


Searching for Innovative Functional Foods: Correlation Between Chemopreventive Potential and Bioactive Compounds Accumulation in Brassica Sprouts Grown Under Altered Gravity Conditions
 , , , and
, , , and
Abstract
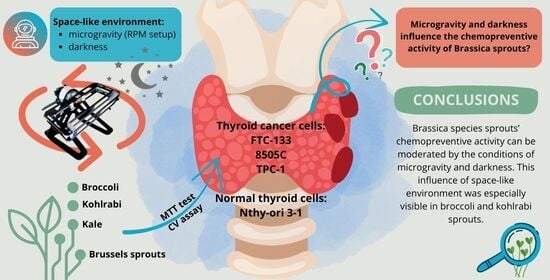
1. Introduction
2. Results and Discussion
2.1. Cytotoxic Activity Results
2.1.1. Cytotoxicity of Broccoli Sprouts
2.1.2. Cytotoxicity of Kale Sprouts
2.1.3. Cytotoxicity of Kohlrabi Sprouts
2.1.4. Cytotoxicity of the Sprouts of Brussels Sprouts
2.2. Antiproliferative Activity Results
2.2.1. Antiproliferative Activity of Broccoli Sprouts
2.2.2. Antiproliferative Activity of Kale Sprouts
2.2.3. Antiproliferative Activity of Kohlrabi Sprouts
2.2.4. Antiproliferative Activity of Brussels Sprouts’ Sprouts
2.3. Chemometric Analysis
3. Materials and Methods
3.1. Plant Material
3.2. Microgravity Setup
3.3. Extraction
3.4. Cell Cultures
3.5. Cytotoxic Activity
3.6. Antiproliferative Activity
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ferreira, P.M.P.; Rodrigues, L.A.R.L.; de Alencar Carnib, L.P.; de Lima Sousa, P.V.; Nolasco Lugo, L.M.; Nunes, N.M.F.; do Nascimento Silva, J.; da Silva Araûjo, L.; de Macêdo Gonçalves Frota, K. Cruciferous Vegetables as Antioxidative, Chemopreventive and Antineoplasic Functional Foods: Preclinical and Clinical Evidences of Sulforaphane Against Prostate Cancers. Curr. Pharm. Des. 2018, 24, 4779–4793. [Google Scholar] [CrossRef] [PubMed]
- Mitsiogianni, M.; Koutsidis, G.; Mavroudis, N.; Trafalis, D.T.; Botaitis, S.; Franco, R.; Zoumpourlis, V.; Amery, T.; Galanis, A.; Pappa, A.; et al. The Role of Isothiocyanates as Cancer Chemo-Preventive, Chemo-Therapeutic and Anti-Melanoma Agents. Antioxidants 2019, 8, 106. [Google Scholar] [CrossRef]
- Kumar, G.; Tuli, H.S.; Mittal, S.; Shandilya, J.K.; Tiwari, A.; Sandhu, S.S. Isothiocyanates: A Class of Bioactive Metabolites with Chemopreventive Potential. Tumor Biol. 2015, 36, 4005–4016. [Google Scholar] [CrossRef]
- Fiore, M.; Cristaldi, A.; Okatyeva, V.; Lo Bianco, S.; Oliveri Conti, G.; Zuccarello, P.; Copat, C.; Caltabiano, R.; Cannizzaro, M.; Ferrante, M. Dietary Habits and Thyroid Cancer Risk: A Hospital-Based Case–Control Study in Sicily (South Italy). Food Chem. Toxicol. 2020, 146, 111778. [Google Scholar] [CrossRef]
- World Health Organization. Data Platform. Available online: https://platform.who.int/mortality/themes/theme-details/topics/indicator-groups/indicator-group-details/MDB/thyroid-cancer (accessed on 5 August 2025).
- Paśko, P.; Zagrodzki, P.; Okoń, K.; Prochownik, E.; Krośniak, M.; Galanty, A. Broccoli Sprouts and Their Influence on Thyroid Function in Different In Vitro and In Vivo Models. Plants 2022, 11, 2750. [Google Scholar] [CrossRef]
- Paśko, P.; Galanty, A.; Zagrodzki, P.; Żmudzki, P.; Bieniek, U.; Prochownik, E.; Domínguez-Álvarez, E.; Bierła, K.; Łobiński, R.; Szpunar, J.; et al. Varied effect of fortification of kale sprouts with novel organic selenium compounds on the synthesis of sulphur and phenolic compounds in relation to cytotoxic, antioxidant and anti-inflammatory activity. Microchem. J. 2022, 179, 107509. [Google Scholar] [CrossRef]
- Markiewicz, M.; Galanty, A.; Kołodziejczyk, A.; Żmudzki, P.; Prochownik, E.; Zagrodzki, P.; Paśko, P. Bioactive compounds accumulation in Brassica sprouts grown under microgravity and darkness: A novel approach to functional foods. Food Chem. 2025, 491, 145324. [Google Scholar] [CrossRef]
- Grudzińska, M.; Galanty, A.; Prochownik, E.; Kołodziejczyk, A.; Paśko, P. Can Simulated Microgravity and Darkness Conditions Influence the Phytochemical Content and Bioactivity of the Sprouts?—A Preliminary Study on Selected Fabaceae Species. Plants 2024, 13, 1515. [Google Scholar] [CrossRef]
- Nakajima, S.; Ogawa, Y.; Suzuki, T.; Kondo, N. Enhanced antioxidant activity in mung bean seedlings grown under slow clinorotation. Microgravity Sci. Technol. 2019, 31, 395–401. [Google Scholar] [CrossRef]
- Vale, A.P.; Santos, J.; Brito, N.V.; Peixoto, V.; Carvalho, R.; Rosa, E.; Oliveira, M.B.P.P. Light influence in the nutritional composition of Brassica oleracea sprouts. Food Chem. 2015, 178, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Ishida, M.; Hara, H.; Fukino, N.; Kakizaki, T.; Morimitsu, Y. Glucosinolate metabolism, functionality and breeding for the improvement of Brassicaceae vegetables. Breed. Sci. 2014, 64, 48–59. [Google Scholar] [CrossRef]
- Abellan, A.; Domínguez-Perles, R.; Moreno, D.A.; García-Viguera, C. Sorting out the value of cruciferous sprouts as sources of bioactive compounds for nutrition and health. Nutrients 2019, 11, 429. [Google Scholar] [CrossRef]
- Massart, C.; Poirier, C.; Fergelot, P.; Fardel, O.; Gibassier, J. Effect of sodium butyrate on doxorubicin resistance and expression of multidrug resistance genes in thyroid carcinoma cells. Anti-Cancer Drugs 2005, 16, 255–261. [Google Scholar] [CrossRef]
- Su, X.; Liu, J.; Zhang, H.; Gu, Q.; Zhou, X.; Ji, M.; Yao, D. Lenvatinib promotes the antitumor effect of doxorubicin in anaplastic thyroid cancer. OncoTargets Ther. 2020, 13, 11183–11192. [Google Scholar] [CrossRef]
- Tadi, K.; Chang, Y.; Ashok, B.T.; Chen, Y.; Moscatello, A.; Schaefer, S.D.; Schantz, S.P.; Policastro, A.J.; Geliebter, J.; Tiwari, R.K. 3,3′-Diindolylmethane, a cruciferous vegetable derived synthetic anti-proliferative compound in thyroid disease. Biochem. Biophys. Res. Commun. 2005, 337, 1019–1025. [Google Scholar] [CrossRef] [PubMed]
- De Groot, A.P.; Willems, M.I.; De Vos, R.H. Effects of High Levels of Brussels Sprouts in the Diet of Rats. Food Chem. Toxicol. 1991, 29, 829–837. [Google Scholar] [CrossRef] [PubMed]
- Mcmillan, M.; Spinks, E.A.; Fenwick, G.R. Preliminary Observations on the Effect of Dietary Brussels Sprouts on Thyroid Function. Hum. Toxicol. 1986, 5, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, A.; Sharma, U.; Pal Vig, A.; Singh, B.; Arora, S. Free radical scavenging, antiproliferative activities and profiling of variations in the level of phytochemicals in different parts of broccoli (Brassica oleracea italica). Food Chem. 2014, 148, 373–380. [Google Scholar] [CrossRef]
- Le, T.N.; Luong, H.Q.; Li, H.-P.; Chiu, C.-H.; Hsieh, P.-C. Broccoli (Brassica oleracea L. var. italica) Sprouts as the Potential Food Source for Bioactive Properties: A Comprehensive Study on In Vitro Disease Models. Foods 2019, 8, 532. [Google Scholar] [CrossRef]
- Li, Z.; Di, H.; Cheng, W.; Ren, G.; Zhang, Y.; Ma, J.; Ma, W.; Yang, J.; Lian, H.; Li, X.; et al. Effect of the number of dark days and planting density on the health-promoting phytochemicals and antioxidant capacity of mustard (Brassica juncea) sprouts. Plants 2022, 11, 2515. [Google Scholar] [CrossRef]
- Lim, I.; Kang, M.; Kim, B.C.; Ha, J. Metabolomic and transcriptomic changes in mung bean (Vigna radiata (L.) R. Wilczek) sprouts under salinity stress. Front. Plant Sci. 2022, 13, 1030677. [Google Scholar] [CrossRef] [PubMed]
- Sołtys, A.; Galanty, A.; Grabowska, K.; Paśko, P.; Zagrodzki, P.; Podolak, I. Multidirectional Effects of Terpenoids from Sorbus intermedia (EHRH.) PERS Fruits in Cellular Model of Benign Prostate Hyperplasia. Pharmaceuticals 2023, 16, 965. [Google Scholar] [CrossRef] [PubMed]
- Westerhuis, J.; Kourti, T.; Macgregor, J. Analysis of multiblock and hierarchical PCA and PLS models. J. Chemom. 1998, 12, 301–321. [Google Scholar] [CrossRef]
- Janne, K.; Pettersen, J.; Lindberg, N.O.; Lundstedt, T. Hierarchical principal component analysis (PCA) and projection to latent structure (PLS) technique on spectroscopic data as a data pretreatment for calibration. J. Chemom. 2001, 15, 203–213. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 5BL | 6BL | 7BL | 5BLM | 6BLM | 7BLM | 5BD | 6BD | 7BD | 5BDM | 6BDM | 7BDM | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| FTC-133 | ||||||||||||
| 24 h | 113.51 ± 1.29 | 79.65 ± 6.21 | 82.81 ± 7.11 | 90.06 ± 6.3 | 94.44 ± 7.86 | 80.27 ± 3.38 | 83.83 ± 3.53 * | 68.64 ± 8.56 | 71.70 ± 9.65 | 67.72 ± 11.10 * | 94.75 ± 14.81 | 72.82 ± 6.63 |
| 48 h | 98.48 ± 6.02 | 61.19 ± 8.82 | 73.88 ± 6.24 | 71.54 ± 5.06 * | 75.28 ± 9.12 | 88.94 ± 8.96 | 86.67 ± 11.00 | 70.24 ± 5.06 | 72.36 ± 7.35 | 94.20 ± 4.35 | 86.61 ± 5.05 * | 91.46 ± 8.62 |
| 72 h | 40.84 ± 0.48 | 31.62 ± 4.10 | 35.20 ± 5.48 | 68.13 ± 7.13 * | 59.43 ± 4.56 * | 35.34 ± 3.73 | 27.20 ± 3.50 * | 25.31 ± 2.16 | 28.82 ± 5.22 | 32.61 ± 4.78 | 39.57 ± 6.76 | 26.77 ± 1.76 |
| 8505C | ||||||||||||
| 24 h | 88.93 ± 2.22 | 91.28 ± 6.35 | 84.35 ± 3.42 | 90.10 ± 7.82 | 89.25 ± 4.52 | 83.97 ± 9.73 | 80.40 ± 7.10 | 92.66 ± 7.16 | 84.99 ± 3.79 | 86.91 ± 6.44 | 93.41 ± 7.23 | 89.73 ± 1.13 |
| 48 h | 112.72 ± 4.09 | 96.98 ± 12.20 | 78.23 ± 4.00 | 84.14 ± 8.92 * | 81.31 ± 2.70 | 87.99 ± 6.52 | 82.47 ± 3.06 * | 87.48 ± 6.48 | 83.75 ± 6.48 | 87.09 ± 5.39 * | 86.71 ± 7.58 | 105.97 ± 5.34 * |
| 72 h | 64.35 ± 9.62 | 68.43 ± 7.07 | 51.55 ± 4.00 | 61.03 ± 9.89 | 42.60 ± 2.24 * | 43.96 ± 6.26 | 51.96 ± 1.64 | 54.61 ± 4.71 | 52.11 ± 2.95 | 61.18 ± 3.12 | 58.69 ± 6.50 | 66.92 ± 9.06 |
| TPC-1 | ||||||||||||
| 24 h | 91.68 ± 1.84 | 75.02 ± 10.00 | 62.53 ± 2.27 | 92.79 ± 10.30 | 84.50 ± 8.60 | 77.35 ± 9.39 | 63.79 ± 3.07 * | 65.40 ± 8.12 | 74.08 ± 7.78 | 76.29 ± 2.14 | 85.77 ± 1.06 | 91.65 ± 8.45 * |
| 48 h | 62.16 ± 6.86 | 47.30 ± 2.59 | 41.74 ± 2.33 | 92.93 ± 6.54 * | 85.60 ± 6.02 * | 71.80 ± 2.03 * | 44.76 ± 7.34 | 32.26 ± 5.02 | 44.93 ± 4.78 | 46.01 ± 7.31 | 53.47 ± 11.36 | 61.71 ± 6.56 * |
| 72 h | 33.88 ± 3.41 | 32.99 ± 2.3 | 26.13 ± 2.30 | 54.55 ± 6.65 * | 53.40 ± 3.58 * | 53.77 ± 2.27 * | 12.82 ± 0.31 * | 36.62 ± 5.17 | 41.97 ± 3.71 * | 41.60 ± 8.75 | 50.85 ± 1.97 * | 42.36 ± 4.26 * |
| Nthy-ori 3-1 | ||||||||||||
| 24 h | 55.09 ± 8.29 | 46.99 ± 1.82 | 48.11 ± 2.19 | 40.46 ± 6.01 | 34.45 ± 5.71 | 37.04 ± 6.54 | 35.49 ± 7.88 * | 26.21 ± 2.24 * | 28.21 ± 5.56 * | 28.88 ± 3.69 * | 32.96 ± 7.52 | 31.85 ± 5.40 |
| 48 h | 31.36 ± 0.50 | 35.71 ± 4.70 | 34.22 ± 1.78 | 35.87 ± 6.42 | 36.81 ± 5.76 | 33.83 ± 6.12 | 33.20 ± 5.66 | 33.20 ± 4.51 | 34.69 ± 3.37 | 34.93 ± 3.53 | 33.83 ± 4.69 | 38.21 ± 2.83 |
| 72 h | 29.83 ± 3.77 | 29.23 ± 4.11 | 30.53 ± 0.80 | 29.67 ± 5.42 | 25.67 ± 3.24 | 27.40 ± 1.17 | 24.70 ± 1.21 | 23.62 ± 1.75 | 24.49 ± 1.55 | 25.78 ± 0.67 | 25.00 ± 3.75 | 24.81 ± 1.80 |
| 5KaL | 6KaL | 7KaL | 5KaLM | 6KaLM | 7KaLM | 5KaD | 6KaD | 7KaD | 5KaDM | 6KaDM | 7KaDM | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| FTC-133 | ||||||||||||
| 24 h | 85.21 ± 6.71 | 83.68 ± 7.14 | 73.53 ± 8.29 | 76.08 ± 9.66 | 80.37 ± 1.57 | 74.35 ± 0.81 | 93.32 ± 1.73 | 83.22 ± 9.79 | 73.43 ± 14.18 | 77.21 ± 11.30 | 83.32 ± 9.40 | 67.31 ± 10.20 |
| 48 h | 60.65 ± 5.30 | 62.28 ± 6.46 | 51.27 ± 5.60 | 62.49 ± 4.61 | 60.65 ± 6.55 | 56.64 ± 2.68 | 66.99 ± 8.40 | 54.96 ± 5.29 | 53.93 ± 3.61 | 62.66 ± 7.45 | 56.26 ± 3.36 | 56.86 ± 1.88 |
| 72 h | 29.29 ± 6.86 | 33.22 ± 3.48 | 24.59 ± 0.66 | 26.66 ± 4.83 | 24.021 ± 3.16 | 22.61 ± 6.14 | 26.96 ± 3.59 | 18.56 ± 2.91 * | 19.05 ± 3.39 | 24.51 ± 6.52 | 22.94 ± 5.26 | 19.53 ± 0.40 |
| 8505C | ||||||||||||
| 24 h | 112.07 ± 3.20 | 104.93 ± 4.73 | 103.81 ± 13.80 | 115.48 ± 4.07 | 101.73 ± 0.32 | 98.10 ± 7.98 | 99.59 ± 9.69 | 95.22 ± 0.18 | 100.23 ± 2.44 | 97.57 ± 13.60 | 113.56 ± 0.90 | 104.77 ± 4.30 |
| 48 h | 117.53 ± 0.54 | 107.77 ± 2.35 | 109.31 ± 5.89 | 109.06 ± 8.83 | 101.86 ± 6.36 | 91.20 ± 2.35 | 97.62 ± 3.86 * | 101.73 ± 8.09 | 104.30 ± 4.88 | 102.63 ± 4.05 | 106.74 ± 13.70 | 124.66 ± 1.91 |
| 72 h | 85.73 ± 8.09 | 88.14 ± 4.31 | 77.72 ± 5.36 | 71.15 ± 5.49 | 67.07 ± 2.56 * | 54.46 ± 4.53 * | 69.11 ± 9.05 | 78.40 ± 4.90 | 66.77 ± 4.28 | 79.23 ± 7.72 | 79.31 ± 6.17 | 79.53 ± 7.16 |
| TPC-1 | ||||||||||||
| 24 h | 69.61 ± 1.95 | 67.73 ± 5.41 | 62.93 ± 2.62 | 58.32 ± 3.48 | 53.97 ± 4.21 * | 35.17 ± 4.68 * | 71.27 ± 2.23 | 70.61 ± 4.65 | 55.41 ± 1.56 | 73.48 ± 3.94 | 70.14 ± 7.09 | 76.22 ± 8.05 * |
| 48 h | 38.32 ± 4.90 | 43.14 ± 1.40 | 29.89 ± 4.40 | 28.05 ± 0.00 | 29.98 ± 7.93 | 22.72 ± 1.83 | 56.14 ± 8.97 * | 40.27 ± 7.36 | 21.66 ± 4.65 | 39.30 ± 1.62 | 36.06 ± 3.35 | 36.20 ± 2.29 |
| 72 h | 32.59 ± 3.23 | 40.14 ± 2.90 | 43.87 ± 5.97 | 47.22 ± 5.16 * | 35.55 ± 4.47 | 9.83 ± 4.33 * | 43.25 ± 5.01 | 34.14 ± 4.41 | 9.89 ± 0.85 * | 35.71 ± 3.03 | 37.52 ± 5.86 | 38.78 ± 7.87 |
| Nthy-ori 3-1 | ||||||||||||
| 24 h | 61.69 ± 7.56 | 59.24 ± 1.89 | 51.67 ± 4.83 | 44.99 ± 6.65 * | 42.32 ± 4.65 * | 38.45 ± 3.66 * | 32.29 ± 4.60 * | 38.31 ± 4.16 * | 43.80 ± 2.82 | 40.91 ± 2.69 * | 36.90 ± 3.07 * | 39.57 ± 2.94 |
| 48 h | 53.25 ± 5.67 | 50.90 ± 9.05 | 42.52 ± 8.77 | 37.51 ± 10.10 | 41.89 ± 7.69 | 38.06 ± 3.95 | 36.34 ± 4.80 | 40.02 ± 7.11 | 33.99 ± 4.67 | 31.87 ± 1.06 * | 32.58 ± 2.59 | 34.53 ± 1.63 |
| 72 h | 47.36 ± 3.82 | 41.32 ± 4.84 | 37.86 ± 5.88 | 38.35 ± 5.63 | 32.90 ± 1.60 | 29.02 ± 3.91 | 27.18 ± 3.98 * | 29.56 ± 5.15 | 25.94 ± 2.94 | 27.02 ± 6.03 * | 27.62 ± 4.20 * | 27.94 ± 5.66 |
| 5KoL | 6KoL | 7KoL | 5KoLM | 6KoLM | 7KoLM | 5KoD | 6KoD | 7KoD | 5KoDM | 6KoDM | 7KoDM | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| FTC-133 | ||||||||||||
| 24 h | 133.10 ± 10.82 | 117.03 ± 0.22 | 101.58 ± 9.14 | 93.73 ± 5.05 * | 98.06 ± 11.47 * | 101.17 ± 9.22 | 112.29 ± 6.06 * | 108.92 ± 9.09 | 106.48 ± 5.62 | 91.79 ± 4.01 * | 106.02 ± 1.08 | 102.19 ± 3.53 |
| 48 h | 110.81 ± 11.84 | 113.88 ± 11.20 | 94.91 ± 8.39 | 84.39 ± 0.92 * | 104.66 ± 6.69 | 85.80 ± 2.22 | 84.28 ± 9.86 * | 111.49 ± 10.80 | 108.83 ± 2.33 | 100.38 ± 8.54 | 98.54 ± 12.73 | 110.41 ± 4.69 |
| 72 h | 80.53 ± 5.02 | 76.72 ± 7.78 | 85.93 ± 8.53 | 51.55 ± 5.47 * | 54.79 ± 9.48 * | 56.02 ± 8.71 * | 66.20 ± 3.99 | 69.61 ± 4.48 | 67.73 ± 3.58 * | 44.70 ± 0.23 * | 61.62 ± 4.59 | 67.80 ± 0.40 * |
| 8505C | ||||||||||||
| 24 h | 119.00 ± 5.43 | 105.25 ± 9.47 | 106.31 ± 9.33 | 103.11 ± 5.12 | 112.71 ± 5.47 | 102.15 ± 9.94 | 103.97 ± 6.97 | 97.68 ± 7.68 | 102.79 ± 6.66 | 109.40 ± 3.32 | 111.96 ± 6.33 | 99.01 ± 4.30 |
| 48 h | 125.43 ± 0.82 | 105.84 ± 4.05 | 104.62 ± 3.54 | 91.33 ± 1.77 * | 108.16 ± 4.79 | 111.37 ± 7.63 | 113.55 ± 6.63 | 110.60 ± 0.39 | 103.92 ± 4.79 | 101.35 ± 6.06 * | 108.28 ± 0.00 | 96.53 ± 2.00 * |
| 72 h | 101.81 ± 7.40 | 90.11 ± 3.21 | 83.31 ± 6.00 | 71.75 ± 1.76 * | 77.57 ± 4.89 | 97.09 ± 17.50 | 79.98 ± 6.30 * | 82.85 ± 3.51 | 73.34 ± 5.59 | 75.11 ± 5.29 * | 92.22 ± 9.91 | 95.62 ± 9.31 |
| TPC-1 | ||||||||||||
| 24 h | 80.63 ± 4.29 | 63.13 ± 10.30 | 71.21 ± 2.41 | 47.29 ± 1.22 * | 79.69 ± 6.32 | 74.08 ± 8.57 | 85.44 ± 5.53 | 84.30 ± 2.34 * | 88.71 ± 8.52 | 82.77 ± 9.08 | 85.24 ± 8.72 * | 88.44 ± 8.16 |
| 48 h | 66.57 ± 0.67 | 42.08 ± 1.77 | 43.75 ± 5.48 | 22.059 ± 4.88 * | 62.16 ± 8.25 * | 57.43 ± 5.64 * | 55.19 ± 4.94 | 55.82 ± 3.22 * | 59.98 ± 5.10 * | 55.48 ± 2.56 | 50.40 ± 2.83 | 56.25 ± 3.35 |
| 72 h | 49.86 ± 0.32 | 33.91 ± 4.69 | 37.83 ± 9.79 | 13.73 ± 0.97 * | 47.82 ± 3.94 | 51.51 ± 10.12 | 65.94 ± 4.23 | 50.06 ± 3.49 | 49.42 ± 4.36 | 49.24 ± 7.94 | 52.73 ± 6.77 * | 62.22 ± 3.24 * |
| Nthy-ori 3-1 | ||||||||||||
| 24 h | 76.39 ± 6.61 | 68.00 ± 4.59 | 52.41 ± 4.68 | 45.73 ± 6.13 * | 51.60 ± 6.72 * | 43.13 ± 3.21 | 36.75 ± 0.94 * | 44.84 ± 5.39 * | 49.67 ± 5.76 | 41.50 ± 3.76 * | 40.53 ± 1.78 * | 36.97 ± 2.52 * |
| 48 h | 64.53 ± 6.88 | 60.77 ± 1.76 | 47.53 ± 4.76 | 39.47 ± 7.64 * | 41.50 ± 5.82 * | 39.62 ± 3.30 | 39.94 ± 2.49 * | 45.26 ± 2.18 * | 51.53 ± 3.53 | 44.09 ± 2.94 * | 43.46 ± 2.01 * | 48.94 ± 7.09 |
| 72 h | 51.67 ± 7.96 | 44.88 ± 6.85 | 37.22 ± 5.37 | 30.64 ± 3.14 * | 31.55 ± 0.97 | 32.96 ± 2.46 | 28.37 ± 5.57 * | 29.83 ± 2.35 * | 36.73 ± 6.17 | 28.16 ± 2.97 * | 30.20 ± 5.70 * | 29.93 ± 4.86 |
| 5BSL | 6BSL | 7BSL | 5BSLM | 6BSLM | 7BSLM | 5BSD | 6BSD | 7BSD | 5BSDM | 6BSDM | 7BSDM | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| FTC-133 | ||||||||||||
| 24 h | 131.57 ± 13.76 | 83.63 ± 3.68 | 79.96 ± 6.43 | 93.63 ± 13.10 * | 77.82 ± 3.37 | 83.63 ± 7.78 | 96.58 ± 12.80 * | 88.83 ± 9.46 | 79.35 ± 12.90 | 80.88 ± 6.46 * | 81.39 ± 4.86 | 83.02 ± 10.20 |
| 48 h | 112.52 ± 8.74 | 71.54 ± 5.52 | 77.15 ± 1.95 | 95.45 ± 1.84 * | 78.78 ± 0.81 | 92.44 ± 9.08 * | 78.86 ± 11.00 * | 77.72 ± 2.07 | 79.27 ± 8.85 | 99.84 ± 9.16 | 74.47 ± 7.82 | 110.08 ± 2.75 * |
| 72 h | 63.12 ± 8.43 | 32.94 ± 5.55 | 29.29 ± 3.54 | 65.69 ± 9.09 | 27.95 ± 6.12 | 48.28 ± 9.54 | 53.25 ± 4.64 | 78.59 ± 12.70 * | 30.58 ± 5.46 | 62.09 ± 3.11 | 47.99 ± 6.91 | 49.79 ± 3.14 * |
| 8505C | ||||||||||||
| 24 h | 95.81 ± 10.60 | 93.73 ± 8.37 | 86.48 ± 5.69 | 82.11 ± 8.96 | 86.59 ± 6.74 | 86.69 ± 2.42 | 86.27 ± 8.97 | 90.64 ± 6.79 | 82.00 ± 5.69 | 85.63 ± 5.62 | 79.65 ± 5.43 | 90.53 ± 7.82 |
| 48 h | 116.96 ± 9.54 | 100.71 ± 8.69 | 78.61 ± 5.45 | 83.37 ± 3.32 * | 79.77 ± 8.72 * | 103.79 ± 3.58 * | 86.45 ± 7.27 * | 75.92 ± 3.68 * | 78.74 ± 6.27 | 78.23 ± 1.02 * | 74.63 ± 11.40 * | 95.18 ± 9.73 |
| 72 h | 68.66 ± 7.65 | 71.37 ± 0.45 | 63.56 ± 3.68 | 66.09 ± 5.51 | 60.61 ± 11.70 | 43.73 ± 1.92 * | 56.99 ± 2.40 | 52.23 ± 1.76 * | 60.65 ± 4.16 | 49.92 ± 9.11 * | 64.12 ± 7.71 | 68.73 ± 6.61 |
| TPC-1 | ||||||||||||
| 24 h | 92.52 ± 6.31 | 71.14 ± 1.84 | 67.40 ± 2.01 | 90.71 ± 8.37 | 64.73 ± 11.62 | 92.85 ± 3.95 * | 86.17 ± 3.68 | 90.98 ± 6.86 * | 74.35 ± 3.68 | 99.93 ± 1.98 | 101.20 ± 7.81 * | 99.20 ± 11.34 * |
| 48 h | 65.98 ± 5.26 | 58.86 ± 4.76 | 43.98 ± 6.43 | 98.06 ± 7.63 * | 41.74 ± 3.19 | 67.01 ± 9.08 * | 76.07 ± 3.86 | 84.22 ± 9.12 * | 48.51 ± 7.06 | 92.76 ± 7.85 * | 84.56 ± 7.87 * | 93.45 ± 3.15 * |
| 72 h | 36.81 ± 5.36 | 20.61 ± 4.56 | 21.26 ± 5.27 | 59.96 ± 7.06 * | 21.52 ± 4.66 | 45.97 ± 8.62 * | 53.85 ± 3.71 | 59.36 ± 7.97 * | 35.62 ± 2.79 | 52.04 ± 3.18 | 58.57 ± 4.91 * | 55.51 ± 2.27 * |
| Nthy-ori 3-1 | ||||||||||||
| 24 h | 51.52 ± 4.31 | 45.29 ± 1.79 | 39.64 ± 3.28 | 44.25 ± 1.87 | 42.02 ± 6.63 | 43.95 ± 7.02 | 46.92 ± 8.59 | 44.10 ± 8.25 | 34.45 ± 3.79 | 43.88 ± 3.58 | 37.56 ± 9.86 | 36.82 ± 4.48 |
| 48 h | 44.17 ± 4.98 | 46.12 ± 6.10 | 43.46 ± 2.31 | 41.35 ± 3.46 | 40.33 ± 5.73 | 41.93 ± 1.83 | 40.17 ± 6.54 | 38.21 ± 3.64 | 35.32 ± 6.20 | 35.87 ± 3.95 | 36.49 ± 1.11 | 43.93 ± 3.73 |
| 72 h | 41.69 ± 3.64 | 36.14 ± 3.92 | 34.95 ± 7.01 | 34.25 ± 7.36 | 29.61 ± 0.97 | 30.42 ± 2.82 | 33.71 ± 3.33 | 29.83 ± 8.36 | 25.94 ± 2.30 | 26.27 ± 6.40 * | 27.40 ± 3.05 | 27.56 ± 3.65 |
| Pairs of Correlated Parameters | Correlation Coefficient | |
|---|---|---|
| First latent component | ||
| chlorogenic acid | t2 | 0.990 |
| sulforaphane | t2 | −0.879 |
| chlorogenic acid | sulforaphane | −0.938 |
| Second latent component | ||
| glucoiberin | progoitrin | 0.957 |
| caffeic acid | t4 | 0.941 |
| glucoiberin | t4 | −0.948 |
| caffeic acid | progoitrin | −0.951 |
| progoitrin | t4 | −1.000 |
| caffeic acid | glucoiberin | −1.000 |
| First Latent Component; Correlations Implied with Original Parameters | |||||
| Parameter | 72 h 8505C CV50 | 24 h 8505C CV100 | 72 h 8505C CV100 | ||
| chlorogenic acid | −0.297 | −0.327 | −0.297 | ||
| sulforaphane | 0.261 | 0.287 | 0.261 | ||
| Second Latent Component; Correlations Implied with Original Parameters | |||||
| Parameter | TPC-1 MTT 300 | Nthy-ori 3-1 MTT 300 | 24 h 8505C CV50 | 72 h FTC-133 CV50 | FTC-133 MTT 300 |
| caffeic acid | 0.287 | 0.287 | 0.354 | −0.287 | −0.392 |
| glucoiberin | −0.284 | −0.284 | −0.351 | 0.284 | 0.389 |
| progoitrin | −0.300 | −0.300 | −0.370 | 0.300 | 0.410 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Markiewicz, M.; Galanty, A.; Zagrodzki, P.; Kołodziejczyk, A.; Paśko, P. Searching for Innovative Functional Foods: Correlation Between Chemopreventive Potential and Bioactive Compounds Accumulation in Brassica Sprouts Grown Under Altered Gravity Conditions. Int. J. Mol. Sci. 2025, 26, 11287. https://doi.org/10.3390/ijms262311287
Markiewicz M, Galanty A, Zagrodzki P, Kołodziejczyk A, Paśko P. Searching for Innovative Functional Foods: Correlation Between Chemopreventive Potential and Bioactive Compounds Accumulation in Brassica Sprouts Grown Under Altered Gravity Conditions. International Journal of Molecular Sciences. 2025; 26(23):11287. https://doi.org/10.3390/ijms262311287
Chicago/Turabian StyleMarkiewicz, Marta, Agnieszka Galanty, Paweł Zagrodzki, Agata Kołodziejczyk, and Paweł Paśko. 2025. "Searching for Innovative Functional Foods: Correlation Between Chemopreventive Potential and Bioactive Compounds Accumulation in Brassica Sprouts Grown Under Altered Gravity Conditions" International Journal of Molecular Sciences 26, no. 23: 11287. https://doi.org/10.3390/ijms262311287
APA StyleMarkiewicz, M., Galanty, A., Zagrodzki, P., Kołodziejczyk, A., & Paśko, P. (2025). Searching for Innovative Functional Foods: Correlation Between Chemopreventive Potential and Bioactive Compounds Accumulation in Brassica Sprouts Grown Under Altered Gravity Conditions. International Journal of Molecular Sciences, 26(23), 11287. https://doi.org/10.3390/ijms262311287