
Psychological Stress Exacerbates Inflammation of the Ileum via the Corticotropin-Releasing Hormone-Mast Cell Axis in a Mouse Model of Eosinophilic Enteritis

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
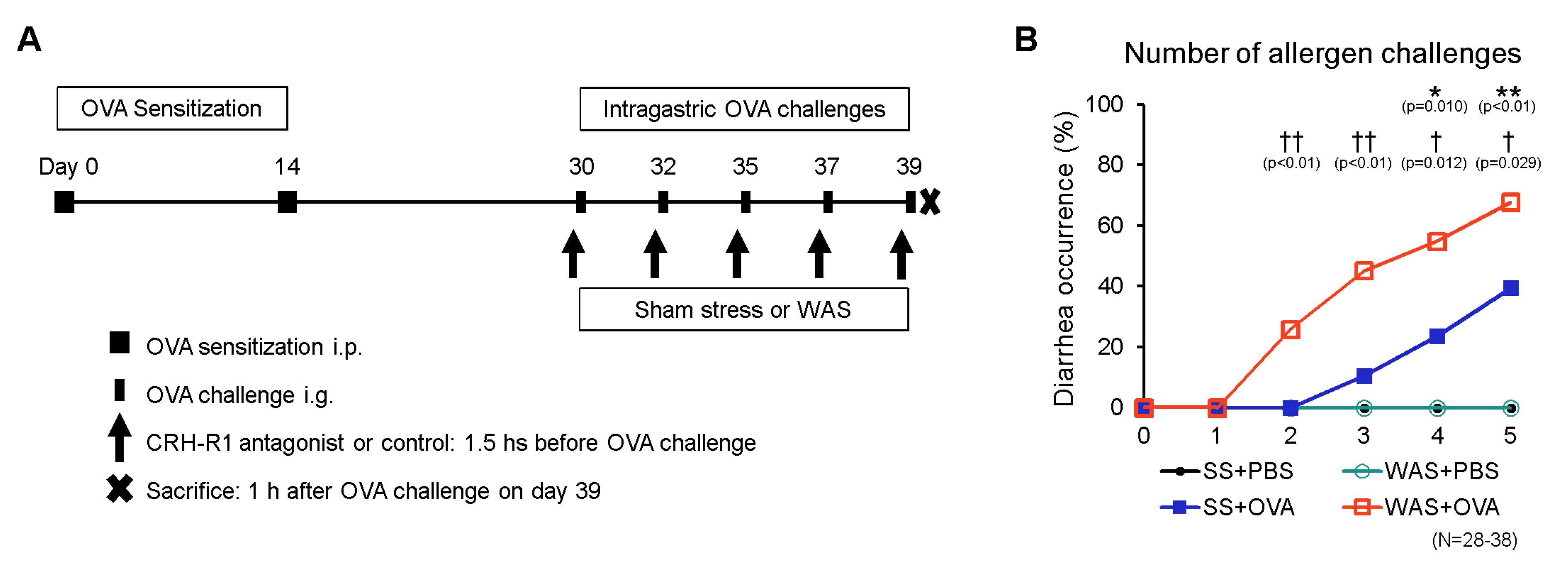
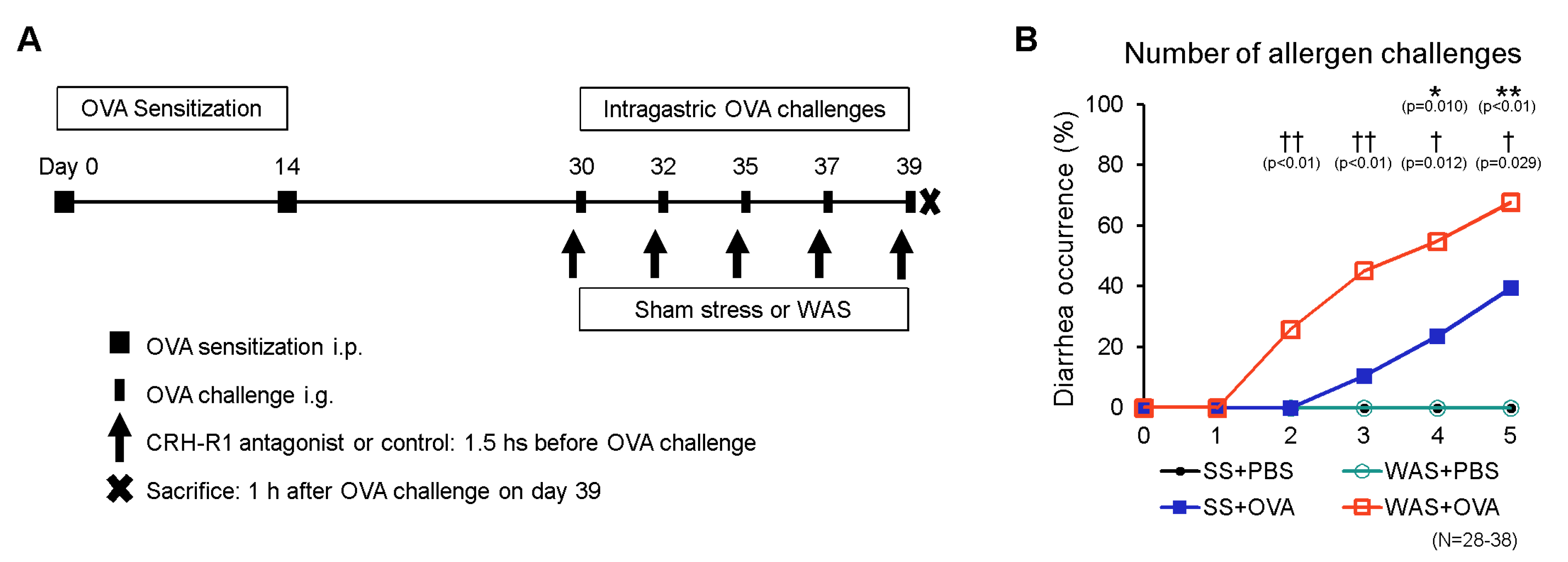
2.1. Establishment of the Oral Allergen-Induced EoN Mouse Model and Effects of Psychological Stress on Diarrhea Occurrence
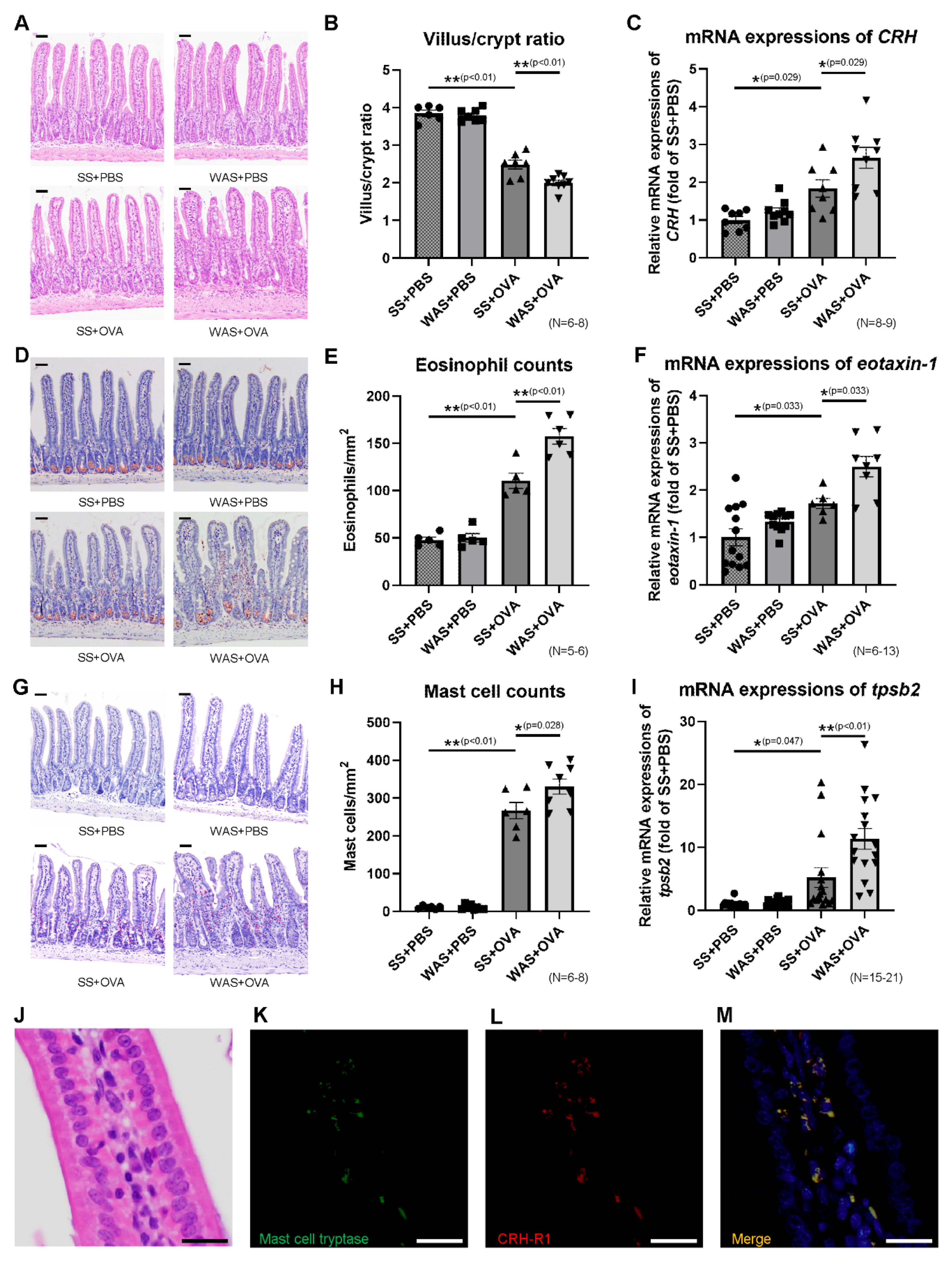
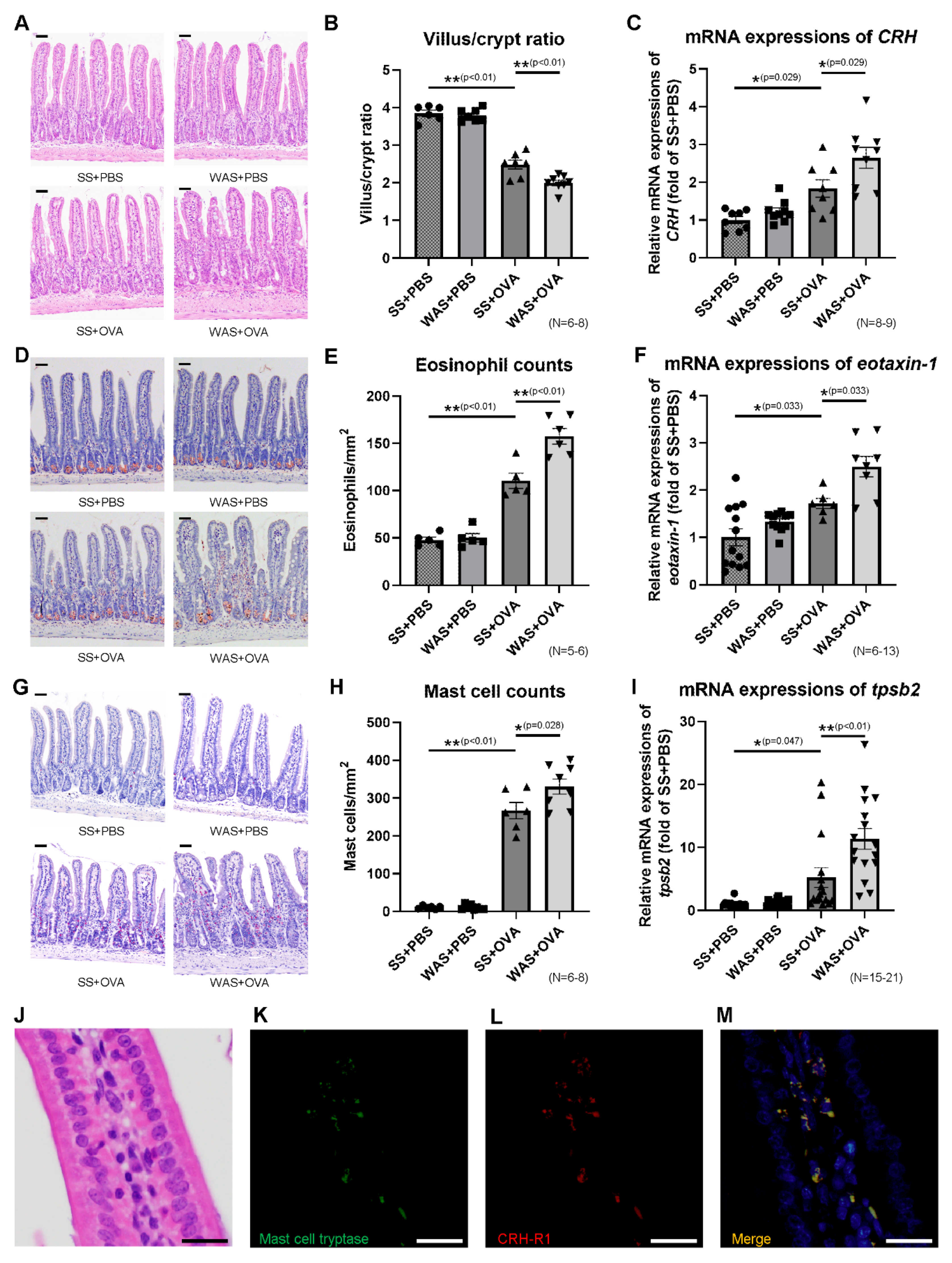
2.2. Effects of Psychological Stress on Villous Atrophy and Crypt Hyperplasia
2.3. Effects of Psychological Stress on the Ileal mRNA Expression of CRH
2.4. Effects of Psychological Stress on the Numbers of Eosinophils and Mast Cells in Ileal Mucosa
2.5. Mast Cell Tryptase and Corticotropin-Releasing Hormone Type 1 Receptor (CRH-R1) Staining
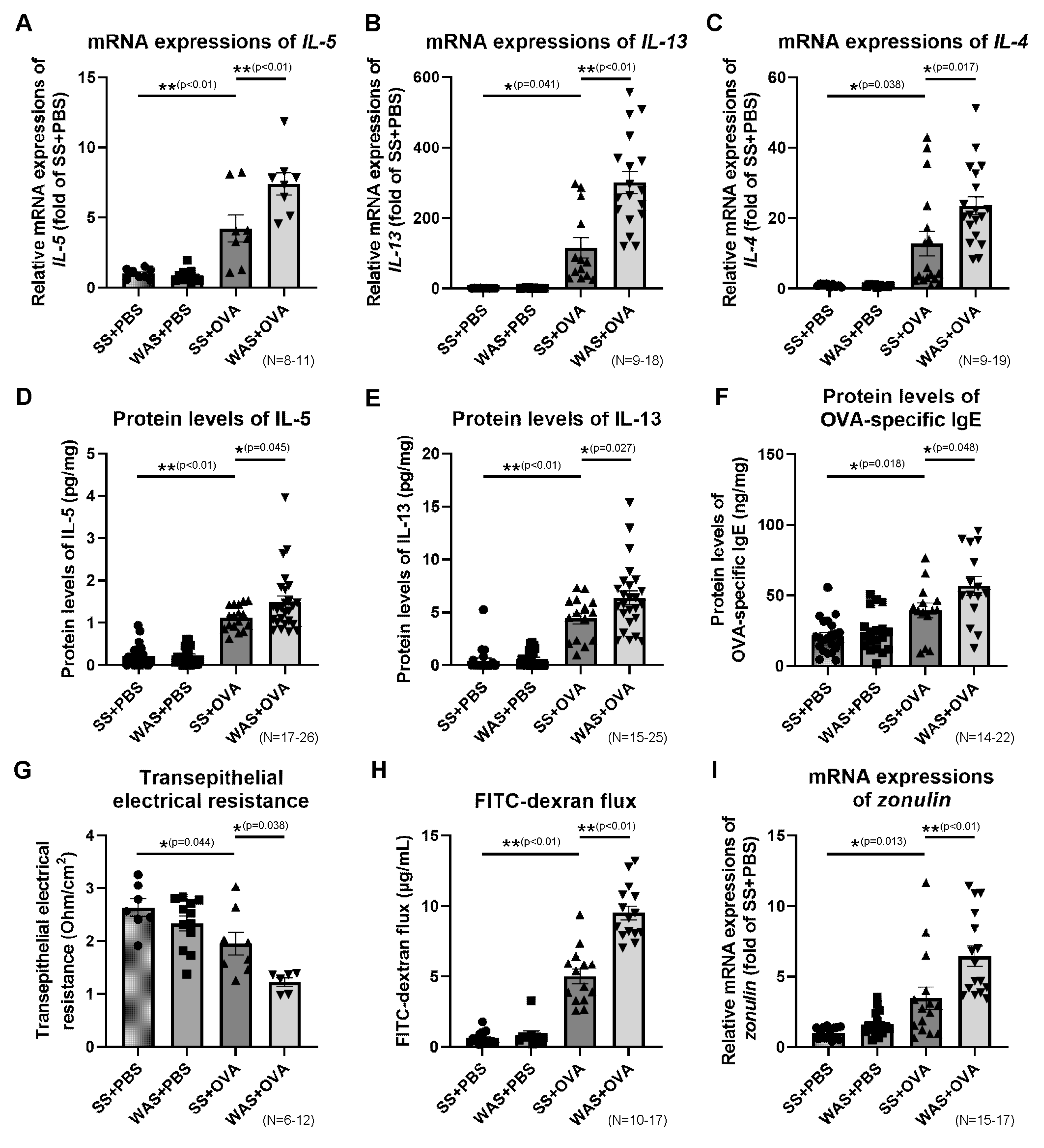
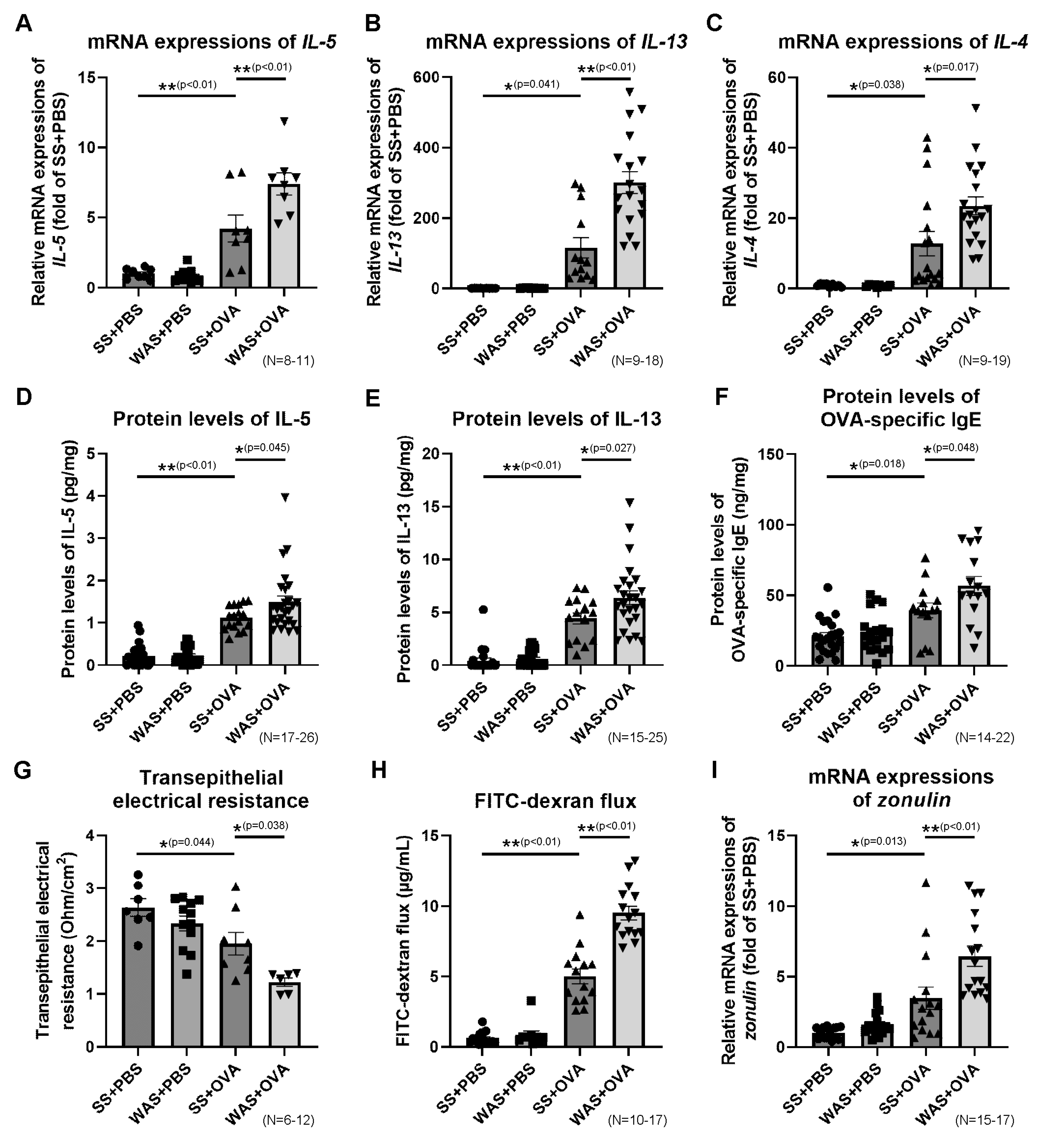
2.6. Effects of Psychological Stress on the Ileal mRNA Expressions of Th2 Cytokines
2.7. Effects of Psychological Stress on the Protein Levels of Ileal Th2 Cytokines and OVA-Specific Immunoglobulin E (IgE)
2.8. Effects of Psychological Stress on Intestinal Permeability
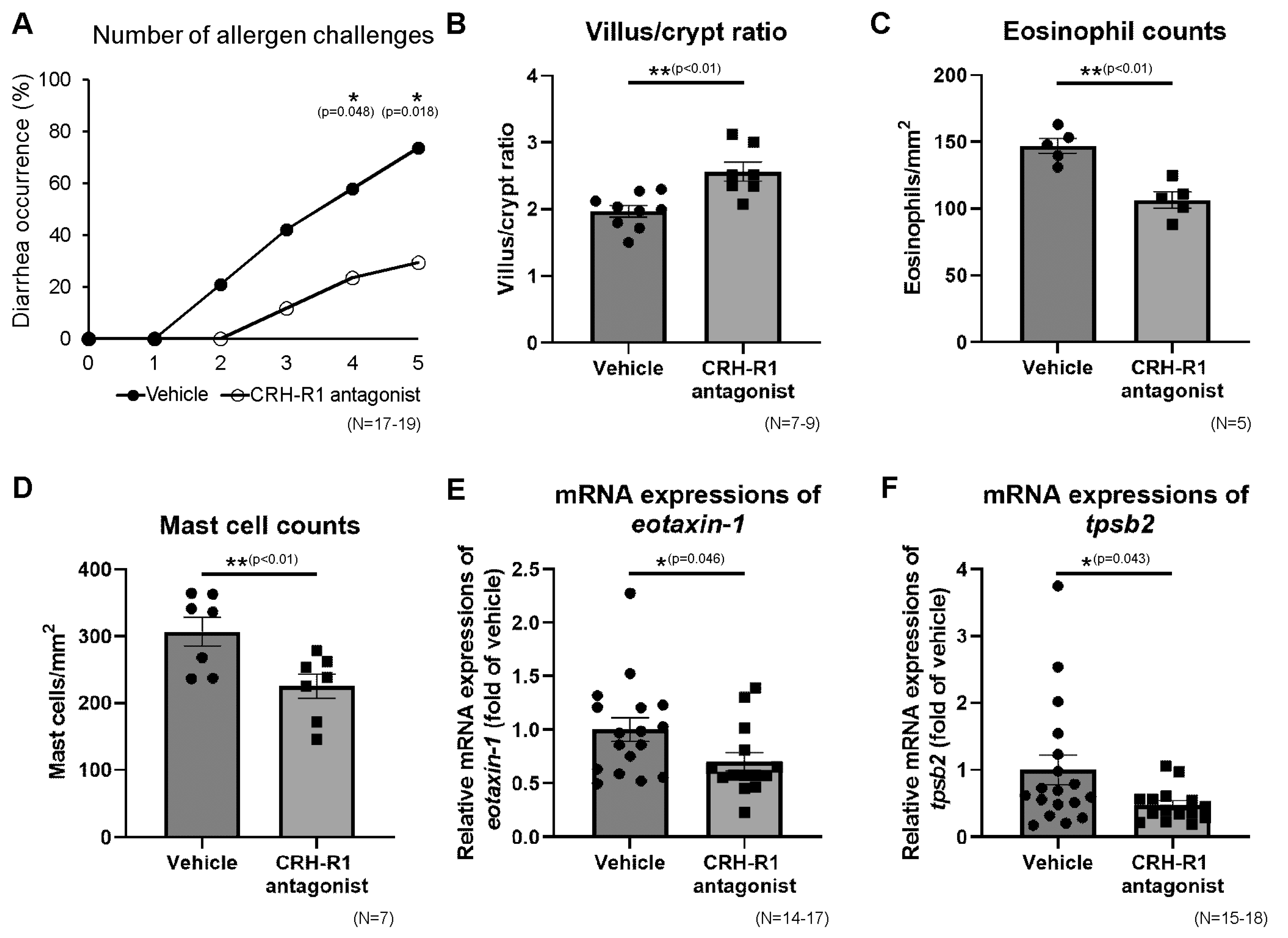
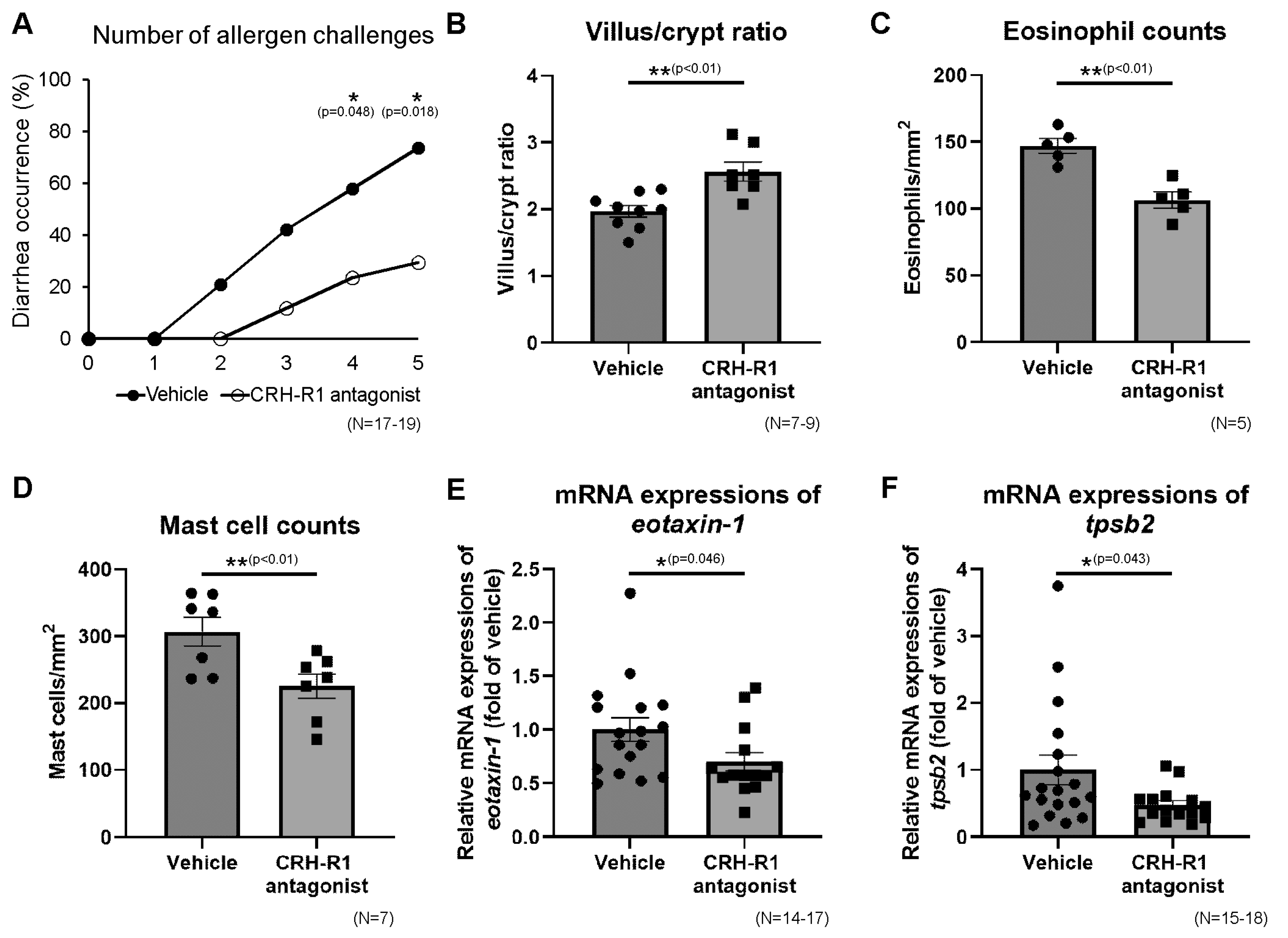
2.9. Effects of CRH-R1 Antagonist on Diarrhea Occurrence, Villus/Crypt Ratio, Eosinophils, and Mast Cells in Ileal Mucosa
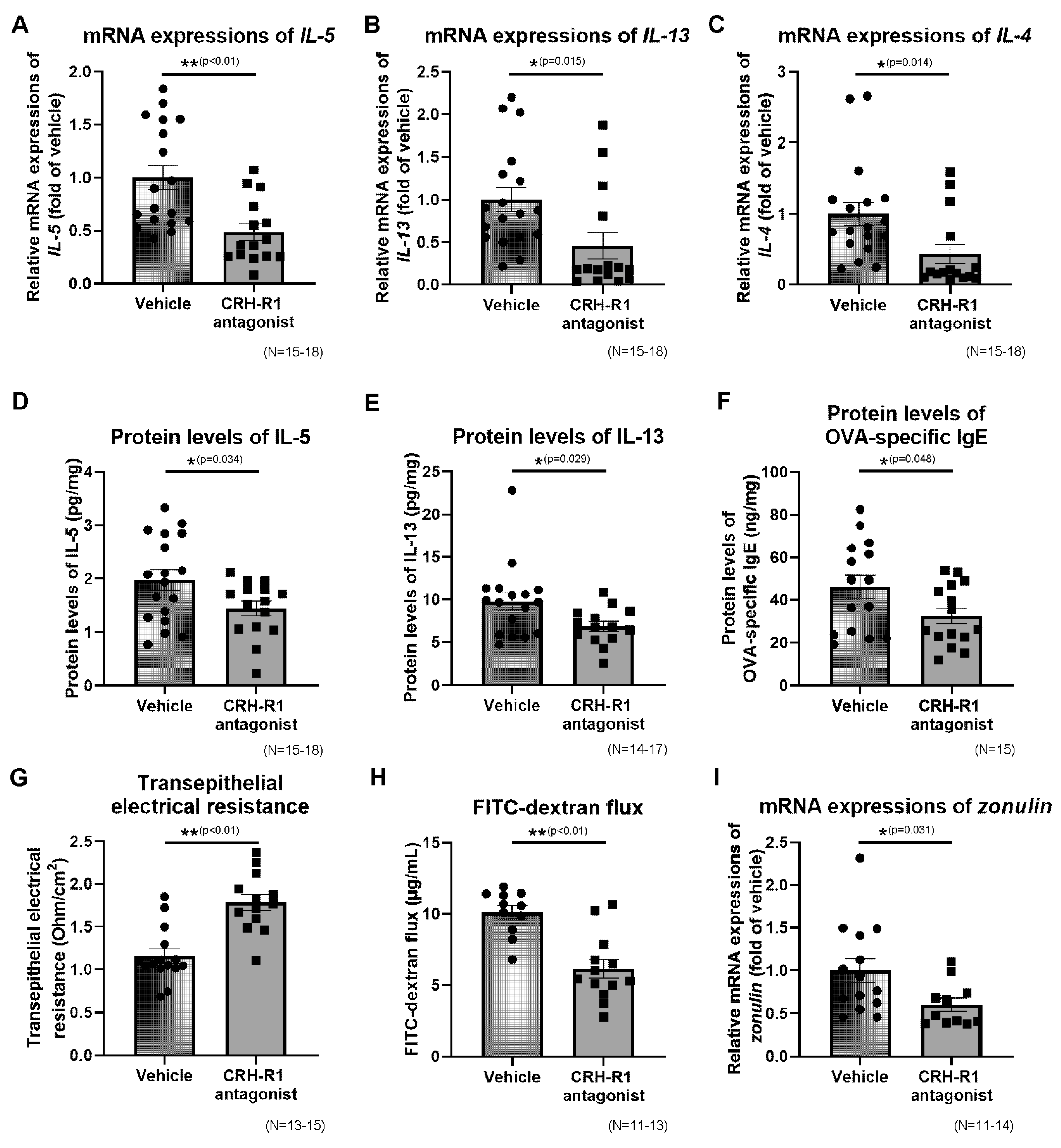
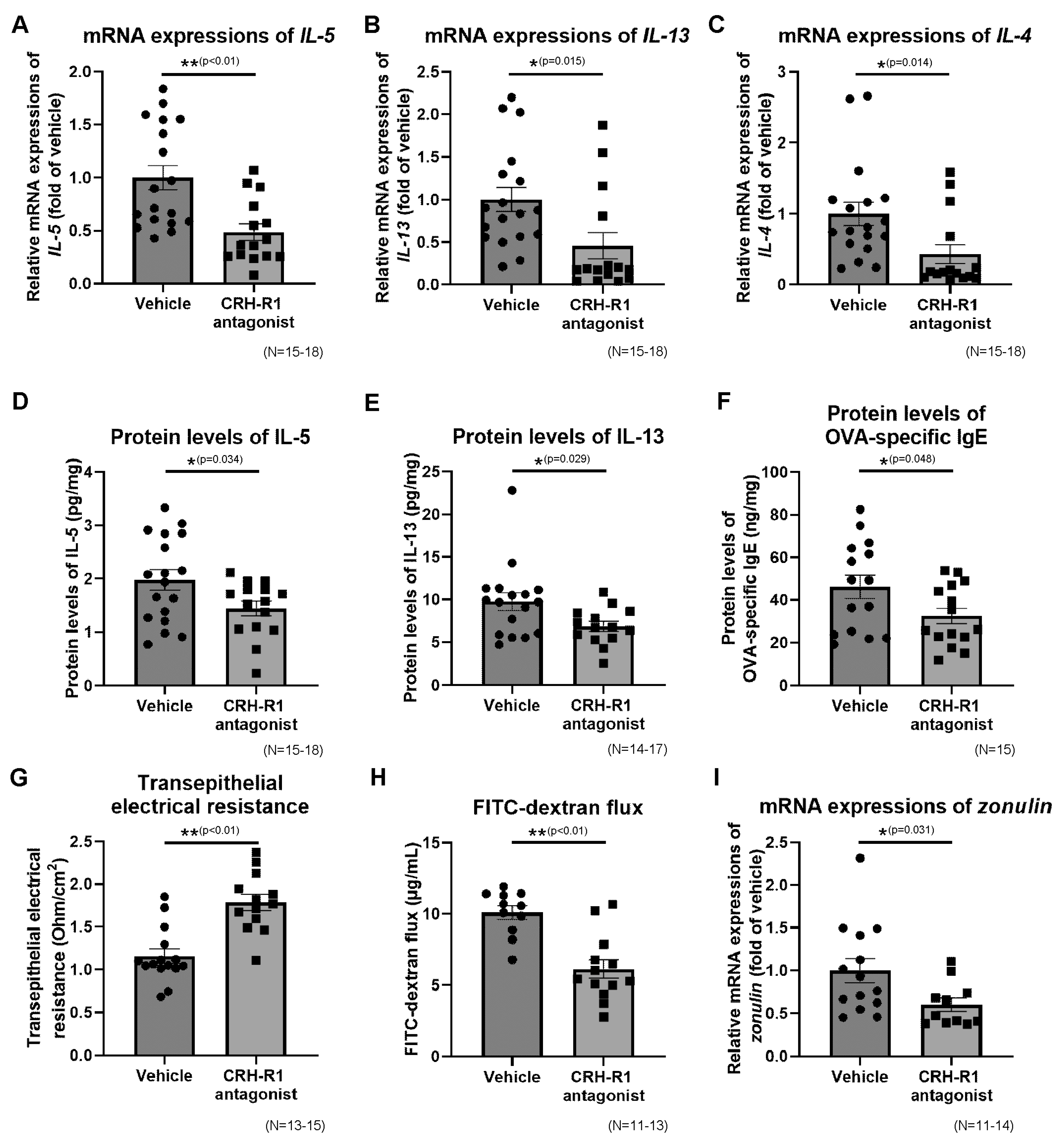
2.10. Effects of CRH-R1 Antagonist on the Ileal mRNA Expressions of Th2 Cytokines, Protein Levels of Th2 Cytokines and OVA-Specific IgE
2.11. Effects of CRH-R1 Antagonist on Intestinal Permeability
3. Discussion
4. Materials and Methods
4.1. Ethics and Animals
4.2. Mouse Model of Oral Allergen-Induced Intestinal Eosinophilic Inflammation and Psychological Stress
4.3. Therapeutic Intervention with the CRH-R1 Antagonist
4.4. Tissue Isolation
4.5. Diarrhea Occurrence
4.6. Microscopic Inflammation in the Ileum
4.7. Ileal Eosinophil and Mast Cell Counts
4.8. Immunofluorescence for Mast Cell Tryptase and CRH-R1
4.9. Quantitative Reverse Transcription-PCR Gene Expression in the Ileum
4.10. Enzyme-Linked Immunosorbent Assay
4.11. Measurement of Ex Vivo Intestinal Permeability, Method 1: Transepithelial Electrical Resistance
4.12. Measurement of Ex Vivo Intestinal Permeability, Method 2: Macromolecular Transport
4.13. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dellon, E.S.; Gonsalves, N.; Abonia, J.P.; Alexander, J.A.; Arva, N.C.; Atkins, D.; Attwood, S.E.; Auth, M.K.H.; Bailey, D.D.; Biederman, L.; et al. International Consensus Recommendations for Eosinophilic Gastrointestinal Disease Nomenclature. Clin. Gastroenterol. Hepatol. 2022. [Google Scholar] [CrossRef]
- Kinoshita, Y.; Furuta, K.; Ishimura, N.; Ishihara, S. Elevated plasma cytokines in Japanese patients with eosinophilic esophagitis and gastroenteritis. Digestion 2012, 86, 238–243. [Google Scholar] [CrossRef] [PubMed]
- Pineton de Chambrun, G.; Dufour, G.; Tassy, B.; Rivière, B.; Bouta, N.; Bismuth, M.; Panaro, F.; Funakoshi, N.; Ramos, J.; Valats, J.C.; et al. Diagnosis, Natural History and Treatment of Eosinophilic Enteritis: A Review. Curr. Gastroenterol. Rep. 2018, 20, 37. [Google Scholar] [CrossRef] [PubMed]
- Pineton de Chambrun, G.; Gonzalez, F.; Canva, J.Y.; Gonzalez, S.; Houssin, L.; Desreumaux, P.; Cortot, A.; Colombel, J.F. Natural history of eosinophilic gastroenteritis. Clin. Gastroenterol. Hepatol. 2011, 9, 950–956.e1. [Google Scholar] [CrossRef]
- Theoharides, T.C.; Enakuaa, S.; Sismanopoulos, N.; Asadi, S.; Papadimas, E.C.; Angelidou, A.; Alysandratos, K.D. Contribution of stress to asthma worsening through mast cell activation. Ann. Allergy Asthma. Immunol. 2012, 109, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Seiffert, K.; Hilbert, E.; Schaechinger, H.; Zouboulis, C.C.; Deter, H.C. Psychophysiological reactivity under mental stress in atopic dermatitis. Dermatology 2005, 210, 286–293. [Google Scholar] [CrossRef]
- Yamanaka-Takaichi, M.; Mizukami, Y.; Sugawara, K.; Sunami, K.; Teranishi, Y.; Kira, Y.; Paus, R.; Tsuruta, D. Stress and Nasal Allergy: Corticotropin-Releasing Hormone Stimulates Mast Cell Degranulation and Proliferation in Human Nasal Mucosa. Int. J. Mol. Sci. 2021, 22, 2773. [Google Scholar] [CrossRef]
- Taft, T.H.; Carlson, D.A.; Simons, M.; Zavala, S.; Hirano, I.; Gonsalves, N.; Pandolfino, J.E. Esophageal Hypervigilance and Symptom-Specific Anxiety in Patients with Eosinophilic Esophagitis. Gastroenterology 2021, 161, 1133–1144. [Google Scholar] [CrossRef] [PubMed]
- Rothenberg, M.E. Eosinophilic gastrointestinal disorders (EGID). J. Allergy Clin. Immunol. 2004, 113, 11–28. [Google Scholar] [CrossRef] [PubMed]
- Cameron, H.L.; Perdue, M.H. Stress impairs murine intestinal barrier function: Improvement by glucagon-like peptide-2. J. Pharmacol. Exp. Ther. 2005, 314, 214–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, P.Y.; Feng, B.S.; Oluwole, C.; Struiksma, S.; Chen, X.; Li, P.; Tang, S.G.; Yang, P.C. Psychological stress induces eosinophils to produce corticotrophin releasing hormone in the intestine. Gut 2009, 58, 1473–1479. [Google Scholar] [CrossRef]
- Vanuytsel, T.; van Wanrooy, S.; Vanheel, H.; Vanormelingen, C.; Verschueren, S.; Houben, E.; Salim Rasoel, S.; Tόth, J.; Holvoet, L.; Farré, R.; et al. Psychological stress and corticotropin-releasing hormone increase intestinal permeability in humans by a mast cell-dependent mechanism. Gut 2014, 63, 1293–1299. [Google Scholar] [CrossRef] [PubMed]
- Wauters, L.; Ceulemans, M.; Schol, J.; Farré, R.; Tack, J.; Vanuytsel, T. The Role of Leaky Gut in Functional Dyspepsia. Front. Neurosci. 2022, 16, 851012. [Google Scholar] [CrossRef]
- Sherrill, J.D.; Kc, K.; Wu, D.; Djukic, Z.; Caldwell, J.M.; Stucke, E.M.; Kemme, K.A.; Costello, M.S.; Mingler, M.K.; Blanchard, C.; et al. Desmoglein-1 regulates esophageal epithelial barrier function and immune responses in eosinophilic esophagitis. Mucosal Immunol. 2014, 7, 718–729. [Google Scholar] [CrossRef]
- Brandt, E.B.; Strait, R.T.; Hershko, D.; Wang, Q.; Muntel, E.E.; Scribner, T.A.; Zimmermann, N.; Finkelman, F.D.; Rothenberg, M.E. Mast cells are required for experimental oral allergen-induced diarrhea. J. Clin. Investig. 2003, 112, 1666–1677. [Google Scholar] [CrossRef] [PubMed]
- Yokooji, T.; Nouma, H.; Matsuo, H. Characterization of ovalbumin absorption pathways in the rat intestine, including the effects of aspirin. Biol. Pharm. Bull. 2014, 37, 1359–1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogawa, T.; Miura, S.; Tsuzuki, Y.; Ogino, T.; Teramoto, K.; Inamura, T.; Watanabe, C.; Hokari, R.; Nagata, H.; Ishii, H. Chronic allergy to dietary ovalbumin induces lymphocyte migration to rat small intestinal mucosa that is inhibited by MAdCAM-1. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 286, G702–G710. [Google Scholar] [CrossRef]
- Alevizos, M.; Karagkouni, A.; Panagiotidou, S.; Vasiadi, M.; Theoharides, T.C. Stress triggers coronary mast cells leading to cardiac events. Ann. Allergy. Asthma. Immunol. 2014, 112, 309–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhuiyan, P.; Wang, Y.W.; Sha, H.H.; Dong, H.Q.; Qian, Y.N. Neuroimmune connections between corticotropin-releasing hormone and mast cells: Novel strategies for the treatment of neurodegenerative diseases. Neural Regen Res 2021, 16, 2184–2197. [Google Scholar]
- Ito, N.; Sugawara, K.; Bodó, E.; Takigawa, M.; van Beek, N.; Ito, T.; Paus, R. Corticotropin-releasing hormone stimulates the in situ generation of mast cells from precursors in the human hair follicle mesenchyme. J. Investig. Dermatol. 2010, 130, 995–1004. [Google Scholar] [CrossRef] [PubMed]
- Kawahito, Y.; Sano, H.; Kawata, M.; Yuri, K.; Mukai, S.; Yamamura, Y.; Kato, H.; Chrousos, G.P.; Wilder, R.L.; Kondo, M. Local secretion of corticotropin-releasing hormone by enterochromaffin cells in human colon. Gastroenterology 1994, 106, 859–865. [Google Scholar] [CrossRef]
- Kawahito, Y.; Sano, H.; Mukai, S.; Asai, K.; Kimura, S.; Yamamura, Y.; Kato, H.; Chrousos, G.P.; Wilder, R.L.; Kondo, M. Corticotropin releasing hormone in colonic mucosa in patients with ulcerative colitis. Gut 1995, 37, 544–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Israeli, E.; Hershcovici, T.; Berenshtein, E.; Zannineli, G.; Wengrower, D.; Weiss, O.; Chevion, M.; Goldin, E. The effect of restraint stress on the normal colon and on intestinal inflammation in a model of experimental colitis. Dig. Dis. Sci. 2008, 53, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Wallon, C.; Yang, P.C.; Keita, A.V.; Ericson, A.C.; McKay, D.M.; Sherman, P.M.; Perdue, M.H.; Söderholm, J.D. Corticotropin-releasing hormone (CRH) regulates macromolecular permeability via mast cells in normal human colonic biopsies in vitro. Gut 2008, 57, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Jacob, C.; Yang, P.C.; Darmoul, D.; Amadesi, S.; Saito, T.; Cottrell, G.S.; Coelho, A.M.; Singh, P.; Grady, E.F.; Perdue, M.; et al. Mast cell tryptase controls paracellular permeability of the intestine. Role of protease-activated receptor 2 and beta-arrestins. J. Biol. Chem. 2005, 280, 31936–31948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, D.; Zhang, X.; Liu, X.; Li, G. Protection of montelukast on OVA-induced eosinophilic gastroenteritis via modulating IL-5, eotaxin-1 and MBP expression. Immunopharmacol. Immunotoxicol. 2013, 35, 625–626. [Google Scholar] [CrossRef] [PubMed]
- Takashima, S.; Tanaka, F.; Kawaguchi, Y.; Usui, Y.; Fujimoto, K.; Nadatani, Y.; Otani, K.; Hosomi, S.; Nagami, Y.; Kamata, N.; et al. Proton pump inhibitors enhance intestinal permeability via dysbiosis of gut microbiota under stressed conditions in mice. Neurogastroenterol. Motil. 2020, 32, e13841. [Google Scholar] [CrossRef] [PubMed]
- Desvignes, C.; Rouquier, L.; Souilhac, J.; Mons, G.; Rodier, D.; Soubrié, P.; Steinberg, R. Control by tachykinin NK(2) receptors of CRF(1) receptor-mediated activation of hippocampal acetylcholine release in the rat and guinea-pig. Neuropeptides 2003, 37, 89–97. [Google Scholar] [CrossRef]
- Friend, D.S.; Ghildyal, N.; Austen, K.F.; Gurish, M.F.; Matsumoto, R.; Stevens, R.L. Mast cells that reside at different locations in the jejunum of mice infected with Trichinella spiralis exhibit sequential changes in their granule ultrastructure and chymase phenotype. J. Cell Biol. 1996, 135, 279–290. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kanamori, A.; Tanaka, F.; Ominami, M.; Nadatani, Y.; Fukunaga, S.; Otani, K.; Hosomi, S.; Kamata, N.; Nagami, Y.; Taira, K.; et al. Psychological Stress Exacerbates Inflammation of the Ileum via the Corticotropin-Releasing Hormone-Mast Cell Axis in a Mouse Model of Eosinophilic Enteritis. Int. J. Mol. Sci. 2022, 23, 8538. https://doi.org/10.3390/ijms23158538
Kanamori A, Tanaka F, Ominami M, Nadatani Y, Fukunaga S, Otani K, Hosomi S, Kamata N, Nagami Y, Taira K, et al. Psychological Stress Exacerbates Inflammation of the Ileum via the Corticotropin-Releasing Hormone-Mast Cell Axis in a Mouse Model of Eosinophilic Enteritis. International Journal of Molecular Sciences. 2022; 23(15):8538. https://doi.org/10.3390/ijms23158538
Chicago/Turabian StyleKanamori, Atsushi, Fumio Tanaka, Masaki Ominami, Yuji Nadatani, Shusei Fukunaga, Koji Otani, Shuhei Hosomi, Noriko Kamata, Yasuaki Nagami, Koichi Taira, and et al. 2022. "Psychological Stress Exacerbates Inflammation of the Ileum via the Corticotropin-Releasing Hormone-Mast Cell Axis in a Mouse Model of Eosinophilic Enteritis" International Journal of Molecular Sciences 23, no. 15: 8538. https://doi.org/10.3390/ijms23158538
APA StyleKanamori, A., Tanaka, F., Ominami, M., Nadatani, Y., Fukunaga, S., Otani, K., Hosomi, S., Kamata, N., Nagami, Y., Taira, K., & Fujiwara, Y. (2022). Psychological Stress Exacerbates Inflammation of the Ileum via the Corticotropin-Releasing Hormone-Mast Cell Axis in a Mouse Model of Eosinophilic Enteritis. International Journal of Molecular Sciences, 23(15), 8538. https://doi.org/10.3390/ijms23158538