
Chemical and Physical Culture Conditions Significantly Influence the Cell Mass and Docosahexaenoic Acid Content of Aurantiochytrium limacinum Strain PKU#SW8


 ,
, 
 and
and 
Abstract
:1. Introduction
2. Results
2.1. Growth Characteristics and Fatty Acid Composition
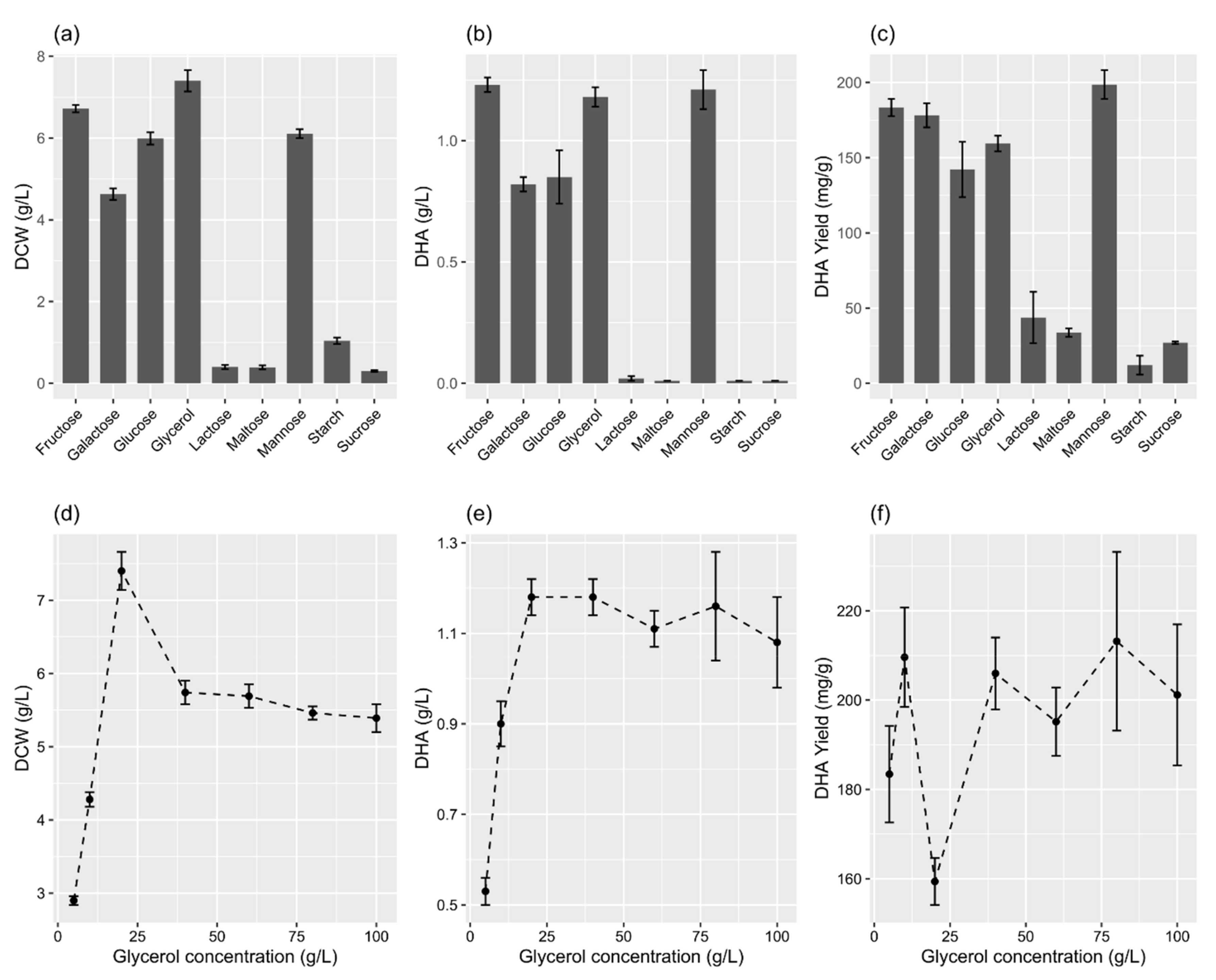
2.2. Effects of Carbon Source
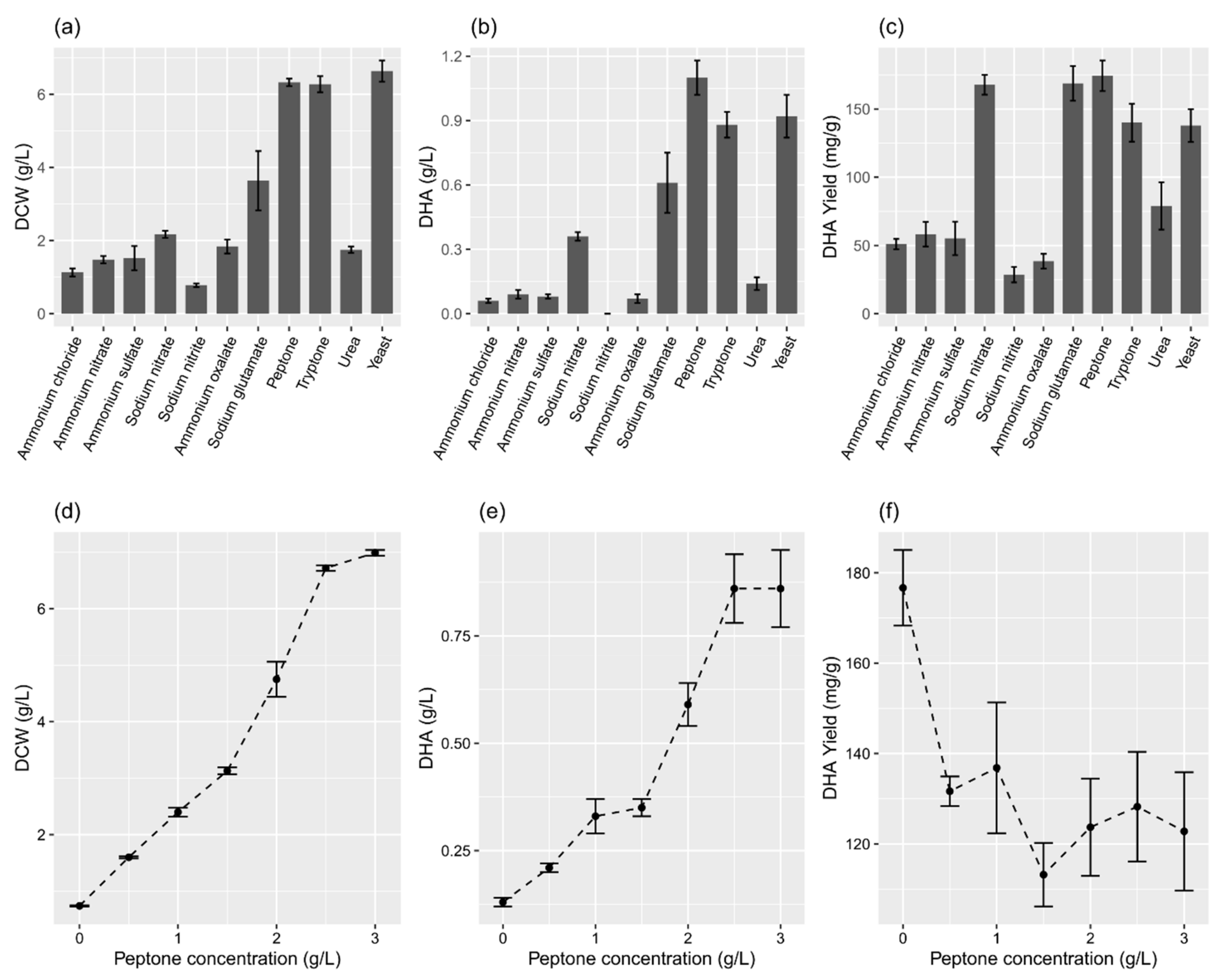
2.3. Effects of Nitrogen Source
2.4. Effects of KH2PO4 and Salinity
2.5. Effects of Environmental Variables
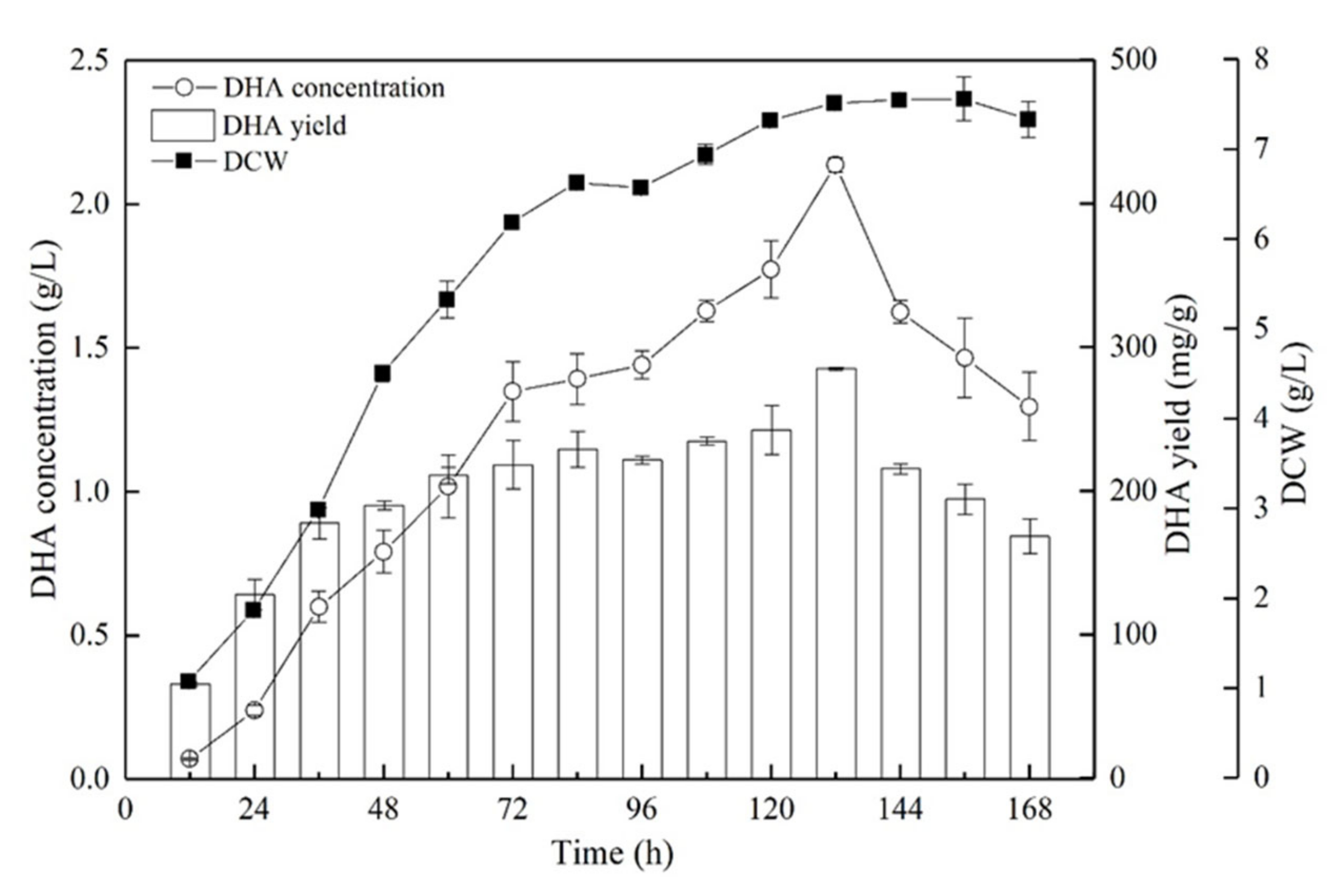
2.6. DHA Production in a 5-L Fermenter
3. Discussion
3.1. Influence of Chemical Culture Conditions
3.2. Influence of Physical Culture Conditions
4. Conclusions
5. Materials and Methods
5.1. Strain and Seed Culture
5.2. Batch Experiments
5.3. Biochemical Analyses
5.4. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Du, F.; Wang, Y.Z.; Xu, Y.S.; Shi, T.Q.; Liu, W.Z.; Sun, X.M.; Huang, H. Biotechnological production of lipid and terpenoid from thraustochytrids. Biotechnol. Adv. 2021, 48, 107725. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.; Karageorgou, D.; Katapodis, P.; Sharma, A.; Rova, U.; Christakopoulos, P.; Matsakas, L. Bioprospecting of thraustochytrids for omega-3 fatty acids: A sustainable approach to reduce dependency on animal sources. Trends Food Sci. Technol. 2021, 115, 433–444. [Google Scholar] [CrossRef]
- Aasen, I.M.; Ertesvåg, H.; Heggeset, T.M.B.; Liu, B.; Brautaset, T.; Vadstein, O.; Ellingsen, T.E. Thraustochytrids as production organisms for docosahexaenoic acid (DHA), squalene, and carotenoids. Appl. Microbiol. Biotechnol. 2016, 100, 4309–4321. [Google Scholar] [CrossRef] [PubMed]
- Lyu, L.; Wang, Q.; Wang, G. Cultivation and diversity analysis of novel marine thraustochytrids. Mar. Life Sci. Technol. 2020, 3, 263–275. [Google Scholar] [CrossRef]
- Marchan, L.F.; Chang, K.J.L.; Nichols, P.D.; Polglase, J.L.; Mitchell, W.J.; Gutierrez, T. Screening of new British thraustochytrids isolates for docosahexaenoic acid (DHA) production. J. Appl. Phycol. 2017, 29, 2831–2843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee Chang, K.J.; Paul, H.; Nichols, P.D.; Koutoulis, A.; Blackburn, S.I. Australian thraustochytrids: Potential production of dietary long-chain omega-3 oils using crude glycerol. J. Funct. Foods 2015, 19, 810–820. [Google Scholar] [CrossRef]
- Leyton, A.; Flores, L.; Shene, C.; Chisti, Y.; Larama, G.; Asenjo, J.A.; Armenta, R.E. Antarctic thraustochytrids as sources of carotenoids and high-value fatty acids. Mar. Drugs 2021, 19, 386. [Google Scholar] [CrossRef]
- Liu, Y.; Singh, P.; Sun, Y.; Luan, S.; Wang, G. Culturable diversity and biochemical features of thraustochytrids from coastal waters of southern China. Appl. Microbiol. Biotechnol. 2014, 98, 3241–3255. [Google Scholar] [CrossRef]
- Sohedein, M.N.A.; Wan-Mohtar, W.A.A.Q.I.; Ilham, Z.; Babadi, A.A.; Hui-Yin, Y.; Siew-Moi, P. Vital parameters for biomass, lipid, and carotenoid production of thraustochytrids. J. Appl. Phycol. 2020, 32, 1003–1016. [Google Scholar] [CrossRef]
- Paparini, A.; Romano-Spica, V. Public health issues related with the consumption of food obtained from genetically modified organisms. Biotechnol. Ann. Rev. 2004, 10, 85–122. [Google Scholar]
- Wang, Q.; Ye, H.; Sen, B.; Xie, Y.; He, Y.; Park, S.; Wang, G. Improved production of docosahexaenoic acid in batch fermentation by newly-isolated strains of Schizochytrium sp. and Thraustochytriidae sp. through bioprocess optimization. Synth. Sys. Biotechnol. 2018, 3, 121–129. [Google Scholar] [CrossRef]
- Chen, X.; He, Y.; Ye, H.; Xie, Y.; Sen, B.; Jiao, N.; Wang, G. Different carbon and nitrogen sources regulated docosahexaenoic acid (DHA) production of Thraustochytriidae sp. PKU#SW8 through a fully functional polyunsaturated fatty acid (PUFA) synthase gene (pfaB). Bioresour. Technol. 2020, 318, 124273. [Google Scholar]
- Song, Z.; Stajich, J.E.; Xie, Y.; Liu, X.; He, Y.; Chen, J.; Hicks, G.R.; Wang, G. Comparative analysis reveals unexpected genome features of newly isolated Thraustochytrids strains: On ecological function and PUFAs biosynthesis. BMC Genomics 2018, 19, 541. [Google Scholar] [CrossRef]
- Huang, J.; Aki, T.; Hachida, K.; Yokochi, T.; Kawamoto, S.; Shigeta, S.; Ono, K.; Suzuki, O. Profile of polyunsaturated fatty acids produced by Thraustochytrium sp. KK17-3. J. Am. Oil Chem. Soc. 2001, 78, 605–610. [Google Scholar] [CrossRef]
- Perveen, Z.; Ando, H.; Ueno, A.; Ito, Y.; Yamamoto, Y.; Yamada, Y.; Takagi, T.; Kaneko, T.; Kogame, K.; Okuyama, H. Isolation and characterization of a novel thraustochytrid-like microorganism that efficiently produces docosahexaenoic acid. Biotechnol. Lett. 2006, 28, 197–202. [Google Scholar] [CrossRef] [Green Version]
- Hong, W.K.; Rairakhwada, D.; Seo, P.S.; Park, S.Y.; Hur, B.K.; Kim, C.H.; Seo, J.W. Production of lipids containing high levels of docosahexaenoic acid by a newly isolated microalga, Aurantiochytrium sp. KRS101. Appl. Biochem. Biotechnol. 2011, 164, 1468–1480. [Google Scholar] [CrossRef]
- Gupta, A.; Singh, D.; Barrow, C.J.; Puri, M. Exploring potential use of Australian thraustochytrids for the bioconversion of glycerol to omega-3 and carotenoids production. Biochem. Eng. J. 2013, 78, 11–17. [Google Scholar] [CrossRef]
- Liu, Y.; Tang, J.; Li, J.; Daroch, M.; Cheng, J.J. Efficient production of triacylglycerols rich in docosahexaenoic acid (DHA) by osmo-heterotrophic marine protists. Appl. Microbiol. Biotechnol. 2014, 98, 9643–9652. [Google Scholar] [CrossRef]
- Manikan, V.; Nazir, M.Y.M.; Kalil, M.S.; Isa, M.H.M.; Kader, A.J.A.; Yusoff, W.M.W.; Hamid, A.A. A new strain of docosahexaenoic acid producing microalga from Malaysian coastal waters. Algal Res. 2015, 9, 40–47. [Google Scholar] [CrossRef]
- Quilodran, B.; Cortinez, G.; Bravo, A.; Silva, D. Characterization and comparison of lipid and PUFA production by native thraustochytrid strains using complex carbon sources. Heliyon 2020, 6, e05404. [Google Scholar] [CrossRef]
- Nakahara, T.; Yokochi, T.; Higashihara, T.; Tanaka, S.; Yaguchi, T.; Honda, D. Production of docosahexaenoic and docosapentaenoic acids by Schizochytrium sp. isolated from yap islands. J. Am. Oil Chem. Soc. 1996, 73, 1421–1426. [Google Scholar] [CrossRef]
- Zhu, L.; Zhang, X.; Ren, X.; Zhu, Q. Effects of culture conditions on growth and docosahexaenoic acid production from Schizochytrium limacinum. J. Ocean Univ. China 2008, 7, 83–88. [Google Scholar] [CrossRef]
- Sen, B.; Li, J.; Lu, L.; Bai, M.; He, Y.; Wang, G. Elemental composition and cell mass quantification of cultured thraustochytrids unveil their large contribution to marine carbon pool. Mar. Drugs 2021, 19, 493. [Google Scholar] [CrossRef] [PubMed]
- Chaung, K.C.; Chu, C.Y.; Su, Y.M.; Chen, Y.M. Effect of culture conditions on growth, lipid content, and fatty acid composition of Aurantiochytrium mangrovei strain BL10. AMB Express 2012, 2, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taoka, Y.; Nagano, N.; Okita, Y.; Izumida, H.; Sugimoto, S.; Hayashi, M. Effect of addition of Tween 80 and potassium dihydrogenphosphate to basal medium on the isolation of marine eukaryotes, thraustochytrids. J. Biosci. Bioeng. 2008, 105, 562–565. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Sen, B.; Liu, X.; He, Y.; Xie, Y.; Wang, G. Enhanced saturated fatty acids accumulation in cultures of newly-isolated strains of Schizochytrium sp. and Thraustochytriidae sp. for large-scale biodiesel production. Sci. Total Environ. 2018, 631–632, 994–1004. [Google Scholar] [CrossRef] [PubMed]
- Raghukumar, S. Thraustochytrid marine protists: Production of PUFAs and other emerging technologies. Mar. Biotechnol. 2008, 10, 631–640. [Google Scholar] [CrossRef]
- Zhang, A.; Mernitz, K.; Wu, C.; Xiong, W.; He, Y.; Wang, G.; Wang, X. ATP drives efficient terpene biosynthesis in marine thraustochytrids. mBio 2021, 12, e0088121. [Google Scholar] [CrossRef]
- Chatdumrong, W.; Yongmanitchail, W.; Limtong, S.; Worawattanamateekul, W. Optimization of docosahexaenoic acid (DHA) production and improvement of astaxanthin content in a mutant Schizochytrium limacinum isolated from mangrove forest in Thailand. Kasetsart J. 2007, 41, 324–334. [Google Scholar]
- Yokochi, T.; Honda, D.; Higashihara, T.; Nakahara, T. Optimization of docosahexaenoic acid production by Schizochytrium limacinum SR21. Appl. Microbiol. Biotechnol. 1998, 49, 72–76. [Google Scholar] [CrossRef]
- Zhu, L.; Zhang, X.; Ji, L.; Song, X.; Kuang, C. Changes of lipid content and fatty acid composition of Schizochytrium limacinum in response to different temperatures and salinities. Process Biochem. 2007, 42, 210–214. [Google Scholar] [CrossRef]
- Półbrat, T.; Konkol, D.; Korczyński, M. Optimization of docosahexaenoic acid production by Schizochytrium sp.—A review. Biocatal. Agric. Biotechnol. 2021, 35, 102076. [Google Scholar] [CrossRef]
- Fan, K.W.; Vrijmoed, L.L.P.; Jones, E.B.G. Physiological studies of subtropical mangrove thraustochytrids. Bot. Mar. 2002, 45, 50–57. [Google Scholar] [CrossRef]
- Wu, S.-T.; Yu, S.-T.; Lin, L.-P. Effect of culture conditions on docosahexaenoic acid production by Schizochytrium sp. S31. Process Biochem. 2005, 40, 3103–3108. [Google Scholar] [CrossRef]
- Yin, F.W.; Zhang, Y.T.; Jiang, J.Y.; Guo, D.S.; Gao, S.; Gao, Z. Efficient docosahexaenoic acid production by Schizochytrium sp. via a two-phase pH control strategy using ammonia and citric acid as pH regulators. Process Biochem. 2019, 77, 1–7. [Google Scholar] [CrossRef]
- Zhao, B.; Li, Y.; Mbifile, M.D.; Li, C.; Yang, H.; Wang, W. Improvement of docosahexaenoic acid fermentation from Schizochytrium sp. AB-610 by staged pH control based on cell morphological changes. Eng. Life Sci. 2017, 17, 981–988. [Google Scholar] [CrossRef] [Green Version]
- Taoka, Y.; Nagano, N.; Okita, Y.; Izumida, H.; Sugimoto, S.; Hayashi, M. Influences of culture temperature on the growth, lipid content and fatty acid composition of Aurantiochytrium sp. strain mh0186. Mar. Biotechnol. 2009, 11, 368–374. [Google Scholar] [CrossRef]
- Zhou, Y.; Han, L.R.; He, H.W.; Sang, B.; Yu, D.L.; Feng, J.T.; Zhang, X. Effects of agitation, aeration and temperature on production of a novel glycoprotein GP-1 by Streptomyces kanasenisi ZX01 and scale-up based on volumetric oxygen transfer coefficient. Molecules 2018, 23, 125. [Google Scholar] [CrossRef] [Green Version]
- Heggeset, T.M.B.; Ertesvåg, H.; Liu, B.; Ellingsen, T.E.; Vadstein, O.; Aasen, I.M. Lipid and DHA-production in Aurantiochytrium sp.—Responses to nitrogen starvation and oxygen limitation revealed by analyses of production kinetics and global transcriptomes. Sci. Rep. 2019, 9, 19470. [Google Scholar] [CrossRef]
- Damare, V.; Raghukumar, S. Morphology and Physiology of the Marine Straminipilan Fungi, the Aplanochytrids Isolated from the Equatorial Indian Ocean. Indian J. Mar. Sci. 2006, 35, 326–340. [Google Scholar]
- Jain, R.; Raghukumar, S.; Tharanathan, R.; Bhosle, N.B. Extracellular polysaccharide production by thraustochytrid protists. Mar. Biotechnol. 2005, 7, 184–192. [Google Scholar] [CrossRef]
- Lepage, G.; Roy, C.C. Improved recovery of fatty acid through direct transesterification without prior extraction or purification. J. Lipid Res. 1984, 25, 1391–1396. [Google Scholar] [CrossRef]
- Zhang, S.; He, Y.; Sen, B.; Chen, X.; Xie, Y.; Keasling, J.D.; Wang, G. Alleviation of reactive oxygen species enhances PUFA accumulation in Schizochytrium sp. through regulating genes involved in lipid metabolism. Metab. Eng. Commun. 2018, 6, 39–48. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acid | Concentration (g/L) | Fraction (%) |
|---|---|---|
| C12:0 | 0.004 ± 0 | 0.2 ± 0.01 |
| C14:0 | 0.095 ± 0.006 | 4.51 ± 0.29 |
| C15:0 | 0.038 ± 0.002 | 1.81 ± 0.12 |
| C15:1 (n-5) | 0.007 ± 0.001 | 0.33 ± 0.06 |
| C16:0 | 0.832 ± 0.056 | 39.67 ± 2.66 |
| C16:1 (n-7) | 0.003 ± 0 | 0.13 ± 0.02 |
| C17:0 | 0.01 ± 0.001 | 0.48 ± 0.03 |
| C18:0 | 0.02 ± 0.001 | 0.93 ± 0.06 |
| C20:3 (n-3) | 0.009 ± 0.002 | 0.52 ± 0.03 |
| C21:0 | 0.011 ± 0.001 | 0.45 ± 0.09 |
| C20:5 (n-3) | 0.057 ± 0.005 | 2.71 ± 0.24 |
| C22:1 (n-9) | 0.009 ± 0.001 | 0.45 ± 0.04 |
| C22:5 (n-6) | 0.152 ± 0.01 | 7.24 ± 0.46 |
| C22:6 (n-3) | 0.839 ± 0.05 | 40.01 ± 2.4 |
| TFA | 2.097 ± 0.127 | 100 |
| SFA | 1.012 ± 0.067 | 48.25 ± 3.2 |
| PUFA | 1.064 ± 0.065 | 50.74 ± 2.95 |
| MUFA | 0.019 ± 0.001 | 0.89 ± 0.07 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, X.; Sen, B.; Zhang, S.; Bai, M.; He, Y.; Wang, G. Chemical and Physical Culture Conditions Significantly Influence the Cell Mass and Docosahexaenoic Acid Content of Aurantiochytrium limacinum Strain PKU#SW8. Mar. Drugs 2021, 19, 671. https://doi.org/10.3390/md19120671
Chen X, Sen B, Zhang S, Bai M, He Y, Wang G. Chemical and Physical Culture Conditions Significantly Influence the Cell Mass and Docosahexaenoic Acid Content of Aurantiochytrium limacinum Strain PKU#SW8. Marine Drugs. 2021; 19(12):671. https://doi.org/10.3390/md19120671
Chicago/Turabian StyleChen, Xiaohong, Biswarup Sen, Sai Zhang, Mohan Bai, Yaodong He, and Guangyi Wang. 2021. "Chemical and Physical Culture Conditions Significantly Influence the Cell Mass and Docosahexaenoic Acid Content of Aurantiochytrium limacinum Strain PKU#SW8" Marine Drugs 19, no. 12: 671. https://doi.org/10.3390/md19120671
APA StyleChen, X., Sen, B., Zhang, S., Bai, M., He, Y., & Wang, G. (2021). Chemical and Physical Culture Conditions Significantly Influence the Cell Mass and Docosahexaenoic Acid Content of Aurantiochytrium limacinum Strain PKU#SW8. Marine Drugs, 19(12), 671. https://doi.org/10.3390/md19120671