1. Introduction
Genes necessary for testis function, particularly in post-meiotic germ cells, are extremely numerous [
1]. However, the function and character of dispensability of these genes is only partially known. Their responsibility in clinical cases of human male infertility is therefore also only partially deciphered.
For example, globozoospermia, a very rare form of male infertility characterized by round-shaped sperm heads that present a complete lack or an abnormal setup of their acrosome, can be explained by alterations of the
DPY19L2 gene in about 60% of reported cases [
2,
3]. Some rare case reports have also revealed mutations of the
SPATA16,
ZPBP1 and
PICK1 genes [
4,
5,
6]. But the defect responsible for some remaining clinical cases is not yet identified and the biogenesis of the acrosome is still not completely elucidated. Animal models can help with suggesting some other genes involved in the process.
Recently, we reported the analysis of a mouse model exhibiting a phenotype of spermatogenesis alterations including anomalies of the acrosome. We identified in an Interspecific Recombinant Congenic Strain (IRCS) a region of mouse chromosome 1 whose presence in a
Mus spretus version within a
Mus musculus domesticus (C57Bl/6) background is associated with signs reminiscent of globozoospermia. This quantitative trait locus (QTL), named
Mafq1 for male fertility QTL chromosome 1, is associated with an interval of approximately 4 Mb between markers
D1Mit438 and
D1Mit305 [
7].
This specific interval contains more than 70 coding genes. Among them, we concentrated our research on the
Spata3 gene (Spermatogenesis associated protein 3, also known as Testis and Spermatogenesis cell Apoptosis Related Gene 1 or
Tsarg1) [
8,
9]. We consider it as one of the best candidates to explain the hypofertility phenotype of the Rc3 strain, on the basis of various arguments.
Spata3 is expressed specifically in the testis both in mice and humans. Precisely, its expression can be detected during spermatogenesis from the round spermatid stage to mature sperm [
7].
Spata3 is conserved in many mammalian species, with mouse and human proteins sharing 55% of their identity.
No data are available on the function of the SPATA3 protein, the SPATA family being a catalogue of proteins sharing an expression in the testis without any similarity of sequence neither function. Furthermore, no specific functional domain can be predicted from its sequence and no mouse model is available to determine its role. However, the human SPATA3 gene has been reported to be among the most down-regulated genes in the testis of infertile men diagnosed with nonobstructive azoospermia [
10]. Rare variants of this gene were also associated with prostate cancer [
11].
Comparison of the sequence of the
Spata3 gene between the
Mus spretus and the C57Bl/6 versions highlights many variations: 14 missense changes in the different isoforms and at least 16 UTR variants (
Table S1). These variations could be responsible for important modifications of the structure and/or the function of the
spretus SPATA3 protein compared to the C57Bl/6 version. This
spretus version of the protein could therefore be inapt to interact/bind/function with any receptor/cofactor/partner in the C57Bl/6 version in the IRCS model to accomplish its normal activity, resulting in the hypofertility phenotype observed in the Rc3 IRCS male mice [
7].
We therefore decided to produce a mouse line invalidated for the Spata3 gene in order to decipher the function of this gene. Though male mice lacking Spata3 expression are fully fertile in vivo, their fertility in vitro is drastically affected and some ultrastructural anomalies can be observed in sperm samples.
3. Discussion
In order to understand which gene within the
Mafq1 QTL interval was responsible for the hypofertility phenotype observed in the Rc3 IRCS mice [
7], we explored the list of candidate genes present in the interval.
Spata3 matched all potential criterion of selection: the specific expression profile, regarding both the timing and location in differentiating sperm cells, the evolutionary conservation, the high number and predicted impact of missense changes between B6 and
spretus SPATA3 proteins. The lack of information about its function left all hypotheses open. We therefore decided to generate a
Spata3 null mouse to determine if the invalidation of the
Spata3 gene could mimic the phenotype of the Rc3 IRCS mouse.
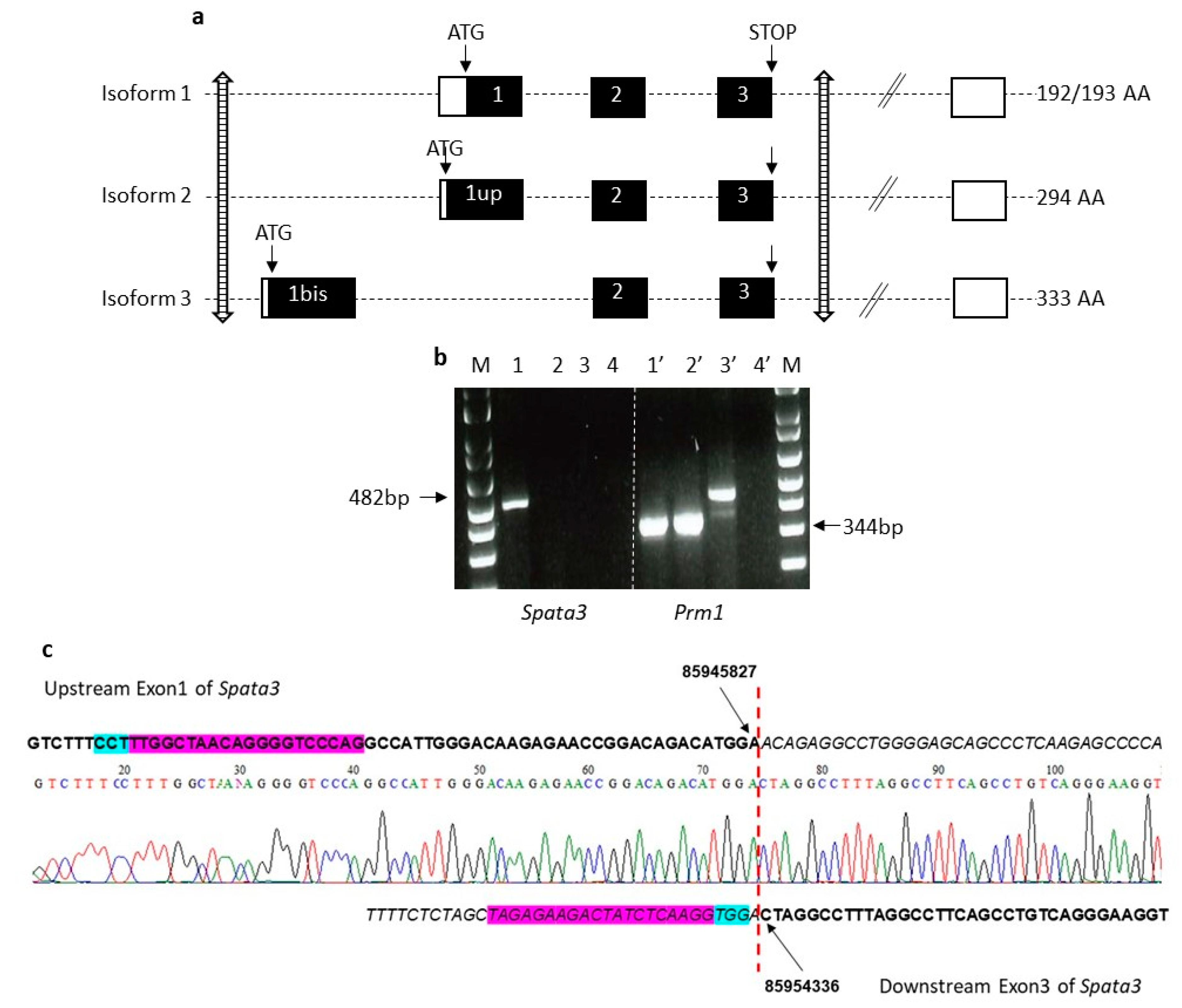
When this project was initiated, the reported structure of the
Spata3 gene was the one reproduced in
Figure 2a, as isoform 1, including 3 coding exons (Exons 1, 2 and 3) and additional variable 3′UTR exons. A CRISPR/Cas9 strategy had been developed to target and affect exon 1 with RNA guides, therefore compromising the full ORF. Two independent KO lines were produced but they were found to be fully fertile in vivo (data not shown).
However, in 2018, databases revealed the existence of so far unsuspected isoforms of the gene. First, an additional isoform starts upstream of the reported translation start, at an ATG located 303 bp earlier, in phase (“exon 1 upstream”,
Figure 2a, isoform 2). Second, a new alternative isoform includes a new upstream first exon (“exon 1 bis”,
Figure 2a, isoform 3), followed by the classic coding exons 2 and 3, in phase. This new messenger therefore encodes a protein that shares the last 92 amino acids but completely differs from the initially known form at the
N-terminus and is not affected by the absence of the “classic” exon 1. This gene organization is conserved, while isoforms 1 and 3 are present in mice and humans. In 2019, Zhou et al. reported new isoforms of the human
SPATA3 gene [
15]. They first identified a new isoform skipping exon 2, leading to a protein of 138 amino acids whose C-terminal end is different because of a frameshift, but whose abundance is unknown. They also described an alternative splicing within exon 1, where 27 bp would be spliced away. However, this is rather a documented polymorphism, referenced as rs 72362780, with different alleles of either 2 or 3 repeats of a 27 bp sequence encoding a QQPSPESTP motif, being almost equally frequent in the human population.
The initial knock-out was therefore considered as a partial KO, suppressing only some of the possible isoforms and probably leaving some not negligible residual SPATA3 activity. Another knock-out strategy was therefore adopted to delete all known coding exons. RNA guides located upstream exon 1bis and downstream exon 3 were used to generate a complete gene deletion of about 8 kb. Two independent KO lines were obtained for this complete invalidation that gave identical results. Sequencing of the locus confirmed the deletion of the coding exons and validated the precise breakpoints. The lack of expression of the
Spata3 cDNA was confirmed by RT-PCR, in the absence of an efficient antibody to detect the SPATA3 protein by Western blot. When mated with WT females, homozygous KO males could sire normally and produce normal litters. The absence of
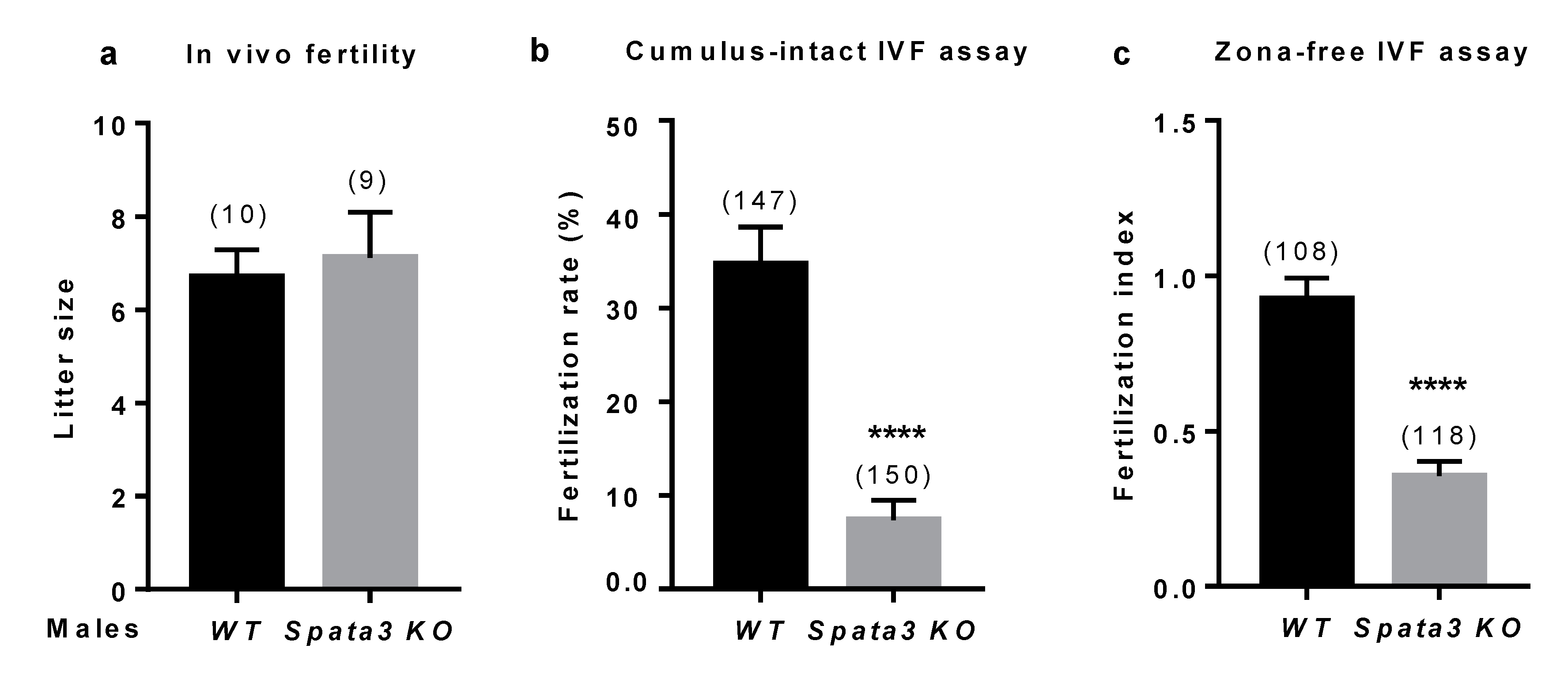
Spata3 did not reproduce the in vivo hypofertility observed in Rc3 males [
7]. However, in IVF assays, sperm lacking
Spata3 expression were less efficient to fertilize WT oocytes. Both the fertilization rate of cumulus-intact oocytes and the fertilization index of zona-free oocytes were drastically reduced. This difference of phenotype between in vivo and in vitro conditions has already been observed. It is probably due to the differences between an optimal in vivo fertilization and the artificial context of IVF where some parameters are not ideally reproduced. Thus, IVF could exacerbate a defect that is compensated for under physiological conditions. This observation has already been encountered for the sperm specific
Itgb1 KO [
16] and for the mouse heterozygous for a
Spaca6 deletion [
17]. This reduction of IVF efficiency was also detected in Rc3 males.
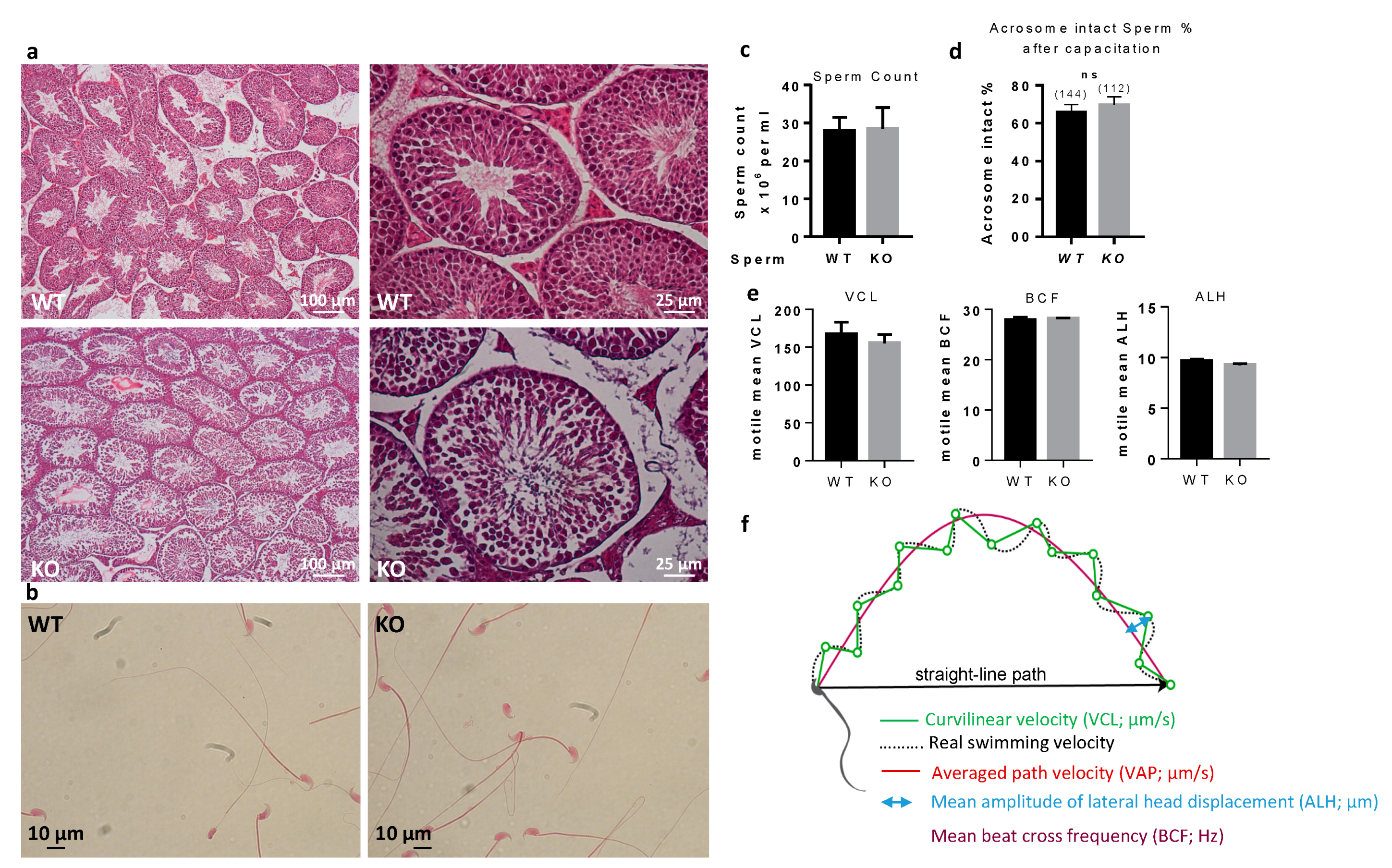
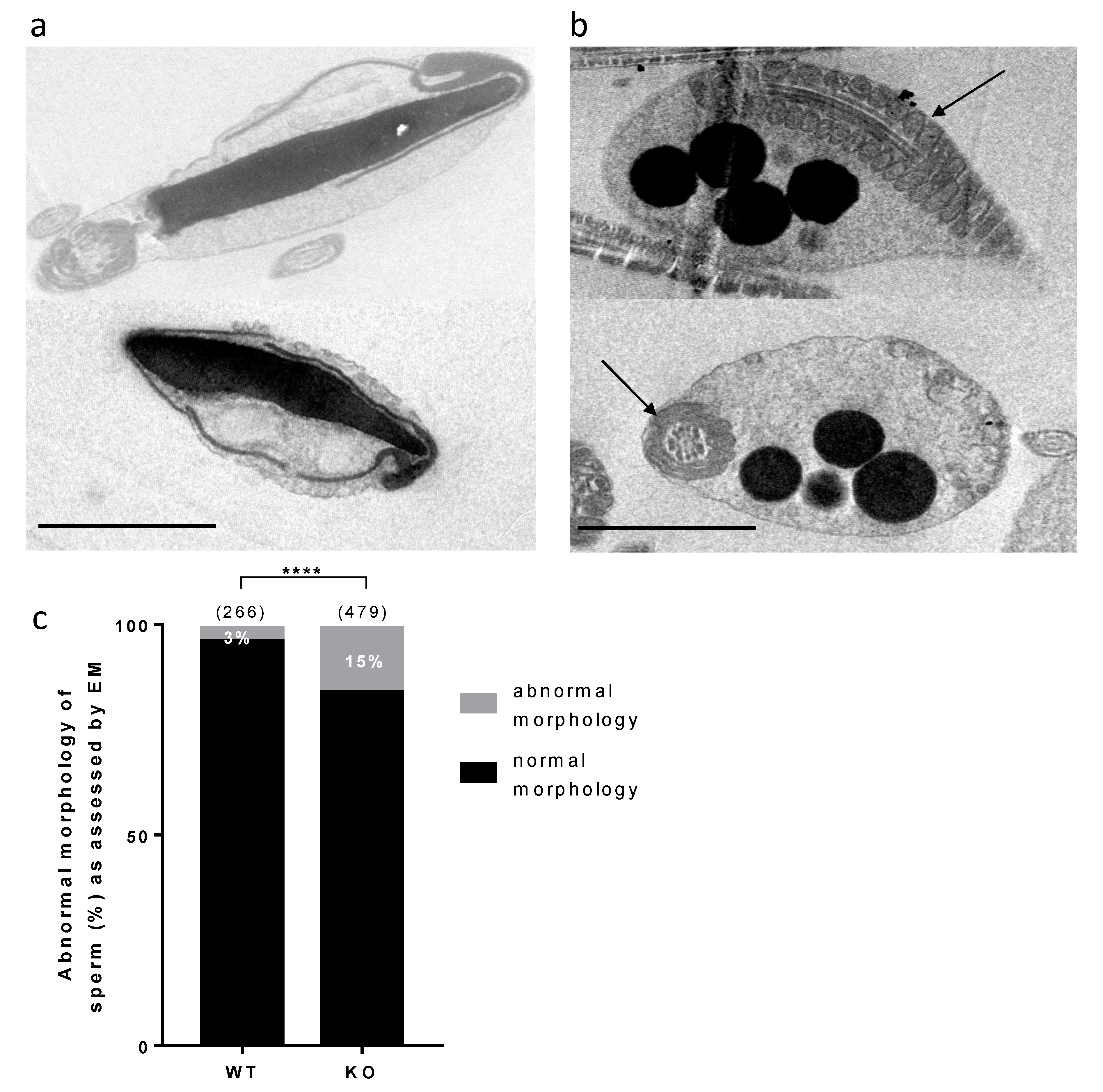
No gross abnormality could be detected in the testicular histology neither in sperm counts, suggesting a normal spermatogenesis. The sperm movement parameters explored by CASA were not affected. A reduction of the motile fraction had been observed in Rc3 males but here the decrease in IVF yield cannot be explained by a reduced motility of sperm. Sperm ultrastructure was explored by electronic microscopy to observe more subtle sperm alterations, as were reported in the Rc3 IRCS model [
7]. Some abnormal sperm could be observed, mainly affecting the acrosome. As observed in the Rc3 line, an acrosome detached from the nucleus is more frequently reported in KO than in WT sperm. The acrosome sometimes looks out of shape, far from the expected symmetric cap aspect. It is very likely that the totally abnormal appearance of the sperm shown in
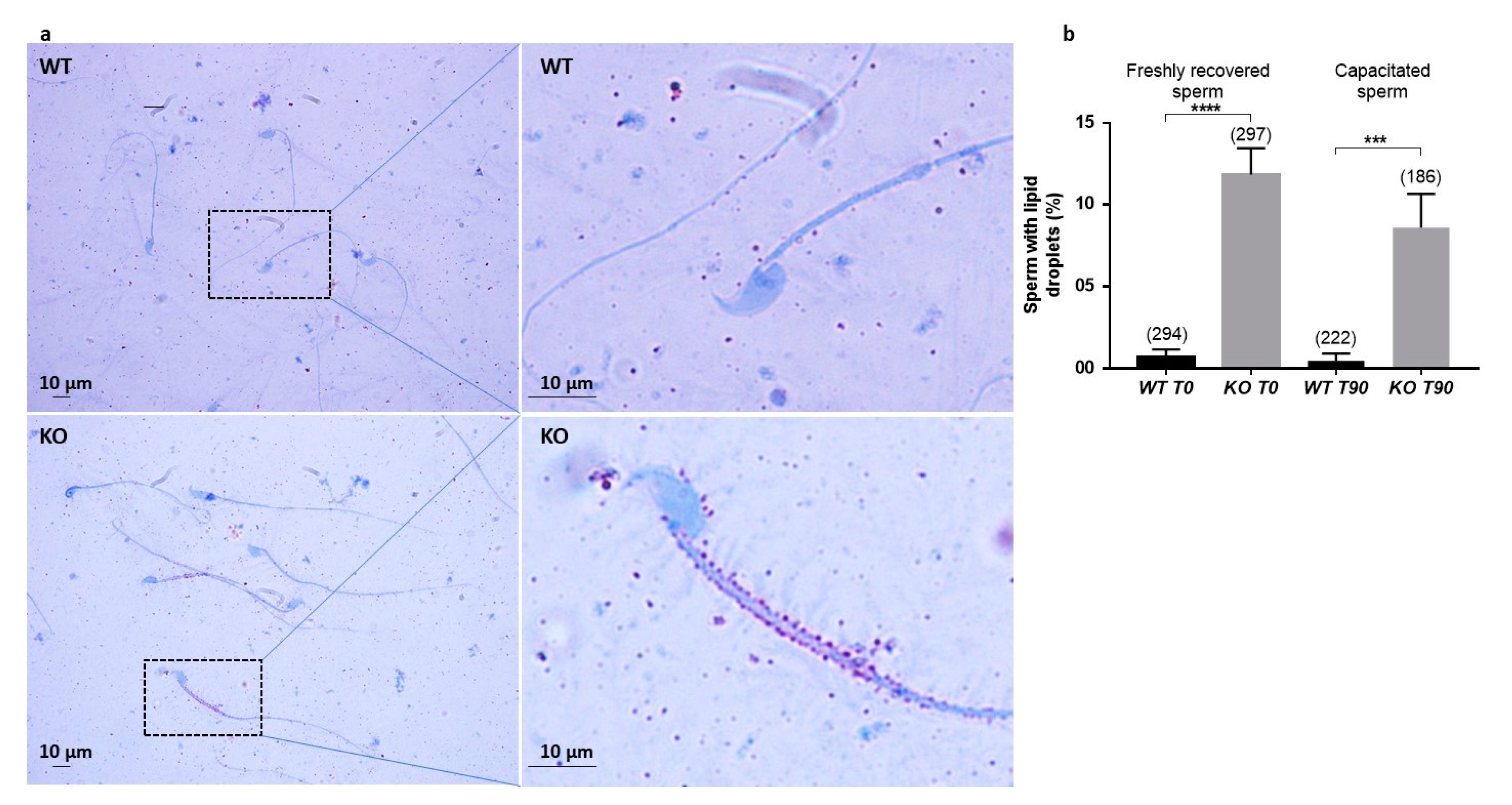
Figure 5a is not really the ejaculated sperm phenotype but rather the result of manipulations which further damage the structure of these KO sperm compared to WT sperm. It is anyway a sign of a particular fragility of KO sperm. In addition, dark droplets in persistent cytoplasmic remnants were also visible. An Oil Red O coloration was performed and confirmed the lipid nature of these droplets contents and that they are much more present in KO sperm. These cytoplasmic bodies should have been removed by Sertoli cells in the last steps of spermiogenesis. It is not clear if the phagocytosis process is altered in KO testes, but SPATA3 has been shown to be expressed in germ cells, not in Sertoli cells. Alternatively, a dysfunction of lipid metabolism could be present in KO sperm, leading to an excess of lipid droplets that fail to be correctly phagocyted by Sertoli cells. Why this process of lipids accumulation happens will need more analyses. It is difficult to estimate exactly what percentage of sperm is abnormal, as, more than 10% showed lipid droplets, 15% appeared fully disorganized at the acrosome level, and some others showed milder defects.
In conclusion, male mice lacking the expression of the Spata3 gene are fertile in vivo, thanks to the optimal context of sperm selection and gamete interaction. They are nevertheless hypofertile in vitro. Moreover, they have a higher frequency of abnormal sperm. Specifically, they have more abnormal acrosomes that tend to untie from the nucleus and more cytoplasmic bodies showing an imperfect spermiogenesis and spermiation.
The Spata3 KO, though modestly affecting sperm capacities, is therefore an incomplete reproduction of the Rc3 IRCS mouse line, that was considered as a model of partial globozoospermia. Two hypotheses can be proposed. It is possible that the complete suppression of SPATA3 protein expression in the invalidated mouse on one side and the expression of a modified SPATA3 protein in the IRCS on the other have different impacts on sperm function. The presence of a deleterious protein could be more damaging than its absence, if for example, some compensatory mechanisms can set up. Some of the polymorphisms existing between spretus and C57Bl/6 strains are indeed individually predicted to affect protein function. However, no prediction model can aggregate the effect of the 14 non-synonymous changes on the integrity of the different protein isoforms, especially on a protein of unknown function. The alternative hypothesis is that another gene within the Mafq1 interval contributes to the complex phenotype of the Rc3 males. One or more gene(s) present in the QTL interval and exhibiting variations in the spretus strain compared to the C57Bl/6 strain, likely missense changes affecting protein function but also possibly regulatory variants affecting protein levels, could also be dysfunctional in the Rc3 line. The cumulative effects of two or more disturbed genes could be necessary to affect more drastically sperm function, either directly if proteins are acting together or indirectly if they have different functions whose anomalies add at different stages or pathways to generate the defects profile at the end. A careful re-analysis of the identity and characteristics of the genes present in the Rc3 interval could suggest other candidates that could participate in the phenotype.
On the human side, SPATA3 and other potential candidate genes from the QTL region should be screened in infertile/hypofertile patients with signs reminiscent of globozoospermia. Furthermore, genes identified in this QTL region will be particularly closely analyzed in our future whole exome sequencing studies of infertile patients.
4. Materials and Methods
4.1. Animals
All animal experiments were performed in agreement with national guidelines for the care and use of laboratory animals. Experimentation was approved by the local ethics committee (C2EA-34, Comité d’éthique en matière d’expérimentation animale Paris Descartes) and the governmental ethical review committees via the APAFiS Application (Autorisation de projet utilisant des animaux à des fins scientifiques), under the registration reference APAFIS #14124-2017072510448522 v26, A. Ziyyat (10/102018-10/10/2023).
Mice were purchased from Janvier Labs (Le Genest-Saint-Isle, France). All animals were maintained at the animal facility of the Cochin Institute (Paris) at a stable temperature (21–23 °C) and 14 h light/10 h dark photoperiods, with free access to food and water.
4.2. Generation of Transgenic Mice
The Crispr/Cas9 approaches were performed at the Transgenesis and Homologous Recombination platform (PRHTEC) of the Cochin Institute, Paris, France. RNA guides were selected, produced by TACGENE (Muséum d’Histoire Naturelle, Paris, France) and tested in vitro (targeting upstream exon 1 bis: ACCGGACAGACATGGAACAG; GCTGGGACCCCTGTTAGCCAA; GTTGTGACATGCTCCAGGGAA- targeting downstream exon 3: GGACCCCAGGCCCACCGTC; GTAGAGAAGACTATCTCAAGG; GGCCCACCGTCCGGCAGGT).
They were preincubated with Cas9 and then microinjected into fertilized C57Bl/6J oocytes.
Embryos that had passed to the 2-cell stage were transferred into the oviducts of pseudopregnant females.
4.3. PCR and RT-PCR
Genotyping was performed on DNA extracted from tail biopsies (NucleoSpin
® Tissue, Macherey-Nagel, Düren, Germany) using the GoTaq Flexi polymerase (Promega, Madison, WI, USA) under standard PCR conditions. Primers are listed in
Table S2. The integrity of the areas targeted by the guides was tested to detect WT alleles. A large amplification encompassing the whole locus was also performed to detect deleted alleles.
RNA was obtained by TRIzol (Thermo Fischer Scientific, Waltham, MA, USA) extraction from WT and KO testes. Five micrograms of total RNA were treated with DNAse I (Promega) and retrotranscribed using the MMLV reverse transcriptase (Thermo Fischer Scientific) following manufacturer’s instructions. RT-PCR was performed using the GoTaq Flexi polymerase to test the expression of Spata3 and a reference gene as normalization control.
4.4. In Vivo and In Vitro Fertility Assessment
Mice, aged from 8 to 12 weeks, were housed as one male and one female per cage. WT females were mated overnight with WT males or homozygous KO males and checked for vaginal plugs the next day. For each group, litter sizes were assessed.
Wild-type 5–8 weeks-old female mice were superovulated with a 10 IU pregnant mare serum gonadotrophin (PMSG) injection, then 48 h later with a 10 IU human chorionic gonadotrophin (hCG) injection (Intervet, Beaucouzé, France). The next day, 14 h post-injection, animals were sacrificed by cervical dislocation. Cumuli oophori were retrieved by tearing the ampulla wall of the oviduct, placed in Ferticult medium (FertiPro, Beernem, Belgium) supplemented with 3% bovine serum albumin (BSA), and maintained at 37 °C under 5% CO2 in air, under mineral oil (Sigma Aldrich, Saint Quentin Fallavier, France).
For zona-free IVF assay, oocytes were released from the cumulus cells by 3–5 min incubation at 37 °C with hyaluronidase (Sigma Aldrich) in M2 medium (Sigma Aldrich), rinsed and kept in Ferticult medium, BSA 3% at 37 °C under a 5% CO2 atmosphere with mineral oil (Sigma Aldrich). The zona pellucida was then dissolved with acidic Tyrode’s (AT) solution (pH 2.5, Sigma Aldrich) under visual monitoring. Zona-free eggs were rapidly rinsed in a Ferticult medium, BSA 3% and kept at 37 °C under a 5% CO2 atmosphere for 2 to 3 h to recover.
Mouse spermatozoa were obtained from the cauda epididymis of control or KO male mice aged 8 to 13-weeks. They were capacitated at 37 °C under a 5% CO2 for 90 min in a 500 µL drop of Ferticult medium supplemented with 3% BSA (30 mg/mL), under mineral oil.
Cumulus-intact or zona-free oocytes were inseminated in a 100 µL drop of Ferticult medium, BSA 3% with capacitated spermatozoa at a final concentration of 1×106/mL or 1×105/mL, respectively, for 3 h. They were then washed and directly mounted in Vectashield/DAPI (Vector Laboratories, Burlingame, CA, USA) for observation under UV light (Zeiss Axioskop 20 microscope, Marly le Roi, France). Oocytes were considered fertilized when a fluorescent decondensed sperm head could be visualized within their cytoplasm.
4.5. Histology, Staining and Immunostaining of Sperm and Testes
Testes from adult WT and KO males were collected. They were fixed overnight in 4% (w/v) paraformaldehyde in PBS, dehydrated and embedded in paraffin. Serial 5 µm thick sections were cut on a microtom and stored at room temperature. Testis sections were rehydrated before Hematoxylin-eosin (H&E) staining.
For Immunofluorescence staining, slides were permeabilized during 10 min in PBS 1× and with triton 0.2% at RT, and were then saturated for 30 min in PBS, BSA 3% and Goat serum 10%. Slides were incubated for 1 h in a polyclonal anti-TSARG1 antibody at 1:20 in PBS (T-18, sc-67901, Santa Cruz Biotechnology, Dallas, TX; reference no longer available), washed three times in PBS, BSA 1%. DAPI (5 µg/mL, Sigma Aldrich) and secondary antibody (anti-Goat-Alexa 488 at 10 µg/mL, ThermoFischer Scientific) were used at RT during 1 h, followed by three washes and slides were mounted with the Dako Fluorescent mounting medium (Agilent, Santa Clara, CA, USA).
For Papanicolaou staining, sperm were retrieved from cauda epididymes in Ferticult medium and spread onto a Superfrost Plus slide (ThermoFischer Scientific). Sperm cells were fixed by incubation with PBS, paraformaldehyde 4% for 10 min and stained following the Papanicolaou protocol (Hematoxylin, OG6, EA50). Briefly, the slides were washed in 95% ethanol and inserted in Harris hematoxylin for 3 min for nucleus counterstaining, and washed, stained with OG-6 dye (RAL diagnostics 05.12013, Martillac, France) and with EA-50 (RAL diagnostics 05.12019, Martillac, France), Then the slides were dehydrated (95% ethanol absolute ethanol and xylene) and mounted with a permanent mounting medium.
Freshly recovered or capacitated sperm were washed in PBS containing 1% BSA, centrifuged at 300 g for 5 min and immediately fixed in 4% Paraformaldehyde (Electron Microscopy Sciences, PA, USA) in PBS-1% BSA for 10 min at room temperature. In order to detect the sperm acrosomal status, after washing, fixed spermatozoa were stained with FITC-conjugated lectin Pisum Sativum Agglutinin (PSA; 25 μg/mL in PBS, Sigma Aldrich) for 15 min. After repeated washing with PBS-BSA 1%, a drop of sperm suspension was smeared on a slide, air-dried, mounted with Vectashield-DAPI. Detection was performed using a Nikon Eclipse E600 microscope Zeiss Axiophot epifluorescence microscope and images were digitally acquired with a camera (Coolpix 4500, Nikon, Champigny sur Marne, France).
For Oil Red O (ORO) staining, a drop of freshly recovered or capacitated sperm was smeared on a slide, air-dried and immediately fixed in PFA 4% and stored at −80 °C until the day of use. Slides were post-fixed for 10 min in PFA 4%, then rinsed and quenched with NH4Cl 50 mM for 10 min. After rinsing, cells were permeabilized for 4 min with triton X-100 0.5%. Finally, cells were stained by an incubation of one hour with Oil-Red-O (Sigma Aldrich) and counter-stained with hematoxylin of Mayer. Lipids appeared as red droplets and nuclei appeared to be blue. The amount of lipid droplets by ORO staining in sperm cells was quantified using a light microscope.
4.6. Mouse Sperm Motility Analysis
Sperm motility was assessed by Computer Aided Sperm Analysis (CASA) using CEROS II apparatus (Hamilton Thorne, Beverly, MA, USA). Briefly, mouse spermatozoa were obtained from the cauda epididymis of Spata3 KO or WT C57BL6/J mice (8 to 10 weeks old) and capacitated at 37 °C under 5% CO2 for 90 min in a 500 μL drop of Ferticult medium supplemented with 3% BSA, under mineral oil. The movements of at least 500 sperm cells per sample were analyzed in 20 µm chambers (Leja Products B.V., Nieuw-Vennep, Netherlands) with Zeiss AX10 Lab. A1 microscope (10× objective), using HT CASAII software.
The settings were as follows: acquisition rate, 60 Hz; number of frames, 45; minimum head brightness, 175; minimum tail brightness, 80; minimum head size, 10 µm2; minimum elongation gate, 1%; maximum elongation gate, 100%; objective magnification factor, 1.2.
The principal motility parameters measured were: curvilinear velocity (VCL), average path velocity (VAP), straight-line velocity (VSL), beat/cross frequency (BCF), amplitude of lateral head displacement (ALH).
4.7. Electronic Microscopy
Mouse spermatozoa from males were prepared as described above (IVF) and fixed by incubation in PBS supplemented with 2% glutaraldehyde (Grade I, Sigma) for 2 h at room temperature. Samples were washed twice in PBS and post-fixed by incubation with 1% osmium tetroxide (Electron Microscopy Sciences), after which they were dehydrated by immersion in a graded series of alcohol solutions and embedded in Epon resin (Polysciences Inc.,Warrington, PA, USA). Ultra-thin sections (90 nm) were cut with a Reichert Ultracut S ultramicrotome (Reichert-Jung AG, Wien, Austria) and were then stained with uranyl acetate and lead citrate. Sections were analyzed with a JEOL 1011 microscope and digital images were acquired with a Gatan Erlangshen CCD camera and Digital Micrograph software. The integrity of sperm organelles was checked including the acrosome, nucleus, flagellum, etc. Acrosomes with a space between the nuclear membrane and the inner acrosomal membrane were considered abnormal. Acrosomes with membranes in close contact were considered normal. The presence of residual bodies and lipid droplets was recorded.
4.8. Statistical Analysis
Results are expressed as mean ± SEM of at least three independent experiments. For statistical analysis, t-Test was performed using GraphPad Prism version 7.00 for Windows, (GraphPad Software, La Jolla, CA, USA). Differences were considered statistically significant when p < 0.05. When data were not normally distributed, a Welch’s correction was applied.


 and
and 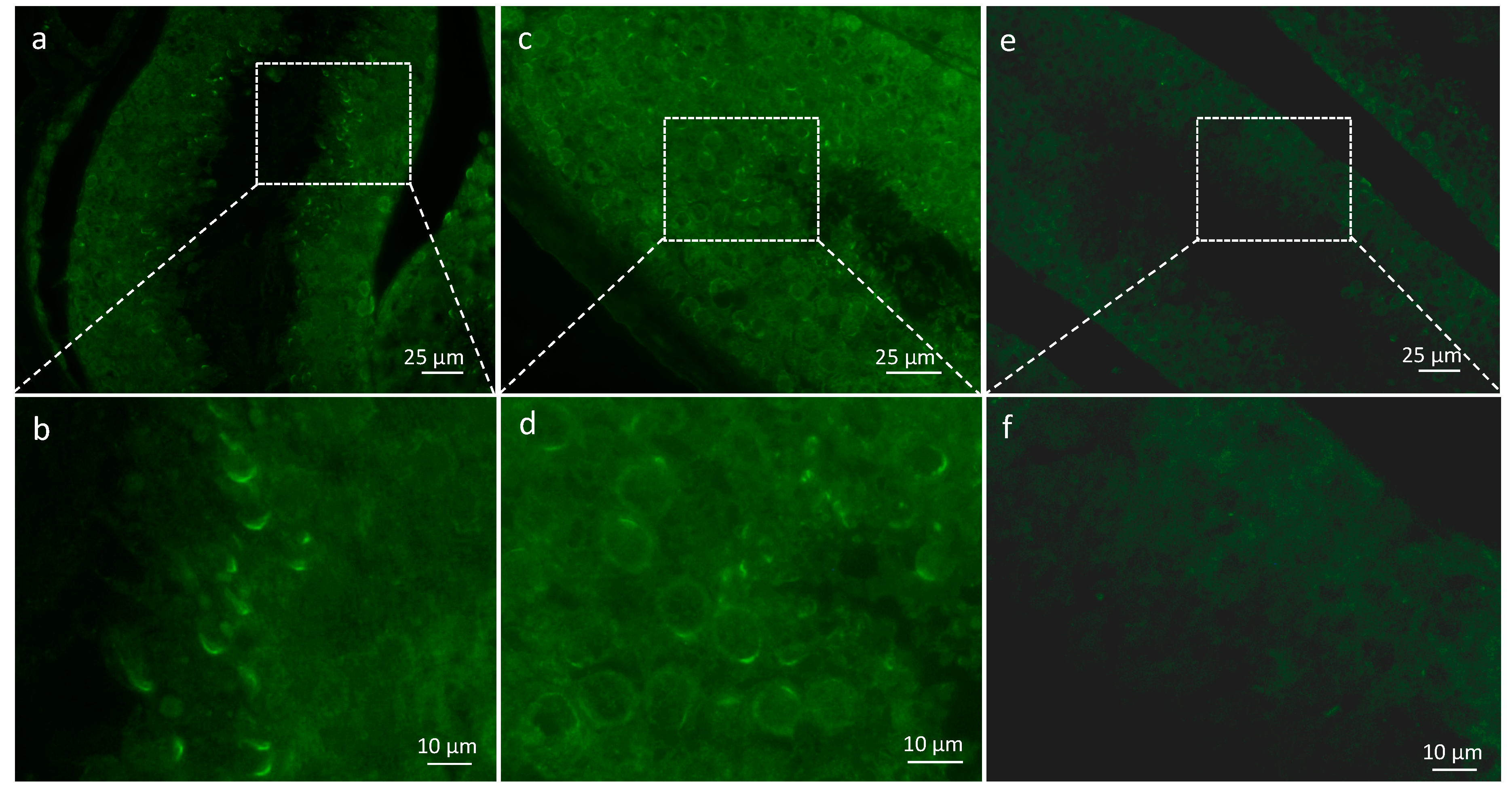{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





