
Na+ and/or Cl− Toxicities Determine Salt Sensitivity in Soybean (Glycine max (L.) Merr.), Mungbean (Vigna radiata (L.) R. Wilczek), Cowpea (Vigna unguiculata (L.) Walp.), and Common Bean (Phaseolus vulgaris L.)


 and
and
Abstract
1. Introduction
2. Results
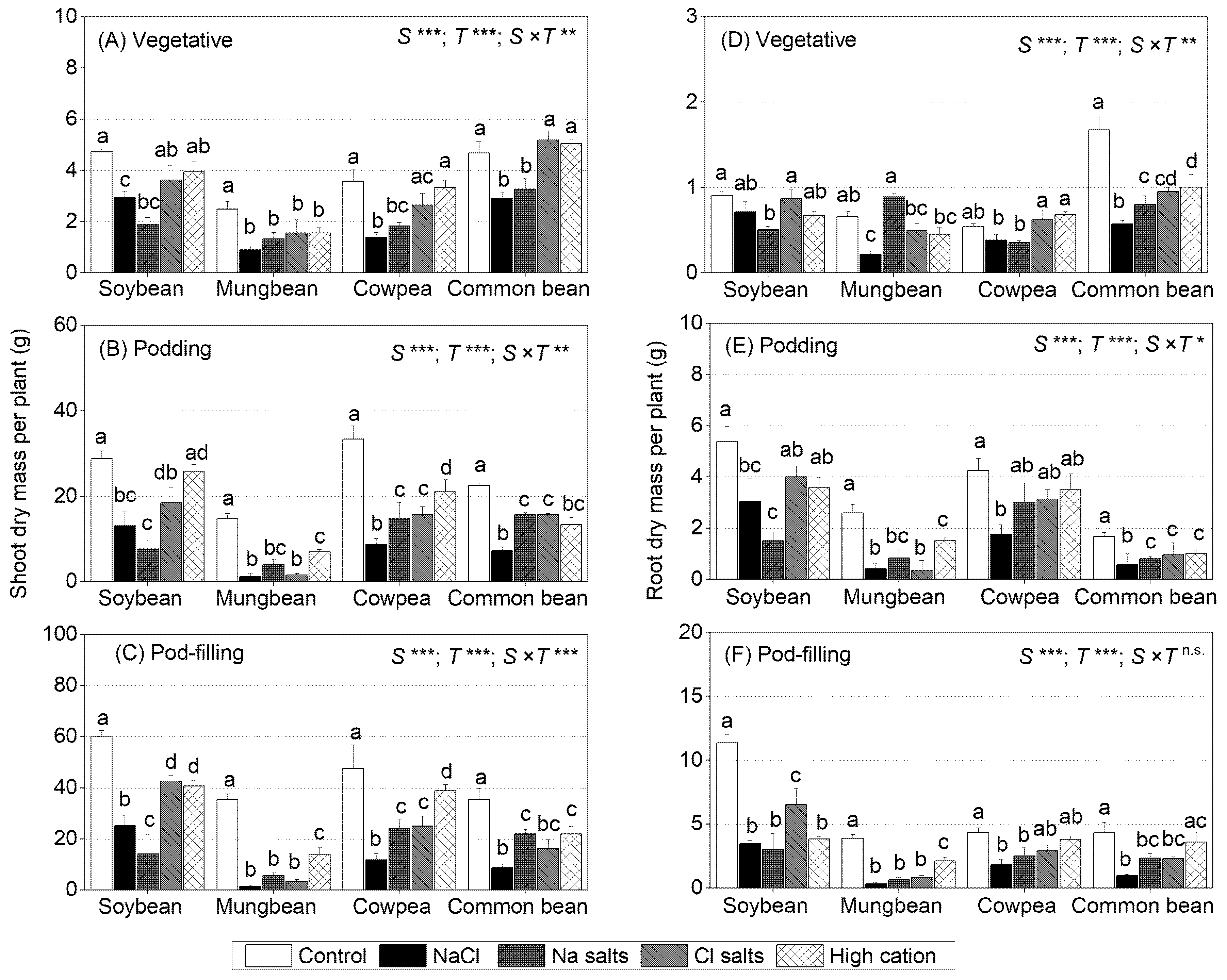
2.1. Treatment Effects on Shoot and Root Dry Mass at Different Growth Stages
2.2. Treatment Effects on Plant Reproductive Attributes
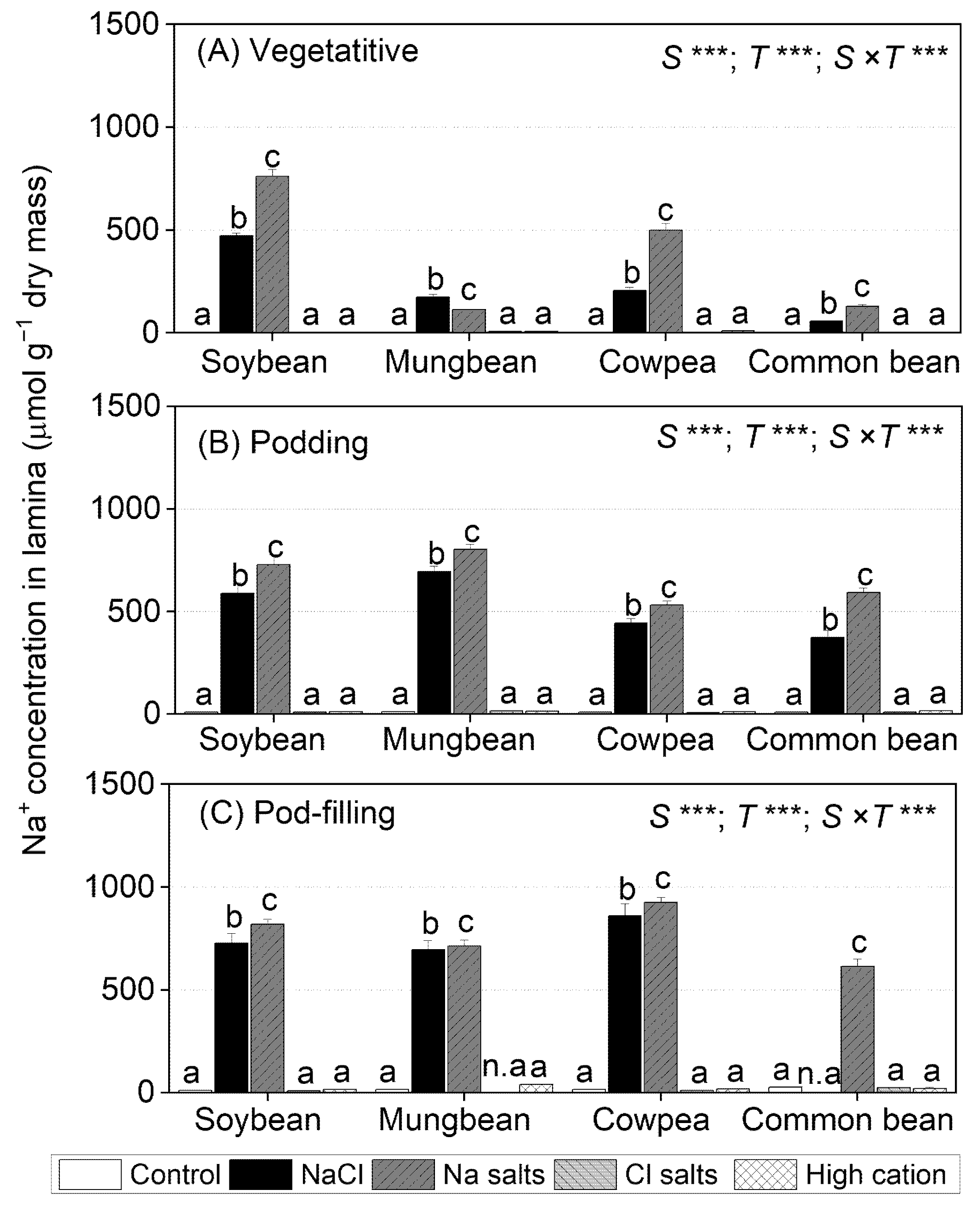
2.3. Treatment Effects on Na+ Concentrations in Tissues
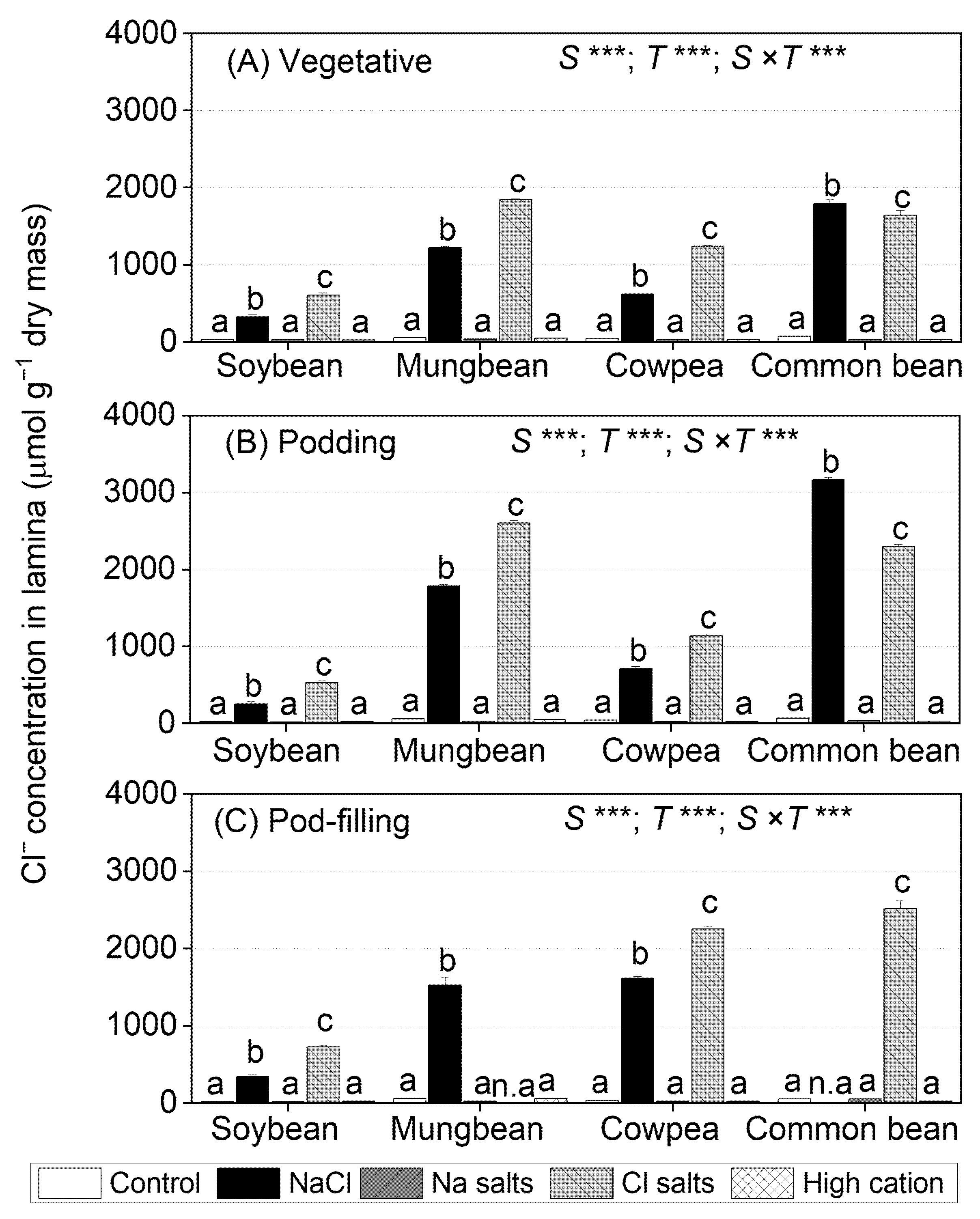
2.4. Treatment Effects on Cl− Concentrations in Tissues
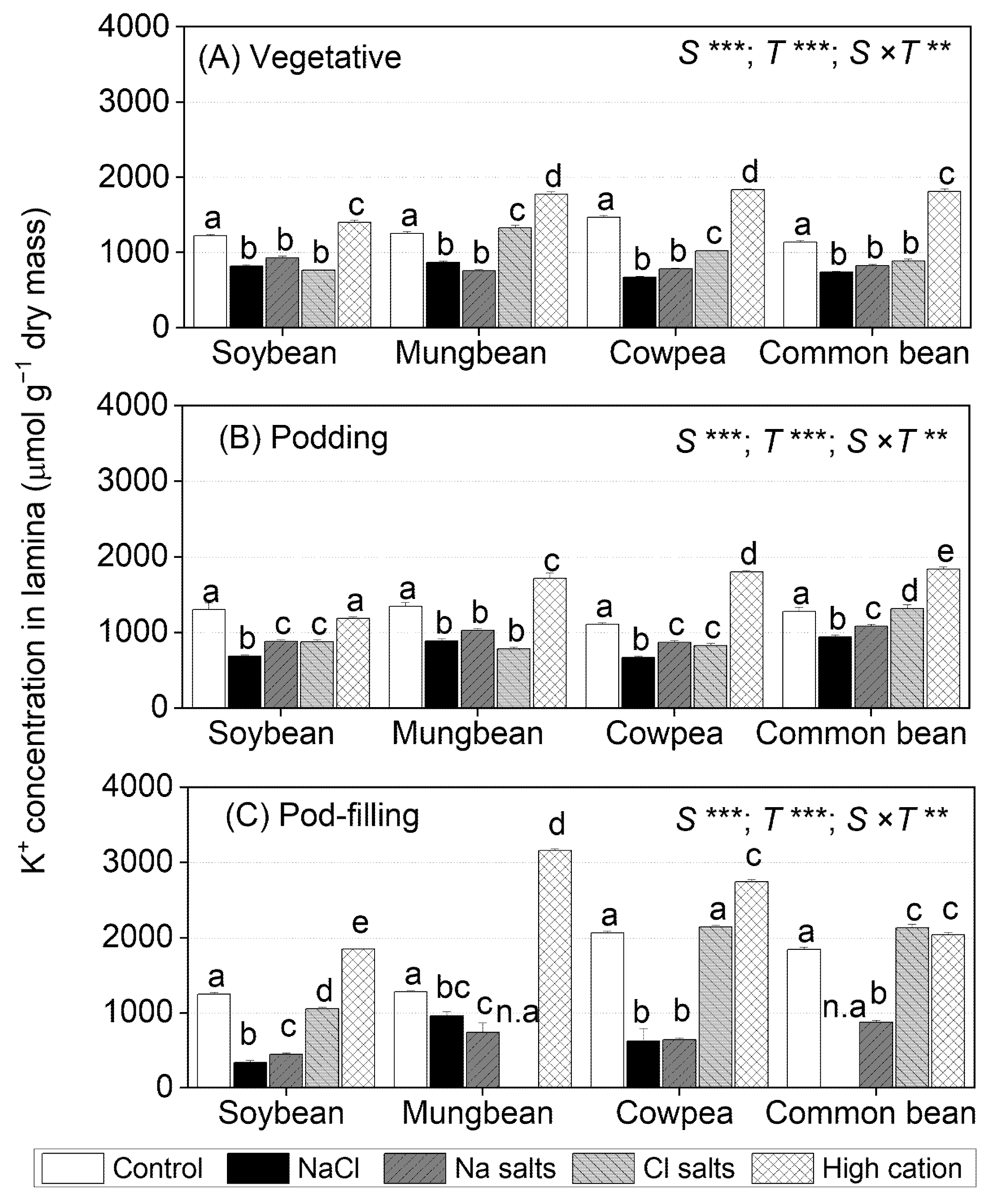
2.5. Treatment Effects on K+ Concentrations in Tissues
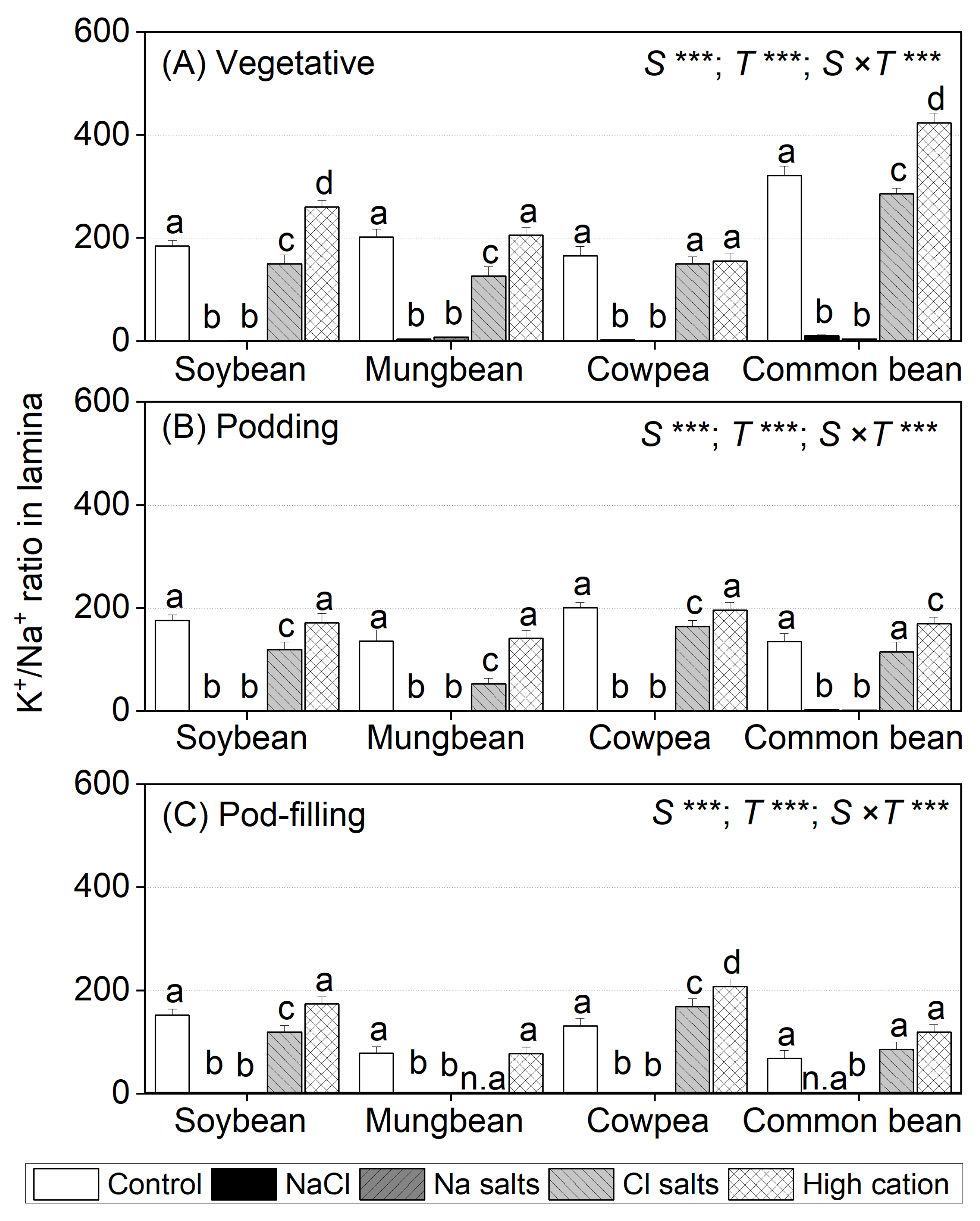
2.6. Treatment Effects on the K+/Na+ Ratio in Tissues
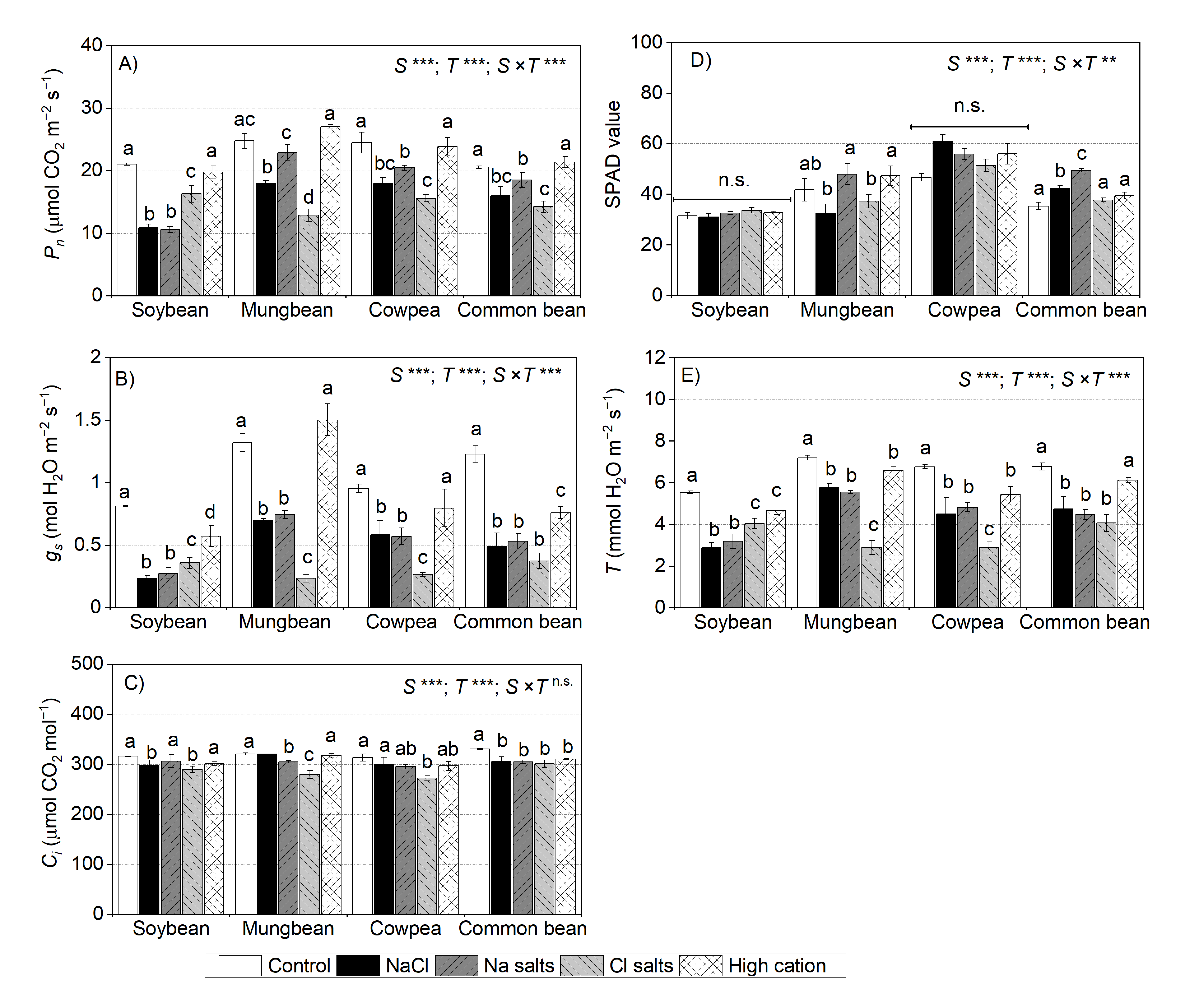
2.7. Treatment Effects on Leaf Gas Exchange at the Vegetative Stage
2.8. Treatment Effects on Leaf Sap Osmotic Potential (Ψπ)
2.9. Relationships of Shoot Dry Mass with Shoot Ion Concentrations
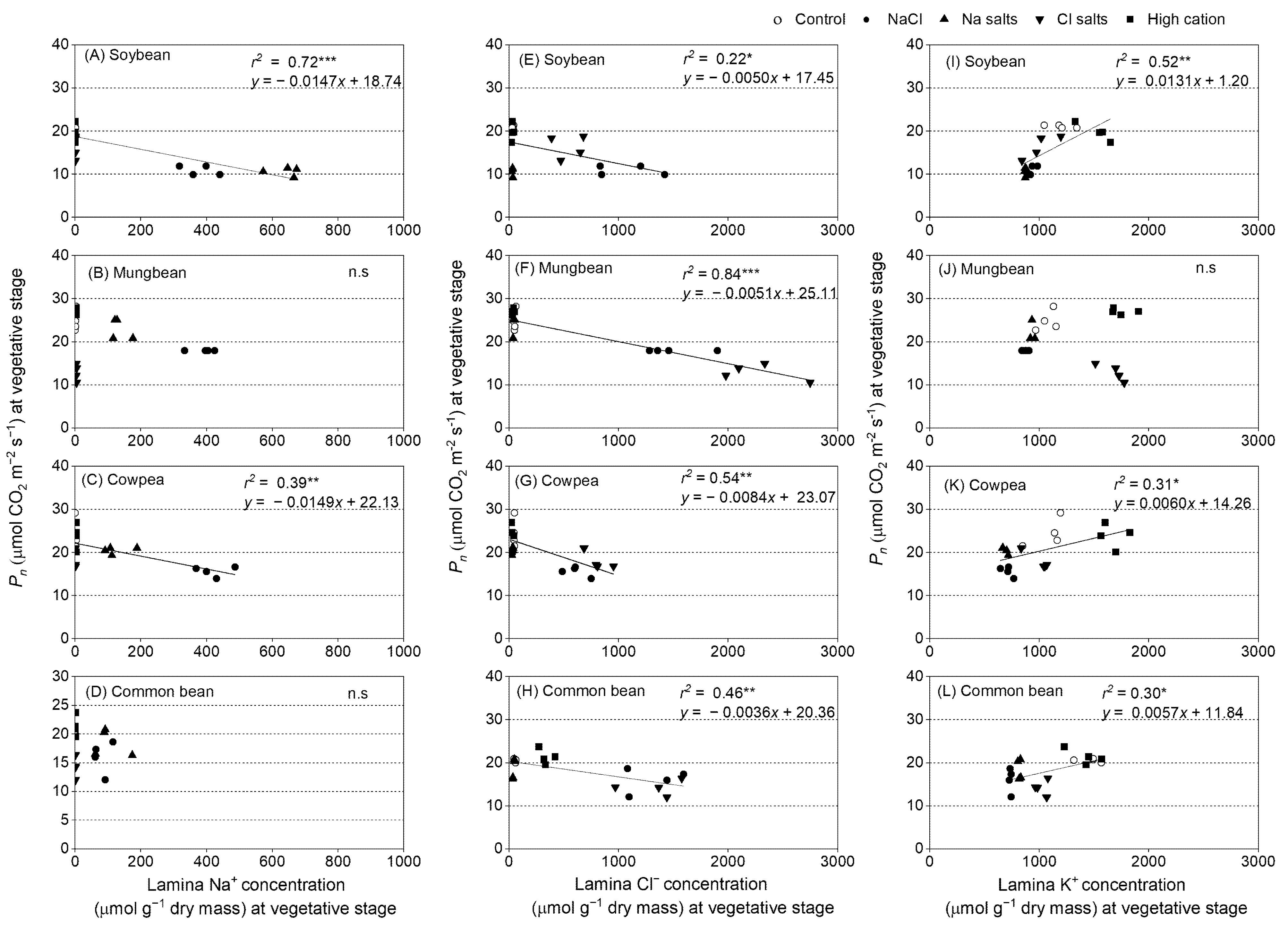
2.10. Relationships of Net Photosynthetic Rate (Pn), Stomatal Conductance (gs), Intercellular CO2 (Ci), and Transpiration Rate (T) with Leaf Ion Concentrations
3. Discussion
3.1. Soybean Is More Sensitive to High Na+ Than High Cl−
3.2. Mungbean Is More Sensitive to High Cl− Than High Na+
3.3. Cowpea Is Sensitive to Both High Na+ and High Cl−
3.4. Common Bean Is Sensitive to Both High Cl− and to High Na+
3.5. Osmotic Contribution to Salinity Tolerance
3.6. Soybean Is More Salinity Tolerant Than the Three Other Grain Legume Species
3.7. Shoots Are More Sensitive to Salinity Stress Than Roots (Three of the Four Species), and the Vegetative Stage Is Less Sensitive to Salinity Than the Reproductive Stage
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Treatments
4.3. Plant Samplings and Measurements
4.4. Leaf Gas Exchange and Chlorophyll Measurements
4.5. Tissue Ion Analysis
4.6. Organic Solutes (Sugars and Sugar Alcohols)
4.7. Leaf Sap Osmotic Potential and Tissue Water Content
4.8. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Messina, M.J. Legumes and soybeans: Overview of their nutritional profiles and health effects. Am. J. Clin. Nutr. 1999, 70, 439–450. [Google Scholar] [CrossRef] [PubMed]
- Shewry, P.R.; Halford, N.G. Cereal seed storage proteins: Structures, properties and role in grain utilization. J. Exp. Bot. 2002, 53, 947–958. [Google Scholar] [CrossRef] [PubMed]
- Sandstrom, B.; Almgren, A.; Kivisto, B.; Cederblad, A. Effect of protein level and protein source on zinc absorption in humans. J. Nutr. 1989, 119, 48–53. [Google Scholar] [CrossRef]
- Fageria, N.K. Green manuring in crop production. J. Plant Nutr. 2007, 30, 691–719. [Google Scholar] [CrossRef]
- Peoples, M.B.; Herridge, D.F.; Ladha, J.K. Biological nitrogen fixation: An efficient source of nitrogen for sustainable agricultural production? Plant Soil 1995, 174, 3–28. [Google Scholar] [CrossRef]
- Graham, P.H.; Vance, C.P. Legumes: Importance and constraints to greater use. Plant Physiol. 2003, 131, 872–877. [Google Scholar] [CrossRef]
- Ahmed, S.; Hasan, M. Legumes: An overview. J. Pharm. Pharm. Sci. 2014, 2, 34–38. [Google Scholar]
- Keating, B.A.; Fisher, M.J. Comparative tolerance of tropical grain legumes to salinity. Aust. J. Agric. Res. 1985, 36, 373–383. [Google Scholar] [CrossRef]
- Flowers, T.J.; Gaur, P.M.; Gowda, C.L.L.; Krishnamurthy, L.; Samineni, S.; Siddique, K.H.M.; Turner, N.C.; Vadez, V.; Varshney, R.K.; Colmer, T.D. Salt sensitivity in chickpea. Plant Cell Environ. 2010, 33, 490–509. [Google Scholar] [CrossRef] [PubMed]
- Deinlein, U.; Stephan, A.B.; Horie, T.; Luo, W.; Xu, G.; Schroeder, J.I. Plant salt-tolerance mechanisms. Trends Plant Sci. 2014, 19, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, P.; Prasad, M.N.V. Abiotic Stress Responses in Plants: Metabolism, Productivity and Sustainability; Springer: New York, NY, USA, 2011. [Google Scholar]
- Munns, R. Physiological processes limiting plant growth in saline soils: Some dogmas and hypotheses. Plant Cell Environ. 1993, 16, 15–24. [Google Scholar] [CrossRef]
- Kotula, L.; Khan, H.A.; Quealy, J.; Turner, N.C.; Vadez, V.; Siddique, K.H.M.; Clode, P.L.; Colmer, T.D. Salt sensitivity in chickpea (Cicer arietinum L.): Lons in reproductive tissues and yield components in contrasting genotypes. Plant Cell Environ. 2015, 38, 1565–1577. [Google Scholar] [CrossRef]
- Khan, H.; Siddique, K.; Colmer, T. Salt sensitivity in chickpea is determined by sodium toxicity. Planta 2016, 244, 623–637. [Google Scholar] [CrossRef]
- Ayoub, A.T.; Ishag, H.M. Sodium toxicity and cation imbalance in dry beans (Phaseolus vulgaris L.). J. Agric. Sci. 1974, 82, 339–342. [Google Scholar] [CrossRef]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Salim, M.; Pitman, M.G. Salinity tolerance of mungbean (Vigna radiata L.): Seed production. Biol. Plant. 1988, 30, 53–57. [Google Scholar] [CrossRef]
- Almeida, D.M.; Oliveira, M.M.; Saibo, N.J.M. Regulation of Na+ and K+ homeostasis in plants: Towards improved salt stress tolerance in crop plants. Genet. Mol. Biol. 2017, 40 (Suppl. S1), 326–345. [Google Scholar] [CrossRef] [PubMed]
- Türkan, I.; Demiral, T. Recent developments in understanding salinity tolerance. Environ. Exp. Bot. 2009, 67, 2–9. [Google Scholar] [CrossRef]
- Phang, T.H.; Shao, G.; Lam, H.M. Salt tolerance in soybean. J. Integr. Plant Biol. 2008, 50, 1196–1212. [Google Scholar] [CrossRef]
- Chang, R.Z.; Chen, Y.W.; Shao, G.H.; Wan, C.W. Effect of salt stress on agronomic characters and chemical quality of seeds in soybean. Soybean Sci. 1994, 13, 101–105. [Google Scholar]
- Ashraf, M.; Wu, L. Breeding for salinity tolerance in plants. Crit. Rev. Plant Sci. 1994, 13, 17–42. [Google Scholar] [CrossRef]
- Singh, R.; Hemantaranjan, A.; Patel, P.; Yadav, D. Morpho-physiological attributes of field pea (Pisum sativum L.) genotypes as influenced by Salicylic acid under salinity stress. Phytomorphology 2013, 63, 105–114. [Google Scholar]
- Lessani, H.; Marschner, H. Relation between salt tolerance and long-distance transport of sodium and Chloride in various crop species. Funct. Plant Biol. 1978, 5, 27–37. [Google Scholar] [CrossRef]
- Manzoor, A.; Sandhu, G. Response of legumes to salt stress: Effect on growth and nitrogen status of soybean. Pak. J. Agric. Res. 1988, 9, 463–468. [Google Scholar]
- Luo, Q.; Yu, B.; Liu, Y. Differential sensitivity to chloride and sodium ions in seedlings of Glycine max and G. soja under NaCl stress. Plant Physiol. 2005, 162, 1003–1012. [Google Scholar] [CrossRef] [PubMed]
- Parker, M.B.; Gaines, T.P.; Gascho, G.J. Chloride effects on corn. Commun. Soil Sci. Plant Anal. 1985, 16, 1319–1333. [Google Scholar] [CrossRef]
- Ghosh, S.; Mitra, S.; Paul, A. Physiochemical studies of Sodium Chloride on mungbean (Vigna radiata L. Wilczek) and its possible recovery with Spermine and Gibberellic Acid. Sci. World J. 2015, 2015, 85. [Google Scholar] [CrossRef]
- Salim, M.; Pitman, M.G. Effects of salinity on ion uptake and growth of mungbean plants (Vigna radiata L.). Aust. J. Plant Physiol. 1983, 10, 395–407. [Google Scholar] [CrossRef]
- Praxedes, S.C.; De Lacerda, C.F.; DaMatta, F.M.; Prisco, J.T.; Gomes-Filho, E. Salt tolerance is associated with differences in ion accumulation, biomass allocation and photosynthesis in cowpea cultivars. J. Agron. Crop Sci. 2009, 196, 193–204. [Google Scholar] [CrossRef]
- Murillo-Amadot, B.; Troyo-Diéguez, E.; López-Aguilar, R.; López-Cortés, A.; Tinoco-Ojanguri, C.L.; Jones, H.G.; Kaya, C. Matching physiological traits and ion concentrations associated with salt stress in cowpea genotypes. Aust. J. Agric. Res. 2002, 53, 1243–1255. [Google Scholar] [CrossRef]
- Ashraf, M.; Bashir, A. Salt stress induced changes in some organic metabolites and ionic relations in nodules and other plant parts of two crop legumes differing in salt tolerance. Flora 2003, 198, 486–498. [Google Scholar] [CrossRef]
- Demiral, T.; Türkan, I. Does exogenous glycinebetaine affect antioxidative system of rice seedlings under NaCl treatment? Plant Physiol. 2004, 161, 1089–1100. [Google Scholar] [CrossRef] [PubMed]
- Khan, H.A.; Siddique, K.H.M.; Colmer, T.D. Vegetative and reproductive growth of salt-stressed chickpea are carbon-limited: Sucrose infusion at the reproductive stage improves salt tolerance. J. Exp. Botany 2016, 68, 2001–2011. [Google Scholar] [CrossRef]
- Grattan, S.R.; Maas, E.V. Effect of salinity on phosphate accumulation and injury in soybean. Plant Soil 1988, 105, 25–32. [Google Scholar] [CrossRef]
- Richard, C.D. Soil Testing and Plant Analysis Part II Plant Analysis; Soil Science Society of America: Madison, WI, USA, 1967. [Google Scholar]
- Flowers, T.J.; Colmer, T.D. Salinity Tolerance in Halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef] [PubMed]
- Reuter, D.; Robinson, J.B. Plant Analysis: An Interpretation Manual; CSIRO Publishing Collingwood: Victoria, Australia, 1997. [Google Scholar]
- Cheeseman, J.M. Mechanisms of salinity tolerance in plants. Plant Physiol. 1988, 87, 547–550. [Google Scholar] [CrossRef] [PubMed]
- Chi, L.C.; Huei, K.C. Relative importance of Na+, Cl−, and abscisic acid in NaCl induced inhibition of root growth of rice seedlings. Plant Soil 2001, 237, 165–171. [Google Scholar] [CrossRef]
- Tavakkoli, E.; Rengasamy, P.; McDonald, G.K. High concentrations of Na+ and Cl− ions in soil solution have simultaneous detrimental effects on growth of faba bean under salinity stress. J. Exp. Bot. 2010, 61, 4449–4459. [Google Scholar] [CrossRef] [PubMed]
- Tavakkoli, E.; Fatehi, F.; Coventry, S.; Rengasamy, P.; McDonald, G.K. Additive effects of Na+ and Cl− ions on barley growth under salinity stress. J. Exp. Bot. 2011, 62, 2189–2203. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; Greenway, H.; Delane, R.; Gibbs, J. Ion concentration and carbohydrate status of the elongating leaf tissue Hordeum vulgare growing at high external NaCl: II. Cause of the growth reduction. J. Exp. Bot. 1982, 33, 574–583. [Google Scholar] [CrossRef]
- Parker, M.B.; Gascho, G.J.; Gaines, T.P. Chloride toxicity of soybeans grown on Atlantic Coast Flatwoods soils. Agron. J. 1983, 75, 439. [Google Scholar] [CrossRef]
- Abel, G.H. Inheritance of the capacity for chloride inclusion and chloride exclusion by soybeans. Crop Sci. 1969, 9, 697–698. [Google Scholar] [CrossRef]
- Pantalone, V.R.; Kenworthy, W.J.; Slaughter, L.H.; James, B.R. Chloride tolerance in soybean and perennial Glycine accessions. Euphytica 1997, 97, 235–239. [Google Scholar] [CrossRef]
- Läuchli, A.; Wieneke, J. Studies on growth and distribution of Na+, K+ and Cl− in soybean varieties differing in salt tolerance. Z. Pflanz. Bodenkd. 1979, 142, 3–13. [Google Scholar] [CrossRef]
- Lauter, D.; Munns, D. Salt sensitivity of chickpea during vegetative growth and at different humidities. Funct. Plant Biol. 1987, 14, 171–180. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- An, P.; Inanaga, S.; Cohen, Y.; Kafkafi, U.; Sugimoto, Y. Salt tolerance in two soybean cultivars. J. Plant Nutr. 2002, 25, 407–423. [Google Scholar] [CrossRef]
- Serraj, R.; Vasquez-Diaz, H.; Drevon, J.J. Effects of salt stress on nitrogen fixation, oxygen diffusion, and ion distribution in soybean, common bean, and alfalfa. J. Plant Nutr. 1998, 21, 475–488. [Google Scholar] [CrossRef]
- Durand, M.; Lacan, D. Sodium partitioning within the shoot of soybean. Physiol. Plant. 1994, 91, 65–71. [Google Scholar] [CrossRef]
- Abel, G.H.; Mackenzie, A.J. Salt tolerance of soybean varieties (Glycine max L. Merrill) during germination and later growth. Crop Sci. 1964, 4, 157–161. [Google Scholar] [CrossRef]
- Weir, R.G. Pastures and field crops. In Plant Nutrient Disorders; Weir, R.G., Cresswell, G.C., Eds.; Inkata Press: Melbourne, Australia, 1994. [Google Scholar]
- Greenway, H. Salt responses of enzymes from species differing in salt tolerance. J. Plant Physiol. 1972, 49, 256–259. [Google Scholar] [CrossRef]
- Munns, R. Na+, K+ and Cl− in xylem sap flowing to shoots of NaCl-treated barley. J. Exp. Bot. 1985, 36, 1032–1042. [Google Scholar] [CrossRef]
- Chen, P.; Yan, K.; Shao, H.; Zhao, S. Physiological mechanisms for high salt tolerance in wild soybean (Glycine soja) from Yellow River Delta, China: Photosynthesis, osmotic regulation, ion flux and antioxidant capacity. PLoS ONE 2013, 8, e83227. [Google Scholar] [CrossRef]
- Yan, K.; Chen, P.; Shao, H.; Zhao, S.; Zhang, L.; Xu, G.; Sun, J. Responses of photosynthesis and photosystem II to higher temperature and salt stress in sorghum. J. Agron. Crop Sci. 2012, 198, 218–225. [Google Scholar] [CrossRef]
- Khan, H.A.; Siddique, K.H.M.; Munir, R.; Colmer, T.D. Salt sensitivity in chickpea: Growth, photosynthesis, seed yield components and tissue ion regulation in contrasting genotypes. Plant Physiol. 2015, 182, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kao, W.; Tsai, T.; Shih, C. Photosynthetic gas exchange and chlorophyll a fluorescence of three wild soybean species in response to NaCl treatments. Photosynthetica 2003, 41, 415–419. [Google Scholar] [CrossRef]
- Lu, K.X.; Cao, B.H.; Feng, X.P.; He, Y.; Jiang, D.A. Photosynthetic response of salt-tolerant and sensitive soybean varieties. Photosynthetica 2009, 47, 381–387. [Google Scholar] [CrossRef]
- Yang, Y.; Yan, C.Q.; Cao, B.H.; Xu, H.X.; Chen, J.P.; Jiang, D.A. Some photosynthetic responses to salinity resistance are transferred into the somatic hybrid descendants from the wild soybean Glycine cyrtoloba ACC547. Physiol. Plant. 2007, 129, 658–669. [Google Scholar] [CrossRef]
- He, Y.; Chen, Y.; Yu, C.L.; Lu, K.X.; Jiang, Q.S.; Fu, J.L.; Wang, G.M.; Jiang, D.A. Photosynthesis and yield traits in different soybean lines in response to salt stress. Photosynthetica 2016, 54, 630–635. [Google Scholar] [CrossRef]
- Salim, M. Effects of salinity and relative humidity on growth and ionic relations of plants. New Phytol. 1989, 113, 13–20. [Google Scholar] [CrossRef]
- Ashraf, M.; Rasul, E. Salt tolerance of mungbean (Vigna radiata (L.) Wilczek) at two growth stages. Plant Soil 1988, 110, 63–67. [Google Scholar] [CrossRef]
- Plaut, Z.; Grieve, C.; Maas, E. Salinity effects on CO2 assimilation and diffusive conductance of cowpea leaves. Physiol. Plant. 1990, 79, 31–38. [Google Scholar] [CrossRef]
- Zhang, J.; Nguyen, H.T.; Blum, A. Genetic analysis of osmotic adjustment in crop plants. J. Exp. Bot. 1999, 50, 291–302. [Google Scholar] [CrossRef]
- Zhu, J.K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol. 2002, 53, 247–273. [Google Scholar] [CrossRef] [PubMed]
- Cachorro, P.; Martínez, R.; Ortiz, A.; Cerdá, A. Abscisic acid and osmotic relations in Phaseolus vulgaris L. shoots under salt stress. J. Plant Growth Regul. 1995, 14, 99–104. [Google Scholar] [CrossRef]
- Schwarz, M.; Gale, J. Maintenance respiration and carbon balance of plants at low levels of sodium chloride salinity. J. Exp. Bot. 1981, 32, 933–941. [Google Scholar] [CrossRef]
- Yeo, A.R. Salinity resistance: Physiologies and prices. Physiol. Plant. 1983, 58, 214–222. [Google Scholar] [CrossRef]
- Lutts, S.; Majerus, V.; Kinet, J.M. NaCl effects on proline metabolism in rice (Oryza sativa) seedlings. Physiol. Plant. 1999, 105, 450–458. [Google Scholar] [CrossRef]
- Streeter, J.G.; Lohnes, D.G.; Fioritto, R.J. Patterns of pinitol accumulation in soybean plants and relationships to drought tolerance. Plant Cell Environ. 2001, 24, 429–438. [Google Scholar] [CrossRef]
- Blum, A. Osmotic adjustment is a prime drought stress adaptive engine in support of plant production. Plant Cell Environ. 2017, 40, 4–10. [Google Scholar] [CrossRef]
- Rathert, G. The influence of high salt stress on starch, sucrose and degradative enzymes of two glycine max varieties that differ in salt tolerance. J. Plant Nutr. 1985, 8, 199–209. [Google Scholar] [CrossRef]
- Maas, E.V.; Grattan, S.R. Crop Yields as Affected by Salinity; Skaggs, R.W., van Schilfgaarde, J., Eds.; Agricultural Drainage Agronomy Monograph No. 38; ASA: Madison, WI, USA, 1999; pp. 55–108. [Google Scholar]
- Essa, T.A. Effect of salinity stress on growth and nutrient composition of three soybean (Glycine max L. Merrill) Cultivars. J. Agron. Crop Sci. 2002, 88, 86–93. [Google Scholar] [CrossRef]
- Seemann, J.R.; Critchley, C. Effects of salt stress on the growth, ion content, stomatal behaviour and photosynthetic capacity of a salt-sensitive species, Phaseolus vulgaris L. Planta 1985, 164, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Ayers, A.D.; Brown, J.W.; Wadleigh, C.H. Salt tolerance of barley and wheat in soil plots receiving several salinization regimes. Agron. J. 1952, 44, 307–310. [Google Scholar] [CrossRef]
- Kotula, L.; Clode, P.L.; Jimenez, J.D.L.C.; Colmer, T.D. Salinity tolerance in chickpea is associated with the ability to ‘exclude’ Na from leaf mesophyll cells. J. Exp. Botany 2019, 70, 4991–5002. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; Wallace, P.A.; Teakle, N.L.; Colmer, T.D. Measuring soluble ion concentrations (Na+, K+, Cl−) in salt-treated plants. Methods Mol. Biol. 2010, 639, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Naidu, B. Separation of sugars, polyols, proline analogues, and betaines in stressed plant extracts by high performance liquid chromatography and quantification by ultraviolet detection. Aust. J. Plant Physiol. 1998, 25, 793–800. [Google Scholar] [CrossRef]
- Colmer, T.D.; Corradini, F.; Cawthray, G.R.; Otte, M.L. Analysis of dimethylsulphoniopropionate (DMSP), betaines and other organic solutes in plant tissue extracts using HPLC. Phytochem. Anal. 2000, 11, 163–168. [Google Scholar] [CrossRef]
- Fan, T.W.; Colmer, T.D.; Lane, A.N.; Higashi, R.M. Determination of metabolites by 1H NMR and GC: Analysis for organic osmolytes in crude tissue extracts. Anal. Biochem. 1993, 214, 260–271. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Ψπ (MPa) | Ion Concentrations (mM) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Na+ | Cl− | K+ | Ca2+ | Mg2+ | SO42− | NO3− | SiO3− | NH4+ | H2PO4− | ||
| Control | −0.05 | 0.2 | 0.05 | 5.0 | 5.0 | 0.4 | 5.4 | 4.4 | 0.1 | 0.6 | 0.2 |
| NaCl (100 mM) | −0.49 | 100.2 | 100.05 | 5.0 | 5.0 | 0.4 | 5.4 | 4.4 | 0.1 | 0.6 | 0.2 |
| Na+ (100 mM, without Cl−): 33.33 mM Na2SO4 + 33.33 mM NaNO3 | −0.41 | 100.2 | 0.05 | 5.0 | 5.0 | 0.4 | 38.7 | 37.7 | 0.1 | 0.6 | 0.2 |
| Cl− (100 mM, without Na+): 16.67 mM CaCl2 + 16.67 mM MgCl2 + 33.33 mM KCl | −0.41 | 0.2 | 100.05 | 38.3 | 16.7 | 17.1 | 5.4 | 4.4 | 0.1 | 0.6 | 0.2 |
| High-cation negative control (equivalent to Cl− treatment): 10.0 mM CaSO4 + 6.67 mM Ca(NO3)2 + 16.67 mM MgSO4 + 16.67 mM K2SO4 | −0.43 | 0.2 | 0.05 | 71.6 | 28.3 | 17.1 | 65.3 | 11.1 | 0.1 | 0.6 | 0.2 |
| Species | Treatment | Pod Dry Mass (g) | % of Control | Seed Dry Mass (g) | % of Control | Seed Number | % of Control |
|---|---|---|---|---|---|---|---|
| Soybean | Control | 21.1 ± 0.8 | - | 10.0 ± 0.5 | 101 ± 5 | - | |
| NaCl | 10.9 ± 1.6 | 52 | 6.2 ± 0.9 | 62 | 41 ± 8 | 41 | |
| Na+ salts | 5.2 ± 2.9 | 25 | 3.0 ± 1.6 | 30 | 49 ± 5 | 48 | |
| Cl− salts | 15.6 ± 3.1 | 74 | 6.3 ± 1.5 | 63 | 65 ± 6 | 64 | |
| High cation | 15.5 ± 1.4 | 73 | 10.4 ± 1.6 | 104 | 84 ± 9 | 83 | |
| LSD (5%) | 6.4 *** | - | 3.9 ** | - | 53 * | - | |
| Mungbean | Control | 15.5 ± 1.0 | 10.9 ± 0.9 | 151 ± 10 | - | ||
| NaCl | 0.4 ± 0.2 | 3 | 0.2 ± 0.1 | 2 | 3 ± 1 | 2 | |
| Na+ salts | 1.8 ± 0.8 | 12 | 1.3 ± 0.6 | 12 | 18 ± 6 | 12 | |
| Cl− salts | 1.1 ± 0.2 | 7 | 0.3 ± 0.04 | 3 | 6.0 ± 2 | 4 | |
| High cation | 6.1 ± 1.3 | 39 | 4.3 ± 1.1 | 39 | 67 ± 10 | 44 | |
| LSD (5%) | 3.4 *** | - | 2.9 *** | - | 47 *** | - | |
| Cowpea | Control | 26.2 ± 2.5 | - | 22.0 ± 1.6 | - | 372 ± 12 | - |
| NaCl | 6.3 ± 1.0 | 24 | 4.7 ± 1.3 | 21 | 71 ± 15 | 19 | |
| Na+ salts | 11.3 ± 2.0 | 43 | 9.7 ± 1.6 | 44 | 166 ± 11 | 45 | |
| Cl− salts | 12.7 ± 2.0 | 48 | 10.6 ± 1.6 | 48 | 167 ± 12 | 45 | |
| High cation | 18.6 ± 0.9 | 71 | 15.6 ± 0.7 | 71 | 240 ± 14 | 64 | |
| LSD (5%) | 5.3 *** | - | 4.2 *** | - | 89 *** | - | |
| Common bean | Control | 17.9 ± 3.6 | 13.5 ± 3.3 | 94 ± 12 | |||
| NaCl | 1.4 ± 0.7 | 8 | 0.6 ± 0.1 | 4 | 6 ± 2 | 6 | |
| Na+ salts | 9.4 ± 0.9 | 52 | 6.5 ± 1.0 | 48 | 46 ± 11 | 49 | |
| Cl− salts | 4.8 ± 2.8 | 27 | 2.8 ± 1.9 | 21 | 56 ± 11 | 59 | |
| High cation | 8.2 ± 2.5 | 46 | 5.5 ± 1.8 | 41 | 54 ± 11 | 57 | |
| LSD (5%) | 7.6 ** | - | 5.9 ** | - | 59 * | - | |
| LSD (5%) | S | 2.5 *** | - | 1.8 *** | - | 31 *** | - |
| T | 2.8 *** | - | 2.1 *** | - | 35 *** | - | |
| S × T | 5.3 *** | - | 4.1 * | - | 71 *** | - |
| Species | Treatment | Pod-Filling Stage | |||||
|---|---|---|---|---|---|---|---|
| Ψπsap (MPa) | Change in Ψπsap (MPa) | Ψπsap (MPa) | Change in Ψπsap (MPa) | Ψπsap (MPa) | Change in Ψπsap (MPa) | ||
| Soybean | Control | −1.01 ± 0.10 | - | −0.94 ± 0.05 | - | −1.11 ± 0.11 | - |
| NaCl | −1.63 ± 0.17 | 0.62 | −1.19 ± 0.04 | 0.25 | −1.35 ± 0.07 | 0.25 | |
| Na+ salts | −1.10 ± 0.05 | 0.09 | −1.14 ± 0.10 | 0.20 | −1.76 ± 0.11 | 0.65 | |
| Cl− salts | −1.18 ± 0.08 | 0.17 | −1.13 ± 0.01 | 0.19 | −1.39 ± 0.04 | 0.28 | |
| High cation | −1.10 ± 0.10 | 0.09 | −1.15 ± 0.11 | 0.21 | −1.52 ± 0.10 | 0.41 | |
| LSD (5%) | 0.32 *** | - | n.s. | - | 0.27 ** | - | |
| Mungbean | Control | −0.76 ± 0.05 | - | −0.77 ± 0.05 | - | −0.94 ± 0.05 | - |
| NaCl | −1.88 ± 0.25 | 1.12 | −1.24 ± 0.01 | 0.46 | - | - | |
| Na+ salts | −0.91 ± 0.08 | 0.15 | −1.01 ± 0.07 | 0.23 | −1.77 ± 0.04 | 0.82 | |
| Cl− salts | −2.05 ± 0.11 | 1.29 | −1.62 ± 0.01 | 0.85 | - | - | |
| High cation | −1.11 ± 0.02 | 0.35 | −1.17 ± 0.09 | 0.39 | −1.33 ± 0.07 | 0.39 | |
| LSD (5%) | 0.39 *** | - | 0.15 *** | - | 0.5 *** | - | |
| Cowpea | Control | −0.83 ± 0.03 | - | −0.85 ± 0.08 | - | −1.00 ± 0.11 | - |
| NaCl | −1.12 ± 0.03 | 0.29 | −1.07 ± 0.05 | 0.22 | −1.08 ± 0.02 | 0.08 | |
| Na+ salts | −1.10 ± 0.06 | 0.27 | −1.13 ± 0.08 | 0.28 | −1.35 ± 0.23 | 0.34 | |
| Cl− salts | −1.12 ± 0.06 | 0.29 | −1.03 ± 0.06 | 0.17 | −1.25 ± 0.07 | 0.25 | |
| High cation | −1.13 ± 0.07 | 0.30 | −1.00 ± 0.04 | 0.14 | −1.05 ± 0.05 | 0.04 | |
| LSD (5%) | 0.16 ** | - | n.s. | - | n.s. | - | |
| Common bean | Control | −0.98 ± 0.02 | - | −0.75 ± 0.03 | - | −0.95 ± 0.16 | - |
| NaCl | −1.25 ± 0.06 | 0.27 | −1.36 ± 0.11 | 0.61 | −1.75 ± 0.04 | 0.80 | |
| Na+ salts | −1.28 ± 0.10 | 0.30 | −1.11 ± 0.06 | 0.36 | −1.38 ± 0.09 | 0.43 | |
| Cl− salts | −1.20 ± 0.03 | 0.22 | −1.11 ± 0.09 | 0.36 | −1.74 ± 0.10 | 0.79 | |
| High cation | −1.10 ± 0.03 | 0.12 | −1.04 ± 0.04 | 0.39 | −1.12 ± 0.05 | 0.17 | |
| LSD (5%) | 0.16 * | - | 0.22 *** | 0.37 * | |||
| LSD (5%) | S | 0.11 *** | 0.08 *** | 0.36 ** | |||
| T | 0.13 *** | 0.09 *** | 0.40 *** | ||||
| S × T | 0.24 *** | 0.18 *** | 0.80 ** | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Le, L.T.T.; Kotula, L.; Siddique, K.H.M.; Colmer, T.D. Na+ and/or Cl− Toxicities Determine Salt Sensitivity in Soybean (Glycine max (L.) Merr.), Mungbean (Vigna radiata (L.) R. Wilczek), Cowpea (Vigna unguiculata (L.) Walp.), and Common Bean (Phaseolus vulgaris L.). Int. J. Mol. Sci. 2021, 22, 1909. https://doi.org/10.3390/ijms22041909
Le LTT, Kotula L, Siddique KHM, Colmer TD. Na+ and/or Cl− Toxicities Determine Salt Sensitivity in Soybean (Glycine max (L.) Merr.), Mungbean (Vigna radiata (L.) R. Wilczek), Cowpea (Vigna unguiculata (L.) Walp.), and Common Bean (Phaseolus vulgaris L.). International Journal of Molecular Sciences. 2021; 22(4):1909. https://doi.org/10.3390/ijms22041909
Chicago/Turabian StyleLe, Ly Thi Thanh, Lukasz Kotula, Kadambot H. M. Siddique, and Timothy D. Colmer. 2021. "Na+ and/or Cl− Toxicities Determine Salt Sensitivity in Soybean (Glycine max (L.) Merr.), Mungbean (Vigna radiata (L.) R. Wilczek), Cowpea (Vigna unguiculata (L.) Walp.), and Common Bean (Phaseolus vulgaris L.)" International Journal of Molecular Sciences 22, no. 4: 1909. https://doi.org/10.3390/ijms22041909
APA StyleLe, L. T. T., Kotula, L., Siddique, K. H. M., & Colmer, T. D. (2021). Na+ and/or Cl− Toxicities Determine Salt Sensitivity in Soybean (Glycine max (L.) Merr.), Mungbean (Vigna radiata (L.) R. Wilczek), Cowpea (Vigna unguiculata (L.) Walp.), and Common Bean (Phaseolus vulgaris L.). International Journal of Molecular Sciences, 22(4), 1909. https://doi.org/10.3390/ijms22041909