
The Effect of Cyanine Dye NK-4 on Photoreceptor Degeneration in a Rat Model of Early-Stage Retinitis Pigmentosa


Abstract

1. Introduction
2. Results
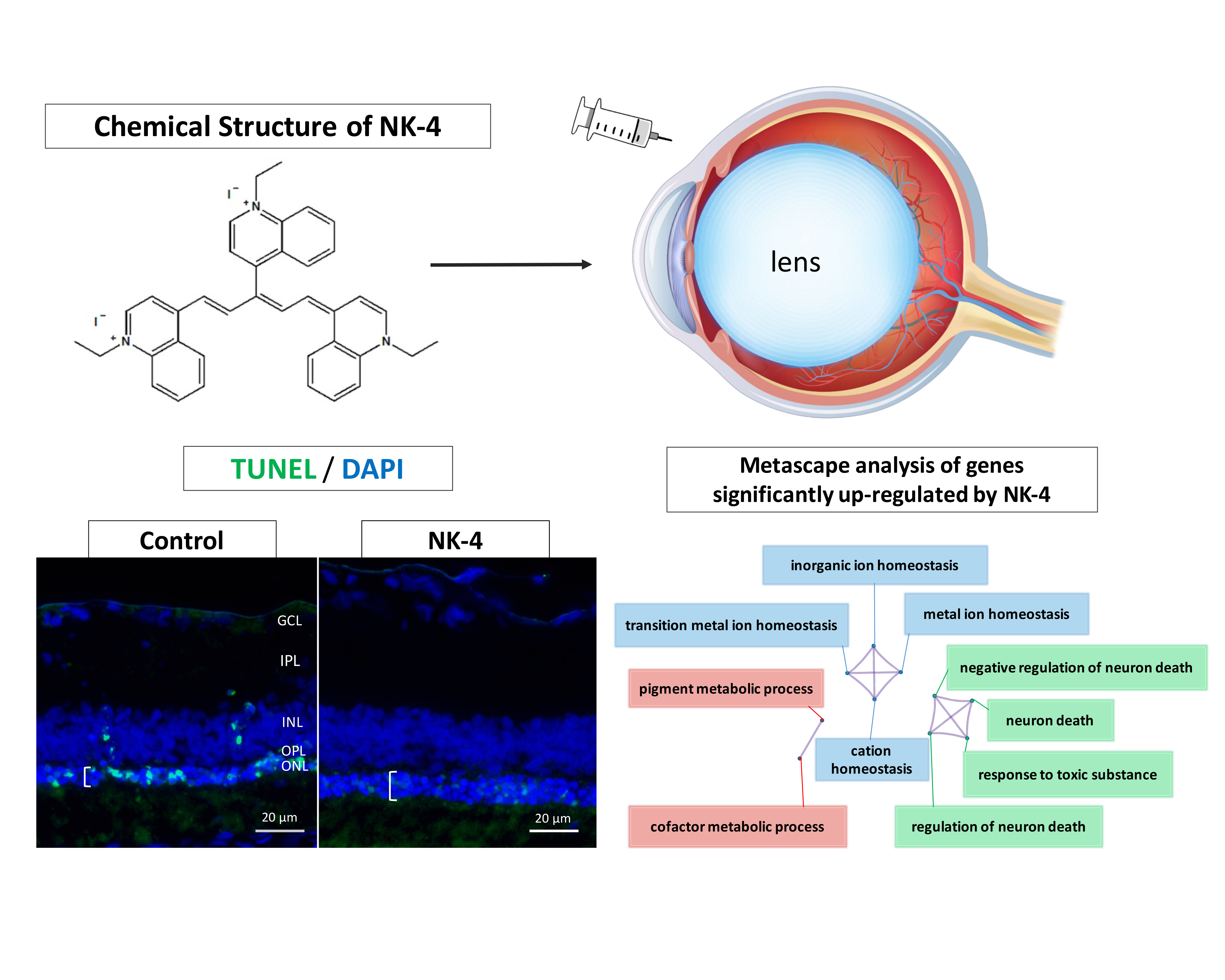
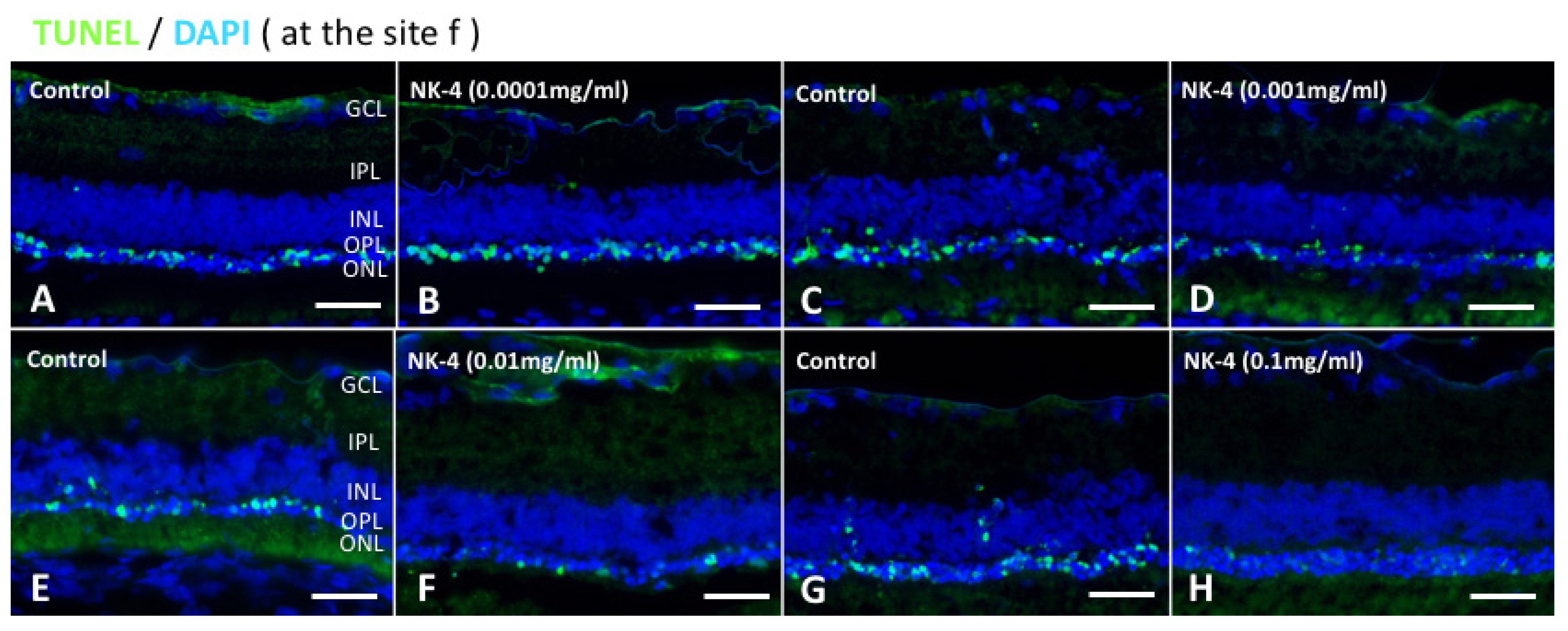
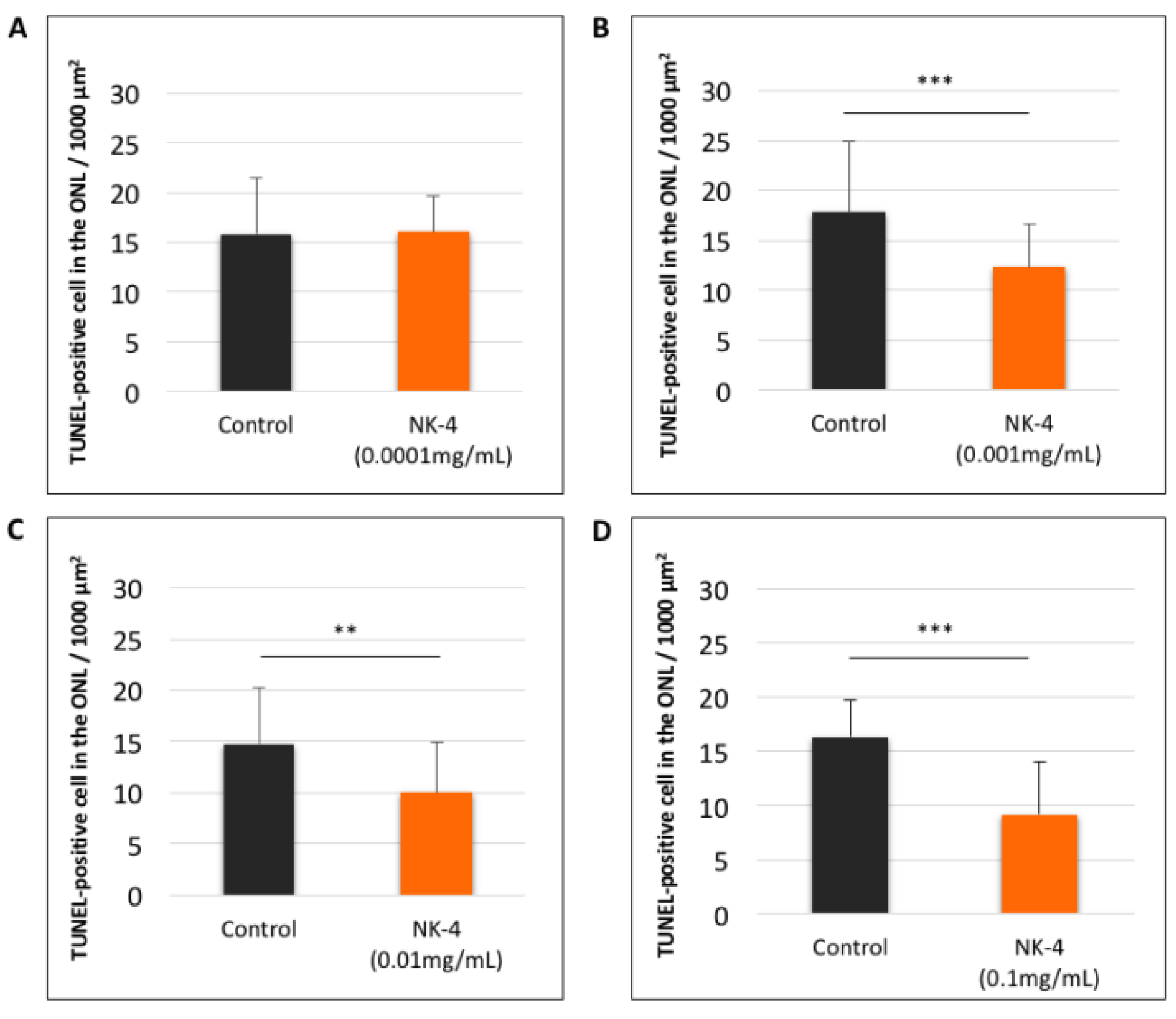
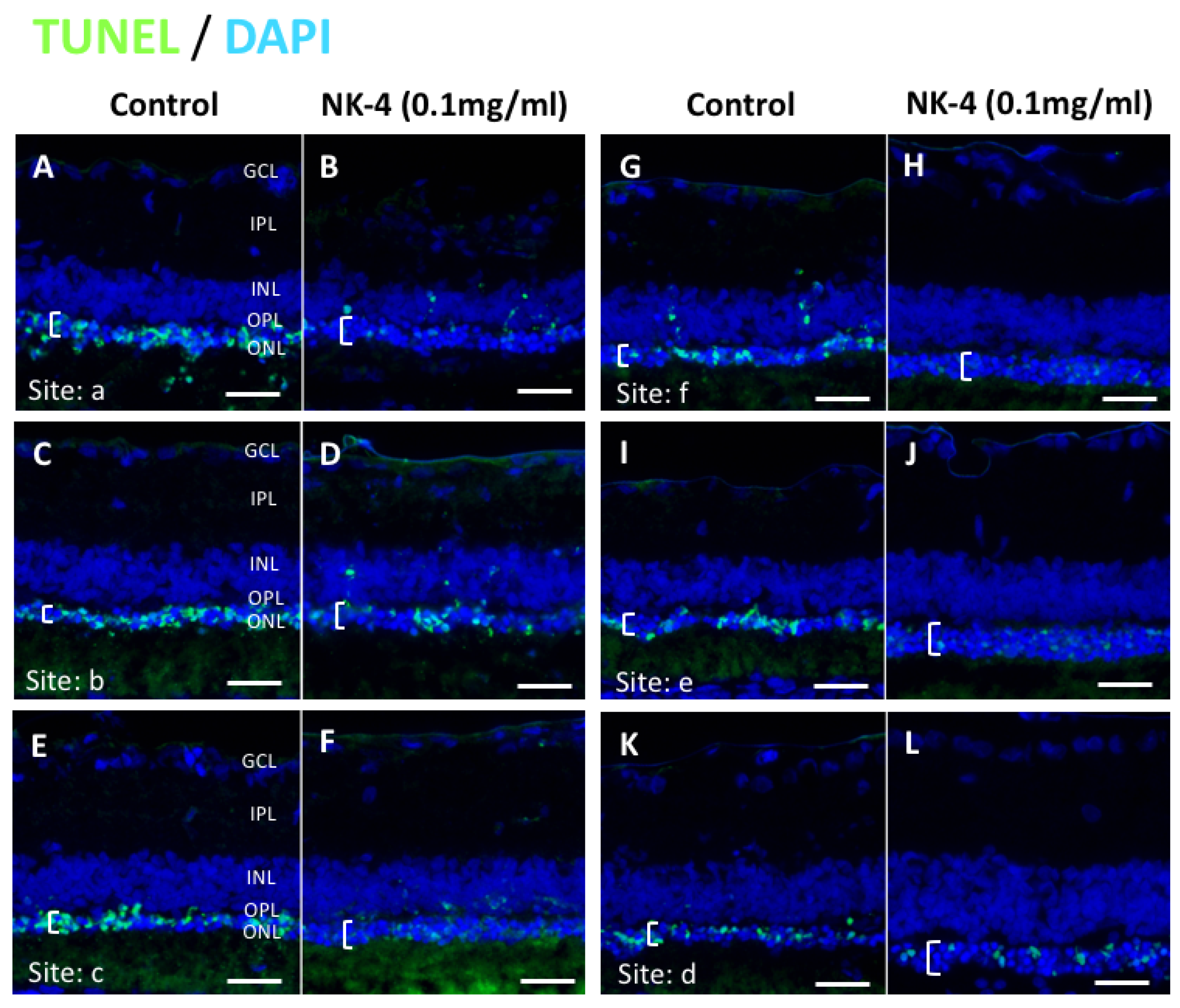
2.1. TUNEL Staining and ONL Thickness
2.2. Screening of Differentially Expressed Genes in The Eyes Injected with NK-4
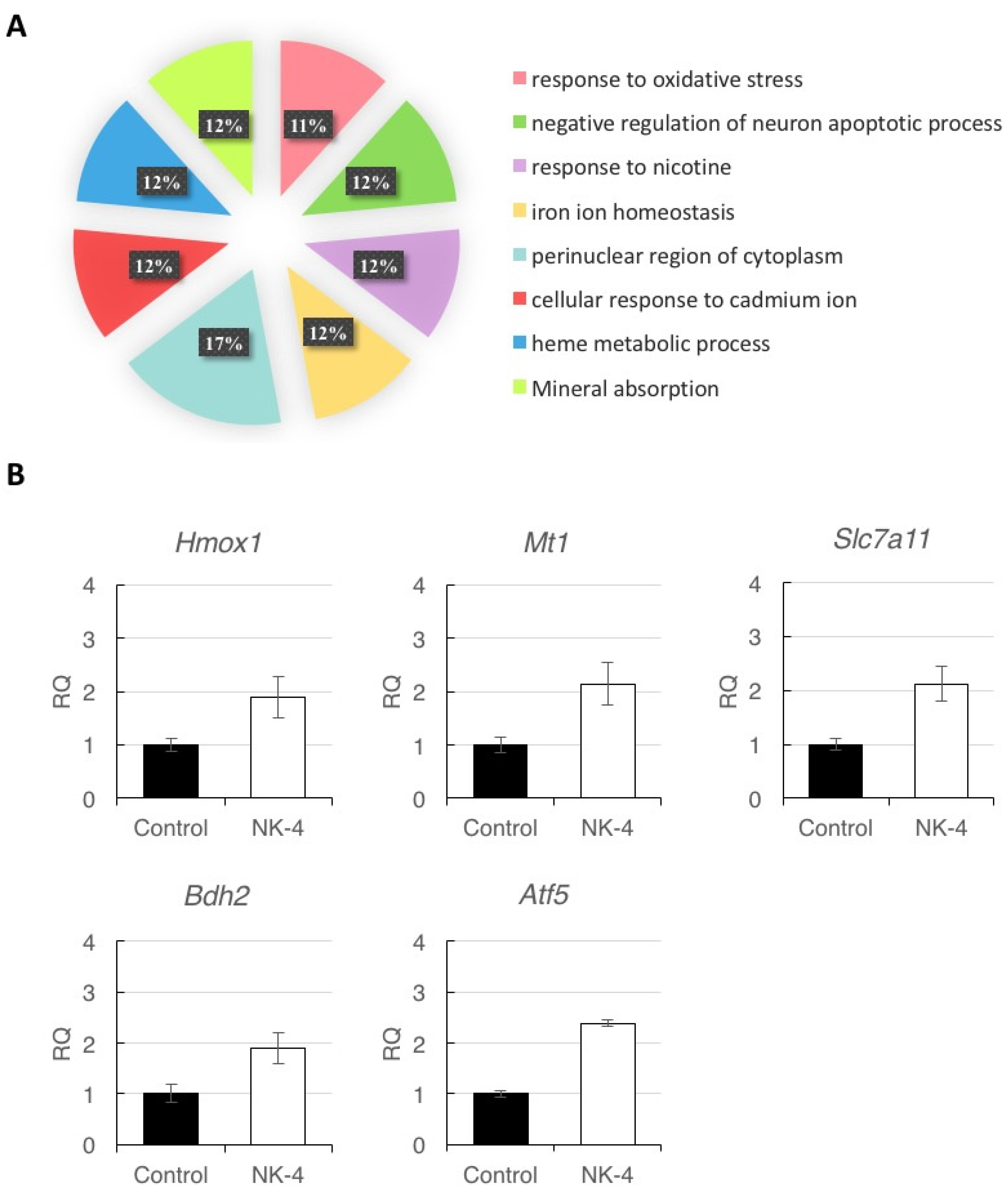
2.3. Bioinformatics Analysis of Differentially Expressed Genes in the Eyes Injected with NK-4
2.4. Validation of RNA-seq Data by RT-qPCR
3. Discussion
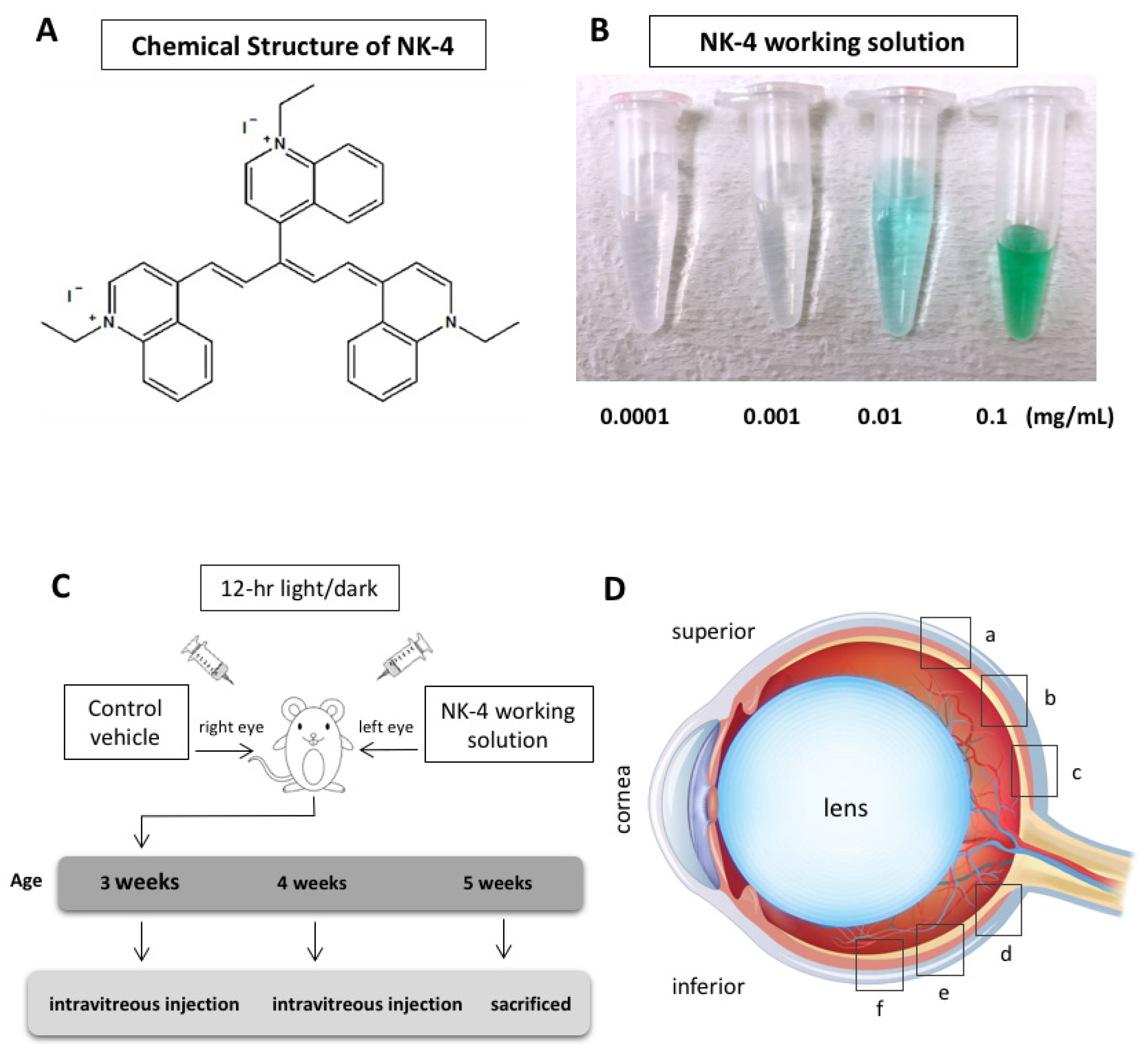
4. Materials and Methods
4.1. Chemicals and Preparations
4.2. Animals
4.3. TUNEL Staining
4.4. RNA Extraction
4.5. RNA Sequencing
4.6. Bioinformatics Analysis
4.7. Real-Time Quantitative PCR (RT-qPCR)
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nishiguchi, K.M.; Tearle, R.G.; Liu, Y.P.; Oh, E.C.; Miyake, N.; Benaglio, P.; Harper, S.; Koskiniemi-Kuendig, H.; Venturini, G.; Sharon, D.; et al. Whole genome sequencing in patients with retinitis pigmentosa reveals pathogenic DNA structural changes and NEK2 as a new disease gene. Proc. Natl. Acad. Sci. USA 2013, 110, 16139–16144. [Google Scholar] [CrossRef]
- Phelan, J.K.; Bok, D. A brief review of retinitis pigmentosa and the identified retinitis pigmentosa genes. Mol. Vis. 2000, 6, 116–124. [Google Scholar]
- Chen, J.; Poulaki, V.; Kim, S.-J.; Eldred, W.D.; Kane, S.; Gingerich, M.; Shire, D.B.; Jensen, R.; DeWalt, G.; Kaplan, H.J.; et al. Implantation and extraction of penetrating electrode arrays in minipig retinas. Transl. Vis. Sci. Technol. 2020, 9, 19. [Google Scholar] [CrossRef]
- Matsuo, T.; Uchida, T.; Sakurai, J.; Yamashita, K.; Matsuo, C.; Araki, T.; Yamashita, Y.; Kamikawa, K. Visual evoked potential recovery by subretinal implantation of photoelectric dye-coupled thin film retinal prosthesis in monkey eyes with macular degeneration. Artif. Organs 2018, 42, E186–E203. [Google Scholar] [CrossRef]
- Luo, Y.H.-L.; da Cruz, L. The Argus(®) II retinal prosthesis system. Prog. Retin Eye Res. 2016, 50, 89–107. [Google Scholar] [CrossRef]
- Ramsden, C.M.; Powner, M.B.; Carr, A.-J.; Smart, M.J.K.; da Cruz, L.; Coffey, P.J. Stem cells in retinal regeneration: Past, present and future. Development 2013, 140, 2576–2585. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Galicia, C.; Stenkamp, D.L. Transcripts within rod photoreceptors of the Zebrafish retina. BMC Genom. 2018, 19, 127. [Google Scholar] [CrossRef]
- Rezaie, T.; McKercher, S.R.; Kosaka, K.; Seki, M.; Wheeler, L.; Viswanath, V.; Chun, T.; Joshi, R.; Valencia, M.; Sasaki, S.; et al. Protective effect of carnosic acid, a pro-electrophilic compound, in models of oxi-dative stress and light-induced retinal degeneration. Invest. Ophthalmol. Vis. Sci. 2012, 53, 7847–7854. [Google Scholar] [CrossRef]
- Sanz, M.; Johnson, L.; Ahuja, S.; Ekström, P.; Romero, J.; van Veen, T. Significant photoreceptor rescue by treatment with a combination of antioxidants in an animal model for retinal degeneration. Neuroscience 2007, 145, 1120–1129. [Google Scholar] [CrossRef]
- Cehajic-Kapetanovic, J.; Xue, K.; de la Camara, C.M.-F.; Nanda, A.; Davies, A.; Wood, L.J.; Salvetti, A.P.; Fischer, M.D.; Aylward, J.W.; Barnard, A.R.; et al. Initial results from a first-in-human gene therapy trial on X-linked retinitis pigmentosa caused by mutations in RPGR. Nat. Med. 2020, 26, 354–359. [Google Scholar] [CrossRef]
- D’Cruz, P.M.; Yasumura, D.; Weir, J.; Matthes, M.T.; Abderrahim, H.; Lavail, M.M.; Vollrath, D. Mutation of the receptor tyrosine kinase gene Mertk in the retinal dystrophic RCS rat. Hum. Mol. Genet. 2000, 9, 645–651. [Google Scholar] [CrossRef] [PubMed]
- Nandrot, E.; Dufour, E.M.; Provost, A.C.; Péquignot, M.O.; Bonnel, S.; Gogat, K.; Marchant, D.; Rouillac, C.; de Condé, B.S.; Bihoreau, M.-T.; et al. Homozygous deletion in the coding sequence of the c-mer gene in RCS rats unravels general mechanisms of physiological cell adhesion and apoptosis. Neurobiol. Dis. 2000, 7, 586–599. [Google Scholar] [CrossRef] [PubMed]
- Newton, F.; Megaw, R. Mechanisms of photoreceptor death in retinitis pigmentosa. Genes 2020, 11, 1120. [Google Scholar] [CrossRef] [PubMed]
- Gal, A.; Li, Y.; Thompson, D.; Weir, J.; Orth, U.; Jacobson, S.; Apfelstedt-Sylla, E.; Vollrath, D. Mutations in Mertk, the human orthologue of the RCS rat retinal dystrophy gene, cause retinitis pigmentosa. Nat. Genet. 2000, 26, 270–271. [Google Scholar] [CrossRef]
- Al-Khersan, H.; Shah, K.P.; Jung, S.C.; Rodriguez, A.; Madduri, R.K.; Grassi, M.A. A novel MERTK mutation causing retinitis pigmentosa. Graefe’s Arch. Clin. Exp. Ophthalmol. 2017, 255, 1613–1619. [Google Scholar] [CrossRef] [PubMed]
- Portera-Cailliau, C.; Sung, C.H.; Nathans, J.; Adler, R. Apoptotic photoreceptor cell death in mouse models of retinitis pigmentosa. Proc. Natl. Acad. Sci. USA 1994, 91, 974–978. [Google Scholar] [CrossRef]
- Kohno, K.; Koya-Miyata, S.; Harashima, A.; Tsukuda, T.; Katakami, M.; Ariyasu, T.; Ushio, S.; Iwaki, K. Inflammatory M1-like macrophages polarized by NK-4 undergo enhanced phenotypic switching to an anti-inflammatory M2-like phenotype upon co-culture with apoptotic cells. Journal of Inflammation. J. Inflamm. 2021, 18, 2. [Google Scholar] [CrossRef]
- Kunikata, T.; Ishihara, T.; Ushio, S.; Iwaki, K.; Ikeda, M.; Kurimoto, M. Lumin, a cyanine dye, enhances interleukin 12-dependent inter-feron gamma production by lipopolysaccharide-stimulated mouse splenocytes. Biol. Pharm. Bull. 2002, 25, 1018–1021. [Google Scholar] [CrossRef]
- Sasaki, Y.; Nagai, N.; Okimura, T. Immunopharmacological actions of lumin. (II). Effect of lumin administration in NZB*NZW (B/W) F1 mice. Folia Pharmacol. Jpn. 1987, 89, 9–13. [Google Scholar] [CrossRef]
- Ushio, C.; Ariyasu, H.; Ariyasu, T.; Arai, S.; Ohta, T.; Fukuda, S. Suppressive effects of a cyanine dye against herpes simplex virus (HSV)-1 infection. Biomed. Res. 2009, 30, 365–368. [Google Scholar] [CrossRef][Green Version]
- Matsuo, T.; Liu, S.; Uchida, T.; Onoue, S.; Nakagawa, S.; Ishii, M.; Kanamitsu, K. Photoelectric dye, NK-5962, as a potential drug for preventing retinal neurons from apoptosis: Pharmacokinetic studies based on review of the evidence. Life 2021, 11, 591. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, Y.; Nagai, N.; Okimura, T.; Yamamoto, I. Immunopharmacological actions of lumin. (I). Anti-allergic actions of lumin. Folia Pharmacol. Jpn. 1987, 89, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kohno, K.; Koya-Miyata, S.; Harashima, A.; Ariyasu, T.; Ushio, S. NK-4 exerts selective regulatory effects on the activation and function of allergy-related Th2 cells. PLoS ONE 2018, 13, e0199666. [Google Scholar] [CrossRef] [PubMed]
- Koya-Miyata, S.; Ohta, H.; Akita, K.; Arai, S.; Ohta, T.; Kawata, T.; Fukuda, S. Cyanine dyes attenuate cerebral ischemia and reperfusion injury in rats. Biol. Pharm. Bull. 2010, 33, 1872–1877. [Google Scholar] [CrossRef][Green Version]
- Ohta, H.; Arai, S.; Akita, K.; Ohta, T.; Fukuda, S. Neurotrophic effects of a cyanine dye via the PI3K-Akt pathway: Attenuation of motor discoordination and neurodegeneration in an ataxic animal model. PLoS ONE 2011, 6, e17137. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Matsuo, T.; Hosoya, O. NK-4 dye reduces the apoptosis of photoreceptor cells. IOVS 2017, 58, 277. [Google Scholar]
- Liu, S.; Matsuo, T.; Miyaji, M.; Hosoya, O. Prevention of photoreceptor apoptosis by a Cryptocyanine drug (NK-4), in RCS rats. IOVS 2020, 61, 4945. [Google Scholar]
- Poss, K.D.; Tonegawa, S. Reduced stress defense in heme oxygenase 1-deficient cells. Proc. Natl. Acad. Sci. USA 1997, 94, 10925–10930. [Google Scholar] [CrossRef]
- Ito, Y.; Tanaka, H.; Hara, H. The potential roles of metallothionein as a therapeutic target for cerebral is-chemia and retinal diseases. Curr. Pharm. Biotechnol. 2013, 14, 400–407. [Google Scholar] [CrossRef]
- Lu, H.; Hunt, D.M.; Gantia, R.; Davisb, A.; Duttc, K.; Alamd, J.; Hunt, R.C. Metallothionein protects retinal pigment epithelial cells against apoptosis and oxidative stress. Exp. Eye Res. 2002, 74, 83–92. [Google Scholar] [CrossRef]
- Ananth, S.; Babu, E.; Veeranan-Karmegam, R.; Baldowski, B.R.B.; Boettger, T.; Martin, P.M. Induction of the cystine/glutamate exchanger SLC7A11 in retinal pigment epithelial cells by the antipsoriatic drug monomethylfumarate. Investig. Opthalmol. Vis. Sci. 2013, 54, 1592–1602. [Google Scholar] [CrossRef]
- Ananth, S.; Gnana-Prakasam, J.P.; Bhutia, Y.D.; Veeranan-Karmegam, R.; Martin, P.M.; Smith, S.B.; Ganapathy, V. Regulation of the cholesterol efflux transporters ABCA1 and ABCG1 in retina in hemochromatosis and by the endogenous siderophore 2,5-dihydroxybenzoic acid. Biochim. Biophys. Acta Bioenerg. 2014, 1842, 603–612. [Google Scholar] [CrossRef]
- Key, J.; Sen, N.E.; Arsović, A.; Krämer, S.; Hülse, R.; Khan, N.N.; Meierhofer, D.; Gispert, S.; Koepf, G.; Auburger, G. Systematic surveys of iron homeostasis mechanisms reveal ferritin superfamily and nucleotide surveillance regulation to be modified by PINK1 absence. Cells 2020, 9, 2229. [Google Scholar] [CrossRef]
- Torres-Peraza, J.F.; Engel, T.; Martín-Ibáñez, R.; Sanz-Rodríguez, A.; Fernández-Fernández, M.R.; Esgleas, M.; Canals, J.M.; Henshall, D.C.; Lucas, J.J. Protective neuronal induction of ATF5 in endoplasmic reticulum stress induced by status epilepticus. Brain 2013, 136, 1161–1176. [Google Scholar] [CrossRef] [PubMed]
- Ohta, H.; Akita, K.; Ohta, T. Effects of NK-4, a cyanine dye with antioxidant activities: Attenuation of neuronal deficits in animal models of oxidative stress-mediated brain ischemia and neurodegenerative diseases. Biol. Pharm. Bull. 2012, 35, 1831–1835. [Google Scholar]
- Dowling, J.E.; Sidman, R.L. Inherited Retinal Dystrophy in the Rat. J. Cell Biol. 1962, 14, 73–109. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.W.; Wubben, T.J.; Besirli, C.G. Photoreceptor metabolic reprogramming: Current understanding and therapeutic im-plications. Commun. Biol. 2021, 4, 245. [Google Scholar] [CrossRef]
- Eckhert, C.D. Elevated body zinc in rats with inherited retinal dystrophy. J. Hered. 1981, 72, 130. [Google Scholar] [CrossRef]
- Yefimova, M.G.; Jeanny, J.-C.; Keller, N.; Sergeant, C.; Guillonneau, X.; Beaumont, C.; Courtois, Y. Impaired retinal iron homeostasis associated with defective phagocytosis in Royal College of Surgeons rats. Investig. Ophthalmol. Vis. Sci. 2002, 43, 537–545. [Google Scholar]
- Battaglia, K.B.-; Organisciak, D.T.; Albert, A.D. RCS rat retinal rod outer segment membranes exhibit different cholesterol distributions than those of normal rats. Exp. Eye Res. 1994, 58, 293–300. [Google Scholar] [CrossRef]
- Sergeant, C.; Llabador, Y.; Devès, G.; Vesvres, M.; Simonoff, M.; Yefimova, M.; Courtois, Y.; Jeanny, J. Iron and other elements (Cu, Zn, Ca) contents in retina of rats during development and hereditary retinal degeneration. Nucl. Instruments Methods Phys. Res. Sect. B Beam Interactions Mater. Atoms 2001, 181, 533–538. [Google Scholar] [CrossRef]
- Komeima, K.; Rogers, B.S.; Campochiaro, P.A. Antioxidants slow photoreceptor cell death in mouse models of retinitis pig-mentosa. J. Cell Physiol. 2007, 213, 809–815. [Google Scholar] [CrossRef]
- Kang, K.; Tarchick, M.J.; Yu, X.; Beight, C.; Bu, P.; Yu, M. Carnosic acid slows photoreceptor degeneration in the Pde6b rd10 mouse model of retinitis pigmentosa. Sci. Rep. 2016, 6, 22632. [Google Scholar] [CrossRef]
- Faktorovich, E.G.; Steinberg, R.H.; Yasumura, D.; Matthes, M.T.; Lavail, M.M. Photoreceptor degeneration in inherited retinal dystrophy delayed by basic fibroblast growth factor. Nat. Cell Biol. 1990, 347, 83–86. [Google Scholar] [CrossRef]
- Corrochano, S.; Barhoum, R.; Boya, P.; Arroba, A.I.; Rodríguez-Muela, N.; Gómez-Vicente, V.; Bosch, F.; de Pablo, F.; de La Villa, P.; de La Rosa, E.J.; et al. Attenuation of vision loss and delay in apoptosis of photoreceptors induced by proinsulin in a mouse model of retinitis pigmentosa. Investig. Opthalmol. Vis. Sci. 2008, 49, 4188–4194. [Google Scholar] [CrossRef]
- Hartong, D.T.; Berson, E.L.; Dryja, T.P. Retinitis pigmentosa. Lancet 2006, 368, 1795–1809. [Google Scholar] [CrossRef]
- Berson, E.L.; Rosner, B.; Sandberg, M.A.; Weigel-DiFranco, C.; Willet, W.C. ω-3 intake and visual acuity in patients with retinitis pigmentosa receiving vitamin A. Arch. Ophthalmol. 2012, 130, 707–711. [Google Scholar] [CrossRef] [PubMed]
- Aleman, T.S.; Duncan, J.L.; Bieber, M.L.; de Castro, E.; Marks, D.A.; Gardner, L.M.; Steinberg, J.D.; Cideciyan, A.V.; Maguire, M.G.; Jacobson, S. Macular pigment and lutein supplementation in retinitis pigmentosa and Usher syndrome. Investig. Ophthalmol. Vis. Sci. 2001, 42, 1873–1881. [Google Scholar]
- Piano, I.; D’Antongiovanni, V.; Testai, L.; Calderone, V.; Gargini, C. A nutraceutical strategy to slowing down the progression of cone death in an animal model of retinitis pigmentosa. Front. Neurosci. 2019, 13, 461. [Google Scholar] [CrossRef]
- Tsuruma, K.; Yamauchi, M.; Inokuchi, Y.; Sugitani, S.; Shimazawa, M.; Hara, H. Role of oxidative stress in retinal photoreceptor cell death in N-methyl-N-nitrosourea–treated mice. J. Pharmacol. Sci. 2012, 118, 351–362. [Google Scholar] [CrossRef]
- Masuda, T.; Shimazawa, M.; Hara, H. Retinal diseases associated with oxidative stress and the effects of a free radical scavenger (edaravone). Oxidative Med. Cell. Longev. 2017, 2017, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Neeraj, K.G.; Sharad, M.; Tejram, S.; Abhinav, M.; Suresh, P.V.; Rajeev, K.T. Evaluation of anti–apoptotic activity of different dietary antioxidants in renal cell carcinoma against hydrogen peroxide. Asian Pac. J. Trop. Biomed. 2011, 1, 57–63. [Google Scholar] [CrossRef]
- Komeima, K.; Rogers, B.S.; Lu, L.; Campochiaro, P.A. Antioxidants reduce cone cell death in a model of retinitis pigmentosa. Proc. Natl. Acad. Sci. USA 2006, 103, 11300–11305. [Google Scholar] [CrossRef]
- Campochiaro, P.A.; Strauss, R.W.; Lu, L.; Hafiz, G.; Wolfson, Y.; Shah, S.M.; Sophie, R.; Mir, T.A.; Scholl, H.P. Is there excess oxidative stress and damage in eyes of patients with retinitis pigmentosa? Antioxidants Redox Signal. 2015, 23, 643–648. [Google Scholar] [CrossRef]
- Campochiaro, P.A.; Mir, T.A. The mechanism of cone cell death in retinitis pigmentosa. Prog. Retin. Eye Res. 2018, 62, 24–37. [Google Scholar] [CrossRef]
- Limoli, P.G.; Vingolo, E.M.; Limoli, C.; Nebbioso, M. Antioxidant and biological properties of mesenchymal cells used for therapy in retinitis pigmentosa. Antioxidants 2020, 9, 983. [Google Scholar] [CrossRef]
- Fan, B.; Tam, P.O.S.; Choy, K.W.; Wang, D.Y.; Lam, D.S.C.; Pang, C.P. Molecular diagnostics of genetic eye diseases. Clin. Biochem. 2006, 39, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Matsuo, T.; Hosoya, O.; Uchida, T. Photoelectric dye used for Okayama University-type retinal prosthesis reduces the apoptosis of photoreceptor cells. J. Ocul. Pharmacol. Ther. 2017, 33, 149–160. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Miyaji, M.; Furuta, R.; Hosoya, O.; Sano, K.; Hara, N.; Kuwano, R.; Kang, J.; Tateno, M.; Tsutsui, K.M.; Tsutsui, K. Topoisomerase IIβ targets DNA crossovers formed between distant homologous sites to modulate chromatin structure and gene expression. bioRxiv 2019, 484956. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Description | Refseq_ID | Locus | FPKM | LogFC | p Value | |
|---|---|---|---|---|---|---|---|
| Control | NK-4 | ||||||
| Hmox1 | Heme Oxygenase 1 | NM_012580 | chr19:13963122-13969964 | 4.36 | 7.60 | 0.802 | 5.75 × 10−3 |
| Mt1 | Metallothionein 1 | NM_138826 | chr19:11261630-11262647 | 58.30 | 114.76 | 0.977 | 1.00 × 10−4 |
| Slc7a11 | Solute carrier family 7 member 11 | NM_001107673 | chr2:139241141-139317101 | 12.74 | 21.53 | 0.756 | 5.00 × 10−5 |
| Bdh2 | 3-hydroxybutyrate dehydrogenase, type 2 | NM_001106473 | chr2:232765048-232785694 | 2.01 | 3.72 | 0.890 | 2.46 × 10−2 |
| Atf5 | Activating transcription factor 5 | NM_172336 | chr1:95284510-95286786 | 21.24 | 37.17 | 0.807 | 6.00 × 10−4 |
| Gene | Sequence (5’ -> 3’) Forward Primer | Sequence (5’ -> 3’) Reverse Primer |
|---|---|---|
| Hmox1 | cacgcatatacccgctacct | aaggcggtcttagcctcttc |
| Mt1 | acctcctgcaagaagagctg | aaactgggtggaggtgtacg |
| Scl7a11 | agcagtcccgatctttgttg | aacagctggcagaggagtgt |
| Bdh2 | aaccacagagaacggacctg | gacgatgactttcccttcca |
| Atf5 | agagggcagagtcagtggaa | ggaagtgaaatggagggaca |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; Matsuo, T.; Miyaji, M.; Hosoya, O. The Effect of Cyanine Dye NK-4 on Photoreceptor Degeneration in a Rat Model of Early-Stage Retinitis Pigmentosa. Pharmaceuticals 2021, 14, 694. https://doi.org/10.3390/ph14070694
Liu S, Matsuo T, Miyaji M, Hosoya O. The Effect of Cyanine Dye NK-4 on Photoreceptor Degeneration in a Rat Model of Early-Stage Retinitis Pigmentosa. Pharmaceuticals. 2021; 14(7):694. https://doi.org/10.3390/ph14070694
Chicago/Turabian StyleLiu, Shihui, Toshihiko Matsuo, Mary Miyaji, and Osamu Hosoya. 2021. "The Effect of Cyanine Dye NK-4 on Photoreceptor Degeneration in a Rat Model of Early-Stage Retinitis Pigmentosa" Pharmaceuticals 14, no. 7: 694. https://doi.org/10.3390/ph14070694
APA StyleLiu, S., Matsuo, T., Miyaji, M., & Hosoya, O. (2021). The Effect of Cyanine Dye NK-4 on Photoreceptor Degeneration in a Rat Model of Early-Stage Retinitis Pigmentosa. Pharmaceuticals, 14(7), 694. https://doi.org/10.3390/ph14070694