Abstract
Mollusks are the macroinvertebrates most commonly introduced into fresh water. In invaded reservoirs, alien mollusks form a large biomass due to their large size. Climate change, water level regulation, and anthropogenic impacts on the environment lead to the drying up of water bodies and the death of littoral macroinvertebrates. To assess the impact of invasive snail mass mortality on water quality, laboratory experiments on the snail tissue decomposition were performed, the potential release of nutrients into aquatic ecosystems was calculated, and the predicted concentrations of nutrients were verified by field studies. The laboratory experiment showed quick decomposition of the common river snail Viviparus viviparus tissues with release into the environment of ammonium and total phosphorus of 2.72 ± 0.14 mg and 0.10 ± 0.02 mg, respectively, per gram of decomposing tissue. The concentrations of ammonium, nitrates, and total phosphorus at the site of snail death reached 2.70 ± 0.10, 3.13 ± 0.38 and 0.30 ± 0.02 mg/L, respectively. This indicates local contamination of the Novosibirsk reservoir littoral with decomposition products. The aquatic management, water level regulation, and control of undesirable species should take into account the likelihood of water quality decreasing as a result of macroinvertebrate mass mortality.
1. Introduction
The development of water transport, aquaculture, increased trade in hydrobionts and various human-induced transformations of ecosystems have greatly contributed to the expansion of species beyond their natural habitats. After establishment, alien species often reach very high abundance in new locations, causing undesirable ecological and economic consequences. After an outbreak, alien species’ abundance and biomass often drop sharply and then stabilize at a lower level [1,2]. A rapid increase in invader abundance followed by its sharp decline occurs in aquatic and terrestrial species as well as in plants and animals [3]. The decrease in abundance can be caused by competition aggravation, diseases, and exhaustion of available resources or varying environmental factors [4,5,6]. In the case of rapidly changing habitat conditions, the decline in species abundance is accompanied by mass die-offs and the release of decomposition products into aquatic ecosystems. Decomposing tissues release toxicants (e.g., ammonium) into water, thus causing oppression and even mortality of particularly sensitive species [7].
Despite the high volumes of nutrients released into water during mollusk decomposition, only a few related investigations have been carried out. They reported on the ability of dead mollusks to be embedded in trophic networks of terrestrial ecosystems [8]. Previous studies also demonstrated a toxic effect of ammonium released due to mollusk decomposition on their larvae and other macroinvertebrates [7].
Common river snail Viviparus viviparus (Linnaeus, 1758) is a mollusk of European origin; in recent decades it has actively settled in the reservoirs of West Siberia. The biomass of V. viviparus in the Novosibirsk reservoir is much higher (up to 16,000 g/m2) than in its native habitat [9,10]. Peculiar seasonal migration of river snails contributes to the formation of extremely high biomass on the dried littoral of the Novosibirsk reservoir. In autumn, V. viviparus migrates to depth of 2–3 m and greater to overwinter; accordingly, their number is growing here [11]. Adverse hydrological conditions leading to a drop in water stages and soil drying induce its mass death in the coastal zone. The research conducted at the Novosibirsk reservoir in 2005 is evidence of a 90% mortality rate of V. viviparus in the drained littoral during spring freezing [12].
The present study is aimed at the assessment of consequences of alien mollusk die-offs in the Novosibirsk reservoir ecosystem. To determine the impact of snail mass mortality on water quality, (1) laboratory experiments to estimate the rate of snail tissue decomposition and to quantify the amount of nutrients released were conducted, (2) potential nutrient concentrations (in cases of mass die-off) were calculated, and (3) field studies were carried out to assess the number of dead individuals and to measure nutrient concentrations at sites of mass mortality events.
2. Materials and Methods
2.1. Study Area
The Novosibirsk reservoir was built in the upper section of the Ob River in 1957 (Figure 1). It is a seasonally regulated reservoir of approximately 200 km length, with a water area of 1070 km2 at a full reservoir level, a volume of 8.8 km3, and an average depth of 9 m [13]. The reservoir is used for power production, navigation, fishing, recreation, tourism, and drinking water supply.
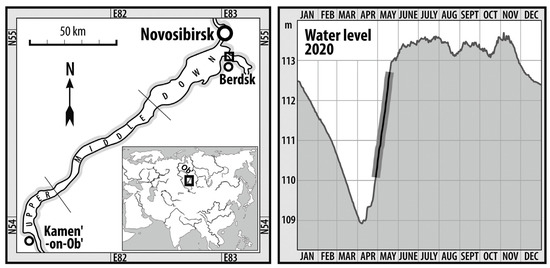
Figure 1.
Sampling location and changes in water level of the Novosibirsk reservoir in 2020. Changes in the water level during the field study are highlighted in bold.
The Novosibirsk reservoir is a mesotrophic-eutrophic reservoir with a characteristic longitudinal distribution of phytoplankton, the density of which is high in the upper part (chlorophyll a concentration of 20 mg/m3 in the surface water layer), drastically reduced in the middle part (less than 10 mg/m3) and increased in the lower part [14]. Maximum concentrations of chlorophyll a are registered in Berd Bay (more than 50 mg/m3) [14]. Hydrochemical indicators vary seasonally and depend on substance inflow from the Ob River. The average concentration of ammonium is within 0.1 mg/L, that of nitrates is within 0.3 mg/L, that of nitrites is within 0.01 mg/L, and that of phosphates is within 0.04 mg/L [13]. Waters of Berd Bay are distinguished by increased concentrations of ammonium and total phosphorus (on average, 0.17 and 0.06 mg/L in spring, respectively) [15]. Waters of the Novosibirsk reservoir are slightly alkaline (pH 7.3–8.6) with a favorable oxygen regime (5.1–12.0 mg/L) [16]. The TDS concentration reaches its maximum in winter (up to 350 mg/L), whereas it does not exceed 100–150 mg/L in spring [13].
One of the purposes of the reservoir is seasonal regulation of the Ob River flow. The water level in the reservoir falls during the autumn–winter period, with further draining of about 30% of its area. Filling of the reservoir begins in the middle of April and continues until the end of June. An ice-covered period in the reservoir usually lasts from early November to late April, while a snow cover at the catchment occurs from early November to mid-April. In April, air temperature can be negative (−7 °C on 18 April 2019) or positive (+30 °C on 23 April 2020) here. When sampling, air temperature varied from +20 to +28 °C, except for 6 May 2020 (+9 °C). During investigations, water temperature rose from +12 to +20 °C. Low-wind sunny weather was observed on 27 April 2020, 10 May, and 18 May 2020. Moderate wind was noted on 30 April 2020, whereas strong wind and waves were recorded on 6 May 2020.
2.2. Laboratory Experiment
The purpose of the laboratory experiment was to evaluate biogeochemical flows of matter formed due to mollusk tissue decomposition. Snails were collected on 6 June 2019, thoroughly treated to remove sediment and periphyton, and stored in a freezer at a temperature of −5 °C. Then, 50 g of mollusks with the addition of 600 mL of water (from the Novosibirsk reservoir) were placed in experimental containers of 0.95 L capacity and exposed to 21 °C. Reservoir water without snails was used as a control treatment. At 3, 7, 14, and 38 days after the start of the experiment, water was taken from three experimental and three control containers to analyze nutrient content. To quantify NH4, NO3, and Ptotal concentrations, the photometric method was used. The accredited laboratory of Verkhneobregionvodkhoz and the Chemical Analytical Center of the Institute for Water and Environmental Problems, SB RAS, performed the hydrochemical analysis of water. In this experiment, the amount of nutrients released per gram of mollusk decomposition was defined.
The Mann–Whitney U-test was used to assess the statistical significance of differences in nutrient concentration between control and experiment. Differences were considered significant at p < 0.05. Data analysis was performed using Statistica 6.0.
2.3. Field Study
In April–May 2020, field studies were arranged to assess the impact of the mass mortality of snails on the water quality of the Novosibirsk reservoir of Berd Bay. To undertake these studies, three sites, each 0.05 m2 in area, chosen as a result of a water level drop in the Novosibirsk reservoir of Berd Bay, were randomly selected. All common river snail individuals from each site were collected and measured. The mass of dead mollusks was calculated using the equation of body mass as a function of the shell height. The drawdown area covered with dead snails was measured on 27 April 2020. At the next survey of the reservoir (30 April 2020), all dead snails were already covered with water due to the reservoir filling.
During the reservoir filling, water was sampled to determine nutrient concentrations at the site with dead snails and at the control site. The latter was a place in the reservoir of Berd Bay with only live individuals. Water sampling was performed on 27 April 2020, 30 April 2020, 6 May 2020, 10 May 2020, and 18 May 2020 in the coastal zone; the depth of the reservoir at the water sampling site was 0.1 m. On 27 and 30 April 2020, samples were taken in areas where dead snails were found to be present. As the reservoir filled further, the area of the temporarily dry zone was decreased, and the sampling site was distanced from the zone with dead mollusks. By the end of the field research (18 May 2020), the site with dead mollusks was located at a depth of 1 m and at a distance of 40 m from the water sampling site.
In situ study results and calculations of potential nutrient concentrations in the areas of mass mortality were compared using the following formula:
where Cnutrient is the potential concentration of nutrients (mg/m3), Bdied is the biomass of recently dead mollusks (g/m2), Vnutrient is the amount of nutrients (mg) released during decomposition of 1 g of snails, and h is the total depth of reservoir above the decomposed mollusks (m). As the reservoir fills, the height of the water column above the mollusks increases, decomposition products are diluted with water, and nutrient concentrations decrease. When calculating the potential concentration of nutrients, it was assumed that all snails died simultaneously and decomposed completely.
3. Results
3.1. Abundance and Biomass of Dead Mollusks in the Dried Zone of the Reservoir
In April 2020, a mass mortality of V. viviparus was observed in an area of 25,000 m2 that dried up as a result of a water level drop (Figure S1). The density of dead mollusks was 2970 ± 280 ind./m2, whereas the calculated wet biomass with shells was 11,410 ± 1050 g/m2. Some of the large snails with shell heights of 32–38 mm, which managed to dig themselves into wet ground, survived. Live snails amounted to 850 ± 30 ind./m2. In dried soils, recently dead snails were found lying over alive ones; their shells contained decomposing soft tissue. The number of recently dead snails reached 430 ± 70 ind./m2, and their biomass was calculated as 1640 ± 350 g/m2. The surface layer contained empty mollusk shells (2540 ± 340 ind./m2).
3.2. Laboratory Experiment
Experiments with V. viviparus tissue decomposition suggested that the values of all analyzed indicators were higher for containers with snails than for the controls (Figure 2). Over the course of the experiment, the latter showed a fourfold decrease in ammonium concentrations, i.e., from 0.59 ± 0.02 mg/L to 0.14 ± 0.01 mg/L. In containers with snails, in the 3 days after starting the experiment, the ammonium concentration reached 59.3 ± 8.9 mg/L showing its maximum (226.7 ± 40.7 mg/L) on the 14th day. The concentration of NO3 in the control, on the contrary, was increased fivefold (from 0.23 ± 0.01 mg/L to 1.15 ± 0.05 mg/L). When the tissue decomposed, this trend (i.e., a slight increase throughout the experiment) was similar for control containers. Despite the similar trends, the NO3 concentration differed significantly between the control and experiment (Mann–Whitney U test, p = 0.002). Throughout the experiment, the concentration of phosphorus (Ptotal) in controls remained the same. In containers with snails, the maximum phosphorus concentrations were marked on day 14. After reaching its maximum, this indicator decreased insignificantly until the end of the experiment.
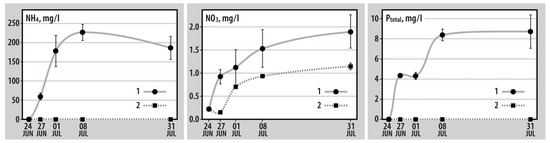
Figure 2.
Dynamics of NH4, NO3, and Ptotal concentrations (mg/L) in experimental (with decomposing snail tissues) (1) and in control containers (2).
According to the laboratory experiment, 1 g of decomposing tissue of V. viviparus released 2.72 ± 0.14 mg of ammonium and 0.10 ± 0.02 mg of total phosphorus. By the end of the experiment, only unbroken shells remained in containers; undecomposed mollusk bodies were not visually marked.
To calculate the impact on nutrient inflow to the reservoir (when water rises to the full reservoir level and fills the zone of temporary drainage with dead snails), the data on the dead mass of mollusks from the reservoir littoral were combined with the experimental data on decomposition-induced release of nutrients. Calculations of probable concentrations of NH4, NO3, and Ptotal in the reservoir in case of mass die-offs of snails showed that aquatic ecosystems can be exposed to extremely high concentrations of nutrients (Table 1). According to these calculations, only NO3 remained within the limits of water quality standards; its levels did not differ from that characteristic for Berd Bay of the Novosibirsk reservoir. All other indicators were demonstrated to be in excess of both national water quality standards and average values for the Novosibirsk reservoir. Thus, the high mortality of snails drastically deteriorates water quality in the littoral of the Novosibirsk reservoir. The nutrient inflow caused by mollusk decomposition may lead to a 110-fold increase in the Russian water quality standards for ammonium.

Table 1.
Nutrient concentrations in water of the Novosibirsk reservoir.
3.3. Field Data on Nutrient Concentrations
The in-situ studies of Berd Bay water revealed a significant increase in nutrient concentrations (as compared to background) at sites with dead snails. Maximum concentrations were observed at the beginning of the study in hot weather with little wind. On 27 April 2020, ammonium concentrations were 2.7 mg/L, thus exceeding the maximum allowable concentration (MAC) and background values by factors of 6.8 and 27, respectively (Table 1). Concentrations of nitrate and total phosphorus were eightfold and fivefold higher than background values, respectively. In 3 days, a twofold drop in ammonium concentrations was recorded. In 9 days, its content was within the MAC. During this period, the reservoir level rose by 179 cm, thereby contributing to the dilution of decomposition products. In addition, windy weather provided better water mixing and dilution of decomposition products. Over 3 weeks of observations, a 10-fold decrease in the concentrations of all nutrients, as well as a decline in most indicators to background values occurred. Only ammonium concentrations were twofold higher than background values. According to the field data, concentrations of ammonium, total phosphorus, and phosphates in the water over dead snails were much lower than predicted ones. This is probably due to the fact that in situ studies were performed when the water level had already risen by 100 cm, which greatly influenced the dilution of decomposition products and potentially led to peak concentrations being missed.
4. Discussion
Mollusks are long-lived organisms; they have a relatively large body and are able to form clusters with a biomass of several kilograms per square meter. Furthermore, numerous mollusk species are very sensitive to environmental impacts. Rising temperatures, reduced oxygen concentrations, reservoir pollution and extreme droughts can initiate mass die-offs [7,17]. In contrast to native mollusks, invaders are often less adapted to extremely varying environmental factors and are, thus, more exposed to high mortality [18]. In the last decade, mass die-offs of alien mollusks (i.e., Corbicula fluminea, Sinanodonta woodiana, Belamyia chinnensis, and Dreissena polymorpha) were registered in the USA, Canada, France, Portugal, Hungary, Switzerland, and the Netherlands [6].
4.1. Ecological Mechanisms of Population Decline
Factors able to regulate invader populations primarily include an abundance of natural enemies and competitors, interactions with later introduced invaders, human activities to control their number, environmental transformations caused by these species, and delayed genetic effects [2]. All of these factors have little impact in the form of reduction in V. viviparus abundance in the Novosibirsk reservoir. Recorded cases of local collapse of the V. viviparus population have been caused by cast ashore mollusks during severe storms, significant and long-term water level drop as a result of human regulation, and essential decreases in air temperature on the dried littoral [12]. In water bodies of the temperate zone, including the Novosibirsk reservoir, a decrease in reservoir storage occurs during winter that ensures macroinvertebrate survival until spring in wet soils under the ice during the ice-covered period. After spring snowmelt, mollusks find themselves on dried soils of the drawdown area. The ability of snails to bury into the ground, cover their shell mouth with operculum, and persist throughout unfavorable periods in moistened soils allows them to survive. Viviparidae are able to survive for several weeks without water [19,20]. The studied areas of the Novosibirsk reservoir remain dry for 2–3 months, which leads to the death of snails. Littoral drying is one of the most common causes of mass mortality of mollusks, especially in small rivers and lakes characterized by unstable hydrological regime, along with intermittent dry (rain-free) and hot weather.
The post-mortem decomposition of snail soft tissues occurs much faster than that of vertebrate skeletons and is accompanied by a release of nitrogen and phosphorus into the water column and interstitial space. Naturally, this creates local environmental problems. The decomposition rate determines the rate of inflow of nutrients and toxicants into the reservoir, thus representing an important indicator for predicting pollution. In this study, peak concentrations of nutrients and organic substances were noted 1–2 weeks after the beginning of the experiment. In experiments with fish, decomposition can last several months, whereas the decomposition of large mammals can last several years [21,22,23]. Such a rapid process initiated by massive die-offs of snails forms areas with extremely high concentrations of nutrients and organic substances. Therefore, high mortality is particularly dangerous for small and relatively isolated bays of the reservoir with a limited dilution of decomposition products.
4.2. Assessment of Nutrient Release
Decomposition can provide a release of toxic substances into the water, among which ammonium is particularly harmful. Toxic ammonium concentrations are different for various species [7]. Bivalves are most sensitive to ammonium; for example, the median lethal concentration (LC50) for juvenile freshwater clams is within 0.04–0.28 mg/L, that for crustaceans Hyalella azteca is within 0.19–6.09 mg/L, and that for rainbow trout Oncorhynchus mykiss is within 0.26–1.04 mg/L [24]. At the site of mass death of V. viviparus in the Novosibirsk reservoir, the concentration of ammonium reached 2.7 mg/L, which significantly exceeds both the tolerance limits of the most sensitive hydrobionts and the national water quality standards of the Russian Federation.
The NH4/NO3 ratio was significantly lower in the natural conditions of the Novosibirsk reservoir compared to the experiment. Apparently, this was due to the partial decomposition of the snail tissues in the air, in contrast to the laboratory experiment where the decomposition occurred completely in water. When decomposed in air, some of the gaseous decomposition products (including NH3) are blown with the wind, whereas, when decomposed in water, they dissolve. In addition, lower air temperatures at night may contribute to a slower mineralization of organic matter. The optimal temperature of ammonification is known to be significantly higher than that of nitrification [25]. Low night temperatures and colder temperatures in the spring of 2020 contributed to a decrease in the rate of mineralization of the snail tissues, while these temperature values were optimal for nitrification. Further studies of the influence of temperature on the decomposition rate are necessary for a more accurate assessment of the nitrogen release after mass mortality events.
Snail decomposition also greatly contributes to phosphorus increase in aquatic ecosystems. In freshwater ecosystems, algae are limited by low phosphorus concentrations [26]. V. viviparus have a reduced phosphorus excretion, which contributes to an increase in the N:P ratio in the water and a decrease in the amount of biologically available P, thus having a strong effect on the structure of algal communities [27]. In addition, river snails are able to obtain food by both grazing and filter-feeding [28]. The high biomass of filter-feeding snails, along with the low excretion of P, provides phosphorus retention and reduces the probability of the reservoir bloom. Mollusk die-offs can provide ecosystems with a large phosphorus concentration (up to 6.3 ± 1.3 mg Ptotal/L). The increased phosphorus release due to animal decomposition can cause reservoir bloom. An increase in phytoplankton density during mollusk tissue decomposition was registered both in laboratory experiments and in the natural reservoir [29]. In shallow and rapidly warming areas of the Novosibirsk reservoir (e.g., Ordynsky Bay), abundant algae and even their bloom can be observed as early as in June. Phosphorus arrival during mollusk tissue decomposition is among the causes of algae bloom in the reservoir.
Field studies confirmed that the mass death of snails has a strong impact on water quality of the Novosibirsk reservoir. In situ measurements of nutrient concentrations at sites of mass die-off of V. viviparus suggested much lower values than those calculated from experimental data. This may have been due to a stronger dilution of decomposition products under wind-wave action, the non-simultaneous death of snails, and the partial decomposition of snails in air before reservoir filling. During the research period, there was no extreme drop in air temperature able to cause the simultaneous death of most of the population. In the littoral of the reservoir, both empty mollusk shells and recently dead individuals at various stages of decay were found. This was indicative of gradual dying off as the soil dried and, accordingly, gradual nutrients release into water. The impact of extreme weather conditions, as well as an unfavorable combination of various factors, can result in local environmental disasters. The discrepancy between predicted values and field observations has also been noted when assessing the consequences of mass mortality of Corbicula fluminea in US rivers [18].
Unlike fish and mammalian skeletons, which serve as a relatively long source of nutrients in aquatic ecosystems [23], mollusk bodies provide a rapid release of nitrogen and phosphorus, thus resulting in a short-term but more profound effect. In this paper, only a short-term pulse of nutrients entering the ecosystems due to decomposition of soft tissues of dead snails was calculated. It should be noted that shells remain undecomposed in the aquatic environment over a long period, representing a long-term source of C, N and P for aquatic ecosystems [6,30].
The findings of this investigation hold significance in understanding the role of mass mortality of mollusks in aquatic and terrestrial trophic networks. It is common knowledge that dead animals can be included in food webs as food for scavengers. McDowell and Sousa [6] noted that soft tissues of dead Corbicula can be consumed by estuarine fish. However, there is little evidence of the use of dead snails by scavengers in aquatic ecosystems [6]. Beached shellfish are also considered as a trophic subsidy for terrestrial communities [31]. Dead mollusks can become involved in terrestrial trophic chains when consumed by terrestrial vertebrates such as crows, near-water birds, and rats [6,32]. The transfer of trophic resources beyond the habitat boundaries is an important process that ensures nutrient circulation in ecosystems and the interaction of aquatic and terrestrial ecosystems [32,33]. These studies suggest that the mass death of V. viviparus can also lead to an inflow of large amounts of pollutants into aquatic ecosystems, followed by a deterioration of water quality and ecosystem services. In the case of large volumes of dead mass appearing in trophic networks, aquatic and terrestrial scavengers do not have time for consumption due to high rates of decomposition of mollusk soft tissue. Keeping in mind the sedentary lifestyle of V. viviparus and their localization in certain areas of the reservoir bottom, their periodic cleaning and removal of invaders may have a positive effect on the ecosystem.
5. Conclusions
Mass die-offs among alien snails caused by adverse environmental conditions contribute to a rapid release of high concentrations of nutrients. When entering aquatic ecosystems, this can significantly affect the structure of communities, causing the mortality of particularly sensitive species and contributing to increased algae abundance. When managing a reservoir, regulating its level, and using it for water consumption purposes, one should keep in mind that water discharge can bring about the mass death of mollusks with high biomass and a drastic deterioration in water quality caused by nutrients released during tissue decomposition. In the near future, the number of extreme climatic events is predicted to increase. Thus, mass die-offs of hydrobionts will undoubtedly occur more frequently. As such, a further, more in-depth study of the effect of their decomposition products on aquatic ecosystems is required.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/d13080362/s1, Figure S1: Viviparus viviparus die-off in the temporary drained littoral of the Novosibirsk reservoir in the Berd Bay (27 April 2020).
Funding
This study was carried out as part of State Task (registration no. 121031200178-8).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data is included within the article.
Acknowledgments
The author would like to thank Anton V. Kotovshchikov and Laura M. Kipriyanova for their assistance with macroinvertebrate sampling.
Conflicts of Interest
The author declares no conflict of interest.
References
- Zaiko, A.; Minchin, D.; Olenin, S. “The day after tomorrow”: Anatomy of an ‘r’ strategist aquatic invasion. Aquat. Invasions 2014, 92, 145–155. [Google Scholar] [CrossRef]
- Strayer, D.L.; D’Antonio, C.M.; Essl, F.; Fowler, M.S.; Geist, J.; Hilt, S.; Jeschke, J.M. Boom-bust dynamics in biological invasions: Towards an improved application of the concept. Ecol. Lett. 2017, 20, 1337–1350. [Google Scholar] [CrossRef]
- Fey, S.B.; Siepielski, A.M.; Nusslé, S.; Cervantes-Yoshida, K.; Hwan, J.L.; Huber, E.R.; Carlson, S.M. Recent shifts in the occurrence, cause, and magnitude of animal mass mortality events. Proc. Natl. Acad. Sci. USA 2015, 112, 1083–1088. [Google Scholar] [CrossRef]
- Strayer, D.L.; Malcom, H.M. Long-term demography of a zebra mussel (Dreissena polymorpha) population. Freshw. Biol. 2006, 51, 117–130. [Google Scholar] [CrossRef]
- Moore, J.W.; Herbst, D.B.; Heady, W.N.; Carlson, S.M. Stream community and ecosystem responses to the boom and bust of an invading snail. Biol. Invasions 2012, 14, 2435–2446. [Google Scholar] [CrossRef]
- McDowell, W.G.; Sousa, R. Mass mortality events of invasive freshwater bivalves: Current understanding and potential directions for future research. Front. Ecol. Evol. 2019, 7, 331. [Google Scholar] [CrossRef]
- Cherry, D.S.; Scheller, J.L.; Cooper, N.L.; Bidwell, J.R. Potential effects of Asian clam (Corbicula fluminea) die-offs on native freshwater mussels (Unionidae) I: Water-column ammonia levels and ammonia toxicity. J. N. Am. Benthol. Soc. 2005, 24, 369–380. [Google Scholar] [CrossRef]
- Wesner, J.S.; Walters, D.M.; Zuellig, R.E. Pulsed salmonfly emergence and its potential contribution to terrestrial detrital pools. Food Webs 2019, 18, e00105. [Google Scholar] [CrossRef]
- Yanygina, L.V.; Vizer, A.M. Long-term dynamics and current distribution of the River snail (Viviparus viviparus) in the Novosibirsk reservoir. Tomsk. State Univ. J. Biol. 2020, 49, 149–165. [Google Scholar] [CrossRef]
- Yanygina, L.V.; Kotovshchikov, A.V.; Kipriyanova, L.M.; Volgina, D.D. Factors of spatial distribution and risk assessment of Viviparus viviparus L. invasion in aquatic ecosystems of the Ob River basin. Contemp. Probl. Ecol. 2020, 13, 162–171. [Google Scholar]
- Jakubik, B. Life strategies of Viviparidae (Gastropoda: Caenogastropoda: Architaenioglossa) in various aquatic habitats: Viviparus viviparus (Linnaeus, 1758) and V. contectus (Millet, 1813). Folia Malacol. 2012, 20, 145–179. [Google Scholar] [CrossRef][Green Version]
- Vizer, A.M. Zoobenthos of the drainage zone of Novosibirsk Reservoir. Contemp. Probl. Ecol. 2011, 4, 50–55. [Google Scholar] [CrossRef]
- Savkin, V.M.; Dvurechenskaya, S.Y.; Yermolaeva, N.I.; Kipriyanova, L.M.; Kirillov, V.V.; Romanov, R.E.; Popov, P.; Shlychkov, V.A.; Yanygina, L.V.; Atavin, A.A.; et al. Long-Term Dynamics of Water and Ecological Regime of Novosibirsk Reservoir; Publishing House of SB RAS: Novosibirsk, Russia, 2014. [Google Scholar]
- Kotovshchikov, A.V.; Yanygina, L.V. Spatial heterogeneity of chlorophyll a content in Novosibirsk reservoir. Bull. AB RGS 2018, 3, 46–52. [Google Scholar]
- Savkin, V.M.; Dvurechenskaya, S.Y. Influence of long-term complex use of water resources of the ecosystem of the Novosibirsk. Water Ecol. 2018, 73, 71–82. [Google Scholar] [CrossRef]
- Vasilyev, O.F.; Savkin, V.M.; Dvurechenskaya, S.Y. Ecological state of the Novosibirsk reservoir. Sibirskij Ehkologicheskij Zhurnal 2000, 2, 149–163. [Google Scholar]
- Sousa, R.; Ferreira, A.; Carvalho, F.; Lopes-Lima, M.; Varandas, S.; Teixeira, A. Die-offs of the endangered pearl mussel Margaritifera margaritifera during an extreme drought. Aquat. Conserv. 2018, 1–5. [Google Scholar] [CrossRef]
- McDowell, W.G.; McDowell, W.H.; Byers, J.E. Mass mortality of a dominant invasive species in response to an extreme climate event: Implications for ecosystem function. Limnol. Oceanogr. 2017, 62, 177–188. [Google Scholar] [CrossRef]
- Unstad, K.M.; Uden, D.R.; Allen, C.R.; Chaine, N.M.; Haak, D.M.; Kill, R.A.; Pope, K.L.; Stephen, B.J.; Wong, A. Survival and behavior of Chinese mystery snails (Bellamya chinensis) in response to simulated water body drawdowns and extended air exposure. Manag. Biol. Invasions 2013, 4, 123–127. [Google Scholar] [CrossRef]
- Havel, J.E.; Bruckerhoff, L.A.; Funkhouser, M.A.; Gemberling, A.R. Resistance to desiccation in aquatic invasive snails and implications for their overland dispersal. Hydrobiologia 2014, 741, 89–100. [Google Scholar] [CrossRef]
- Chidami, S.; Amyot, M. Fish decomposition in boreal lakes and biogeochemical implications. Limnol. Oceanogr. 2008, 53, 1988–1996. [Google Scholar] [CrossRef]
- Premke, K.; Fischer, P.; Hempel, M.; Rothhaupt, K.-O. Ecological studies on the decomposition rate of fish carcasses by benthic organisms in the littoral zone of Lake Constance, Germany. Ann. Limnol. 2010, 46, 157–168. [Google Scholar] [CrossRef]
- Nobre, R.L.G.; Carneiro, L.S.; Panek, S.E.; González, M.J.; Vanni, M.J. Fish, including their carcasses, are net nutrient sources to the water column of a eutrophic lake. Front. Ecol. Evol. 2019, 7. [Google Scholar] [CrossRef]
- Newton, T.J. The effects of ammonia on freshwater unionid mussels. Environ. Toxicol. Chem. 2003, 22, 2543–2544. [Google Scholar] [CrossRef] [PubMed]
- Thangarajan, R.; Bolan, N.S.; Naidu, R.; Surapaneni, A. Effects of temperature and amendments on nitrogen mineralization in selected Australian soils. Environ. Sci. Pollut. Res. 2013, 22, 8843–8854. [Google Scholar] [CrossRef]
- Vaughn, C.C.; Atkinson, C.L.; Julian, J.P. Drought-induced changes in flow regimes lead to long-term losses in mussel-provided ecosystem services. Ecol. Evol. 2015, 5, 1291–1305. [Google Scholar] [CrossRef] [PubMed]
- Johnson, P.T.J.; Olden, J.D.; Solomon, C.; Vander Zanden, M.J. Interactions among invaders: Community and ecosystem effects of multiple invasive species in an experimental aquatic system. Oecologia 2009, 159, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Höckelmann, C.; Pusch, M. The respiration and filter-feeding rates of the snail Viviparus viviparus (Gastropoda) under simulated stream conditions. Arch. Hydrobiol. 2000, 149, 553–568. [Google Scholar] [CrossRef]
- DuBose, T.P.; Atkinson, C.L.; Vaughn, C.C.; Golladay, S.W. Drought-induced, punctuated loss of freshwater mussels alters ecosystem function across temporal scales. Front. Ecol. Evol. 2019, 7, 274. [Google Scholar] [CrossRef]
- Ilarri, M.I.; Souza, A.T.; Sousa, R. Contrasting decay rates of freshwater bivalves’ shells: Aquatic versus terrestrial habitats. Limnologica 2015, 51, 8–14. [Google Scholar] [CrossRef]
- Bódis, E.; Tóth, B.; Sousa, R. Massive mortality of invasive bivalves as a potential resource subsidy for the adjacent terrestrial food web. Hydrobiologia 2014, 735, 253–262. [Google Scholar] [CrossRef]
- Schlacher, T.A.; Strydom, S.; Connolly, R.M. Multiple scavengers respond rapidly to pulsed carrion resources at the land–ocean interface. Acta Oecol. 2013, 48, 7–12. [Google Scholar] [CrossRef]
- Novais, A.; Souza, A.T.; Ilarri, M.; Pascoal, C.; Sousa, R. From water to land: How an invasive clam may function as a resource pulse to terrestrial invertebrates. Sci. Total. Environ. 2015, 538, 664–671. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).