
Anti-Inflammatory Activity of 4-((1R,2R)-3-Hydroxy-1-(4-hydroxyphenyl)-1-methoxypropan-2-yl)-2-methoxyphenol Isolated from Juglans mandshurica Maxim. in LPS-Stimulated RAW 264.7 Macrophages and Zebrafish Larvae Model

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
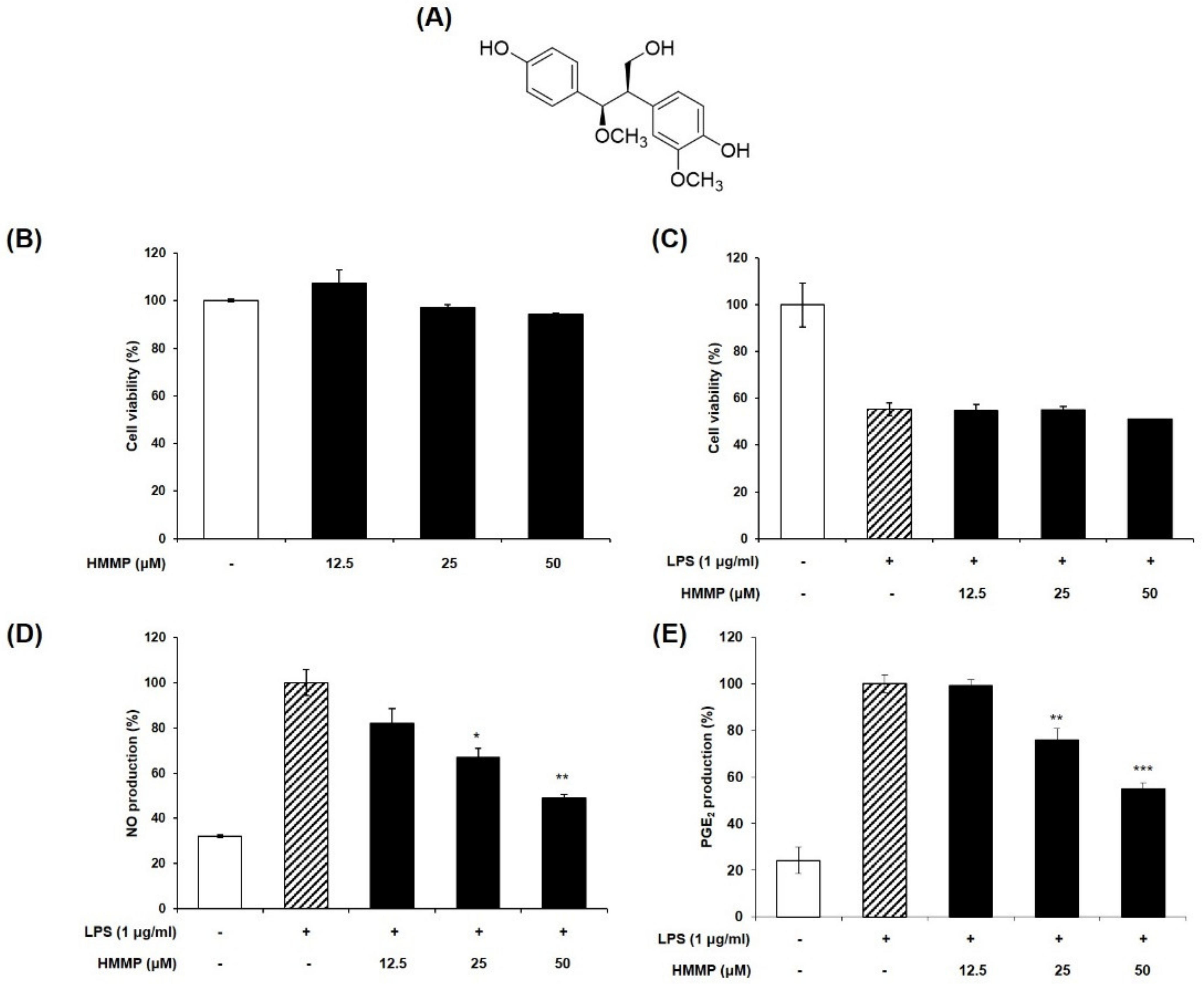
2.1. Effect of HHMP on LPS-Induced NO and PGE2 Production and Cell Viability in RAW 264.7 Cells
2.2. Effect of HHMP on LPS-Induced iNOS and COX-2 Protein Expression in RAW 264.7 Cells
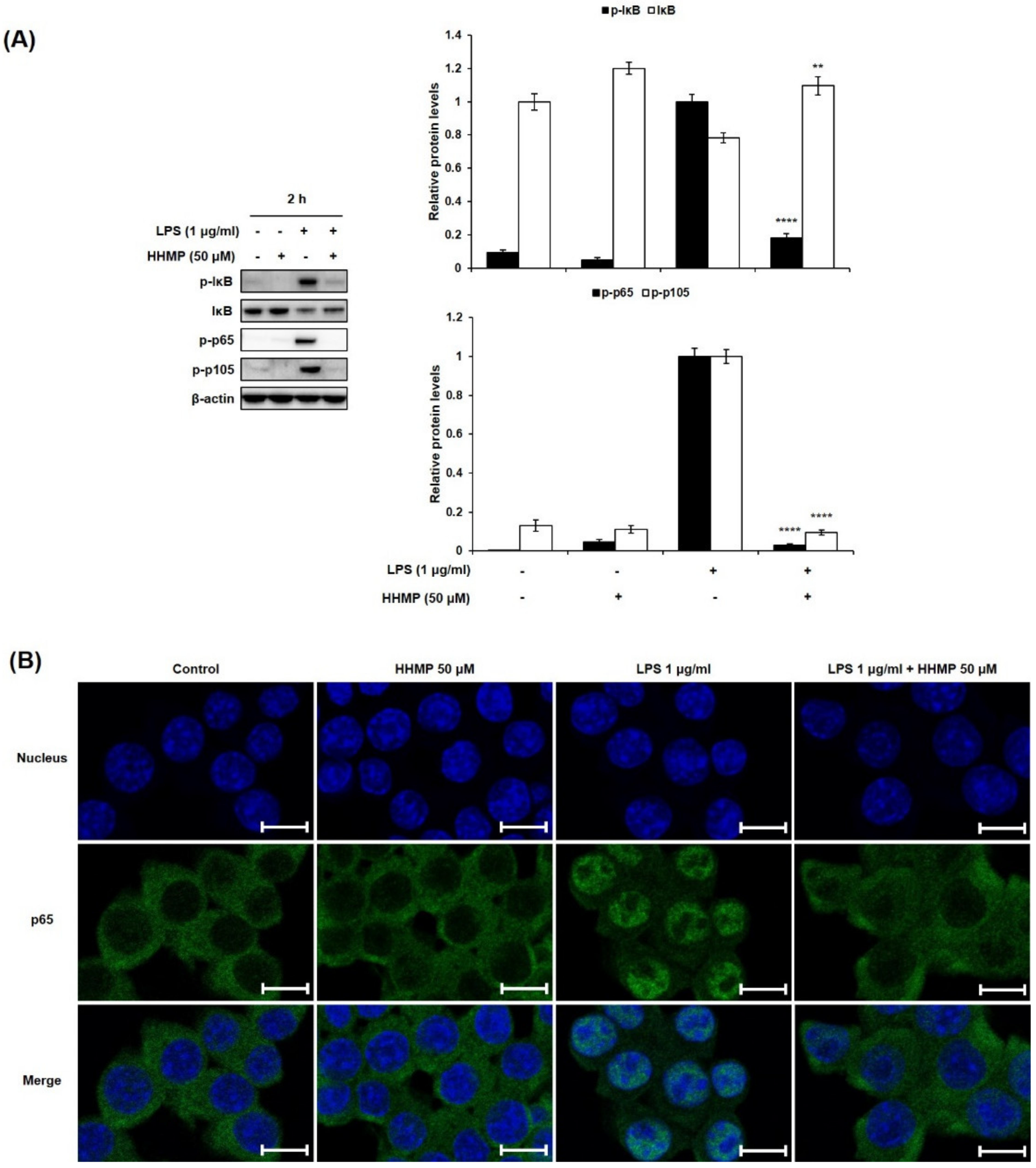
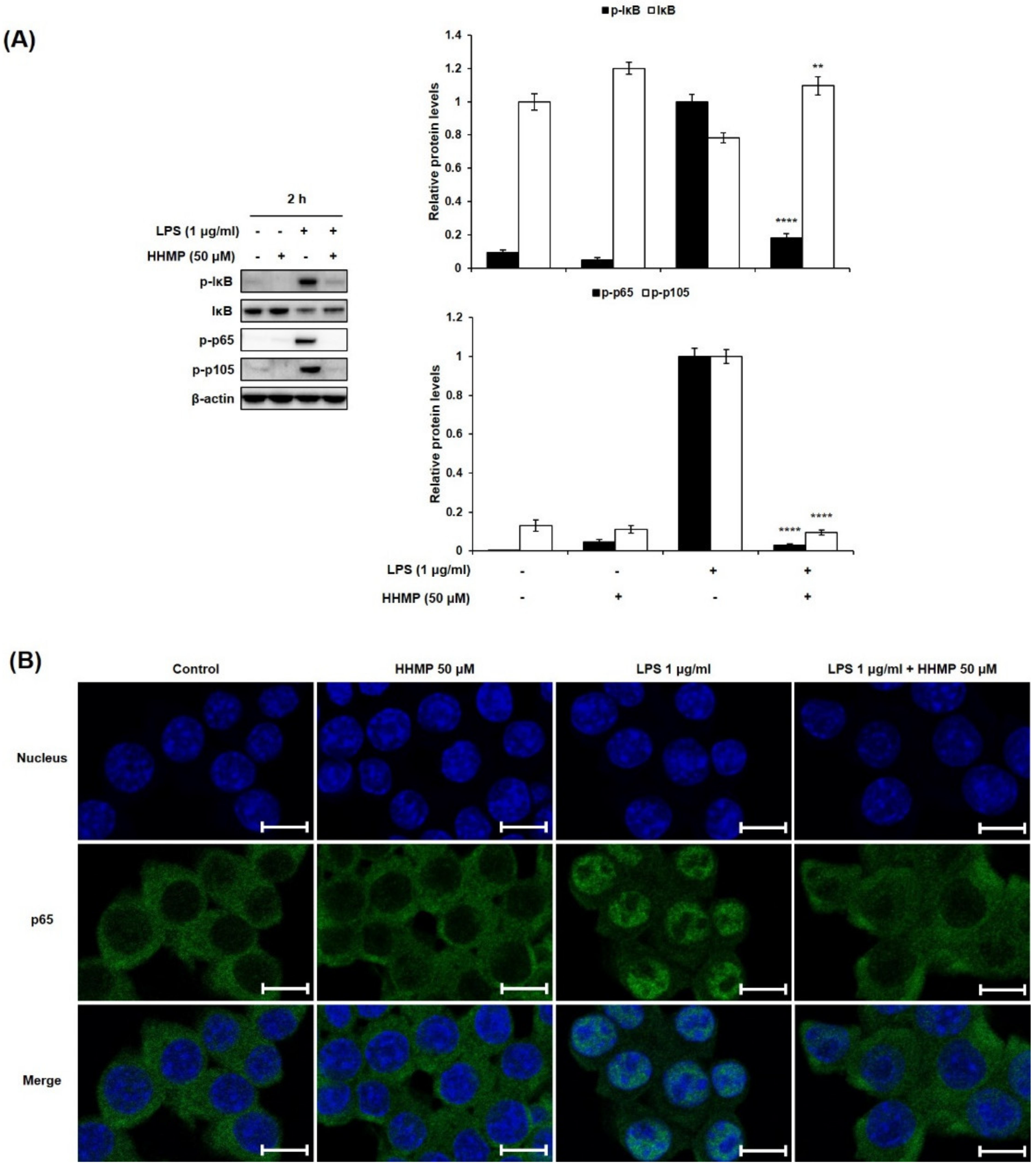
2.3. Effect of HHMP on LPS-Induced NF-κB Activation in RAW 264.7 Cells
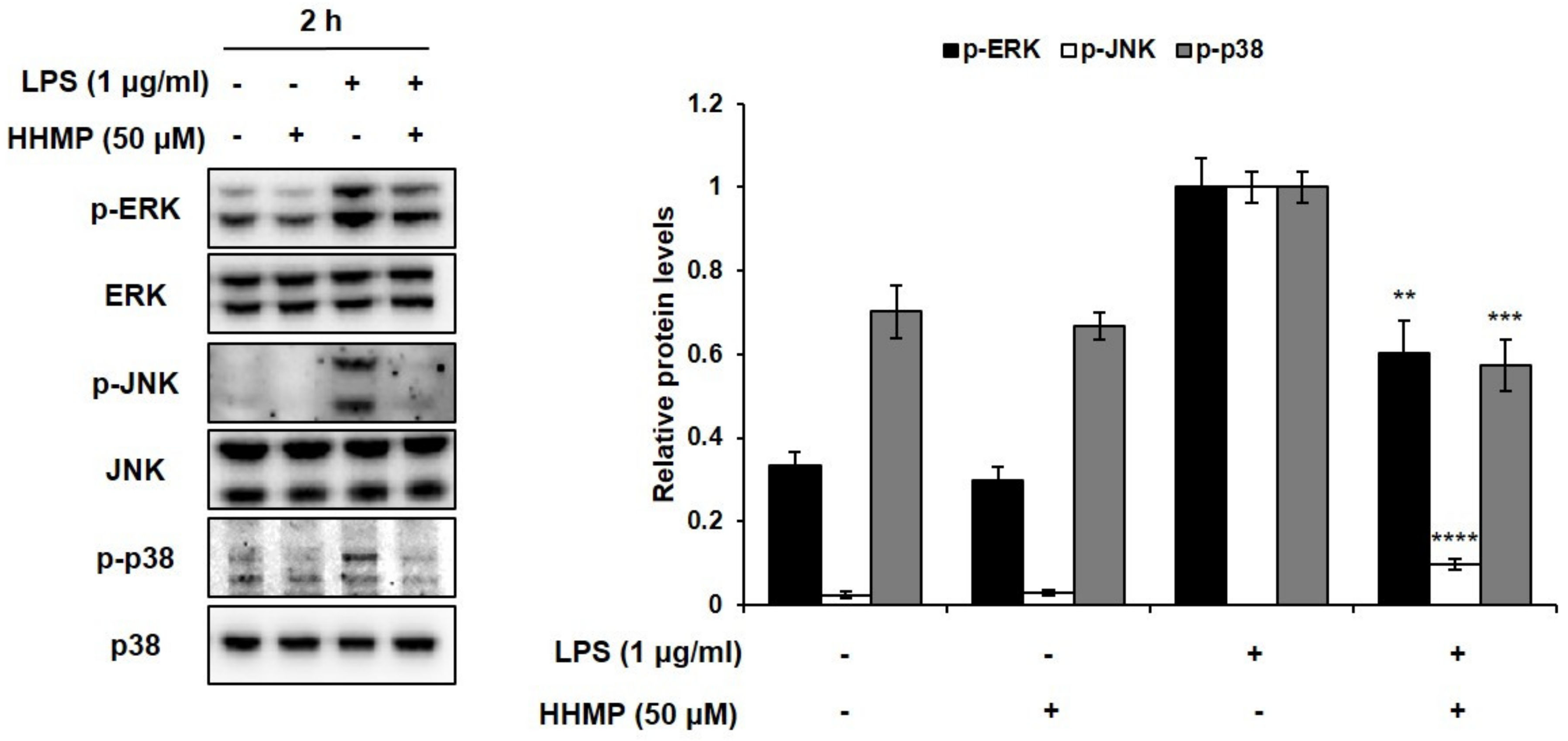
2.4. Effect of HHMP on LPS-Induced MAPK Activation in RAW 264.7 Cells
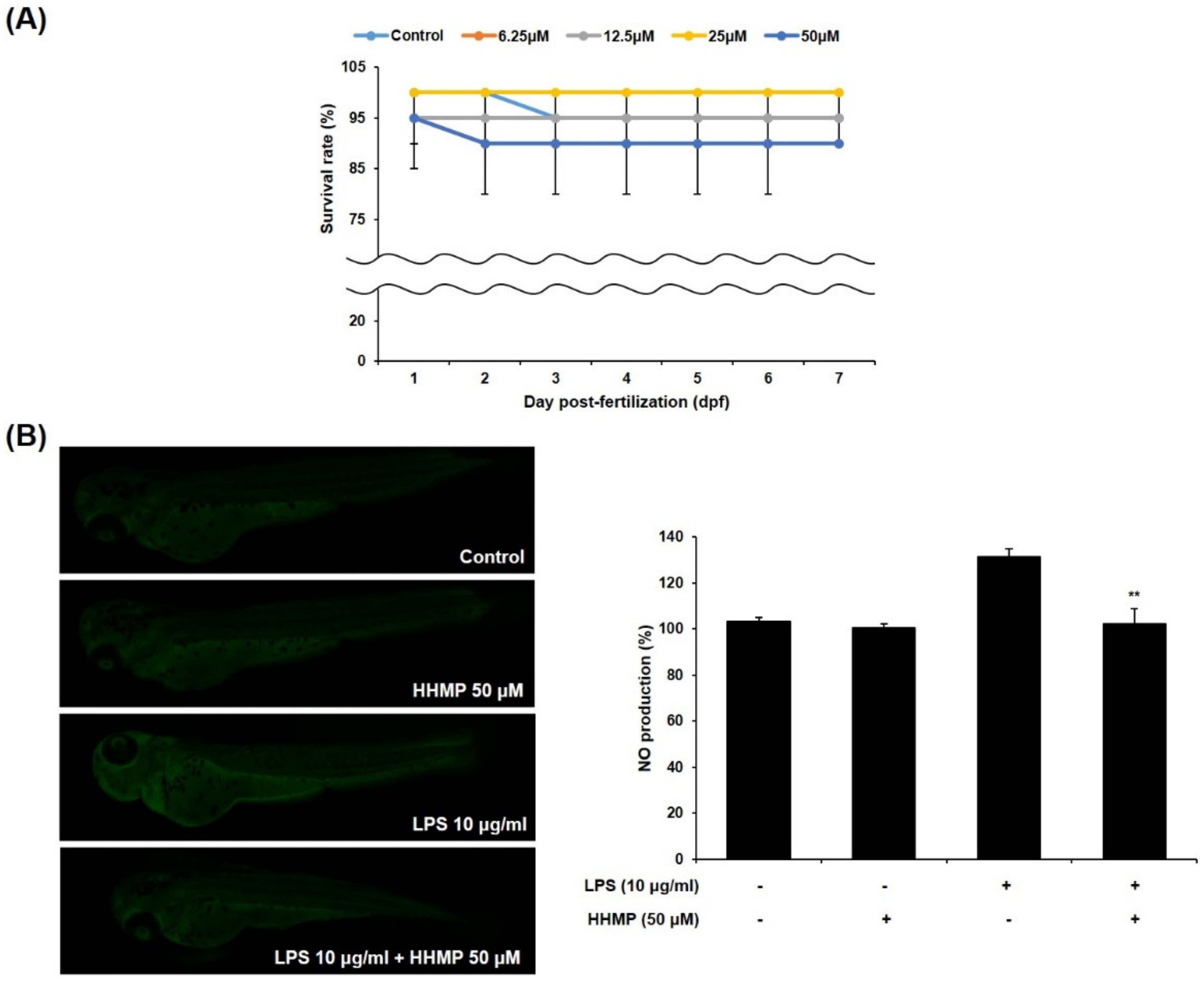
2.5. Effect of HHmP on Survival Rate and NO Production in LPS-Stimulated Zebrafish Larvae
3. Discussion
4. Materials and Methods
4.1. Extraction and Isolation of HHMP from J. mandshurica
4.2. Cell Culture
4.3. Measurement of Cell Viability
4.4. Nitric Oxide (NO) Assay
4.5. Measurement of Prostaglandin E2 (PGE2) Production
4.6. Western Blot Analysis
4.7. Immunofluorescence Staining
4.8. Maintenance of Adult Zebrafish and Survival Rate Measurement
4.9. Maintenance of NO Production in Zebrafish Larvae
5. Statistical Analysis
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Soonthornsit, N.; Pitaksutheepong, C.; Hemstapat, W.; Utaisincharoen, P.; Pitaksuteepong, T. In Vitro Anti-Inflammatory Activity of Morus alba L. Stem Extract in LPS-Stimulated RAW 264.7 Cells. Evid. Based Complement. Altern. Med. 2017, 2017, 3928956. [Google Scholar] [CrossRef] [Green Version]
- Sanjeewa, K.K.A.; Fernando, I.P.S.; Kim, E.A.; Ahn, G.; Jee, Y.; Jeon, Y.J. Anti-inflammatory activity of a sulfated polysaccharide isolated from an enzymatic digest of brown seaweed Sargassum horneri in RAW 264.7 cells. Nutr. Res. Pract. 2017, 11, 3–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, H.Y.; Shin, E.M.; Guo, L.Y.; Youn, U.J.; Bae, K.; Kang, S.S.; Zou, L.B.; Kim, Y.S. Anti-inflammatory activity of 4-methoxyhonokiol is a function of the inhibition of iNOS and COX-2 expression in RAW 264.7 macrophages via NF-κB, JNK and p38 MAPK inactivation. Eur. J. Pharmacol. 2008, 586, 340–349. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.-H.; Lai, C.-S.; Ho, C.-T. Anti-inflammatory activity of natural dietary flavonoids. Food Funct. 2010, 1, 15–31. [Google Scholar] [CrossRef]
- Xie, Z.; Wang, Y.; Huang, J.; Qian, N.; Shen, G.; Chen, L. Anti-inflammatory activity of polysaccharides from Phellinus linteus by regulating the NF-κB translocation in LPS-stimulated RAW 264.7 macrophages. Int. J. Biol. Macromol. 2019, 129, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Clarke, D.; Damera, G.; Sukkar, M.B.; Tliba, O. Transcriptional regulation of cytokine function in airway smooth muscle cells. Pulm. Pharmacol. Ther. 2009, 22, 436–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, C.; Hui, T.; Ting, F.; Jianbo, X. Agrimonolide from Agrimonia pilosa suppresses inflammatory responses through down-regulation of COX-2/iNOS and inactivation of NF-κB in lipopolysaccharide-stimulated macrophages. Phytomedicine 2016, 23, 846–855. [Google Scholar]
- Li, Q.; Verma, I.M. NF-κB regulation in the immune system. Nat. Rev. Immunol. 2002, 2, 725–734. [Google Scholar] [CrossRef]
- Oeckinghaus, A.; Ghosh, S. The NF-κB family of transcription factors and its regulation. Cold Spring Harb. Perspect. Biol. 2009, 1, a000034. [Google Scholar] [CrossRef]
- Hasnat, M.A.; Pervin, M.; Cha, K.M.; Kim, S.K.; Lim, B.O. Anti-inflammatory activity on mice of extract of Ganoderma lucidum grown on rice via modulation of MAPK and MF-κB pathways. Phytochemistry 2015, 114, 125–136. [Google Scholar] [CrossRef]
- Lv, H.; Ren, W.; Zheng, Y.; Wang, L.; Lu, G.; Yi, P.; Ci, X. Tenuigenin exhibits anti-inflammatory activity via inhibiting MAPK and NF-κB and inducing Nrf2/HO-1 signaling in macrophages. Food Funt. 2016, 7, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Wongrakpanich, S.; Wonggrakpanich, A.; Melhado, K.; Rangaswami, J. A comprehensive review of non-steroidal anti-inflammatory drug use in the elderly. Aging Dis. 2018, 9, 143–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.M.; Park, S.H. Risk and benefit of steroid therapy. Korean J. Intern. Med. 2009, 77, 298–303. [Google Scholar]
- Yao, D.L.; Zhang, C.H.; Luo, J.; Jin, M.; Zheng, M.S.; Cui, J.M.; Son, J.K.; Li, G. Chemical constituents from the leaves of Juglans mandshurica. Arch. Pharm. Res. 2014, 38, 480–484. [Google Scholar] [CrossRef] [PubMed]
- Park, G.; Oh, M.S. Inhibitory effects of Juglans mandshurica leaf on allergic dermatitis-like skin lesions-induced by 2,4-dinitrochlorobenzene in mice. Exp. Toxicol. Pathol. 2014, 66, 97–101. [Google Scholar] [CrossRef]
- Liu, L.; Li, W.; Koike, K.; Zhang, S.; Nikaido, T. New α-tetralonyl glucosides from the fruit of Juglans mandshurica. Chem. Pharm. Bull. 2004, 52, 566–569. [Google Scholar] [CrossRef] [Green Version]
- Xin, N.; Li, Y.; Hasan, M.; Li, W. Juglans mandshurica Maxim extracts exhibit antitumor activity on HeLa cells in vitro. Mol. Med. Rep. 2014, 9, 1313–1318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Du, K.; Fang, L.; Liu, C.; Min, W.; Liu, J. Evaluation of the antidiabetic activity of hydrolyzed peptides derived from Juglans mandshurica Maxim. fruits in insulin-resistant HepG2 cells and type 2 diebetic mice. J. Food Biochem. 2018, 42, e12518. [Google Scholar] [CrossRef]
- Park, S.J.; Kim, N.; You, G.; Kim, S.N.; Kwon, H.J.; Jung, K.; Oh, O.C.; Lee, Y.H.; Kim, S.H. Phenolic and neolignans isolated from the fruits of Juglans mandshurica Maxim. and their effect on lipolysis in adipocyte. Phytochemistry 2017, 137, 87–93. [Google Scholar] [CrossRef]
- Kim, J.Y.; Lee, E.J.; Ahn, Y.; Park, S.J.; Kim, S.H.; Oh, S.H. A chemical compound from fruit extract of Juglans mandshurica inhibits melanogenesis through p-ERK-associated MITF degradation. Phytomedicine 2019, 57, 57–64. [Google Scholar] [CrossRef]
- Yang, B.Y.; Liu, Y.; Jiang, H.B.; Xu, Z.P.; Guo, R.; Wang, R.; Li, X.M.; Kuang, H.X. Phenylpropanoids from the fruits of Nicandra physaloides and their anti-inflammatory activities. Nat. Prod. Res. 2017, 31, 2634–2640. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yin, X.; Sun, Y.; Liu, Y.; Lu, D.; Zhou, Y.; Adnan, M.A.; Pan, J.; Guan, W.; Yang, B.; et al. A new phenylpropanoid from the roots of Solanum melongena L. and evaluation of anti-inflammatory activities. Rec. Nat. Prod. 2021, 15, 261–266. [Google Scholar] [CrossRef]
- Kang, M.A.; Kim, M.B.; Kim, J.H.; Ko, Y.H.; Lim, S.B. Integral antioxidative capacity and antimicrobacterial activity of pressurized liquid extracts from 40 selected plant species. J. Korean Soc. Food Sci. Nutr. 2010, 39, 1249–1256. [Google Scholar] [CrossRef]
- Kim, K.-H.; Kim, A.R.; Cho, E.-J.; Joo, S.-J.; Park, J.-H.; Moon, J.-Y.; Yum, J.-H.; Kim, T.H.; Kwon, H.-J.; Lee, H.-T.; et al. Antibacterial activity of Rhus javanica against the fish pathogens Vibrio ichthyoenteri and Streptococcus iniae. Korean J. Fish Aquat. Sci. 2014, 47, 18–22. [Google Scholar]
- Lee, J.H. Anti-oxidant and anti-inflammatory effects of Diospytos kaki Tumb leaves extracts. Korean J. Aesthet. Cosmetol. 2014, 12, 719–724. [Google Scholar]
- Thi, N.N.; Song, H.S.; Oh, E.J.; Lee, Y.G.; Ko, J.H.; Kwon, J.E.; Kang, S.C.; Lee, D.Y.; Jung, I.H.; Baek, N.I. Phenylpropanoids from Lilium Asiatic hybrid flowers and their anti-inflammatory activities. Appl. Biol. Chem. 2017, 60, 517–533. [Google Scholar] [CrossRef]
- Han, J.M.; Lee, E.K.; Gong, S.Y.; Sohng, J.K.; Kang, Y.J.; Jung, H.J. Sparassis crispa exerts anti-inflammatory activity via suppression of TLR-mediated NF-κB and MAPK signaling pathways in LPS-induced RAW 264.7 macrophage cells. J. Ethnopharmacol. 2019, 231, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Laskin, D.L.; Pendino, K.J. Macrophges and inflammatory mediators in tissue injury. Annu. Rev. Toxicol. 1995, 35, 655–677. [Google Scholar] [CrossRef]
- Sharma, J.N.; Al-Omran, A.; Parvathy, S.S. Role of nitric oxide in inflammatory diseases. Inflammopharmacology 2007, 15, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Korns, D.; Frasch, S.C.; Fernandez-Boyanapalli, R.; Henson, P.M.; Bratton, D.L. Modulation of macrophage efferocytosis in inflammation. Front. Immunol. 2011, 2, 57. [Google Scholar] [CrossRef] [Green Version]
- Spilioti, E.; Holmbom, B.; Papavassiliou, A.G.; Moutsatsou, P. Lignans 7-hydroxymatairesinol and 7-hydroxymatairesinol 2 exhibit anti-inflammatory activity in human aortic endothelial cells. Mol. Nutr. Food Res. 2014, 58, 749–759. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.-M.; Kwon, O.-K.; Shin, I.-S.; Jeon, C.-M.; Shin, N.-R.; Lee, J.; Park, S.-H.; Bach, T.T.; Van Hai, D.; Oh, S.-R.; et al. Anti-inflammatory effects of methanol extract of Canarium lyi C.D. Dai & Yakovlev in RAW 264.7 macrophages and a murine model of lipopolysaccharide-induced lung injury. Int. J. Mol. Med. 2015, 35, 1403–1410. [Google Scholar] [PubMed] [Green Version]
- Kim, H.S.; Park, J.W.; Kwon, O.K.; Kim, J.H.; Oh, S.R.; Lee, H.K.; Bach, T.T.; Quang, B.H.; Ahn, K.S. Anti-inflammatory activity of a methanol extract from Ardisia tinctoria on mouse macrophages and paw edema. Mol. Med. Rep. 2014, 9, 1388–1394. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Kim, H.; Choi, H.; Jo, A.; Kang, H.; Yun, H.; Im, S.; Choi, C. Anti-inflammatory effects of a Stauntonia hexaphylla fruit extract in lipopolysaccharide-activated RAW-264.7 macrophages and rats by carrageenan-induced hind paw swelling. Nutrients 2018, 10, 110. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2017, 9, 7204–7218. [Google Scholar] [CrossRef] [Green Version]
- Ko, E.Y.; Cho, S.H.; Kang, K.; Kim, G.; Lee, J.H.; Jeon, Y.J.; Kim, D.K.; Ahn, G.; Kim, K.N. Anti-inflammatory activity of hydrosols from Tetragonia tetragonoides in LPS-induced RAW 264.7 cells. EXCLI J. 2017, 16, 521–530. [Google Scholar]
- Kim, K.-N.; Ko, Y.-J.; Yang, H.-M.; Ham, Y.-M.; Roh, S.W.; Jeon, Y.-J.; Ahn, G.; Kang, M.-C.; Yoon, W.-J.; Kim, D.; et al. Anti-inflammatory effect of essential oil and its constituents from fingered citron (Citrus medica L. var. sarcodactylis) through blocking JNK, ERK and NF-κB signaling pathways in LPS-activated RAW 264.7 cells. Food Chem. Toxicol. 2013, 57, 126–131. [Google Scholar]
- Barnes, P.J.; Karin, M. Nuclear factor-κB—A pivotal transcription factor in chronic inflammatory diseases. N. Engl. J. Med. 1997, 336, 1066–1071. [Google Scholar] [CrossRef]
- Karavtsova-Ivantsiv, Y.; Ciechanover, A. The ubiquitin-proteasome system and activation of NF-κB: Involvement of the ubiquitin ligase KPC1 in p105 processing and tumor suppression. Mol. Cell. Oncol. 2015, 2, e1054552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Liu, H.T. MAPK signal pathways in the regulation of cell proliferation in mammalian cells. Cell Res. 2002, 12, 9–18. [Google Scholar] [CrossRef]
- Ko, Y.J.; Ahn, G.; Ham, Y.M.; Song, S.M.; Ko, E.Y.; Cho, S.H.; Yoon, W.J.; Kim, K.N. Anti-inflammatory effect and mechanism of action of Lindera erythrocarpa essential oil in lipopolysaccharide-stimulated RAW 264.7 cells. EXCLI J. 2017, 16, 1103–1113. [Google Scholar]
- Beck, I.M.S.; Berghe, W.V.; Vermeulen, L.; Yamamoto, K.R.; Haegeman, G.; Bosscher, K.D. Crosstalk in inflammation: The interplay of glucocorticoid receptor-based mechanisms and kinases and phosphatases. Endocr. Rev. 2009, 30, 830–882. [Google Scholar] [CrossRef]
- Kim, Y.S.; Ahn, C.B.; Je, J.Y. Anti-inflammatory action of high molecular weight Mytilus edulis hydrolysates fraction in LPS-induced RAW 264.7 macrophage via NF-κB and MAPK pathways. Food Chem. 2016, 202, 9–14. [Google Scholar] [CrossRef]
- Cho, S.H.; Heo, S.J.; Yang, H.W.; Ko, E.Y.; Jung, M.S.; Cha, S.H.; Ahn, C.; Jeon, Y.J.; Kim, K.N. Protective effect of 3-bromo-4,5-dihydroxybenzaldehyde from Polysiphonia morrowii Harvey against hydrogen perxoxide-induced oxidative stress in vitro and in vivo. J. Microbiol. Biotechnol. 2019, 29, 1193–1203. [Google Scholar] [CrossRef] [PubMed]
- Rothenbücher, T.S.P.; Ledin, J.; Gibbs, D.; Engqvist, H.; Persson, C.; Hulsart-Billström, G. Zebrafish embryos as a replacement model for initial biocompatibility studies of biomaterials and drug delivery systems. Acta Biomater. 2019, 100, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.Y.; Kang, M.C.; Lee, J.H.; Kang, N.; Lee, W.; Oh, J.Y.; Yang, H.W.; Lee, J.S.; Jeon, Y.J. Protective effect of marine brown algal polyphenols against oxidative stressed zebrafish with high glucose. RSC Adv. 2015, 5, 25738–25746. [Google Scholar] [CrossRef]
- Lee, S.H. Developmental toxicity and anti-inflammatory effect of the soft coral Dendronephthya gigantea collected Jeju Island in zebrafish model. Fish Aquat. Sci. 2017, 20, 32. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Li, X.; Chu, Q.; Jia, R.; Chen, W.; Wang, Y.; Yu, X.; Zheng, X. Russula alutacea Fr. polysaccharide ameliorates inflammation in both RAW 264.7 and zebrafish (Danio rerio) larvae. J. Biol. Macromol. 2020, 145, 740–749. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Takagi, N.; Yuan, B.; Zhou, Y.; Si, N.; Wang, H.; Yang, J.; Wei, X.; Zhao, H.; Bian, B. The protection of indolealkylamines from LPS-induced inflammation in zebrafish. J. Ethnopharmacol. 2019, 243, 112122. [Google Scholar] [CrossRef]
- Cho, S.H.; Kim, H.S.; Lee, W.; Han, E.J.; Kim, S.Y.; Fernando, I.P.S.; Ahn, G.; Kim, K.N. Eckol from Ecklonia cava ameliorates TNF-α/IFN-γ-induced inflammatory responses via regulating MAPKs and NF-κB signaling pathway in HaCaT cells. Int. Immunopharmacol. 2020, 82, 106146. [Google Scholar] [CrossRef]
- Ham, Y.M.; Cho, S.H.; Song, S.M.; Yoon, S.A.; Lee, Y.B.; Kim, C.S.; Kwon, S.H.; Jeong, M.S.; Yoon, W.J.; Kim, K.N. Litsenolide A2: The major anti-inflammatory activity compound in Litsea japonica fruit. J. Funct. Foods 2017, 39, 168–174. [Google Scholar] [CrossRef]
- Ko, E.Y.; Cho, S.H.; Kwon, S.H.; Eom, C.Y.; Jeong, M.S.; Lee, W.; Kim, S.Y.; Heo, S.J.; Ahn, G.; Lee, K.P.; et al. The roles of NF-κB and ROS in regulation of pro-inflammatory mediators of inflammation induction in LPS-stimulated zebrafish embryos. Fish Shellfish. Immun. 2017, 68, 525–529. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, S.-H.; Park, S.; Jeong, H.; Kim, S.-R.; Jeong, M.S.; Choi, M.; Kim, S.H.; Kim, K.-N. Anti-Inflammatory Activity of 4-((1R,2R)-3-Hydroxy-1-(4-hydroxyphenyl)-1-methoxypropan-2-yl)-2-methoxyphenol Isolated from Juglans mandshurica Maxim. in LPS-Stimulated RAW 264.7 Macrophages and Zebrafish Larvae Model. Pharmaceuticals 2021, 14, 771. https://doi.org/10.3390/ph14080771
Cho S-H, Park S, Jeong H, Kim S-R, Jeong MS, Choi M, Kim SH, Kim K-N. Anti-Inflammatory Activity of 4-((1R,2R)-3-Hydroxy-1-(4-hydroxyphenyl)-1-methoxypropan-2-yl)-2-methoxyphenol Isolated from Juglans mandshurica Maxim. in LPS-Stimulated RAW 264.7 Macrophages and Zebrafish Larvae Model. Pharmaceuticals. 2021; 14(8):771. https://doi.org/10.3390/ph14080771
Chicago/Turabian StyleCho, Su-Hyeon, SeonJu Park, Hoibin Jeong, Song-Rae Kim, Myeong Seon Jeong, Miri Choi, Seung Hyun Kim, and Kil-Nam Kim. 2021. "Anti-Inflammatory Activity of 4-((1R,2R)-3-Hydroxy-1-(4-hydroxyphenyl)-1-methoxypropan-2-yl)-2-methoxyphenol Isolated from Juglans mandshurica Maxim. in LPS-Stimulated RAW 264.7 Macrophages and Zebrafish Larvae Model" Pharmaceuticals 14, no. 8: 771. https://doi.org/10.3390/ph14080771
APA StyleCho, S.-H., Park, S., Jeong, H., Kim, S.-R., Jeong, M. S., Choi, M., Kim, S. H., & Kim, K.-N. (2021). Anti-Inflammatory Activity of 4-((1R,2R)-3-Hydroxy-1-(4-hydroxyphenyl)-1-methoxypropan-2-yl)-2-methoxyphenol Isolated from Juglans mandshurica Maxim. in LPS-Stimulated RAW 264.7 Macrophages and Zebrafish Larvae Model. Pharmaceuticals, 14(8), 771. https://doi.org/10.3390/ph14080771