Antiprotozoal Activities of Organic Extracts from French Marine Seaweeds

Abstract
:1. Introduction
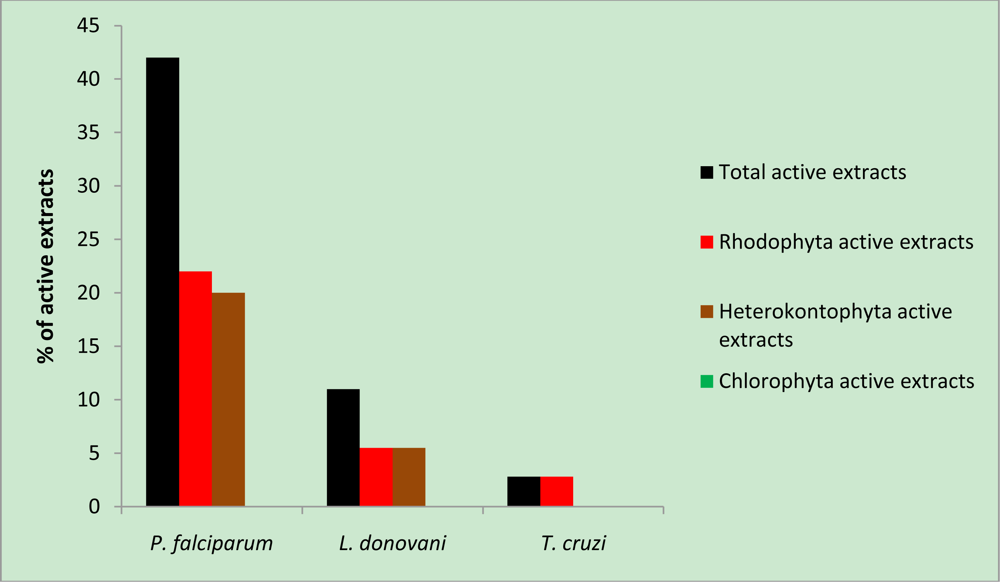
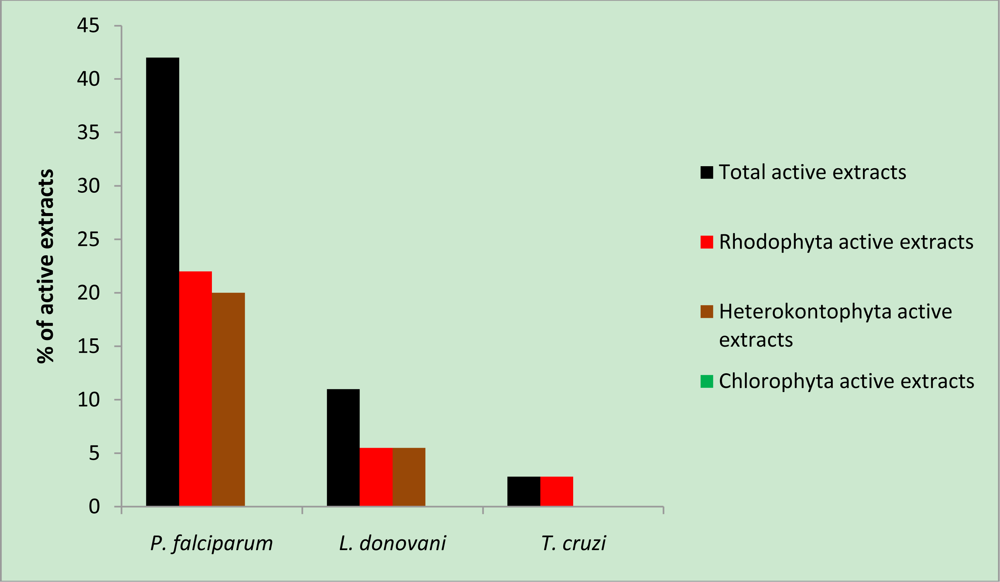
2. Results and Discussion
3. Experimental Section
3.1. Algae Collection and Identification
3.2. Preparation of Crude Extracts
3.3. In Vitro Antiprotozoal Assays
3.3.1. Activity against P. Falciparum
3.3.2. Activity against Trypanosoma cruzi
3.3.3. Activity against Leishmania Donovani
3.4. Cytotoxicity against L6 cells
3.5. Calculation of IC50
3.6. Selectivity Index Determination
4. Conclusions
Acknowledgments
References
- World Health Organization. Working to Overcome the Global Impact of Neglected Tropical Diseases. First WHO report on neglected tropical diseases, 12 October 2010. Available online: http://www.who.int/neglected_diseases/2010report/NTD_2010report_web.pdf (accessed on 14 January 2011).
- Dondorp, AM; Nosten, F; Yi, P; Das, D; Phyo, AP; Tarning, J; Lwin, KM; Ariey, F; Hanpithakpong, W; Lee, SJ; Ringwald, P; Silamut, K; Imwong, M; Chotivanich, K; Lim, P; Herdman, T; An, SS; Yeung, S; Singhasivanon, P; Day, NP; Lindegardh, N; Socheat, D; White, NJ. Artemisinin resistance in Plasmodium falciparum malaria. N Engl J Med 2009, 361, 455–467. [Google Scholar]
- Maude, RJ; Pontavornpinyo, W; Saralamba, S; Aguas, R; Yeung, S; Dondorp, AM; Day, NP; White, NJ; White, LJ. The last man standing is the most resistant: Eliminating artemisinin-resistant malaria in Cambodia. Malar J 2009, 8, 31. [Google Scholar]
- Ekland, EH; Fidock, DA. In vitro evaluations of antimalarial drugs and their relevance to clinical outcomes. Int J Parasitol 2008, 38, 743–747. [Google Scholar]
- Lin, AS; Stout, EP; Prudhomme, J; Le Roch, K; Fairchild, CR; Franzblau, SG; Aalbersberg, W; Hay, ME; Kubanek, J. Bioactive bromophycolides R–U from the Fijian red alga. Callophycus serratus J Nat Prod 2010, 73, 275–278. [Google Scholar]
- Afolayan, AF; Mann, M; Lategan, CA; Smith, PJ; Bolton, JJ; Beukes, DR. Antiplasmodial halogenated monoterpenes from the marine red alga. Plocamium cornutum Phytochemistry 2009, 70, 597–600. [Google Scholar]
- Assreuy, AMS; Gomes, DM; da Silva, MSJ; Torres, VM; Siqueira, RCL; Pires, A; Criddle, DN; de Alencar, NMN; Cavada, BS; Sampaio, AH; Farias, WRL. Biological effects of a sulfated polysaccharide isolated from the marine red algae. Champia feldmannii Biol Pharm Bull 2008, 31, 691–695. [Google Scholar]
- Mayer, AM; Rodríguez, AD; Berlinck, RG; Hamann, MT. Marine pharmacology in 2005–2006: Marine compounds with anthelmintic, antibacterial, anticoagulant, antifungal, antiinflammatory, antimalarial, antiprotozoal, antituberculosis, and antiviral activities; affecting the cardiovascular, immune and nervous systems, and other miscellaneous mechanisms of action. Biochem Biophys Acta 2009, 1790, 283–308. [Google Scholar]
- Chakraborty, K; Lipton, AP; Paulraj, R; Chakraborty, RD. Guaiane sesquiterpenes from seaweed Ulva fasciata Delile and their antibacterial properties. Eur J Med Chem 2010, 45, 2237–2244. [Google Scholar]
- Moo-Puc, R; Robledo, D; Freile-Pelegrin, Y. Evaluation of selected tropical seaweeds for in vitro anti-trichomonal activity. J Ethnopharmacol 2008, 120, 92–97. [Google Scholar]
- Orhan, I; Sener, B; Atici, T; Brun, R; Perozzo, R; Tasdemir, D. Turkish freshwater and marine macrophyte extracts show in vitro antiprotozoal activity and inhibit FabI, a key enzyme of Plasmodium falciparum fatty acid biosynthesis. Phytomedicine 2006, 13, 388–393. [Google Scholar]
- Kubanek, J; Jensen, PR; Keifer, PA; Sullards, MC; Collins, DO; Fenical, W. Seaweed resistance to microbial attack: A targeted chemical defense against marine fungi. Proc Natl Acad Sci USA 2003, 100, 6916–6921. [Google Scholar]
- Nara, T; Kamei, Y; Tsubouchi, A; Annoura, T; Hirota, K; Iizumi, K; Dohmoto, Y; Ono, T; Aoki, T. Inhibitory action of marine algae extracts on the Trypanosoma cruzi dihydroorotate dehydrogenase activity and on the protozoan growth in mammalian cells. Parasitol Int 2005, 54, 59–64. [Google Scholar]
- Freile-Pelegrin, Y; Robledo, D; Chan-Bacab, MJ; Ortega-Morales, BO. Antileishmanial properties of tropical marine algae extracts. Fitoterapia 2008, 79, 374–377. [Google Scholar]
- Spavieri, J; Allmendinger, A; Kaiser, M; Casey, R; Hingley-Wilson, S; Lalvani, A; Guiry, MD; Blunden, G; Tasdemir, D. Antimycobacterial, antiprotozoal and cytotoxic potential of twenty-one brown algae (Phaeophyceae) from British and Irish waters. Phytother Res 2010, 24, 1724–1729. [Google Scholar]
- Spavieri, J; Kaiser, M; Casey, R; Hingley-Wilson, S; Lalvani, A; Blunden, G; Tasdemir, D. Antiprotozoal, antimycobacterial and cytotoxic potential of some british green algae. Phytother Res 2010, 24, 1095–1098. [Google Scholar]
- El Hattab, M; Ben Mesaoud, M; Daoudi, M; Ortalo-Magne, A; Culioli, G; Valls, R; Piovetti, L. Trihydroxylated linear diterpenes from the brown alga Bifurcaria bifurcata (Fucales, Phaeophyta). Biochem Syst Ecol 2008, 36, 484–489. [Google Scholar]
- Ortalo-Magné, A; Culioli, G; Valls, R; Pucci, B; Piovetti, L. Polar acyclic diterpenoids from Bifurcaria bifurcata (Fucales, Phaeophyta). Phytochemistry 2005, 66, 2316–2323. [Google Scholar]
- Culioli, G; Ortalo-Magné, A; Daoudi, M; Thomas-Guyon, H; Valls, R; Piovetti, L. Trihydroxylated linear diterpenes from the brown alga. Bifurcaria bifurcata Phytochemistry 2004, 65, 2063–2069. [Google Scholar]
- Valls, R; Banaigs, B; Piovetti, L; Archavlis, A; Artaud, J. Linear diterpene with antimitotic activity from the brown alga. Bifurcaria bifurcata Phytochemistry 1993, 34, 1585–1588. [Google Scholar]
- O’Neill, MJ; Bray, DH; Boardman, P; Phillipson, JD; Warhurst, DC. Plants as sources of antimalarian drugs. In vitro test method for the evaluation of crude extracts from plants. 1985, 394–398. [Google Scholar]
- Benoit, F; Valentin, A; Pelissier, Y; Diafouka, F; Marion, C; Kone-Bamba, D; Kone, M; Mallie, M; Yapo, A; Bastide, JM. In vitro antimalarial activity of vegetal extracts used in west african traditional medicine. Am J Trop Med Hyg 1996, 54, 67–71. [Google Scholar]
- Baumeister, S; Winterberg, M; Przyborski, JM; Lingelbach, K. The malaria parasite Plasmodium falciparum: Cell biological peculiarities and nutritional consequences. Protoplasma 2010, 240, 3–12. [Google Scholar]
- Lane, AL; Stout, EP; Lin, AS; Prudhomme, J; Le Roch, K; Fairchild, CR; Franzblau, SG; Hay, ME; Aalbersberg, W; Kubanek, J. Antimalarial bromophycolides J–Q from the Fijian red alga Callophycus serratus. J Org Chem 2009, 74, 2736–2742. [Google Scholar]
- Houghton, PJ; Raman, A. Laboratory Handbook for the Fractionation of Natural Extracts, 1st ed; Chapman & Hall: London, UK, 1998; p. 49. [Google Scholar]
- Scala, F; Fattorusso, E; Menna, M; Taglialatela-Scafati, O; Tierney, M; Kaiser, M; Tasdemir, D. Bromopyrrole alkaloids as lead compounds against protozoan parasites. Mar Drugs 2010, 8, 2162–2174. [Google Scholar]
- Thaithong, S; Beale, GH. Resistance of ten Thai isolates of Plasmodium falciparum to chloroquine and pyrimethamine by in vitro tests. Trans Roy Soc Trop Med Hyg 1981, 75, 271–273. [Google Scholar]
- Buckner, FS; Verlinde, CL; La Flamme, AC; Van Voorhis, WC. Efficient technique for screening drugs for activity against Trypanosoma cruzi using parasites expressing beta-galactosidase. Antimicrob Agents Chemother 1996, 40, 2592–2597. [Google Scholar]
- Huber, W; Koella, JC. A comparison of the three methods of estimating EC50 in studies of drug resistance of malaria parasites. Acta Trop 1993, 55, 257–261. [Google Scholar]

{kind=link}
| Species | Family | Collection site | Collection time |
|---|---|---|---|
| Chlorophyta | |||
| Codium tomentosum Stackhouse | Codiaceae | Cap Lévy (Manche) | June 2007 |
| Ulva lactuca (Linnaeus) | Ulvaceae | Luc-sur-Mer (Calvados) | October 2006 |
| Ulva clathrata (Roth) C. Agardh | Ulvaceae | Anse St Martin (Manche) | June 2007 |
| Heterokontophyta | |||
| Bifurcaria bifurcata R. Ross | Sargassaceae | Cap Lévy (Manche) | June 2007 |
| Dictyopteris polypodioides (A.P. de Candolle) | Dictyotaceae | Barneville (Calvados) | October 2007 |
| J.V. Lamouroux | June 2007 | ||
| Dictyota dichotoma (Hudson) J.V. Lamouroux | Dictyotaceae | Anse St Martin (Manche) | June 2007 |
| Fucus serratus (Linnaeus) | Fucaceae | Luc-sur-mer (Calvados) | November 2005 |
| Himanthalia elongata (Linnaeus) | Himanthaliaceae | Cap Lévy (Manche) | June 2006 |
| Laminaria digitata (Linnaeus) J.V. Lamouroux | Laminariaceae | Langrunes-sur-Mer (Calvados) | January 2007 |
| Pelvetia canaliculata Decaisne & Thuret | Fucaceae | Cap Lévy (Manche) | June 2006 |
| Sargassum muticum (Yendo) Fensholt | Sargassaceae | Cap Lévy (Manche) | June 2006 |
| Rhodophyta | |||
| Calliblepharis jubata (Goodenough & woodward) Kützing | Cystocloniaceae | Cap Lévy (Manche) | June 2007 |
| Chondrus crispus Stackhouse | Gigartinaceae | Cap Lévy (Manche) | June 2007 |
| Dilsea carnosa (Schmidel) Kuntze | Dumontiaceae | Langrune-sur-Mer (Calvados) | January 2007 |
| Gelidium latifolium Bornet ex Hauck | Gelidiaceae | Cap Lévy (Manche) | June 2006 |
| Gracilaria gracilis (Stackhouse) Steentoft, L.M. Irvine & Farnham | Gracilariaceae | Anse St Martin (Manche) | June 2007 |
| Grateloupia turuturu Yamada | Halymeniaceae | St Vaast-la-Hougue (Manche) | September 2007 |
| Halurus flosculosus (J. Ellis) Maggs & Hommersand | Ceramiaceae | Anse St Martin (Manche) | June 2007 |
| Mastocarpus stellatus (Stackhouse) Guiry | Phyllophoraceae | Cap Lévy (Manche) | June 2006 |
| Palmaria palmata (Linnaeus) Kuntze | Palmariaceae | Luc-sur-Mer (Calvados) | November 2005 |
| Parasite growth inhibition (%) | |||||||
|---|---|---|---|---|---|---|---|
| P. falciparum Erythrocytic stages | T. cruzi Trypomastigotes | L. donovani Axenic amastigotes | |||||
| Species | Extract | 1.6 μg/mL | 9.7μg/mL | 1.6 μg/mL | 9.7 μg/mL | 1.6 μg/mL | 9.7 μg/mL |
| B. bifurcata | E | 0 | 25 | 18 | 0 | 16 | 40 |
| A | 36 | 100 | 0 | 78 | 31 | 100 | |
| C. jubata | E | 0 | 0 | 11 | 0 | 13 | 34 |
| A | 3 | 71 | 19 | 0 | 20 | 40 | |
| C. tomentosum | E | 0 | 2 | 0 | 0 | 15 | 29 |
| A | 0 | 0 | 0 | 0 | 0 | 0 | |
| C. crispus | E | 0 | 0 | 0 | 0 | 13 | 95 |
| A | 28 | 92 | 1 | 0 | 0 | 12 | |
| D. polypodioides | E | nd | |||||
| A | 8 | 81 | 6 | 7 | 14 | 41 | |
| D. dichotoma | E | nd | |||||
| A | 19 | 98 | 21 | 17 | 27 | 83 | |
| D. carnosa | E | 0 | 6 | 0 | 0 | 0 | 15 |
| A | 9 | 71 | 10 | 14 | 9 | 31 | |
| F. serratus | E | 0 | 0 | 0 | 0 | 0 | 15 |
| A | 0 | 42 | 0 | 3 | 0 | 14 | |
| G. latifolium | E | nd | |||||
| A | 1 | 93 | 0 | 0 | 18 | 49 | |
| G. gracilis | E | 0 | 0 | 0 | 9 | 21 | 29 |
| A | 5 | 92 | 0 | 0 | 19 | 36 | |
| G. turuturu | E | nd | |||||
| A | 42 | 97 | 15 | 14 | 9 | 33 | |
| H. flosculosus | E | nd | |||||
| A | 19 | 94 | 17 | 21 | 22 | 49 | |
| H. elongata | E | 0 | 5 | 17 | 13 | 20 | 40 |
| A | 14 | 81 | 0 | 18 | 13 | 43 | |
| L. digitata | E | 0 | 0 | 0 | 9 | 0 | 11 |
| A | 0 | 64 | 0 | 3 | 0 | 8 | |
| M. stellatus | E | 0 | 0 | 0 | 4 | 11 | 20 |
| A | 42 | 94 | 8 | 0 | 22 | 39 | |
| P. palmata | E | 0 | 0 | 0 | 0 | 0 | 0 |
| A | 0 | 1 | 17 | 36 | 0 | 10 | |
| P. canaliculata | E | 15 | 30 | 0 | 0 | 7 | 32 |
| A | 0 | 41 | 0 | 13 | 14 | 37 | |
| S. muticum | E | 0 | 0 | 0 | 1 | 8 | 37 |
| A | 50 | 96 | 0 | 9 | 4 | 48 | |
| U. lactuca | E | 0 | 0 | 0 | 3 | 0 | 26 |
| A | 0 | 28 | 0 | 0 | 0 | 11 | |
| U. clathrata | E | 0 | 0 | 0 | 0 | 0 | 0 |
| A | 0 | 13 | 0 | 0 | 0 | 7 | |
| Standard drugs | 0.003 μg/mL | 0.018 μg/mL | 0.5 μg/mL | 2.4 μg/mL | 0.2 μg/mL | 1.2 μg/mL | |
| Artemisinin | 68 | 100 | |||||
| Benznidazole | 45 | 91 | |||||
| Miltefosine | 59 | 85 | |||||
| IC50 (μg/mL) | Selectivity index (SI) | |||
|---|---|---|---|---|
| Antiprotozoal activity | Cytotoxic activity | |||
| Species | P. falciparum | L. donovani | L6 cells | |
| B. bifurcata | >5 | 3.8 | 6 | 1.6 b |
| C. jubata | 5 | nd | 71 | 14 a |
| C. crispus | 2.9 | nd | 84 | 29 a |
| D. polypodioides | nd | 10.8 | 87 | 8 b |
| D. dichotoma | 3.1 | 8.8 | 27 | 9 a |
| D. carnosa | 3.9 | 9.5 | 74 | 19 a |
| G. latifolium | 3.4 | nd | 62 | 18 a |
| G. gracilis | 3.3 | nd | 71 | 21 a |
| G. turuturu | 3.1 | nd | 71 | 23 a |
| H. flosculosus | 4.6 | nd | 58 | 12 a |
| H. elongata | 3.5 | nd | 88 | 25 a |
| M. stellatus | 2.8 | nd | >90 | >30 a |
| P. canaliculata | nd | nd | 87 | 11 a |
| S. muticum | 2.9 | nd | 63 | 11 a |
| Standards | ||||
| Chloroquine | 0.069 | - | - | - |
| Miltefosine | - | 0.181 | - | - |
| Podophyllotoxin | - | - | 0.007 | - |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Vonthron-Sénécheau, C.; Kaiser, M.; Devambez, I.; Vastel, A.; Mussio, I.; Rusig, A.-M. Antiprotozoal Activities of Organic Extracts from French Marine Seaweeds. Mar. Drugs 2011, 9, 922-933. https://doi.org/10.3390/md9060922
Vonthron-Sénécheau C, Kaiser M, Devambez I, Vastel A, Mussio I, Rusig A-M. Antiprotozoal Activities of Organic Extracts from French Marine Seaweeds. Marine Drugs. 2011; 9(6):922-933. https://doi.org/10.3390/md9060922
Chicago/Turabian StyleVonthron-Sénécheau, Catherine, Marcel Kaiser, Isabelle Devambez, Antoine Vastel, Isabelle Mussio, and Anne-Marie Rusig. 2011. "Antiprotozoal Activities of Organic Extracts from French Marine Seaweeds" Marine Drugs 9, no. 6: 922-933. https://doi.org/10.3390/md9060922
APA StyleVonthron-Sénécheau, C., Kaiser, M., Devambez, I., Vastel, A., Mussio, I., & Rusig, A.-M. (2011). Antiprotozoal Activities of Organic Extracts from French Marine Seaweeds. Marine Drugs, 9(6), 922-933. https://doi.org/10.3390/md9060922