
The Role of NF-κB in Endometrial Diseases in Humans and Animals: A Review

 , ,
, ,  , and
, and
Abstract
1. Introduction
2. NF-κB Signaling in Endometrial Disease
2.1. NF-κBa Activation
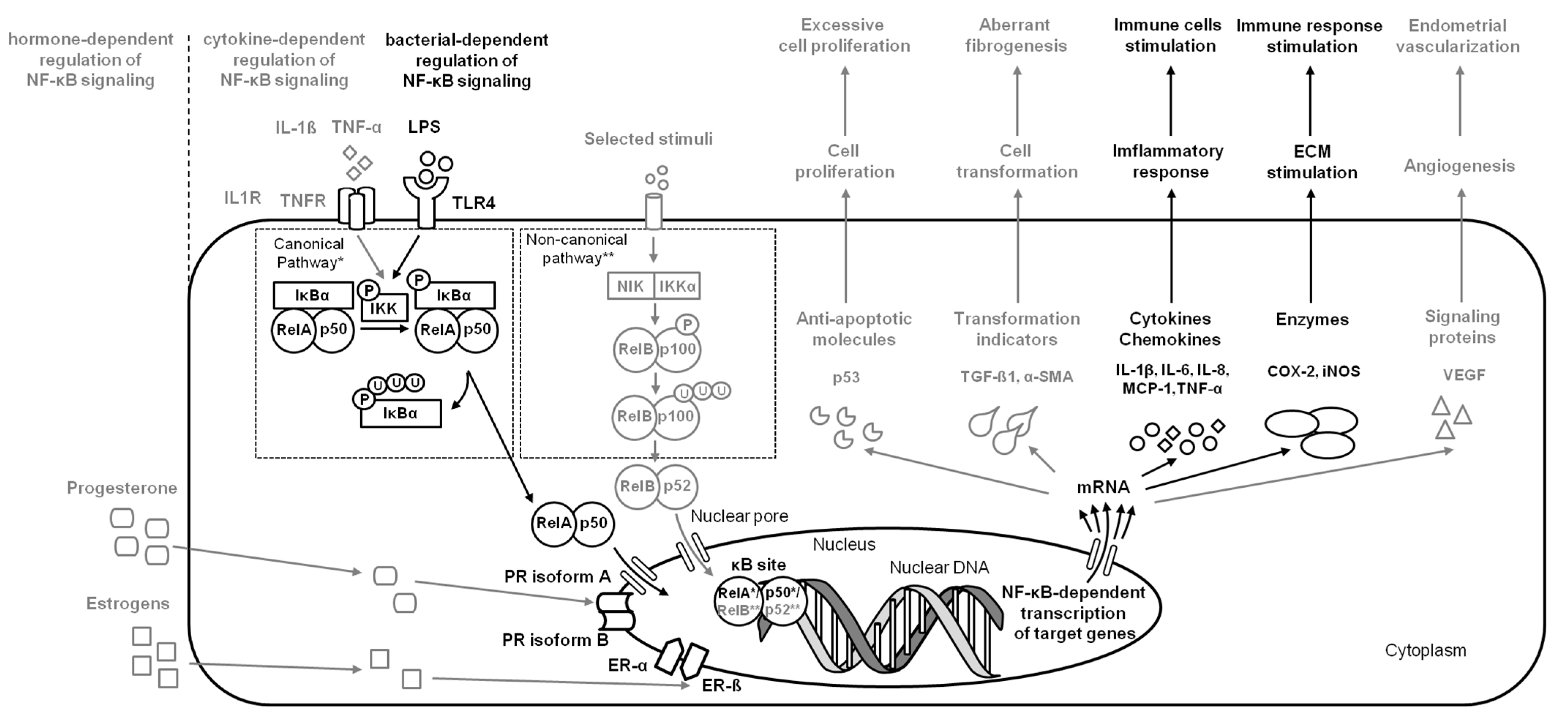
2.2. Endometritis
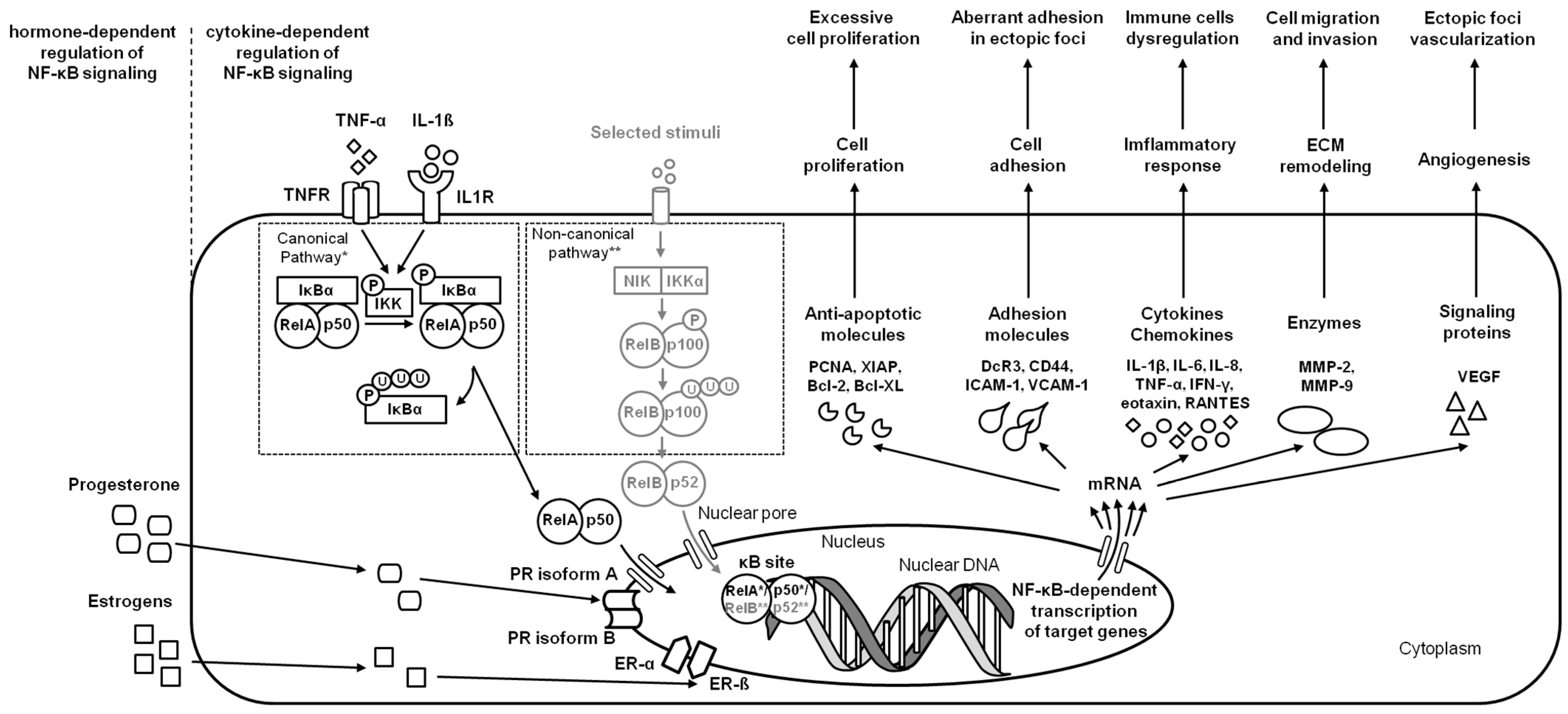
2.3. Endometriosis
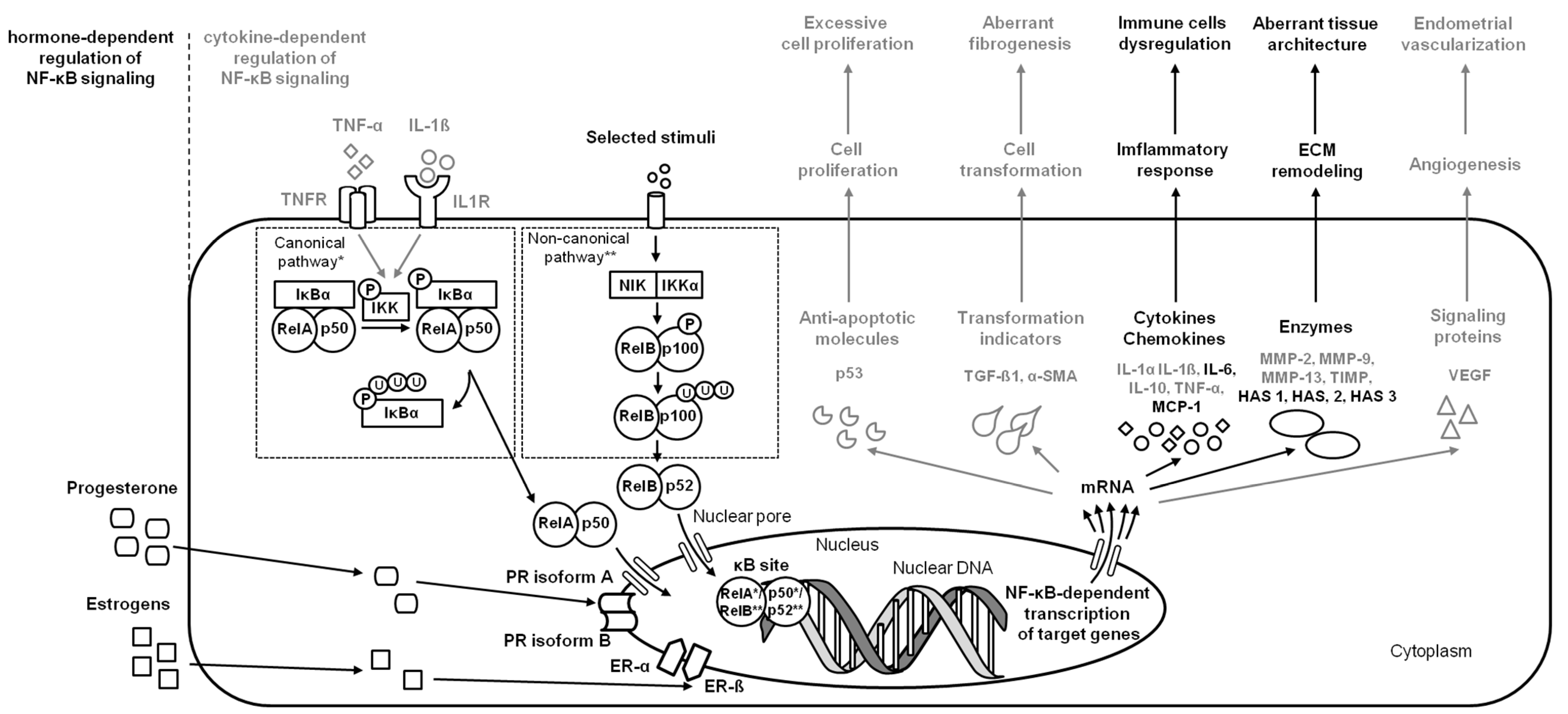
2.4. Endometrosis
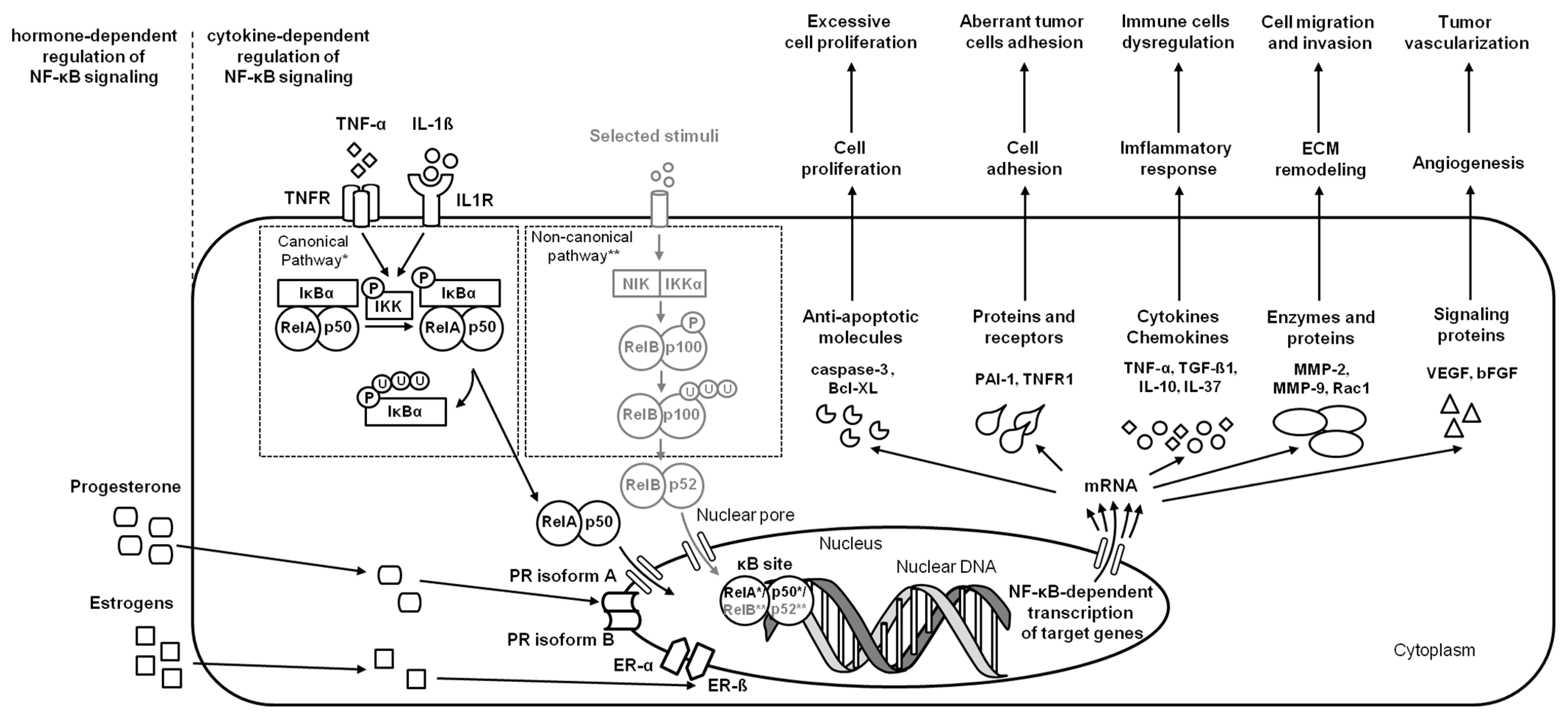
2.5. Endometrial Carcinoma
3. Regulation of NF-κB Signaling in Endometrium
3.1. Progesterone’s Regulation
3.2. Estrogen’s Regulation
4. Perspectives for NF-κB Signaling in Endometrium
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, Y.; Wang, J.; Zhang, X. An Update on the Multifaceted Role of NF-kappaB in Endometriosis. Int. J. Biol. Sci. 2022, 18, 4400–4413. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J. Current uses of mushrooms in cancer treatment and their anticancer mechanisms. Int. J. Mol. Sci. 2022, 23, 10502. [Google Scholar] [CrossRef] [PubMed]
- Mothes, J.; Busse, D.; Kofahl, B.; Wolf, J. Sources of dynamic variability in NF-κB signal transduction: A mechanistic model. BioEssays 2015, 37, 452–462. [Google Scholar] [CrossRef]
- García-García, V.A.; Alameda, J.P.; Page, A.; Casanova, M.L. Role of NF-κB in ageing and age-related diseases: Lessons from genetically modified mouse models. Cells 2021, 10, 1906. [Google Scholar] [CrossRef] [PubMed]
- O’Dea, E.; Hoffmann, A. NF-κB signaling. Wiley Interdiscip. Rev. Syst. Biol. Med. 2009, 1, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, I.; Banerjee, S.; Driss, A.; Xu, W.; Mehrabi, S.; Nezhat, C.; Sidell, N.; Taylor, R.N.; Thompson, W.E. Curcumin attenuates proangiogenic and proinflammatory factors in human eutopic endometrial stromal cells through the NF-κB signaling pathway. J. Cell. Physiol. 2019, 234, 6298–6312. [Google Scholar] [CrossRef]
- Cui, L.; Wang, H.; Lin, J.; Wang, Y.; Dong, J.; Li, J.; Li, J. Progesterone inhibits inflammatory response in E. coli-or LPS-Stimulated bovine endometrial epithelial cells by NF-κB and MAPK pathways. Dev. Comp. Immunol. 2020, 105, 103568. [Google Scholar] [CrossRef]
- Domino, M.; Jasinski, T.; Kautz, E.; Juszczuk-Kubiak, E.; Ferreira-Dias, G.; Zabielski, R.; Sady, M.; Gajewski, Z. Expression of genes involved in the NF-κB-dependent pathway of the fibrosis in the mare endometrium. Theriogenology 2020, 147, 18–24. [Google Scholar] [CrossRef]
- Jasiński, T.; Zdrojkowski, Ł.; Kautz, E.; Juszczuk-Kubiak, E.; Ferreira-Dias, G.; Domino, M. Equine endometrosis pathological features: Are they dependent on NF-κB Signaling pathway? Animals 2021, 11, 3151. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. The hallmarks of cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef]
- Jasiński, T.; Zdrojkowski, Ł.; Ferreira-Dias, G.; Kautz, E.; Juszczuk-Kubiak, E.; Domino, M. Molecular Mechanism of Equine Endometrosis: The NF-κB-Dependent Pathway Underlies the Ovarian Steroid Receptors’ Dysfunction. Int. J. Mol. Sci. 2022, 23, 7360. [Google Scholar] [CrossRef]
- Oeckinghaus, A.; Ghosh, S. The NF-kappaB family of transcription factors and its regulation. Cold Spring Harb. Perspect. Biol. 2009, 1, a000034. [Google Scholar] [CrossRef] [PubMed]
- Hayden, M.S.; Ghosh, S. NF-κB, the first quarter-century: Remarkable progress and outstanding questions. Genes Dev. 2012, 26, 203–234. [Google Scholar] [CrossRef] [PubMed]
- May, M.J.; Ghosh, S. Signal transduction through NF-κB. Trends Immunol. 1998, 19, 80–88. [Google Scholar] [CrossRef]
- Umezawa, K. Possible role of peritoneal NF-kB in peripheral inflammation and cancer: Lessons from the inhibitor DHMEQ. Biomed. Pharm. 2011, 65, 252–259. [Google Scholar] [CrossRef]
- Pallares, J.; Martínez-Guitarte, J.L.; Dolcet, X.; Llobet, D.; Rue, M.; Palacios, J.; Prat, J.; Matias-Guiu, X. Abnormalities in the NF-κB family and related proteins in endometrial carcinoma. J. Pathol. 2004, 204, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Mita, S.; Shimizu, Y.; Notsu, T.; Imada, K.; Kyo, S. Dienogest inhibits Toll-like receptor 4 expression induced by costimulation of lipopolysaccharide and high-mobility group box 1 in endometrial epithelial cells. Fertil. Steril. 2011, 96, 1485–1489. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Qu, Y.; Li, J.; Cui, L.; Wang, Y.; Lin, J.; Wang, H. Cortisol inhibits NF-κB and MAPK pathways in LPS activated bovine endometrial epithelial cells. Int. Immunopharmacol. 2018, 56, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Cui, L.; Huang, X.; Wang, S.; Li, D.; Li, L.; Sun, Y.; Du, M. Activation of Rev-erbα attenuates lipopolysaccharide-induced inflammatory reactions in human endometrial stroma cells via suppressing TLR4-regulated NF-κB activation. Acta Biochim. Biophys. Sin. 2019, 51, 908–914. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, X.; Wang, X.; Wan, L.; Liu, Y.; Shi, Y.; Zhang, L.; Fang, Z.; Wei, Z. PDCD4 suppresses proliferation, migration, and invasion of endometrial cells by inhibiting autophagy and NF-κB/MMP2/MMP9 signal pathway. Biol. Reprod. 2018, 99, 360–372. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Gong, Z.; Zhou, S.; Yang, S.; Wang, D.; Chen, X.; Wu, J.; Liu, L.; Zhong, S.; Zhao, J.; et al. Liposomal curcumin targeting endometrial Cancer through the NF-κB pathway. Cell. Physiol. Biochem. 2018, 48, 569–582. [Google Scholar] [CrossRef] [PubMed]
- Besso, M.J.; Rosso, M.; Lapyckyj, L.; Moiola, C.P.; Matos, M.L.; Mercogliano, M.F.; Schilacci, R.; Reventos, J.; Colas, E.; Gil-Moreno, A.; et al. FXYD5/Dysadherin, a biomarker of endometrial cancer myometrial invasion and aggressiveness: Its relationship with TGF-β1 and NF-κB pathways. Front. Oncol. 2019, 9, 1306. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wei, Z.; Tang, Z.; Xue, C.; Yu, H.; Zhang, D.; Li, Y.; Liu, X.; Shi, Y.; Zhang, L.; et al. IL-37bΔ1-45 suppresses the migration and invasion of endometrial cancer cells by targeting the Rac1/NF-κB/MMP2 signal pathway. Lab. Investig. 2021, 101, 760–774. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Huang, O.; Zhang, W.; Liu, L.; Xu, C. Astragaloside IV exerts anti-inflammatory role in endometriosis by downregulating TLR4/NF-κB pathway. Trop. J. Pharm. Res. 2019, 18, 539–545. [Google Scholar] [CrossRef]
- Napetschnig, J.; Wu, H. Molecular basis of NF-κB signaling. Annu. Rev. Biophys. 2013, 42, 443–468. [Google Scholar] [CrossRef]
- Elweza, A.E.; Ezz, M.A.; Acosta, T.J.; Talukder, A.K.; Shimizu, T.; Hayakawa, H.; Shimada, M.; Imakawa, K.; Zaghlouls, A.H.; Miyamoto, A. A proinflammatory response of bovine endometrial epithelial cells to active sperm in vitro. Mol. Reprod. Dev. 2018, 85, 215–226. [Google Scholar] [CrossRef]
- Morris, L.H.A.; McCue, P.M.; Aurich, C. Equine endometritis: A review of challenges and new approaches. Reproduction 2020, 160, 95–110. [Google Scholar] [CrossRef]
- Katila, T.; Ferreira-Dias, G. Evolution of the Concepts of Endometrosis, Post Breeding Endometritis, and Susceptibility of Mares. Animals 2022, 12, 779. [Google Scholar] [CrossRef]
- Kimura, F.; Takebayashi, A.; Ishida, M.; Nakamura, A.; Kitazawa, J.; Morimune, A.; Hirata, K.; Takahashi, A.; Tsuji, S.; Takashima, A.; et al. Review: Chronic endometritis and its effect on reproduction. J. Obstet. Gynaecol. Res. 2019, 45, 951–960. [Google Scholar] [CrossRef]
- Schöniger, S.; Schoon, H.A. The healthy and diseased equine endometrium: A review of morphological features and molecular analyses. Animals 2020, 10, 625. [Google Scholar] [CrossRef]
- Witkowski, M.; Katkiewicz, M.; Zając, S.; Kochan, J. Effect of Long-term Hyperimmunization on the Presence of Mast Cells in the Endometrium of the Mare. J. Equine Vet. Sci. 2015, 35, 569–572. [Google Scholar] [CrossRef]
- Sheldon, I.M.; Cronin, J.G.; Bromfield, J.J. Tolerance and innate immunity shape the development of postpartum uterine disease and the impact of endometritis in dairy cattle. Annu. Rev. Anim. Biosci. 2019, 7, 361–384. [Google Scholar] [CrossRef]
- Hedia, M.; Ibrahim, S.; Mahmoud, K.; Ahmed, Y.; Ismail, S.; El-Belely, M. Hemodynamic changes in cytokines, chemokines, acute phase proteins and prostaglandins in mares with subclinical endometritis. Theriogenology 2021, 171, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Bonizzi, G.; Karin, M. The two NF-kappaB activation pathways and their role in innate and adaptive immunity. Trends Immunol. 2004, 25, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Tsai, H.W.; Huang, M.T.; Wang, P.H.; Huang, B.S.; Chen, Y.J.; Hsieh, S.L. Decoy receptor 3 promotes cell adhesion and enhances endometriosis development. J. Pathol. 2018, 244, 189–202. [Google Scholar] [CrossRef] [PubMed]
- Nasiri, N.; Babaei, S.; Moini, A.; Eftekhari-Yazdi, P. Controlling Semi-Invasive Activity of Human Endometrial Stromal Cells by Inhibiting NF-kB Signaling Pathway Using Aloe-emodin and Aspirin. J. Reprod. Fertil. 2021, 22, 227–240. [Google Scholar] [CrossRef]
- Rebordão, M.R.; Galvão, A.; Szóstek, A.; Amaral, A.; Mateus, L.; Skarzynski, D.J.; Ferreira-Dias, G. Physiopathologic mechanisms involved in mare endometrosis. Reprod. Dom. Anim. 2014, 49, 82–87. [Google Scholar] [CrossRef]
- Alpoim-Moreira, J.; Fernandes, C.; Rebordão, M.R.; Amaral, A.; Pinto-Bravo, P.; Bliebernicht, M.; Skarzynski, D.J.; Ferreira-Dias, G. Collagens and DNA methyltransferases in mare endometrosis. Reprod. Domest. Anim. 2019, 54, 46–52. [Google Scholar] [CrossRef]
- Szóstek, A.Z.; Lukasik, K.; Galvão, A.M.; Ferreira-Dias, G.M.; Skarzynski, D.J. Impairment of the interleukin system in equine endometrium during the course of endometrosis. Biol. Reprod. 2013, 89, 1–13. [Google Scholar] [CrossRef]
- Szóstek-Mioduchowska, A.Z.; Lukasik, K.; Skarzynski, D.J.; Okuda, K. Effect of transforming growth factor-β1 on α-smooth muscle actin and collagen expression in equine endometrial fibroblasts. Theriogenology 2019, 124, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Szóstek-Mioduchowska, A.Z.; Baclawska, A.; Okuda, K.; Skarzynski, D.J. Effect of proinflammatory cytokines on endometrial collagen and metallopeptidase expression during the course of equine endometrosis. Cytokine 2019, 123, 154767. [Google Scholar] [CrossRef]
- Szóstek-Mioduchowska, A.; Słowinska, M.; Pacewicz, J.; Skarzynski, D.J.; Okuda, K. Matrix metallopeptidase expression and modulation by transforming growth factor-1 in equine endometrosis. Sci. Rep. 2020, 10, 1119. [Google Scholar] [CrossRef]
- Szóstek-Mioduchowska, A.; Leciejewska, N.; Zelmanska, B.; Staszkiewicz-Chodor, J.; Ferreira-Dias, G.; Skarzynski, D. Lysophosphatidic acid as a regulator of endometrial connective tissue growth factor and prostaglandin secretion during estrous cycle and endometrosis in the mare. BMC Vet. Res. 2020, 16, 343. [Google Scholar] [CrossRef]
- Skarzynski, D.J.; Szóstek-Mioduchowska, A.Z.; Rebordão, M.R.; Jalali, B.M.; Piotrowska-Tomala, K.K.; Leciejewska, N.; Łazarczyk, M.; Ferreira-Dias, G.M. Neutrophils, monocytes and other immune components in the equine endometrium: Friends or foes? Theriogenology 2020, 150, 150–157. [Google Scholar] [CrossRef]
- Witkowskia, M.; Duliban, M.; Rak, A.; Profaska-Szymik, M.; Gurgul, A.; Arent, Z.J.; Galuszka, A.; Kotula-Balak, M. Next-Generation Sequencing analysis discloses genes implicated in equine endometrosis that may lead to tumorigenesis. Theriogenology 2022, 189, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Piersma, B.; Hayward, M.K.; Weaver, V.M. Fibrosis and cancer: A strained relationship. Biochim. Biophys. Acta Rev. Cancer 2020, 1873, 188356. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Zhao, T.; Yan, H.; Guo, K.; Liu, Z.; Wei, L.; Qiu, C.; Liang, J. Fatostatin reverses progesterone resistance by inhibiting the SREBP1-NF-κB pathway in endometrial carcinoma. Cell Death Dis. 2021, 12, 544. [Google Scholar] [CrossRef] [PubMed]
- Gunson, D.E.; Gillette, D.M.; Beech, J.; Orsini, J. Endometrial adenocarcinoma in a mare. Vet. Pathol. 1980, 17, 776–780. [Google Scholar] [CrossRef] [PubMed]
- Chan, M.K.; Fuentealba, I.C.; Schmitz, D.G.; Read, W.K. Endometrial adenocarcinoma in a mare. Cornell Vet. 1990, 80, 65–73. [Google Scholar]
- Thompson, R.; Armién, A.G.; Rasmussen, J.M.; Wolf, T.M. Uterine adenocarcinoma in a Przewalski’s wild horse (Equus ferus przewalskii). J. Zoo Wildl. Med. 2014, 45, 441–445. [Google Scholar] [CrossRef] [PubMed]
- Katkiewicz, M.; Witkowski, M. Gruczolakorak błony śluzowej macicy klaczy-opis przypadku. Życie Wet. 2016, 91, 852–853. [Google Scholar]
- Lopez, C.; Ciccarelli, M.; Gold, J.R.; Tibary, A. Uterine adenocarcinoma in Quarter horse mare. Equine Vet. Educ. 2017, 30, 640–644. [Google Scholar] [CrossRef]
- Garcia-Iglesias, M.J.; Bravo-Moral, A.M.; Perez-Martinez, C.; Ferreras-Estrada, M.C.; Martinez-Rodriguez, J.M.; Escudero-Diez, A. Incidence and pathomorphology of uterine tumours in the cow. J. Vet. Med. Sci. 1995, 42, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Bockman, J.V. Two pathogenic types of endometrial carcinoma. Gynecol. Oncol. 1983, 15, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; McDonnell, P.C.; Lehr, R.; Tierney, L.; Tzimas, M.N.; Griswold, D.E.; Capper, E.A.; Tal-Singer, R.; Wells, G.I.; Doyle, M.L.; et al. Identification and initial characterization of four novel members of the interleukin-1 family. J. Biol. Chem. 2000, 275, 10308–10314. [Google Scholar] [CrossRef] [PubMed]
- Ding, V.A.; Zhu, Z.; Mantz, A.A.; Xiao, H.; Wakefield, M.R.; Bai, Q.; Fang, Y. The role of IL-37 in non-cancerous diseases. Pathol. Oncol. Res. 2017, 23, 463–470. [Google Scholar] [CrossRef]
- Ding, V.A.; Zhu, Z.; Xiao, H.; Wakefield, M.R.; Bai, Q.; Fang, Y. The role of IL-37 in cancer. Med. Oncol. 2016, 33, 68. [Google Scholar] [CrossRef]
- Bhatnagar, P.C.; Chaudhary, J.L.; Bhardwaj, B.; Shakhar, C.; Gupta, L.; Sharma, D.K. Effect of different hormonal protocols and nutrient supplementation on reproductive performance of cattle under different field conditions. Int. J. Curr. Microbiol. Appl. Sci. 2020, 9, 1511–1522. [Google Scholar] [CrossRef]
- Crabtree, J. Update on the management of anoestrus and transitional phase in horses. In Pract. 2021, 43, 457–466. [Google Scholar] [CrossRef]
- Vercellini, P.; Viganò, P.; Somigliana, E.; Fedele, L. Endometriosis: Pathogenesis and treatment. Nat. Rev. Endocrinol. 2014, 10, 261–275. [Google Scholar] [CrossRef]
- Beagley, K.W.; Gockel, C.M. Regulation of innate and adaptive immunity by the female sex hormones oestradiol and progesterone. FEMS Immunol. Med. Microbiol. 2003, 38, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Bommer, I.; Muzzio, D.O.; Zygmunt, M.; Jensen, F. Progesterone and estradiol exert an inhibitory effect on the production of anti-inflammatory cytokine IL-10 by activated MZ B cells. J. Reprod. Immunol. 2016, 116, 113–116. [Google Scholar] [CrossRef] [PubMed]
- Kalkhoven, E.; Wissink, S.; van der Saag, P.T.; van der Burg, B. Negative interaction between the RelA (p65) subunit of NF-kappaB and the progesterone receptor. J. Biol. Chem. 1996, 271, 6217–6224. [Google Scholar] [CrossRef]
- Mote, P.A.; Arnett-Mansfield, R.L.; Gava, N.; Defazio, A.; Mulac-Jericevic, B.; Conneely, O.M.; Clarke, C.L. Overlapping and distinct expression of progesterone receptors A and B in mouse uterus and mammary gland during the estrus cycle. Endocrinology 2006, 147, 5503–5512. [Google Scholar] [CrossRef] [PubMed]
- Silva, E.S.M.; Scoggin, K.E.; Canisso, I.F.; Troedsson, M.H.T.; Squires, E.L.; Ball, B.A. Expression of receptors for ovarian steroids and prostaglandin E2 in the endometrium and myometrium of mares during estrus, diestrus and early pregnancy. Anim. Reprod. Sci. 2014, 151, 169–181. [Google Scholar] [CrossRef]
- DeFranco, D.B. Navigating steroid hormone receptors through the nuclear compartment. Mol. Endocrinol. 2002, 16, 1449–1455. [Google Scholar] [CrossRef] [PubMed]
- Hartt, L.S.; Carling, S.J.; Joyce, M.M.; Johnson, G.A.; Vanderwall, D.K.; Ott, T.L. Temporal and spatial associations of oestrogen receptor alpha and progesterone receptor in the endometrium of cyclic and early pregnant mares. Reproduction 2005, 130, 241–250. [Google Scholar] [CrossRef]
- González-Ramos, R.; Rocco, J.; Rojas, C.; Sovino, H.; Poch, A.; Kohen, P.; Alvarado-Diaz, C.; Devoto, L. Physiologic activation of nuclear factor kappa-B in the endometrium during the menstrual cycle is altered in endometriosis patients. Fertil. Steril. 2012, 97, 645–651. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.H.; Cui, L.J.; Li, A.Y.; Zhang, J.P.; Liu, Y.; Zhao, J.S.; Xu, X.B.; He, B.; Wang, J.D.; Chu, L.; et al. Endometrial breakdown with sustained progesterone release involves NF-κB-mediated functional progesterone withdrawal in a mouse implant model. Mol. Reprod. Dev. 2016, 83, 780–791. [Google Scholar] [CrossRef] [PubMed]
- Shen, F.; Wang, Y.; Lu, Y.; Yuan, L.; Liu, X.; Guo, S.W. Immunoreactivity of progesterone receptor isoform B and nuclear factor kappa-B as biomarkers for recurrence of ovarian endometriomas. Am. J. Obstet. Gynecol. 2008, 199, 486.e1–486.e10. [Google Scholar] [CrossRef]
- Han, A.R.; Lee, T.H.; Kim, S.; Lee, H.Y. Risk factors and biomarkers for the recurrence of ovarian endometrioma: About the immunoreactivity of progesterone receptor isoform B and nuclear factor kappa B. Gynecol. Endocrinol. 2017, 33, 70–74. [Google Scholar] [CrossRef]
- Enmark, E.; Pelto-Huikko, M.; Grandien, K.; Lagercrantz, S.; Lagercrantz, J.; Fried, G.; Nordenskjold, M.; Gustafsson, J.A. Human estrogen receptor b-gene structure, chromosomal localization, and expression pattern. J. Clin. Endocr. Metab. 1997, 82, 4258–4265. [Google Scholar] [CrossRef] [PubMed]
- Kayisli, U.A.; Guzeloglu-Kayisli, O.; Arici, A. Endocrine-immune interactions in human endometrium. Ann. N. Y. Acad. Sci. 2004, 1034, 50–63. [Google Scholar] [CrossRef]
- Guzeloglu-Kayisli, O.; Halis, G.; Taskiran, S.; Kayisli, U.A.; Arici, A. DNA-binding ability of NF-kappaB is affected differently by ERalpha and ERbeta and its activation results in inhibition of estrogen responsiveness. Reprod. Sci. 2008, 15, 493–505. [Google Scholar] [CrossRef]
- Mei, J.; Zhu, X.Y.; Jin, L.P.; Duan, Z.L.; Li, D.J.; Li, M.Q. Estrogen promotes the survival of human secretory phase endometrial stromal cells via CXCL12/CXCR4 up-regulation-mediated autophagy inhibition. Hum. Reprod. 2015, 30, 1677–1689. [Google Scholar] [CrossRef] [PubMed]
- Han, S.J.; Lee, J.E.; Cho, Y.J.; Park, M.J.; O’Malley, B.W. Genomic Function of Estrogen Receptor β in Endometriosis. Endocrinology 2019, 160, 2495–2516. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Yuan, Y.; He, L.; Yao, X.; Chen, J. Involvement of angiotensin II receptor type 1/NF-κB signaling in the development of endometriosis. Exp. Ther. Med. 2020, 20, 3269–3277. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Song, H.; Lu, Y.; Chen, H.; Jiang, S.; Li, L. Effects of estradiol on VEGF and bFGF by Akt in endometrial cancer cells are mediated through the NF-κB pathway. Oncol. Rep. 2016, 36, 705–714. [Google Scholar] [CrossRef] [PubMed]
- Nishio, H.; Yaguchi, T.; Sugiyama, J.; Sumimoto, H.; Umezawa, K.; Iwata, T.; Susumu, N.; Fujii, T.; Kawamura, N.; Kobayashi, A.; et al. Immunosuppression through constitutively activated NF-κB signalling in human ovarian cancer and its reversal by an NF-κB inhibitor. Br. J. Cancer 2014, 110, 2965–2974. [Google Scholar] [CrossRef]
- Schlein, L.J.; Thamm, D.H. NF-kB activation in canine cancer. Vet. Pathol. 2022, 59, 724–732. [Google Scholar] [CrossRef]
- Jing, H.; Lee, S. NF-κB in cellular senescence and cancer treatment. Mol. Cells 2014, 37, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Mak, I.W.; Evaniew, N.; Ghert, M. Lost in translation: Animal models and clinical trials in cancer treatment. Am. J. Transl. Res. 2014, 6, 114–118. [Google Scholar] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Disease | Species | Tissue | Pathways | Targets/Mechanisms | Reference |
|---|---|---|---|---|---|
| endometritis | Human | hEEC | NF-κB | increase TLR4, IL-6, IL-8, and MCP-1 expression | [13] |
| endometritis | Cattle | BEEC | NF-κB (RelA (p65), IκBα) and MAPK | increase IL-1β, IL-6, IL-8, TNF-α, COX-2, and iNOS expression | [14] |
| endometritis | Human | hESC | NF-κB (pNF-κB, NF-κB) | increase IL-1β, IL-6, IL-8, IL-18, and TNFαexpression | [15] |
| endometritis | Cattle | BEEC | NF-κB (RelA (p65), IκBα) and MAPK | increase IL-1β, IL-6, IL-8, and TNF-α expression | [4] |
| endometriosis | Human | hESC | NF-κB (pNF-κB, NF-κB, pIKKβ, IKKβ, pIKKα, IKKα), STAT3 and JNK | increase IL-1α, IL-1β, IL-4, IL-6, IL-7, IL-8, IL-13, IL-15, IL-17, eotaxin, FGF, G-CSF, INF-γ, IP-10, MCP-1, MIP-1α, MIP-1β, RANTES, TNF-α, and VEGF expression | [3] |
| endometriosis | Human | endometrium | NF-κB (RelA (p65)) | increase MMP-2 and MMP-9 expression | [16] |
| endometriosis | Mouse Model | endometrium | NF-κB (RelA (p65)) | increase IL-1β, IL-6, MCP-1, and TNF-α expression | [17] |
| endometrosis | Horses | endometrium | NF-κB (RelA (p65), NF-κB1 (p50/p105), NF-κB2 (p52/p100)) | decrease IL-6 and increase HAS 1 and HAS 3 expression | [5] |
| endometrosis | Horses | endometrium | NF-κB (RelA (p65), NF-κB1 (p50/p105)) | increase IL-6, MCP-1, and HAS 2 expression | [6] |
| endometrosis | Horses | endometrium | NF-κB (RelA (p65), NF-κB1 (p50/p105), NF-κB2 (p52/p100)) | decrease ER-α, ER-ß, and PR expression | [18] |
| endometrial carcinoma | Human | endometrium | NF-κB (RelA (p65), NF-κB1 (p50/p105), NF-κB2 (p52/p100), cRel, RelB, IκBα, IκBβ, IκBε, and Bcl-3) | increase Bcl-XLexpression | [12] |
| endometrial carcinoma | Human | hECC | NF-κB (RelA (p65)) | decrease caspase-3 activation and increase MMP-9 expression | [19] |
| endometrial carcinoma | Human | endometrium | NF-κB (RelA (p65), pRelA (p65), IκBα) | increase TNF-α, PAI-1, TGF-ß1, and TNFR1 expression | [20] |
| endometrial carcinoma | Human | endometrium, hECC | NF-κB (pIKKα, pIKKβ, IκBα), JNK, and MAPK | decrease IL-37 expression and increase MCP-2, MCP-9, and Rac1 expression | [24] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zdrojkowski, Ł.; Jasiński, T.; Ferreira-Dias, G.; Pawliński, B.; Domino, M. The Role of NF-κB in Endometrial Diseases in Humans and Animals: A Review. Int. J. Mol. Sci. 2023, 24, 2901. https://doi.org/10.3390/ijms24032901
Zdrojkowski Ł, Jasiński T, Ferreira-Dias G, Pawliński B, Domino M. The Role of NF-κB in Endometrial Diseases in Humans and Animals: A Review. International Journal of Molecular Sciences. 2023; 24(3):2901. https://doi.org/10.3390/ijms24032901
Chicago/Turabian StyleZdrojkowski, Łukasz, Tomasz Jasiński, Graça Ferreira-Dias, Bartosz Pawliński, and Małgorzata Domino. 2023. "The Role of NF-κB in Endometrial Diseases in Humans and Animals: A Review" International Journal of Molecular Sciences 24, no. 3: 2901. https://doi.org/10.3390/ijms24032901
APA StyleZdrojkowski, Ł., Jasiński, T., Ferreira-Dias, G., Pawliński, B., & Domino, M. (2023). The Role of NF-κB in Endometrial Diseases in Humans and Animals: A Review. International Journal of Molecular Sciences, 24(3), 2901. https://doi.org/10.3390/ijms24032901