
Antimicrobial Properties of Palladium and Platinum Nanoparticles: A New Tool for Combating Food-Borne Pathogens

 , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. The Effect of Palladium Nanoparticles
2.2. The Effect of Platinum Nanoparticles
2.3. Colony Plate Counting and Inhibitory Rate Method
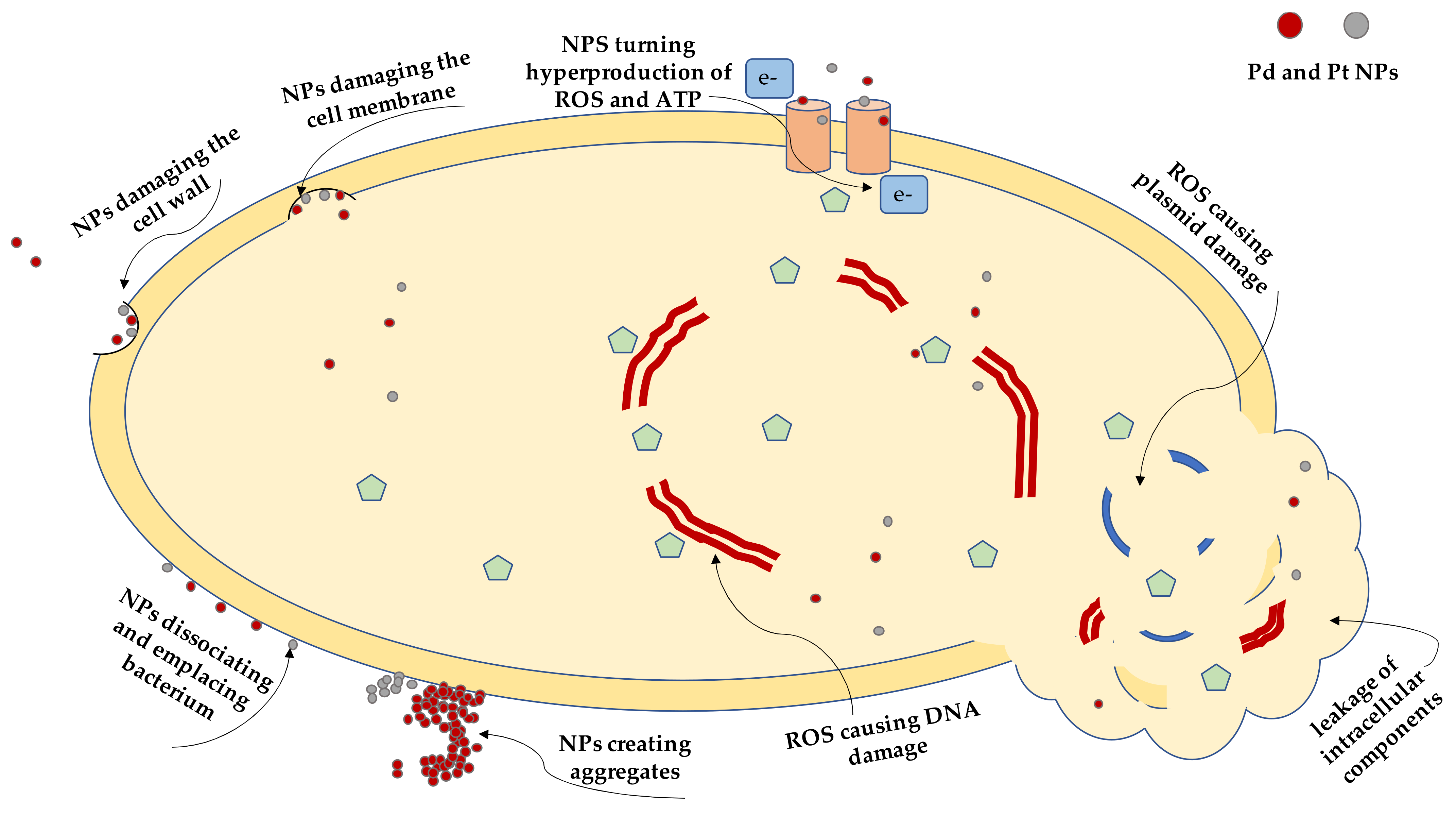
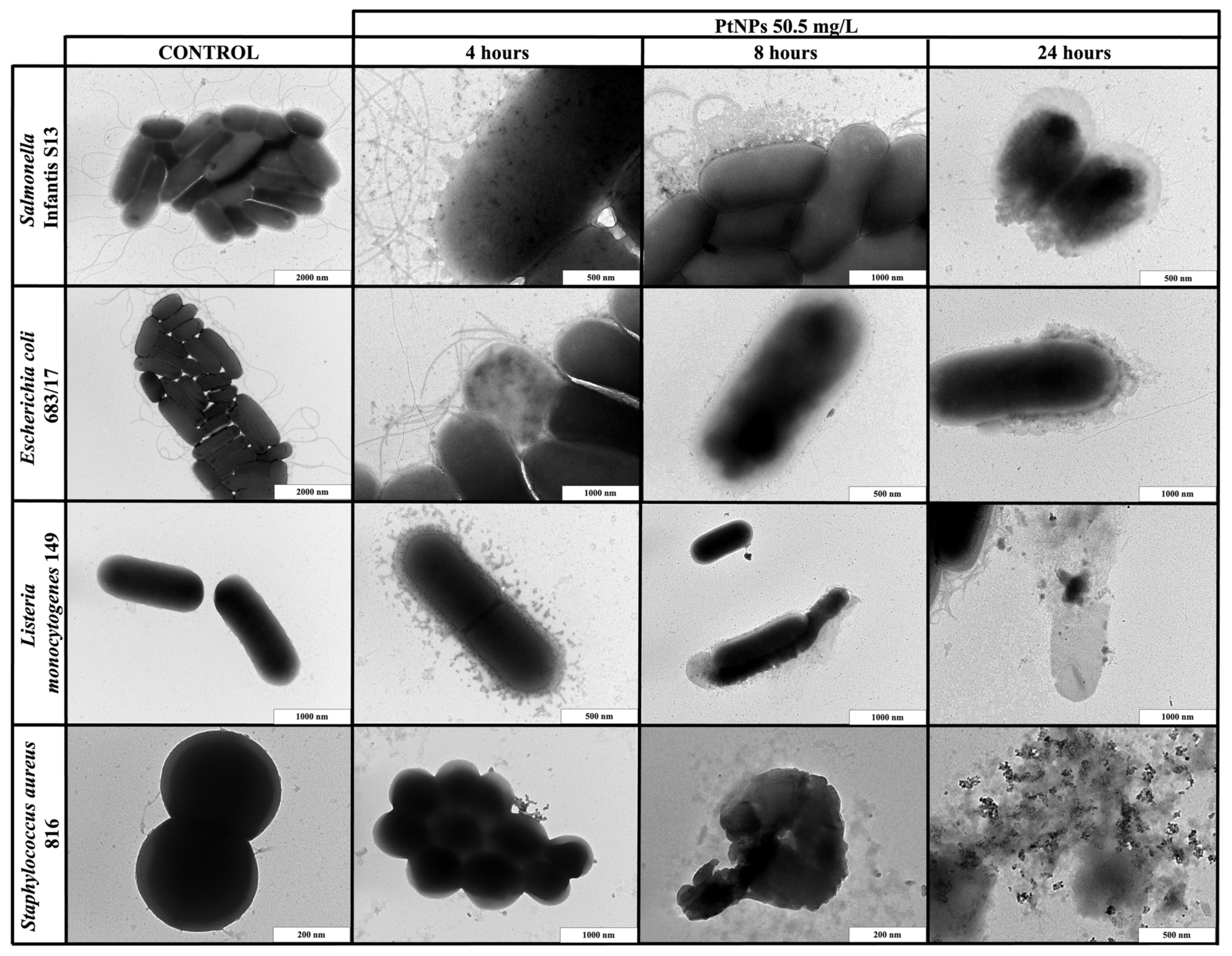
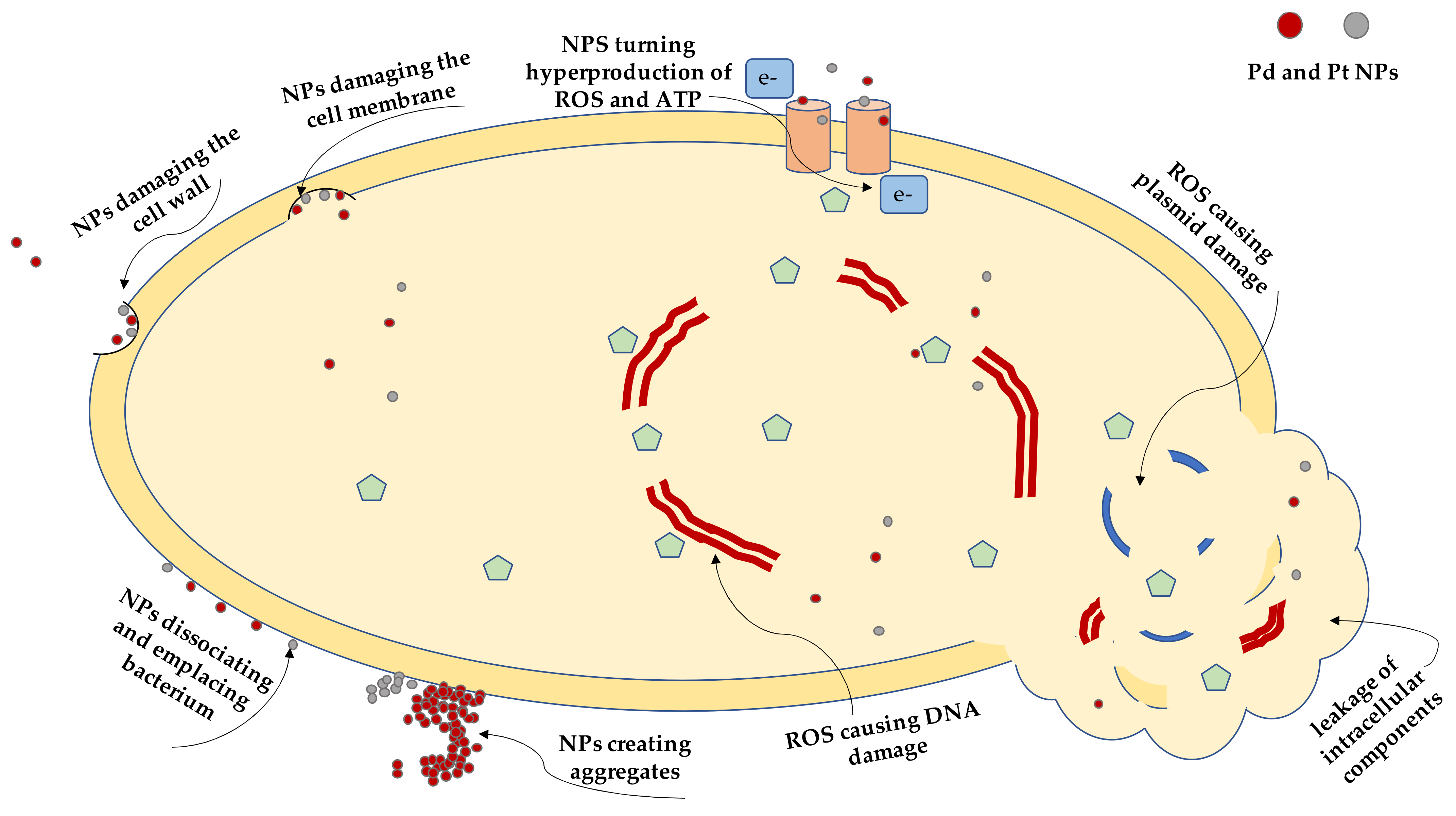
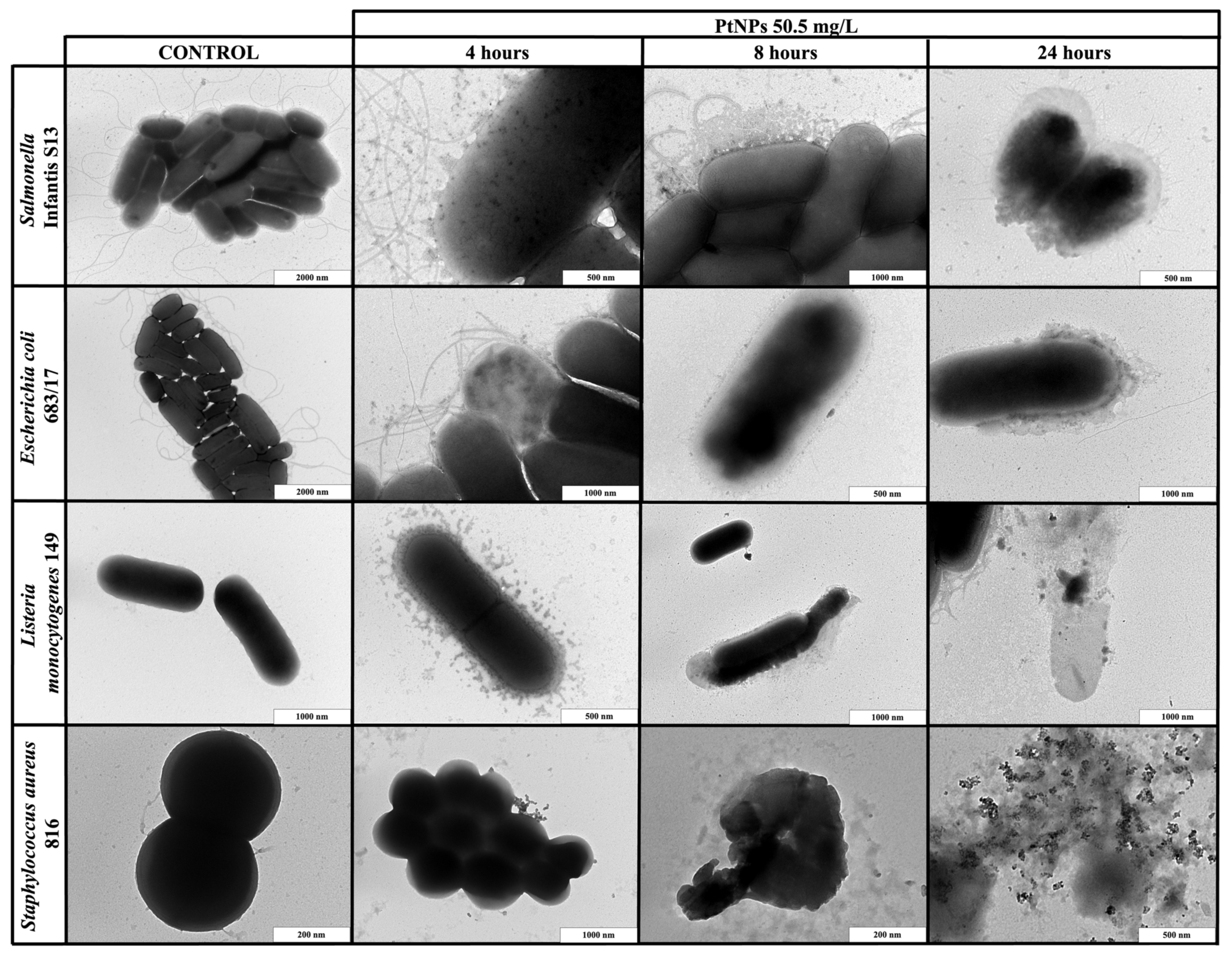
2.4. Transmission Electron Microscopy Imaging
2.5. Acute Cytotoxicity of Metallic Nanoparticles
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Preparation of Metallic Nanoparticles
4.3. Bacterial Strains
4.4. Cell Lines and Cell Cultures
4.5. Bacterial Stock Cultures Preparation
4.6. Inoculum Preparation and Preparation of Dilution Series for Metallic Nanoparticles
4.7. Determination of Minimum Inhibitory Concentrations
4.7.1. Evaluation of Planktonic Cells Growth
4.7.2. Quantification of Biofilm Formation
4.7.3. Evaluation of Metabolic Activity
4.7.4. Evaluation of Nanoparticles Effect on Preformed Biofilms
4.8. Evaluation of Growth Inhibition Using the Plate Counting Agar
4.9. Transmission Electron Microscopy Imaging
4.10. Cytotoxicity Assay
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bintsis, T. Foodborne pathogens. AIMS Microbiol. 2017, 3, 529–563. [Google Scholar] [CrossRef]
- Schirone, M.; Visciano, P.; Tofalo, R.; Suzzi, G. Editorial: Foodborne Pathogens: Hygiene and Safety. Front. Microbiol. 2019, 10, 1974. [Google Scholar] [CrossRef] [Green Version]
- Nespolo, N.M. The Behavior of Consumers and Producers of Food of Animal Origin and Their Impacts in One Health. Front. Vet. Sci. 2021, 8, 607. [Google Scholar] [CrossRef]
- Berlanga, M.; Guerrero, R. Living together in biofilms: The microbial cell factory and its biotechnological im-plications. Microb Cell Fact. 2016, 15, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, A.L.S.D.; Galdino, A.C.M.; Mello, T.P.; Ramos, L.S.; Branquinha, M.H.; Bolognese, A.M.; Columbano Neto, J.; Roudbary, M. What are the advantages of living in a community? A microbial biofilm perspective! Mem. Inst. Oswaldo Cruz 2018, 113, e180212. [Google Scholar] [PubMed] [Green Version]
- Gebreyohannes, G.; Nyerere, A.; Bii, C.; Sbhatu, D.B. Challenges of intervention, treatment, and antibiotic resistance of biofilm-forming microorganisms. Heliyon 2019, 5, e02192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rumbaugh, K.P.; Sauer, K. Biofilm dispersion. Nat. Rev. Microbiol. 2020, 18, 571–586. [Google Scholar] [CrossRef] [PubMed]
- Awad, T.S.; Asker, D.; Hatton, B.D. Food-safe modification of stainless steel food-processing surfaces to re-duce bacterial biofilms. ACS Appl. Mater. Interfaces 2018, 10, 22902–22912. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhao, F.; Wang, J.; Zhong, N. Biofilm formation and control strategies of foodborne pathogens: Food safety perspectives. RSC Adv. 2017, 7, 36670–36683. [Google Scholar] [CrossRef] [Green Version]
- Chew, S.; Yang, L. Biofilms. In Encyclopedia of Food and Health; Academic Press: Cambridge, MA, USA, 2016; pp. 407–415. [Google Scholar] [CrossRef]
- Flemming, H.-C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef]
- Han, C.; Romero, N.; Fischer, S.; Dookran, J.; Berger, A.; Doiron, A.L. Recent developments in the use of nanoparticles for treatment of biofilms. Nanotechnol. Rev. 2017, 6, 383–404. [Google Scholar] [CrossRef]
- González, A.; Riego, A.; Vega, V.; García, J.; Galié, S.; del Río, I.G.; de Yuso, M.M.; Villar, C.; Lombó, F.; De la Prida, V. Functional Antimicrobial Surface Coatings Deposited onto Nanostructured 316L Food-Grade Stainless Steel. Nanomaterials 2021, 11, 1055. [Google Scholar] [CrossRef]
- Marriott, N.G.; Schilling, M.W.; Gravani, R.B. Principles of Food Sanitation; Springer International Publishing: Cham, Switzerland, 2018. [Google Scholar]
- Lahiri, D.; Nag, M.; Sheikh, H.I.; Sarkar, T.; Edinur, H.A.; Pati, S.; Ray, R.R. Microbiologically-Synthesized Nanoparticles and Their Role in Silencing the Biofilm Signaling Cascade. Front. Microbiol. 2021, 12, 180. [Google Scholar] [CrossRef] [PubMed]
- Qayyum, S.; Khan, A.U. Nanoparticles vs. biofilms: A battle against another paradigm of antibiotic resistance. MedChemComm 2016, 7, 1479–1498. [Google Scholar] [CrossRef]
- Wang, L.; Hu, C.; Shao, L. The antimicrobial activity of nanoparticles: Present situation and prospects for the future. Int. J. Nanomed. 2017, 12, 1227–1249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahira, S.; Jain, A.; Khan, W.; Domb, A.J. Chapter 1: Antimicrobial Materials—An Overview. In Antimicrobial Materials for Biomedical Applications; Royal Society of Chemistry: London, UK, 2019; pp. 1–37. [Google Scholar]
- Lee, H.; Lee, D.G. A Novel Strategy for Antimicrobial Agents: Silver Nanoparticles, In Metal Nanoparticles in Pharma; Springer: Cham, Switzerland, 2017; pp. 139–153. [Google Scholar]
- Masri, A.; Anwar, A.; Khan, N.A.; Siddiqui, R. The Use of Nanomedicine for Targeted Therapy against Bacterial Infections. Antibiotics 2019, 8, 260. [Google Scholar] [CrossRef] [Green Version]
- McClements, D.J.; Xiao, H. Is nano safe in foods? Establishing the factors impacting the gastrointestinal fate and toxicity of organic and inorganic food-grade nanoparticles. npj Sci. Food 2017, 1, 1–13. [Google Scholar] [CrossRef]
- Ameta, S.K.; Rai, A.K.; Hiran, D.; Ameta, R.; Ameta, S.C. Use of nanomaterials in food science. In Biogenic Nano-Particles and Their Use in Agro-Ecosystems; Ghor-banpour, M., Bhargava, P., Varma, A., Choudhary, D., Eds.; Springer: Singapore, 2020. [Google Scholar]
- Pandhi, S.; Mahato, D.K.; Kumar, A. Overview of Green Nanofabrication Technologies for Food Quality and Safety Applications. Food Rev. Int. 2021, 1–21. [Google Scholar] [CrossRef]
- Otari, S.V.; Patel, S.K.S.; Lee, J.H.; Jeong, J.-H. A green chemistry approach for synthesizing thermostable antimicrobial peptide-coated gold nanoparticles immobilized in an alginate biohydrogel. RSC Adv. 2016, 6, 86808–86816. [Google Scholar] [CrossRef]
- Samuggam, S.; Chinni, S.; Mutusamy, P.; Gopinath, S.; Anbu, P.; Venugopal, V.; Reddy, L.; Enugutti, B. Green Synthesis and Characterization of Silver Nanoparticles Using Spondias mombin Extract and Their Antimicrobial Activity against Biofilm-Producing Bacteria. Molecules 2021, 26, 2681. [Google Scholar] [CrossRef]
- Mittal, D.; Kaur, G.; Singh, P.; Yadav, K.; Ali, S.A. Nanoparticle-based sustainable agriculture and food Science: Recent advances and future uutlook. Front. Nanotechnol. 2020, 2, 10. [Google Scholar] [CrossRef]
- Lencova, S.; Zdenkova, K.; Jencova, V.; Demnerova, K.; Zemanova, K.; Kolackova, R.; Hozdova, K.; Stiborova, H. Benefits of Polyamide Nanofibrous Materials: Antibacterial Activity and Retention Ability for Staphylococcus Aureus. Nanomaterials 2021, 11, 480. [Google Scholar] [CrossRef]
- Halicka, K.; Cabaj, J. Electrospun Nanofibers for Sensing and Biosensing Applications—A Review. Int. J. Mol. Sci. 2021, 22, 6357. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Park, G.D.; Patel, S.K.; Kondaveeti, S.; Otari, S.; Anwar, M.Z.; Kalia, V.C.; Singh, Y.; Kim, S.C.; Cho, B.-K.; et al. SiO2 microparticles with carbon nanotube-derived mesopores as an efficient support for enzyme immobilization. Chem. Eng. J. 2019, 359, 1252–1264. [Google Scholar] [CrossRef]
- Chiari-Andréo, B.G.; de Almeida-Cincotto, M.G.J.; Oshiro, J.A.; Taniguchi, C.Y.Y.; Chiavacci, L.A.; Isaac, V.L.B. Nanoparticles for cosmetic use and its application. In Nanoparticles in Pharmacotherapy; Elsevier BV: Amsterdam, The Netherlands, 2019; pp. 113–146. [Google Scholar]
- Echiegu, E.A. Nanotechnology as a Tool for Enhanced Renewable Energy Application in Developing Countries. J. Fundam. Renew. Energy Appl. 2016, 6. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Pi, C.; Feng, X.; Hou, Y.; Zhao, L.; Wei, Y. The Influence of Nanoparticle Properties on Oral Bioavailability of Drugs. Int. J. Nanomed. 2020, 15, 6295–6310. [Google Scholar] [CrossRef]
- Hassan, A.A.; Mansour, M.K.; El Hamaky, A.M.; El Ahl, R.M.S.; Oraby, N.H. Nanomaterials and nanocomposite applications in veterinary medicine. In Multifunctional Hybrid Nanomaterials for Sustainable Agri-Food and Ecosystems; Elsevier BV: Amsterdam The Netherlands, 2020; pp. 583–638. [Google Scholar]
- Mohajerani, A.; Burnett, L.; Smith, J.V.; Kurmus, H.; Milas, J.; Arulrajah, A.; Horpibulsuk, S.; Kadir, A.A. Nanoparticles in Construction Materials and Other Applications, and Implications of Nanoparticle Use. Materials 2019, 12, 3052. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.K.; Sudarshan, K.; Kadam, R. Optical nanomaterials with focus on rare earth doped oxide: A Review. Mater. Today Commun. 2021, 27, 102277. [Google Scholar] [CrossRef]
- Auría-Soro, C.; Nesma, T.; Juanes-Velasco, P.; Landeira-Viñuela, A.; Fidalgo-Gomez, H.; Acebes-Fernandez, V.; Gongora, R.; Parra, M.J.A.; Manzano-Roman, R.; Fuentes, M. Interactions of Nanoparticles and Biosystems: Microenvironment of Nanoparticles and Biomolecules in Nanomedicine. Nanomaterials 2019, 9, 1365. [Google Scholar] [CrossRef] [Green Version]
- Chlumsky, O.; Purkrtova, S.; Turonova, H.M.; Fuchsova, V.S.; Slepicka, P.; Fajstavr, D.; Ulbrich, P.; Demnerova, K. The effect of gold and silver nanoparticles, chitosan and their combinations on bacterial biofilms of food-borne pathogens. Biofouling 2020, 36, 222–233. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Yanagiuchi, H.; Kitagawa, H.; Honda, Y. Inhibitory effect of platinum nanoparticles on bio-film formation of oral bacteria. Nano. Biomed. 2017, 9, 77–82. [Google Scholar]
- Pedone, D.; Moglianetti, M.; De Luca, E.; Bardi, G.; Pompa, P.P. Platinum nanoparticles in nanobiomedicine. Chem. Soc. Rev. 2017, 46, 4951–4975. [Google Scholar] [CrossRef] [PubMed]
- Hullo, M.; Grall, R.; Perrot, Y.; Mathé, C.; Ménard, V.; Yang, X.; Lacombe, S.; Porcel, E.; Villagrasa, C.; Chevillard, S.; et al. Radiation Enhancer Effect of Platinum Nanoparticles in Breast Cancer Cell Lines: In Vitro and In Silico Analyses. Int. J. Mol. Sci. 2021, 22, 4436. [Google Scholar] [CrossRef] [PubMed]
- Naseer, B.; Srivastava, G.; Qadri, O.; Faridi, S.; Islam, R.; Younis, K. Importance and health hazards of na-noparticles used in the food industry. Nanotechnol. Rev. 2018, 7, 623–641. [Google Scholar] [CrossRef]
- Adams, C.P.; Walker, K.A.; Obare, S.O.; Docherty, K.M. Size-dependent antimicrobial effects of novel palla-dium nanoparticles. PLoS ONE 2014, 9, e85981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manikandan, V.; Velmurugan, P.; Park, J.-H.; Lovanh, N.; Seo, S.-K.; Jayanthi, P.; Park, Y.-J.; Cho, M.; Oh, B.-T. Synthesis and antimicrobial activity of palladium nanoparticles from Prunus × yedoensis leaf extract. Mater. Lett. 2016, 185, 335–338. [Google Scholar] [CrossRef]
- Staszek, M.; Siegel, J.; Kolarova, K.; Rimpelova, S.; Švorčík, V. Formation and antibacterial action of Pt and Pd nanoparticles sputtered into liquid. Micro Nano Lett. 2014, 9, 778–781. [Google Scholar] [CrossRef]
- Slavin, Y.N.; Asnis, J.; Häfeli, U.O.; Bach, H. Metal nanoparticles: Understanding the mechanisms behind an-tibacterial activity. J. Nanobiotechnol. 2017, 15, 65. [Google Scholar] [CrossRef] [PubMed]
- Qin, N.; Tan, X.; Jiao, Y.; Liu, L.; Zhao, W.; Yang, S.; Jia, A. RNA-Seq-based transcriptome analysis of methicillin-resistant Staphylococcus aureus biofilm inhibition by ursolic acid and resveratrol. Sci. Rep. 2015, 4, 5467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Microchem Laboratory. Log and Percent Reductions in Microbiology and Antimicrobial Testing. Available online: http://archive.today/2021.05.04-100145/https://microchemlab.com/information/log-and-percent-reductions-microbiology-and-antimicrobial-testing (accessed on 4 May 2021).
- Tran, V.N.; Viktorova, J.; Augustynkova, K.; Jelenova, N.; Dobiasova, S.; Rehorova, K.; Fenclova, M.; Stranska-Zachariasova, M.; Vitek, L.; Hajslova, J.; et al. In Silico and In Vitro Studies of Mycotoxins and Their Cocktails; Their Toxicity and Its Mitigation by Silibinin Pre-Treatment. Toxins 2020, 12, 148. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BACTERIAL STRAIN | PdNPs (mg/L) | |||||
|---|---|---|---|---|---|---|
| MICPC80 | A620 Inhibition (%) | MICBF80 | A595 Inhibition (%) | MICBM80 | A595 Inhibition (%) | |
| S. aureus 816 | >22.25 | 61.4 ± 3.8 | 22.25 | 90.2 ± 4.1 | >22.25 | 47.0 ± 16.6 |
| S. aureus 1241 | >22.25 | 61.4 ± 4.2 | 22.25 | 80.5 ± 2.0 | >22.25 | 34.8 ± 16.2 |
| L. monocytogenes 149 | >22.25 | 51.0 ± 3.3 | >22.25 | 14.8 ± 6.4 | >22.25 | 18.1 ± 10.1 |
| L. monocytogenes 164 | >22.25 | 28.6 ± 6.9 | >22.25 | 39.4 ± 4.6 | >22.25 | 43.0 ± 11.5 |
| E. coli 683/17 | 22.25 | 83.0 ± 0.6 | >22.25 | 22.3 ± 1.2 | >22.25 | 19.1 ± 0.6 |
| E. coli 693/17 | 22.25 | 92.0 ± 0.1 | >22.25 | 9.6 ± 4.1 | >22.25 | 3.3 ± 0.3 |
| E. coli 815 | >22.25 | 66.0 ± 7.3 | >22.25 | 47.0 ± 3.0 | >22.25 | 35.8 ± 1.8 |
| E. coli 859 | >22.25 | 74.2 ± 0.1 | >22.25 | 50.4 ± 2.4 | >22.25 | 52.1 ± 2.8 |
| S. Infantis S13 | >22.25 | 41.1 ± 2.2 | >22.25 | 17.0 ± 2.0 | >22.25 | 11.0 ± 0.0 |
| S. Infantis S59 | >22.25 | 47.3 ± 0.8 | >22.25 | 12.0 ± 1.8 | >22.25 | 51.5 ± 2.3 |
| BACTERIAL STRAIN | PtNPs (mg/L) | |||||
|---|---|---|---|---|---|---|
| MICPC80 | A620 Inhibition (%) | MICBF80 | A595 Inhibition (%) | MICBM80 | A595 Inhibition (%) | |
| S. aureus 816 | >50.5 | 61.8 ± 5.6 | 50.5 | 88.8 ± 3.9 | >50.5 | 53.7 ± 14.8 |
| S. aureus 1241 | >50.5 | 63.1 ± 5.9 | >50.5 | 54.9 ± 7.5 | >50.5 | 37.1 ± 6.1 |
| L. monocytogenes 149 | >50.5 | 49.2 ± 1.7 | >50.5 | 12.7 ± 0.7 | >50.5 | 21.5 ± 13.6 |
| L. monocytogenes 164 | >50.5 | 28.9 ± 5.7 | >50.5 | 15.3 ± 0.3 | >50.5 | 59.4 ± 10.6 |
| E. coli 683/17 | >50.5 | 76.8 ± 2.9 | >50.5 | 21.2 ± 1.5 | >50.5 | 5.8 ± 0.3 |
| E. coli 693/17 | >50.5 | 77.8 ± 1.6 | >50.5 | 6.0 ± 1.7 | >50.5 | 24.6 ± 8.9 |
| E. coli 815 | >50.5 | 67.6 ± 3.0 | >50.5 | 5.3 ± 0.3 | >50.5 | 32.3 ± 6.2 |
| E. coli 859 | >50.5 | 69.0 ± 7.2 | >50.5 | 43.0 ± 5.4 | >50.5 | 36.0 ± 4.9 |
| S. Infantis S13 | >50.5 | 51.6 ± 1.6 | >50.5 | 19.4 ± 7.4 | >50.5 | 23.8 ± 14.2 |
| S. Infantis S59 | >50.5 | 51.5 ± 1.9 | >50.5 | 8.0 ± 0.2 | >50.5 | 64.3 ± 0.7 |
| BACTERIAL STRAIN | PdNPs (mg/L) | |||||
|---|---|---|---|---|---|---|
| MICBC80 | A620 Inhibition (%) | MICBR80 | A595 Inhibition (%) | MICMPB80 | A595 Inhibition (%) | |
| S. aureus 816 | 44.5 | 98.0 ± 3.6 | 44.5 | 87.1 ± 7.4 | 44.5 | 96.5 ± 1.4 |
| S. aureus 1241 | 44.5 | 94.0 ± 3.2 | >44.5 | 75.7 ± 8.2 | 44.5 | 97.2 ± 0.6 |
| L. monocytogenes 149 | 44.5 | 89.7 ± 5.2 | >44.5 | 38.3 ± 0.3 | 44.5 | 100.0 ± 4.0 |
| L. monocytogenes 164 | 44.5 | 85.3 ± 7.7 | >44.5 | 29.1 ± 25.4 | 44.5 | 100.0 ± 0.6 |
| E. coli 683/17 | 22.25 | 84.5 ± 5.6 | >44.5 | 23.8 ± 11.7 | 44.5 | 100.0 ± 0.1 |
| E. coli 693/17 | 22.25 | 86.6 ± 3.0 | >44.5 | 20.3 ± 9.2 | 44.5 | 100.0 ± 0.2 |
| E. coli 815 | 22.25 | 84.7 ± 3.8 | >44.5 | 20.2 ± 3.8 | 44.5 | 93.3 ± 6.7 |
| E. coli 859 | 22.25 | 90.5 ± 3.7 | >44.5 | 52.5 ± 8.8 | 44.5 | 99.5 ± 1.0 |
| S. Infantis S13 | 44.5 | 96.6 ± 0.1 | 44.5 | 83.6 ± 1.8 | 44.5 | 97.2 ± 0.0 |
| S. Infantis S59 | 44.5 | 95.1 ± 0.2 | 44.5 | 82.7 ± 6.7 | 44.5 | 90.5 ± 2.1 |
| BACTERIAL STRAIN | PtNPs (mg/L) | |||||
|---|---|---|---|---|---|---|
| MICBC80 | A620 Inhibition (%) | MICBR80 | A595 Inhibition (%) | MICMPB80 | A595 Inhibition (%) | |
| S. aureus 816 | 101 | 87.3 ± 5.4 | 101 | 84.5 ± 0.5 | 101 | 100.0 ± 0.0 |
| S. aureus 1241 | 101 | 87.2 ± 4.6 | 101 | 83.0 ± 2.3 | 101 | 100.0 ± 0.0 |
| L. monocytogenes 149 | 101 | 98.8 ± 2.3 | >101 | 34.2 ± 5.2 | 101 | 99.6 ± 15.6 |
| L. monocytogenes 164 | 101 | 95.8 ± 1.9 | >101 | 68.1 ± 12.9 | 101 | 97.2 ± 2.8 |
| E. coli 683/17 | 50.5 | 78.0 ± 4.5 | >101 | 14.2 ± 24.3 | 101 | 98.9 ± 2.2 |
| E. coli 693/17 | 50.5 | 80.7 ± 2.3 | >101 | 15.6 ± 21.2 | 101 | 97.8 ± 2.2 |
| E. coli 815 | 101 | 95.4 ± 0.2 | >101 | 3.9 ± 6.6 | 101 | 87.6 ± 12.4 |
| E. coli 859 | 50.5 | 81.8 ± 2.9 | >101 | 53.9 ± 1.4 | 101 | 100.0 ± 0.0 |
| S. Infantis S13 | 101 | 98.9 ± 0.1 | >101 | 31.5 ± 27.0 | 101 | 100.0 ± 0.0 |
| S. Infantis S59 | 101 | 99.1 ± 0.5 | >101 | 42.2 ± 13.2 | 101 | 100.0 ± 2.5 |
| BACTERIAL STRAIN |
Control Log (CFU/mL) | PdNPs (22.25 mg/L) Log (CFU/mL) | Log10 CFU Reduction | Inhibitory Rate (%) |
|---|---|---|---|---|
| S. aureus 816 | 9.8 ± 0.0 | 9.0 ± 0.2 | 0.8 ± 0.2 | 81.5 ± 1.3 |
| S. aureus 1241 | 10.6 ± 0.1 | 8.2 ± 0.2 | 2.4 ± 0.2 | 99.0 ± 0.4 |
| L. monocytogenes 149 | 9.1 ± 0.0 | 8.8 ± 0.0 | 0.3 ± 0.0 | 55.2 ± 0.2 |
| L. monocytogenes 164 | 10.6 ± 0.0 | 9.6 ± 0.1 | 1.0 ± 0.1 | 89.3 ± 9.8 |
| E. coli 683/17 | 9.6 ± 0.0 | 8.7 ± 0.0 | 0.9 ± 0.0 | 87.3 ± 0.1 |
| E. coli 693/17 | 10.4 ± 0.1 | 8.8 ± 0.1 | 1.6 ± 0.1 | 97.0 ± 2.0 |
| E. coli 815 | 10.6 ± 0.1 | 8.9 ± 0.1 | 1.7 ± 0.1 | 97.5 ± 1.5 |
| E. coli 859 | 10.8 ± 0.0 | 9.8 ± 0.1 | 1.0 ± 0.1 | 91.2 ± 7.9 |
| S. Infantis S13 | 10.9 ± 0.0 | 9.7 ± 0.0 | 1.2 ± 0.0 | 93.3 ± 0.1 |
| S. Infantis S59 | 10.9 ± 0.0 | 9.9 ± 0.1 | 1.0 ± 0.1 | 89.3 ± 9.8 |
| BACTERIAL STRAIN |
Control Log (CFU/mL) | PtNPs (50.5 mg/L) Log (CFU/mL) | Log10 CFU Reduction | Inhibitory Rate (%) |
|---|---|---|---|---|
| S. aureus 816 | 9.8 ± 0.0 | 8.1 ± 0.0 | 1.7 ± 0.0 | 97.8 ± 0.2 |
| S. aureus 1241 | 10.6 ± 0.1 | 8.6 ± 0.1 | 2.0 ± 0.1 | 99.0 ± 0.4 |
| L. monocytogenes 149 | 9.1 ± 0.0 | 8.3 ± 0.1 | 0.8 ± 0.1 | 83.8 ± 1.6 |
| L. monocytogenes 164 | 10.6 ± 0.0 | 9.7 ± 0.1 | 0.9 ± 0.1 | 89.3 ± 0.1 |
| E. coli 683/17 | 9.6 ± 0.0 | 8.7 ± 0.1 | 0.9 ± 0.1 | 88.2 ± 0.3 |
| E. coli 693/17 | 10.4 ± 0.1 | 8.8 ± 0.1 | 1.6 ± 0.1 | 97.5 ± 0.4 |
| E. coli 815 | 10.6 ± 0.1 | 8.7 ± 0.1 | 1.9 ± 0.1 | 98.5 ± 0.4 |
| E. coli 859 | 10.8 ± 0.0 | 9.0 ± 0.1 | 1.8 ± 0.1 | 98.4 ± 0.0 |
| S. Infantis S13 | 10.9 ± 0.0 | 9.7 ± 0.0 | 1.2 ± 0.0 | 94.0 ± 0.0 |
| S. Infantis S59 | 10.9 ± 0.0 | 9.9 ± 0.0 | 1.0 ± 0.0 | 90.7 ± 0.1 |
| IC50 (mg/L) | ||
|---|---|---|
| Cell Lines | PdNPs | PtNPs |
| HRTEC | >4.45 | >10.1 |
| HaCat | >4.45 | >10.1 |
| HDF | >4.45 | >10.1 |
| HEK 293 | >4.45 | >10.1 |
| HCAEC | >4.45 | >10.1 |
| Bacterial Strain | Origin |
|---|---|
| Staphylococcus aureus 816 | Sea fish |
| Staphylococcus aureus 1241 | Cow milk |
| Listeria monocytogenes 149 | Pork ham |
| Listeria monocytogenes 164 | Pork ham |
| Escherichia coli 683/17 | Salt bath (cheese industry) |
| Escherichia coli 693/17 | Floor (cheese industry) |
| Escherichia coli 815 | Cheese packaging |
| Escherichia coli 859 | Cheese packaging |
| Salmonella enterica Infantis S13 | Wastewater treatment |
| Salmonella enterica Infantis S59 | Frozen chicken meat |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chlumsky, O.; Purkrtova, S.; Michova, H.; Sykorova, H.; Slepicka, P.; Fajstavr, D.; Ulbrich, P.; Viktorova, J.; Demnerova, K. Antimicrobial Properties of Palladium and Platinum Nanoparticles: A New Tool for Combating Food-Borne Pathogens. Int. J. Mol. Sci. 2021, 22, 7892. https://doi.org/10.3390/ijms22157892
Chlumsky O, Purkrtova S, Michova H, Sykorova H, Slepicka P, Fajstavr D, Ulbrich P, Viktorova J, Demnerova K. Antimicrobial Properties of Palladium and Platinum Nanoparticles: A New Tool for Combating Food-Borne Pathogens. International Journal of Molecular Sciences. 2021; 22(15):7892. https://doi.org/10.3390/ijms22157892
Chicago/Turabian StyleChlumsky, Ondrej, Sabina Purkrtova, Hana Michova, Hana Sykorova, Petr Slepicka, Dominik Fajstavr, Pavel Ulbrich, Jitka Viktorova, and Katerina Demnerova. 2021. "Antimicrobial Properties of Palladium and Platinum Nanoparticles: A New Tool for Combating Food-Borne Pathogens" International Journal of Molecular Sciences 22, no. 15: 7892. https://doi.org/10.3390/ijms22157892
APA StyleChlumsky, O., Purkrtova, S., Michova, H., Sykorova, H., Slepicka, P., Fajstavr, D., Ulbrich, P., Viktorova, J., & Demnerova, K. (2021). Antimicrobial Properties of Palladium and Platinum Nanoparticles: A New Tool for Combating Food-Borne Pathogens. International Journal of Molecular Sciences, 22(15), 7892. https://doi.org/10.3390/ijms22157892