
Isolation and Molecular Identification of Serratia Strains Producing Chitinases, Glucanases, Cellulases, and Prodigiosin and Determination of Their Antifungal Effect against Colletotrichum siamense and Alternaria alternata In Vitro and on Mango Fruit

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Identification of Mango Phytopathogenic Fungi
2.1.1. Isolation of Fungi from Mangoes with Anthracnose
2.1.2. Verification of the Pathogenicity of the Isolated Fungal Strains
Preparation of the Mangoes
Preparation of the Fungal Inoculum
Inoculation of Mangoes and Detection of Anthracnose Signs
2.1.3. Molecular Identification of the Mid Fungal Strain
2.2. Isolation and Identification of Bacteria with Antifungal Activity
2.2.1. Bacterial Isolation
2.2.2. In Vitro Antifungal Effect of Isolated Bacteria
2.2.3. Identification of Bacterial Strains with Antifungal Activity
2.3. Antifungal Effect of S. marcescens and S. nematodiphila Strains against the Conidia of Colletotrichum and Alternaria
2.4. Chitinolytic Activity Assay
2.4.1. Preparation of Colloidal Chitin
2.4.2. Chitinolytic Activity
2.5. Glucanolytic Activity
2.6. Cellulolytic Activity
2.7. Prodigiosin Production
2.8. Antagonistic Effects of S. marcescens and S. nematodiphila against C. siamense and A. alternata on Mango
2.9. Hemolytic Activity of Serratia Marcescens Strains
2.10. Statistical Analyses
3. Results
3.1. Isolation and Identification of Mango Phytopathogenic Fungi
3.2. Antifungal Activity of Isolated Bacteria
Identification of Bacterial Strains with Antifungal Activity
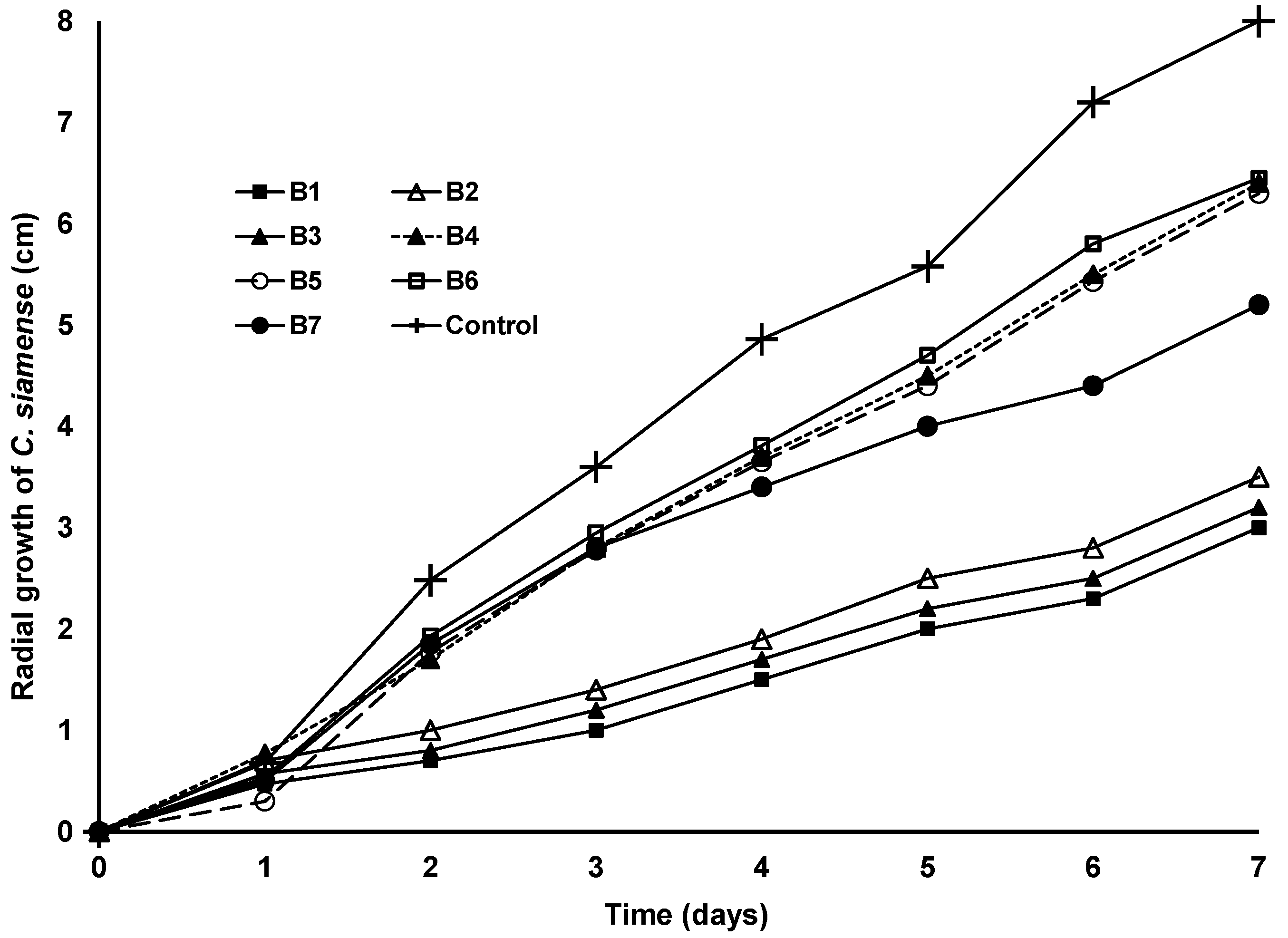
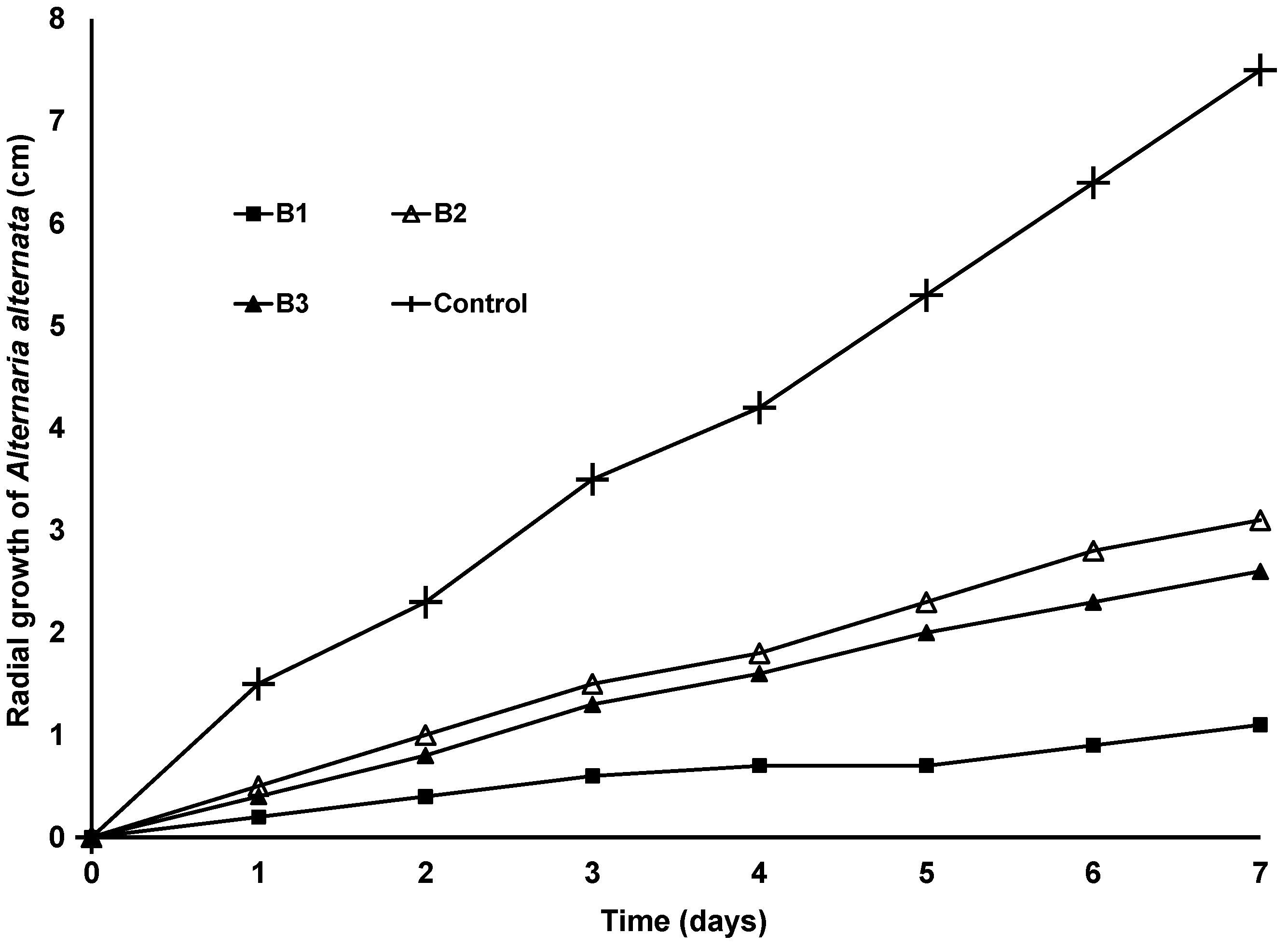
3.3. Antifungal Effect of S. marcescens and S. Nematodiphila Strains against the Conidia of Colletotrichum and Alternaria
3.4. Chitinolytic, Glucanolytic and Cellulolytic Activity
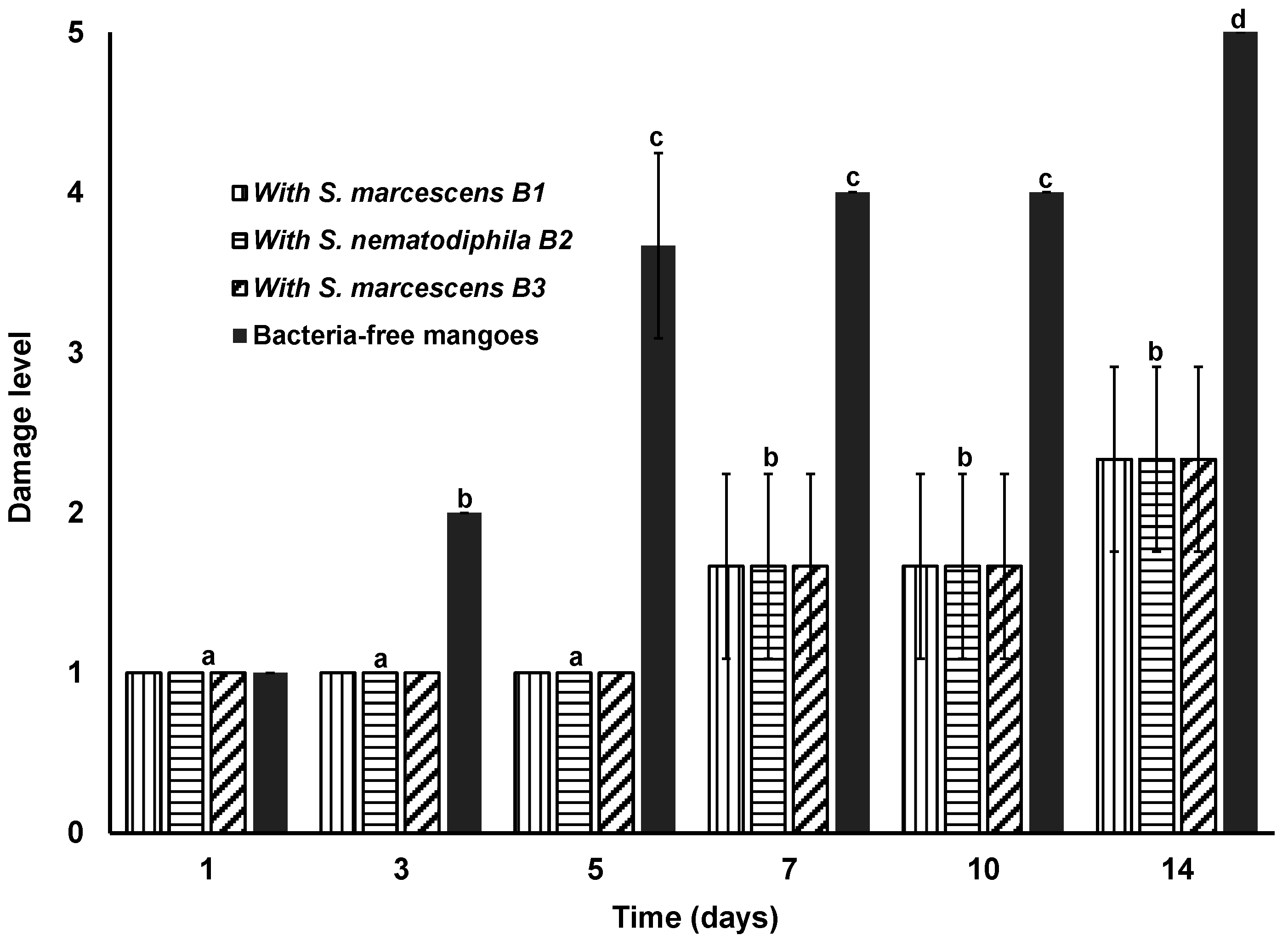
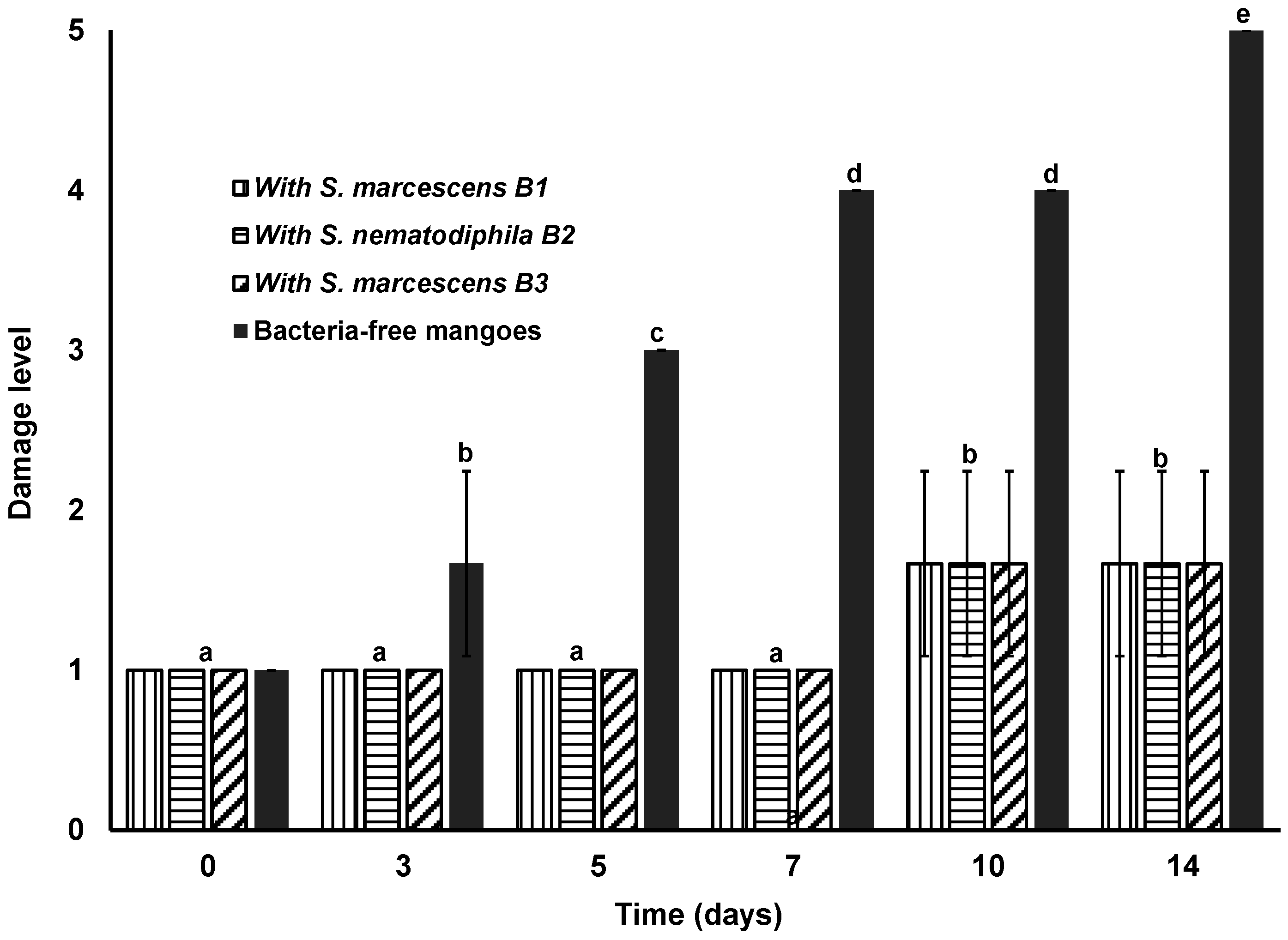
3.5. Antagonistic Effects of S. marcescens and S. nematodiphila on Mango
3.6. Hemolytic Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
- Sequence A1. Nucleotide sequence of the identified Colletotrichum siamense. According to the information contained in the BLAST of the National Center for Biotechnology Information (NCBI), the sequence obtained from the MID strain corresponded to Colletotrichum siamense. The most similar sequence corresponded to C. siamense strain HLX5, with an accession number of MN860116.1, with 100% sequence coverage and 99% sequence identity.
- ATTACTGAGTTTACGCTCTACAACCCTTTGTGAACATACCTATAACTGTTGCTTCGGCGGGTAGGGTCTCCGCGACCCTCCCGGCCTCCCGCCTCCGGGCGGGTCGGCGCCCGCCGGAGGATAACCAAACTCTGATTTAACGACGTTTCTTCTGAGTGGTACAAGCAAATAATCAAAACTTTTAACAACGGATCTCTTGGTTCTGGCATCGATGAAGAACGCAGCGAAATGCGATAAGTAATGTGAATTGCAGAATTCAGTGAATCATCGAATCTTTGAACGCACATTGCGCCCGCCAGCATTCTGGCGGGCATGCCTGTTCGAGCGTCATTTCAACCCTCAAGCTCTGCTTGGTGTTGGGGCCCTACAGCTGATGTAGGCCCTCAAAGGTAGTGGCGGACCCTCTCGGAGCCTCCTTTGCGTAGTAACTTTACGTCTCGCACTGGGATCCGGAGGGACTCTTGCCGTAAAACCCCCCAATTTTCCAAAGGTTGACCTCGGATCAGGT.
- Sequence A2. Nucleotide sequence of the identified Serratia marcescens. The sequence most similar to the B1 strain, according to the BLAST search of the National Center for Biotechnology Information (NCBI), was with an accession number of CP053572.1 with 100% sequence coverage and 99% identity (sequence of the 16S fragment of strain B1 with a total of 1453 nucleotides).
- TGGCGGCAGGCCTAACACATGCAAGTCGAGCGGTAGCACAGGGGAGCTTGCTCCCTGGGTGACGAGCGGCGGACGGGTGAGTAATGTCTGGGAAACTGCCTGATGGAGGGGGATAACTACTGGAAACGGTAGCTAATACCGCATAACGTCGCAAGACCAGAGGGGGACCTTCGGGCCTCTTGCCATCAGATGTGCCCAGATGGGATTAGTGTGGCTCACCTAGGCGACGATCCCTAGCTGGTCTGAGAGGATGACCAGCCACACTGGAACTGAGACACGGTCCAGACTCCTACGGGAGGCAGCAGTGGGGAATATTGCACAATGGGCGCAAGCCTGATGCAGCCATGCCGCGTGTGTGAAGAAGGCCTTCGGGTTGTAAAGCACTTTCAGCGAGGAGGAAGGTGGTGAACTTAATACGTTCATCAATTGACGTTACTCGCAGAAGAAGCACCGGCTAACTCCGTGCCAGCAGCCGCGGTAATACGGAGGGTGCAAGCGTTAATCGGAATTACTGGGCGTAAAGCGCACGCAGGCGGTTTGTTAAGTCAGATGTGAAATCCCCGGGCTCAACCTGGGAACTGCATTTGAAACTGGCAAGCTAGAGTCTCGTAGAGGGGGGTAGAATTCCAGGTGTAGCGGTGAAATGCGTAGAGATCTGGAGGAATACCGGTGGCGAAGGCGGCCCCCTGGACGAAGACTGACGCTCAGGTGCGAAAGCGTGGGGAGCAAACAGGATTAGATACCCTGGTAGTCCACGCTGTAAACGATGTCGATTTGGAGGTTGTGCCCTTGAGGCGTGGCTTCCGGAGCTAACGCGTTAAATCGACCGCCTGG.GGAGTACGGCCGCAAGGTTAAAACTCAAATGAATTGACGGGGGCCCGCACAAGCGGTGGAGCATGTGGTTTAATTCGATGCAACGCGAAGAACCTTACCTACTCTTGACATCCAGAGAACTTAGCAGAGATGCTTTGGTGCCTTCGGGAACTCTGAGACAGGTGCTGCATGGCTGTCGTCAGCTCGTGTTGTGAAATGTTGGGTTAAGTCCCGCAACGAGCGCAACCCTTATCCTTTGTTGCCAGCGGTTCGGCCGGGAACTCAAAGGAGACTGCCAGTGATAAACTGGAGGAAGGTGGGGATGACGTCAAGTCATCATGGCCCTTACGAGTAGGGCTACACACGTGCTACAATGGCATATACAAAGAGAAGCGACCTCGCGAGAGCAAGCGGACCTCATAAAGTATGTCGTAGTCCGGATTGGAGTCTGCAACTCGACTCCATGAAGTCGGAATCGCTAGTAATCGTAGATCAGAATGCTACGGTGAATACGTTCCCGGGCCTTGTACACACCGCCCGTCACACCATGGGAGTGGGTTGCAAAAGAAGTAGGTAGCTTAACCTTCGGGAGGGCGCTTACCACTTTGTGATTCATGACTGGGG.
- Sequence A3. Nucleotide sequence of the identified Serratia nematodiphila. In the case of strain B2, the most similar sequence according to the BLAST search corresponded to Serratia nematodiphila, with an accession number of KP318499.1 with 100% sequence coverage and 99% identity.
- AGMTTGYTSCCCGGGTGACGAGCGSCGGACGGGTGAGYAATGTCTAGGRAAACTGCCTGAAGCTTGCTCCCCGGGTGACGAGCGGCGGACGGGTGAGTAATGTCTGGGAAACTGCCTGATGGAGRGGGATAACTACTGGAAACGGTAGCTAATACCGCATAACGTCGCAAGACCAAAGATGGAGGGGGATAACTACTGGAAACGGTAGCTAATACCGCATAACGTCGCAAGACCAAAGAGGGGGACCTTCGGGCCTCTTGCCATCAGATGTGCCCAGATGGGATTAGCTAGTAGGTGGGGGGGGACCTTCGGGCCTCTTGCCATCAGATGTGCCCAGATGGGATTAGCTAGTAGGTGGGGTAATGGCTCACCTAGGCGACGATCCCTAGCTGGTCTGAGAGGATGACCAGCCACACTGGGTAATGGCTCACCTAGGCGACGATCCCTAGCTGGTCTGAGAGGATGACCAGCCACACTGGAACTGAGACACGGTCCAGACTCCTACGGGAGGCAGCAGTGGGGAATATTGCACAATGGGCAACTGAGACACGGTCCAGACTCCTACGGGAGGCAGCAGTGGGGAATATTGCACAATGGGCGCAAGCCTGATGCAGCCATGCCGCGTGTGTGAAGAAGGCCTTCGGGTTGTAAAGCACTTTGCAAGCCTGATGCAGCCATGCCGCGTGTGTGAAGAAGGCCTTCGGGTTGTAAAGCACTTTCAGCGAGGAGGAAGGTGGTGAGCTTAATACGYTCATCAATTGACGTTACTCGCAGAAGAACAGCGAGGAGGAAGGTGGTGAGCTTAATACGTTCATCAATTGACGTTACTCGCAGAAGAAGCACCGGCTAACTCCGTGCCAGCAGCCGCGGTAATACGGAGGGTGCAAGCGTTAATCGGAGCACCGGCTAACTCCGTGCCAGCAGCCGCGGTAATACGGAGGGTGCAAGCGTTAATCGGAATTACTGGGCGTAAAGCGCACGCAGGCGGTTTGTTAAGTCAGATGTGAAATCCCCGGGCTATTACTGGGCGTAAAGCGCACGCAGGCGGTTTGTTAAGTCAGATGTGAAATCCCCGGGCTCAACCTGGGAACTGCATTTGAAACTGGCAAGCTAGAGTCTCGTAGAGGGGGGTAGAATTCCAACCTGGGAACTGCATTTGAAACTGGCAAGCTAGAGTCTCGTAGAGGGGGGTAGAATTCCAGGTGTAGCGGTGAAATGCGTAGAGATCTGGAGGAATACCGGTGGCGAAGGCGGCCCCCCAGGTGTAGCGGTGAAATGCGTAGAGATCTGGAGGAATACCGGTGGCGAAGGCGGCCCCCTGGACGAAGACTGACGCTCAGGTGCGAAAGCGTGGGGAGCAAACAGGATTAGATACCCTGTGGACGAAGACTGACGCTCAGGTGCGAAAGCGTGGGGAGCAAACAGGATTAGATACCCTGGTAGTCCACGCTGTAAACGATGTCGATTTGGAGGTTGTGCCCTTGAGGCGTGGCTTCCGGGTAGTCCACGCTGTAAACGATGTCGATTTGGAGGTTGTGCCCTTGAGGCGTGGCTTCCGGAGCTAACGCGTTAAATCGACCGCCTGGGGAGTACGGCCGCAAGGTTAAAACTCAAATGAAAGCTAACGCGTTAAATCGACCGCCTGGGGAGTACGGCCGCAAGGTTAAAACTCAAATGAATTGACGGGGGCCCGCACAAGCGGTGGAGCATGTGGTTTAATTCGATGCAACGCGAAGAACTTGACGGGGGCCCGCACAAGCGGTGGAGCATGTGGTTTAATTCGATGCAACGCGAAGAACCTTACCTACTCTTGACATCCAGAGAACTTWCCAGAGATGCWTTGGTGCCTTCGGGAACTCCTTACCTACTCTTGACATCCAGAGAACTTTCCAGAGATGCATTGGTGCCTTCGGGAACTCTGAGACAGGTGCTSCATGGCTGTCGTCAGCTCGTGTTGTGAAATGTTGGGGTTAAGTCCCTGAGACAGGTGCTGCATGGCTGTCGTCAGCTCGTGTTGTGAAATGTTGGGTTAAGTCCCGCAACGAGCGCAACCATATCCTTTGCTGCCAGCGGTCCGTCGGACTCAAGGAGACGCAACGAGCGCAACCCTTATCCTTTGTTGCCAGCGGTTCGGCCGGGAACTCAAAGGAGACTGCCAGTGATAAACTGGAGGAAG.
- Sequence A4. Nucleotide sequence of the identified Serratia marcescens. In the case of strain B3, the most similar sequence according to the BLAST search corresponded to S. marcescens strain S14, with an accession number of MK346258.1 with 100% sequence coverage and 99% identity.
- TGGGTGACGTKYGGGGACGATGAGCAACGTCAGGAAAACTGCCTGATGGAGGGGGATATGGGTGACGAGCGGCGGACGGGTGAGTAATGTCTGGGAAACTGCCTGATGGAGGGGGATAACTACTGGAAACGGTAGCTAATACCGCATAACGTCGCAAGACCAAAGAGGGGGACCTTCGACTACTGGAAACGGTAGCTAATACCGCATAACGTCGCAAGACCAAAGAGGGGGACCTTCGGGCCTCTTGCCATCAGATGTGCCCAGATGGGATTAGCTAGTAGGTGGGGTAATGGCTCACGGCCTCTTGCCATCAGATGTGCCCAGATGGGATTAGCTAGTAGGTGGGGTAATGGCTCACCTAGGCGACGATCCCTAGCTGGKCTCTAGGCGACGATCCCTAGCTGGTCTGAGAGGATGACCAGCCACACTGGAACTGAGACACGTCCMGAMTYCYTACGGGARGSAGCRSKGGGRAWATTGCACAWGGGSGCAGCCTGATGTCCAGACTCCTACGGGAGGCAGCAGTGGGGAATATTGCACAATGGGCGCAAGCCTGATGCAKCMTGSCGCGKGKGTGAAGAGGCTTCSGGTGCAGCCATGCCGCGTGTGTGAAGAAGGCCTTCGGGT.
References
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [PubMed]
- Awang, Y.; Ghani, M.A.A.; Sijam, K.; Mohamad, R.B. Effect of calcium chloride on anthracnose disease and postharvest quality of red-flesh dragon fruit (Hylocereus polyrhizus). Afr. J. Microbiol. Res. 2011, 5, 5250–5259. [Google Scholar] [CrossRef]
- Siddiqui, Y.; Asgar, A. Colletotrichum gloeosporioides (Anthracnose). In Postharvest Decay: Control Strategies; Chapter 11; Bautista-Baños, S., Ed.; Academic Press: Cambridge, MA, USA; Elsevier Inc.: London, UK, 2014; pp. 337–371. [Google Scholar] [CrossRef]
- Díaz-Medina, A.R.; Arboleda-Zapata, T.; Ríos-Osorio, L.A. Biological control strategies used for the management of antracnosis caused by Colletotrichum gloeosporioides in mango fruits: A systematic review. Trop. Subtrop. Agroecosyst. 2019, 22, 595–611. [Google Scholar]
- Tovar-Pedraza, J.M.; Mora-Aguilera, J.A.; Nava-Díaz, C.; Lima, N.B.; Michereff, S.J.; Sandoval-Islas, J.S.; Leyva-Mir, S.G. Distribution and pathogenicity of Colletotrichum species associated with mango anthracnose in Mexico. Plant Dis. 2020, 104, 137–146. [Google Scholar] [CrossRef]
- Köhler, H.R.; Triebskorn, R. Wildlife ecotoxicology of pesticides: Can we track effects to the population level and beyond? Science 2013, 341, 759–765. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Y.; Zhang, M. Spatial and temporal patterns of pesticide use on California almonds and associated risks to the surrounding environment. Sci. Total Environ. 2014, 472, 517–529. [Google Scholar] [CrossRef] [PubMed]
- Touré, Y.; Ongena, M.; Jacques, P.; Guiro, A.; Thonart, P. Role of lipopeptides produced by Bacillus subtilis GA1 in the reduction of grey mould disease caused by Botrytis cinerea on apple. J. Appl. Microbiol. 2004, 96, 1151–1160. [Google Scholar] [CrossRef]
- Kefialew, Y.; Ayalew, A. Postharvest biological control of anthracnose (Colletotrichum gloeosporioides) on mango (Mangifera indica). Postharvest Biol. Technol. 2008, 50, 8–11. [Google Scholar] [CrossRef]
- Guo, Z.; Zhang, X.; Wu, J.; Yu, J.; Xu, M.; Chen, D.; Zhang, Z.; Li, X.; Chi, Y.; Wan, S. In vitro inhibitory effect of the bacterium Serratia marcescens on Fusarium proliferatum growth and fumonisins production. Biol. Control 2020, 19, 202–219. [Google Scholar] [CrossRef]
- Zarei, M.; Aminzadeh, S.; Zolgharnein, H.; Safahieh, A.; Ghoroghi, A.; Motallebi, A.; Daliri, M.; Sahebghadam-Lotfi, A. Serratia marcescens B4A chitinase product optimization using Taguchi approach. Iran. J. Biotechnol. 2010, 8, 252–262. [Google Scholar]
- Fantinel, V.S.; Muniz, M.F.B.; Poletto, T.; Dutra, A.F.; Krahn, J.T.; Favaretto, R.F.; Sarzi, J.S. Biocontrole in vitro de Colletotrichum siamense utilizando Trichoderma spp. e Bacillus thuringiensis var. kurstaki. Rev. Ciência Agríc. 2018, 16, 43–50. [Google Scholar] [CrossRef]
- Xu, J.X.; Li, Z.Y.; Lv, X.; Yan, H.; Zhou, G.Y.; Cao, L.X.; He, Y.H. Isolation and characterization of Bacillus subtilis strain 1-L-29, an endophytic bacteria from Camellia oleifera with antimicrobial activity and efficient plant-root colonization. PLoS ONE 2020, 15, e0232096. [Google Scholar] [CrossRef] [PubMed]
- Živković, S.; Stevanović, M.; Đurović, S.; Ristić, D.; Stošić, S. Antifungal activity of chitosan against Alternaria alternata and Colletotrichum gloeosporioides. Pestic. Phytomed. 2018, 33, 197–204. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press Inc.: New York, NY, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Saha, D.; Purkayastha, G.D.; Ghosh, A.; Isha, M.; Saha, A. Isolation and characterization of two new Bacillus subtilis strains from rhizosphere of eggplant as potential biocontrol agents. J. Plant Pathol. 2012, 94, 109–127. [Google Scholar] [CrossRef]
- Gutiérrez-Román, M.I.; Holguín-Meléndez, F.; Bello-Mendoza, R.; Guillén-Navarro, K.; Dunn, M.F.; Huerta-Palacios, G. Production of prodigiosin and chitinases by tropical Serratia marcescens strains with potential to control plant pathogens. World J. Microbiol. Biotechnol. 2012, 28, 145–153. [Google Scholar] [CrossRef]
- Renwick, A.; Campbell, R.; Coe, S. Assessment of in vivo screening systems for potential biocontrol agents of Gaeumannomyces graminis. Plant Pathol. 1991, 40, 524–532. [Google Scholar] [CrossRef]
- Ashwini, N.; Srividya, S. Potentiality of Bacillus subtilis as biocontrol agent for management of anthracnose disease of chilli caused by Colletotrichum gloeosporioides OGC1. 3 Biotech 2014, 4, 127–136. [Google Scholar] [CrossRef]
- Slater, H.; Crow, M.; Everson, L.; Salmond, G.P. Phosphate availability regulates biosynthesis of two antibiotics, prodigiosin and carbapenem, in Serratia via both quorum-sensing-dependent and-independent pathways. Mol. Microbiol. 2003, 47, 303–320. [Google Scholar] [CrossRef]
- Corkidi, G.; Balderas-Ruíz, K.A.; Taboada, B.; Serrano-Carreón, L.; Galindo, E. Assessing mango anthracnose using a new three-dimensional image-analysis technique to quantify lesions on fruit. Plant Pathol. 2006, 55, 250–257. [Google Scholar] [CrossRef]
- Gerhardt, P.; Murray, R.; Wood, W.; Krieg, N. Methods for General and Molecular Bacteriology; American Society of Microbiology: Washington, DC, USA, 1994; pp. 21–42. [Google Scholar]
- Lima, N.B.; Batista, M.V.D.A.; De Morais, M.A.; Barbosa, M.A.G.; Michereff, S.J.; Hyde, K.D.; Câmara, M.P.S. Five Colletotrichum species are responsible for mango anthracnose in northeastern Brazil. Fungal Divers. 2013, 61, 75–88. [Google Scholar] [CrossRef]
- Sharma, G.; Kumar, N.; Weir, B.S.; Hyde, K.D.; Shenoy, B.D. The ApMat marker can resolve Colletotrichum species: A case study with Mangifera indica. Fungal Divers. 2013, 61, 117–138. [Google Scholar] [CrossRef]
- Du, W.; Yao, Z.; Li, J.; Sun, C.; Xia, J.; Wang, B.; Ren, L. Diversity and antimicrobial activity of endophytic fungi isolated from Securinega suffruticosa in the Yellow River Delta. PLoS ONE 2020, 15, e0229589. [Google Scholar] [CrossRef]
- Lapenda, J.C.; Silva, P.A.; Vicalvi, M.C.; Sena, K.X.F.R.; Nascimiento, S.C. Antimicrobial activity of prodigiosin isolated from Serratia marcescens UFPEDA. World J. Microbiol. Biotechnol. 2015, 31, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Basharat, Z.; Tanveer, F.; Yasmin, A.; Shinwari, Z.K.; He, T.; Tong, Y. Genome of Serratia nematodiphila MB307 offers unique insights into its diverse traits. Genome 2018, 61, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Han, K.I.; Patnaik, B.B.; Cho, A.R.; Lim, H.K.; Lee, J.M.; Jang, Y.G.; Jeong, Y.S.; Yoo, T.K.; Lee, G.S.; Han, M.D. Characterization of chitinase-producing Serratia and Bacillus strains isolated from insects. Entomol. Res. 2014, 44, 109–120. [Google Scholar] [CrossRef]
- Veliz, E.A.; Martínez-Hidalgo, P.; Hirsch, A.M. Chitinase-producing bacteria and their role in biocontrol. AIMS Microbiol. 2017, 3, 689–705. [Google Scholar] [CrossRef] [PubMed]
- Bowman, S.M.; Free, S.J. The structure and synthesis of the fungal cell wall. Bioessays 2006, 28, 799–808. [Google Scholar] [CrossRef]
- Kalbe, C.; Marten, P.; Berg, G. Strains of the genus Serratia as beneficial rhizobacteria of oilseed rape with antifungal properties. Microbiol. Res. 1996, 151, 433–439. [Google Scholar] [CrossRef]
- Sethi, S.; Datta, A.; Gupta, B.L.; Gupta, S. Optimization of cellulase production from bacteria isolated from soil. Int. Sch. Res. Notices 2013, 2013, 985685. [Google Scholar] [CrossRef]
- Darshan, N.; Manonmani, H.K. Prodigiosin and its potential applications. J. Food Sci. Technol. 2015, 52, 5393–5407. [Google Scholar] [CrossRef] [PubMed]
- Manas, N.H.A.; Yee, C.L.; Tesfamariam, Y.M.; Zulkharnain, A.; Mahmud, H.; Mahmod, D.S.A.; Fuzi, S.F.Z.M.; Azelee, N.I.W. Effects of oil substrate supplementation on production of prodigiosin by Serratia nematodiphila for dye-sensitized solar cell. J. Biotechnol. 2020, 317, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Grimont, P.A.; Grimont, F.; Starr, M.P. Serratia species isolated from plants. Curr. Microbiol. 1981, 5, 317–322. [Google Scholar] [CrossRef]
- Zarei, M.; Aminzadeh, S.; Zolgharnein, H.; Safahieh, A.; Daliri, M.; Noghabi, A.; Ghoroghi, A.; Motallebi, A. Characterization of a chitinase with antifungal activity from a native Serratia marcescens B4A. Braz. J. Microbiol. 2011, 42, 1017–1029. [Google Scholar] [CrossRef]
- Kurz, C.L.; Chauvet, S.; Andres, E.; Aurouze, M.; Vallet, I.; Michel, G.P.; Uh, M.; Celli, J.; Filloux, A.; De Bentzmann, S.; et al. Virulence factors of the human opportunistic pathogen Serratia marcescens identified by in vivo screening. EMBO J. 2003, 22, 1451–1724. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (Hours) | Percentage (%) of Conidia Germination of C. siamense and A. alternata | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Cont1C | Cont2C | Cont1A | Cont2A | C + SmB1 | C + SmB3 | C + SnB2 | A + SmB1 | A + SmB3 | A + SnB2 | |
| 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 6 | 41 ± 6 | 0 | 34 ± 6 | 0 | 4 ± 2 | 6 ± 2 | 4 ± 1 | 3 ± 2 | 4 ± 2 | 4 ± 1 |
| 12 | 85 ± 6 | 0 | 75 ± 6 | 0 | 13 ± 3 | 16 ± 5 | 16 ± 4 | 10 ± 2 | 13 ± 2 | 15 ± 3 |
| 18 | 93 ± 5 | 0 | 87 ± 5 | 0 | 15 ± 4 | 17 ± 4 | 18 ± 3 | 14 ± 3 | 16 ± 3 | 17 ± 4 |
| 24 | 96 ± 3 | 0 | 93 ± 4 | 0 | 16 ± 5 | 17 ± 3 | 19 ± 3 | 15 ± 3 | 16 ± 3 | 16 ± 3 |
| Strain | Chitinolytic Activity (mm) | Glucanolytic Activity | Cellulolytic Activity | Prodigiosin Production |
|---|---|---|---|---|
| S. marcescens B1 | (28 ± 2) 1 | + | + | 2.505 ± 0.062 * |
| S. marcescens B3 | (23 ± 2) | + | + | 2.123 ± 0.100 |
| S. nematodiphila B2 | (24 ± 1) | + | + | 1.883 ± 0.099 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trejo-López, J.A.; Rangel-Vargas, E.; Gómez-Aldapa, C.A.; Villagómez-Ibarra, J.R.; Falfán-Cortes, R.N.; Acevedo-Sandoval, O.A.; Castro-Rosas, J. Isolation and Molecular Identification of Serratia Strains Producing Chitinases, Glucanases, Cellulases, and Prodigiosin and Determination of Their Antifungal Effect against Colletotrichum siamense and Alternaria alternata In Vitro and on Mango Fruit. Int. J. Plant Biol. 2022, 13, 281-297. https://doi.org/10.3390/ijpb13030024
Trejo-López JA, Rangel-Vargas E, Gómez-Aldapa CA, Villagómez-Ibarra JR, Falfán-Cortes RN, Acevedo-Sandoval OA, Castro-Rosas J. Isolation and Molecular Identification of Serratia Strains Producing Chitinases, Glucanases, Cellulases, and Prodigiosin and Determination of Their Antifungal Effect against Colletotrichum siamense and Alternaria alternata In Vitro and on Mango Fruit. International Journal of Plant Biology. 2022; 13(3):281-297. https://doi.org/10.3390/ijpb13030024
Chicago/Turabian StyleTrejo-López, J. Alexander, Esmeralda Rangel-Vargas, Carlos A. Gómez-Aldapa, José R. Villagómez-Ibarra, Reyna N. Falfán-Cortes, Otilio A. Acevedo-Sandoval, and Javier Castro-Rosas. 2022. "Isolation and Molecular Identification of Serratia Strains Producing Chitinases, Glucanases, Cellulases, and Prodigiosin and Determination of Their Antifungal Effect against Colletotrichum siamense and Alternaria alternata In Vitro and on Mango Fruit" International Journal of Plant Biology 13, no. 3: 281-297. https://doi.org/10.3390/ijpb13030024
APA StyleTrejo-López, J. A., Rangel-Vargas, E., Gómez-Aldapa, C. A., Villagómez-Ibarra, J. R., Falfán-Cortes, R. N., Acevedo-Sandoval, O. A., & Castro-Rosas, J. (2022). Isolation and Molecular Identification of Serratia Strains Producing Chitinases, Glucanases, Cellulases, and Prodigiosin and Determination of Their Antifungal Effect against Colletotrichum siamense and Alternaria alternata In Vitro and on Mango Fruit. International Journal of Plant Biology, 13(3), 281-297. https://doi.org/10.3390/ijpb13030024