto, H. LAX and SPA : Major regulators of shoot branching in rice. Proc. Natl. Acad. Sci. USA 2003, 100, 11765–11770. [Google Scholar] [CrossRef]
- Shaaf, S.; Bretani, G.; Biswas, A.; Fontana, I.M.; Rossini, L. Genetics of barley tiller and leaf development. J. Integr. Plant Biol. 2019, 61, 226–256. [Google Scholar] [CrossRef]
- Tukey, J.W. Comparing individual means in the analysis of variance. Biometrics 1949, 5, 99–114. [Google Scholar] [CrossRef] [PubMed]
- Gertsch, J.; Leonti, M.; Raduner, S.; Racz, I.; Chen, J.Z.; Xie, X.Q.; Altmann, K.H.; Karsak, M.; Zimmer, A. β-caryophyllene is a dietary cannabinoid. Proc. Natl. Acad. Sci. USA 2008, 105, 9099–9104. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, A.V.; Ribeiro, B.H.; Salgueiro, L.C.L.; Sato, A. Influence of growth regulators in biomass production and volatile profile of in vitro plantlets of Thymus vulgaris L. J. Agric. Food. Chem. 2009, 57, 6392–6395. [Google Scholar] [CrossRef]
- Umehara, M.; Hanada, A.; Yoshida, S.; Akiyama, K.; Arite, T.; Takeda-Kamiya, N.; Magome, H.; Kamiya, Y.; Shirasu, K.; Yoneyama, K. Inhibition of shoot branching by new terpenoid plant hormones. Nature 2008, 455, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Y.; Haider, I.; Jamil, M.; Fiorilli, V.; Saito, Y.; Mi, J.; Baz, L.; Kountche, B.A.; Jia, K.P.; Guo, X. Growth and strigolactone biosynthesis in rice. Nat. Commun. 2018, 2019, 810. [Google Scholar] [CrossRef]
- Mizukami, H.; Okada, Y.; Ohashi, H. Clonal propagation of Lemon Grass (Cymbopogon citratus Stapf) through shoot tip culture. Plant Tissue Cult. Lett. 1989, 6, 22–24. [Google Scholar] [CrossRef][Green Version]
- Quiala, E.; Barbón, R.; Capote, A.; Pérez-Alonso, N. Scaling-up the biomass production of Cymbopogon citratus L. in temporary immersion system. Biotecnol. Veg. 2014, 14, 67–71. [Google Scholar]
- Licea Moreno, R.J.; Fernandez Moreno, M.; Alvarado Ruffo, K.; Gómez-kosky, R. Influencia de la concentración de agar sobre la multiplicación in vitro de Cymbopogon citratus (D.C.) Stapf. Biotecnología Vegetal. 2001, 1, 77–81. [Google Scholar]
- Ribeiro, A.V.; Ribeiro, B.H.; Santos, L.S.; Apparecida, E.M.; Sato, A. Solid phase microextraction (SPME) analysis of volatile compounds produced by in vitro shoots of Lantana camara L. under the influence of auxins and cytokinins. J. Braz. Chem. Soc. 2007, 18, 1504–1508. [Google Scholar]
- Zalabák, D.; Pospíšilová, H.; Šmehilová, M.; Mrízová, K.; Frébort, I.; Galuszka, P. Genetic engineering of cytokinin metabolism: Prospective way to improve agricultural traits of crop plants. Biotechnol. Adv. 2013, 31, 97–117. [Google Scholar] [CrossRef]
- Kamenicka, A.; Valka, J.; Vzarova, G. A comparative study of different cytokinins on the formation of Rhododendron forrestii Bait’. f. ex Diels. axillary shoots in vitro. Acta Phisiologyae Plant. 1998, 20, 167–171. [Google Scholar] [CrossRef]
- Stojičić, D.; Tošić, S.; Stojanović, G.; Slatković, B.; Jovanović, S.; Budimir, S.; Uzelac, B. Volatile organic compound composition and glandular trichome characteristics of in vitro propagated Clinopodium pulegium (Rochel) Bräuchler: Effect of Carbon Source. Plants 2022, 11, 198. [Google Scholar] [CrossRef] [PubMed]
- Kacem, N.S.; Delporte, F.; Muhovski, Y.; Djekoun, A.; Watillon, B. In vitro screening of durum wheat against water-stress mediated through polyethylene glycol. J. Gen. Engin. Biotech. 2017, 15, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Teyssier, C.; Bonhomme, L.; Lomenech, A.; Vallance, M.; Morabito, D.; Label, P. Increased gelling agent concentration promotes somatic embryo maturation in hybrid larch (Larix × eurolepsis): A 2-DE proteomic analysis. Physiol. Plant. 2011, 141, 152–165. [Google Scholar] [CrossRef]
- Klimaszewska, K.; Bernier-Cardou, M.; Cyr, D.R.; Sutton, B.C.S. Influence of gelling agents on culture medium gel strength, water availability, tissue water potential, and maturation response in embryogenic cultures of Pinus strobus L. Cell Develop. Biol.-Plant. 2000, 36, 279–286. [Google Scholar] [CrossRef]
- Seingre, D.; O’Rourke, J.; Gavillet, S.; Moncousin, C. Influence of gelling agent and carbon source on the in vitro proliferation rate of apple rootstock EM IX. Acta Hortic. 1991, 289, 151–156. [Google Scholar] [CrossRef]
- Letham, D.S.; Gollnow, B.I. Regulators of Cell division in plant tissues.XXX. Cytokinin metabolism in relation to radish cotyledon expansion and senescence. J. Plant Growth Regul. 1985, 4, 129–145. [Google Scholar] [CrossRef]
- Balibrea Lara, M.E.; Garcia Gonzalez, M.-C.; Fatima, T.; Ehness, R.; Lee, T.K.; Proels, R.; Tanner, W.; Roitsch, T. Extracellular invertase is an essential component of cytokinin-mediated delay of senescence. Plant Cell 2004, 16, 1276–1287. [Google Scholar] [CrossRef]
- Albacete, A.; Cantero-Navarro, E.; Großkinsky, D.K.; Arias, C.L.; Balibrea, M.E.; Bru, R.; Fragner, L.; Ghanem, M.E.; González, M.d.L.C.; Hernández, J.A.; et al. Ectopic overexpression of the cell wall invertase gene CIN1 leads to dehydration avoidance in tomato. J. Exp. Bot. 2015, 66, 863–878. [Google Scholar] [CrossRef]
- Barbier, F.F.; Dun, E.A.; Kerr, S.C.; Chabikwa, T.G.; Beveridge, C.A. An update on the signals controlling shoot branching. Trends Plant Sci. 2019, 24, 220–236. [Google Scholar] [CrossRef]
- Wang, M.; Gourrierec, L.; Jiao, F.; Demotes-Mainard, S.; Perez-Garcia, M.D.; Ogé, L.; Hamama, L.; Crespel, L.; Bertheloot, J.; Chen, J.; et al. Convergence and divergence of sugar and cytokinin signaling in plant development. Int. J. Mol. Sci. 2021, 1, 1282. [Google Scholar] [CrossRef] [PubMed]
- Trang, D.T.; Van, H.T.K.; Minh, N.T.T.; Van, C.P.; Dang, N.H.; Dang, H.D.; Quang, T.N.; Tien, D.N. Essential oils of lemongrass (Cymbopogon citratus Stapf) induces apoptosis and cell cyclle arrst in A549 lung cancer cells. BioMed. Res. Int. 2020, 2020, 5925856. [Google Scholar] [CrossRef] [PubMed]
- Kim, V.H.T.; Minh, Q.N.; Vinh, H.D.T.; Hai, N.T.; Ly, H.T.; Hien, N.T.; Trang, D.T.; Dang, N.H.; Dat, N.T. Chemical composition and cytotoxic activity of the essential oils of Cymbopogon citratus L. grown in Phu Tho province. J. Biotech. 2016, 14, 683–687. [Google Scholar]
- Stolz, A.; Riefler, M.; Lomin, S.N.; Achazi, K.; Romanov, G.A.; Schmülling, T. The specificity of cytokinin signalling in Arabidopsis thaliana is mediated by differing ligand affinities and expression profiles of the receptors. Plant J. 2011, 67, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, M.; Pischke, M.S.; Mahonen, A.P.; Miyawaki, K.; Hashimoto, Y.; Seki, M.; Kobayashi, M.; Shinozaki, K.; Kato, T.; Tabata, S.; et al. In planta functions of the Arabidopsis cytokinin. Proc. Natl. Acad. Sci. USA 2004, 101, 8821–8826. [Google Scholar] [CrossRef] [PubMed]
- Hare, B.; van Staden, J. Minireview Cytokinin oxidase : Biochemical features and physiological significance. Physiol. Plant. 1994, 91, 128–136. [Google Scholar] [CrossRef]
- Werner, T.; Motika, V.; Strnad, M.S.T. Regulation of plant growth by cytokinin. Proc. Nat. Acad. Sci. USA 2001, 98, 10487–10492. [Google Scholar] [CrossRef]
- Werner, T.; Motyka, V.; Laucou, V.; Smets, R.; Van Onckelen, H. Cytokinin-deficient transgenic arabidopsis plants show functions of cytokinins in the regulation of shoot and root meristem activity. Plant Cell. 2003, 15, 2532–2550. [Google Scholar] [CrossRef]
- Werner, T.; Köllmer, I.; Bartrina, I.; Holst, K.; Schmülling, T. New insights into the biology of cytokinin degradation. Plant Biol. 2006, 8, 371–381. [Google Scholar] [CrossRef]
- Galuszka, P.; Popelkova, H.; Werner, T.; Frebortova, J.; Pospisilova, H.; Vaclav, M.; Kollmer, I.; Schmülling, I.F. Biochemical characterization of Cytokinin Oxidases / Dehydrogenases from Arabidopsis thaliana expressed in Nicotiana tabacum L. J. Plant Growth Regul. 2007, 26, 255–267. [Google Scholar] [CrossRef]
- Cárdenas-Aquino, M.R.; Sarria-Guzmán, Y.; Martínez-Antonio, A. Review: Isoprenoid and aromatic cytokinins in shoot branching. Plant Sci. 2022, 319, 111240. [Google Scholar] [CrossRef] [PubMed]
- Papon, N.; Bremer, J.; Rideau, M. Cytokinin and ethylene control indole alkaloid production at the level of the MEP/Terpenoid pathway in Catharanthus roseus suspension cells. Planta Med. 2005, 71, 572–574. [Google Scholar] [CrossRef] [PubMed]
- Phatak, S.V.; Heble, M.R. Organogenesis and terpenoid synthesis in Mentha arvensis. Fitoterapia 2002, 73, 32–39. [Google Scholar] [CrossRef]
- Danova, K.; Todorova, M.; Trendafilova, A.; Evstatieva, L. Cytokinin and auxin effect on the terpenoid profile of the essential oil and morphological characteristics of shoot cultures of Artimisia alba. Nat. Prod. Commun. 2012, 7, 1075–1076. [Google Scholar] [PubMed]
- Krumova, S.; Motyka, V.; Dobrev, P.; Trendafilova, A.; Evstatieva, L.; Danova, K. Terpenoid profile of Artemisia alba is related to endogenous cytokinins in vitro. Bulg. J. Agric. Sci. 2013, 19 (Suppl. 2), 26–30. [Google Scholar]
- Danova, K.; Motyka, V.; Todorova, M.; Trendafilova, A.; Krumova, S.; Dobrev, P.; Andreeva, T.; Oreshkova, T.; Taneva, S.; Evstatieva, L. Effect of cytokinin and auxin treatments on morphogenesis, terpenoid biosynthesis, photosystem structural organization, and endogenous isoprenoid cytokinin profile in Artemisia alba Turra in vitro. J. Plant Growth Regul. 2018, 37, 403–418. [Google Scholar] [CrossRef]
- Wanke, M.; Skorupinska-Tudek, K.; Swiezewska, E. Isoprenoid biosynthesis via 1-deoxy-D-xylulose5-phosphate/2-C-methyl-erythritol 4-phosphate (DOXP/MEP) pathway. Acta Biochim. Pol. 2001, 48, 663–672. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Non-mevalonate isoprenoide biosynthesis: Enzymes, genes, and inhibitors. Biochem. Soc. Trans. 2000, 28, 785–789. [Google Scholar] [CrossRef]
- Silva, G.D.S.E.; Jesus Márques, J.N.; Moreira Linhares, E.P.; Martinez Bonora, C.; Tosoni Costa, E.; Frota, S.M. Review of anticancer activity of monoterpenoids: Geraniol, nerol, geranial and neral. Chem. Biol. Interact. 2022, 1, 109994. [Google Scholar] [CrossRef]
- Yu, S.G.; Hildebrandt, L.A.; Elson, C.E. Geraniol, an ihibitor of mevalonate biosynthesis, suppresses the growth of hepatomas and melanomas trasplanted to rats and mice. J. Nutr. 1995, 125, 2763–2767. [Google Scholar]
- Pattanayak, M.; Seth, P.K.; Smita, S.; Gupta, S.K. Geraniol and limonene interaction with 3-hydroxy-3-methylglutaryl-CoA (HMG-CoA) reductase for their role as cancer chemo-preventive agents. J. Proteomics Bioinform. 2009, 2, 466–474. [Google Scholar] [CrossRef]
- Izumi, S.; Takashima, O.; Hirata, T. Geraniol is a potent inducer of apoptosis-like cell death in the cultured shoot primordia of Matricaria chamomilla. Biochem. Biophys. Res. Comm. 1999, 259, 519–522. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Kumar, S.U.; Meli, V.S.; Kumar, V.M.; Kumar, A.; Irfan, M.; Chacraborty, N.; Chakraborty, S.; Datta, A. Induction of senescence and identification of differentially expressed genes in tomato in response to monoterpene. PLoS ONE 2013, 8, e76029. [Google Scholar] [CrossRef] [PubMed]
- Chaimovitsh, D.; Abu-Abied, M.; Belausov, E.; Rubin, B.; Dudai, N.; Sadot, E. Microtubules are an intracellular target of the plant terpene citral. Plant J. 2010, 61, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, A.J.; Lehner, K.; Mi, J.; Jia, K.; Mijar, M.; Dinneny, J. β -Cyclocitral is a conserved root growth regulator. Proc. Nat. Acad. Sci. USA 2019, 116, 10563–10567. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).