This is the first randomized, double-blind, placebo-controlled interventional study which investigated the effect of n-3 PUFAs enriched hen eggs consumption on endothelial microvascular function, and their anti-inflammatory potential in young healthy individuals. First very important finding of this study is increased serum concentration of n-3 PUFAs in participants who consumed n-3 PUFAs enriched eggs, which demonstrates successful dietary protocol.
In particular, the novel finding of this study is that consumption of n-3 PUFAs enriched hen eggs improved endothelium-dependent vasodilation of skin microcirculation independently of BP or body composition and fluid status changes in healthy individuals. Furthermore, observed enhancement in endothelial function was accompanied by a significant increase in IL-10 (anti-inflammatory cytokine), and decrease in INFγ (pro-inflammatory cytokine) serum protein concentration. On the other hand, consumption of regular hen eggs did not significantly affect microvascular endothelial function, but significantly increased IL-17A (pro-inflammatory cytokine) and VEGF-A (vascular endothelial growth factor), as well as IL-10 serum protein concentration. Thus, our study suggests that changes in balance between pro-and anti-inflammatory cytokines could be a potential moderator of enhanced endothelial function following increased consumption of n-3 PUFAs in the form of functional food in healthy population. Furthermore, consumption of hen eggs per se, either regular or n-3 PUFAs, even three pieces a day for three weeks, did not have any harmful effects on metabolic (e.g., serum lipid profile; liver function not assessed), inflammatory or functional vascular parameters (LDF measurement of forearm skin microvascular reactivity) in participants of the study and did not increase CV risk in healthy subjects.
3.1. n-3 PUFAs Enriched Hen Eggs and Serum Lipid Profile
n-3 PUFAs supplementation (EPA and DHA) can reduce serum lipids, most notably triglycerides in hyperlipidemic individuals [
31,
32]. In the present study, consumption of n-3 PUFAs enriched hen eggs did not induce significant change in serum lipid profile compared to baseline measurement; triglycerides level decreased by approximately 8% (
p > 0.05). At the end of the protocol, total cholesterol and HDL cholesterol level were slightly higher in Control group than in n-3 PUFAs group. However, this difference is statistically rather than clinically relevant, since both total and HDL cholesterol levels were within the normal reference range. Thus, we conclude that consumption of 3 eggs per day does not pose risk for unfavorable changes in serum lipid profile for healthy individuals. Our results are consistent with the results of a meta-analysis by Leslie et al. who reported that only very high doses of EPA and/or DHA-enriched food sources (≥4 g/day) have the ability to reduce serum triglycerides (by 9–26%) in normolipidemic to borderline hyperlipidemic and otherwise healthy individuals [
33]. Several studies demonstrated that consumption of n-3 PUFAs enriched hen eggs has the potential to decrease serum triglycerides (but not total, HDL and LDL cholesterol levels) in healthy individuals [
18,
19,
34,
35], but the lack of consistent study design for elevating n-3 PUFAs through dietary modifications, particularly functional food, continues to be a limitation in the field. Interestingly, it has been suggested that higher doses of EPA and DHA may be required to decrease serum triglycerides then are needed to improve endothelial dysfunction [
2], and furthermore, endothelial function may be improved in the absence of a decrease in triglycerides [
2], which is in accordance with the present results.
3.2. n-3 PUFA Enriched Hen Eggs, Blood Pressure Level, Body Composition, and Body Fluid Status
n-3 PUFAs supplementation may induce clinically relevant BP decrease in patients with untreated hypertension [
36], in patients with essential hypertension [
37,
38], and in mildly hypercholesterolemic but normotensive individuals [
39]. However, such significant effect of n-3 PUFAs in lowering BP level was not observed in normotensive individuals [
40,
41], and was not observed in the present study. For example, supplementation of n-3 PUFAs in the form of four fish servings per week (∼800 mg/serving EPA+DHA for 8 weeks) or fish oil supplementation (2 g/day EPA+DHA for 12 weeks; 1.7 g/day EPA+DHA for 4 weeks) in normotensive individuals [
42,
43,
44] did not significantly change BP. Still, our earlier study demonstrated that consumption of n-3 PUFAs enriched hen eggs (777 mg/day) for three weeks significantly reduce BP in healthy normotensive individuals, similar to study of Oh et al. [
34]. Surprisingly, BP level significantly decreased in healthy individuals who consumed three regular hen eggs for three weeks. Thus, decrease in BP could not be attributed to n-3 PUFAs in present study.
The influence of n-3 PUFAs intake on body composition is unclear. One systematic review of clinical trials did not find any important effect of dietary n-3 PUFAs intake on body weight in mostly overweight and obese adults [
45]. Another meta-analysis of only randomized clinical trials that explicitly examined body composition-related measures and used only n-3 PUFAs of fish provenience reported minimal non-significant change in body weight (590 g) and other body composition-related outcomes (such as BMI, body fat percentage and waist circumference) between intervention and control groups [
46]. Even consumption of seven n-3 PUFAs enriched hen eggs per week for 24 weeks did not cause any significant change in body weight compared to baseline in healthy volunteers [
47]. Consistently, in the present study we did not observe any significant change in measured body composition parameters and body fluid status following consumption of n-3 PUFAs enriched hen eggs in healthy lean individuals. Interestingly, body fat mass significantly increased, and fat free mass as well as total body water significantly decreased following consumption of regular hen eggs in the Controls, an observation for which the mechanism is still not clear.
Since in the present study n-3 PUFAs consumption was not accompanied by changes in serum lipid profile, BP level, body composition and body fluid, it seems plausible that improved microvascular endothelial function due to n-3 PUFAs intake is a consequence of the unique effect of increased serum n-3 PUFAs concentration, decreased n-6/n-3 PUFAs ratio, and favorable anti-inflammatory milieu rather than systemic hemodynamic and/or obesity-related changes in healthy individuals, which will be further discussed.
3.3. n-3 PUFA Enriched Hen Eggs and Microvascular Endothelial Function
n-3 PUFAs have the potential to modify vascular endothelial function by being incorporated into endothelial cell membrane phospholipids (at the expense of n-6 PUFAs, such as arachidonic acid), resulting in modulation of different factors which determine endothelial function itself (e.g., synthesis of vasoactive mediators derived from endothelium, oxidative stress level, endothelial activation and inflammation etc.) [
48]. Present study demonstrated significant decrease in serum arachidonic acid at the expense of increased n-3 PUFAs, which may be a source for anti-inflammatory and vasoprotective eicosanoids, contributing to enhanced microvascular response, as observed.
A number of multiple epidemiological, experimental and clinical studies in patients with increased CV risks or already existing CV diseases, such as hypertriglyceridemia, peripheral artery disease, diabetes mellitus type 2, healthy smokers etc. reported that n-3 PUFAs may reduce the risk of CV diseases, at least in part, by improving vascular function [
2]. Detailed overview of these studies in CV patients was recently presented in two reviews by Zehr and Walker [
2], and Du et al. [
49], respectively. Importantly, referenced studies utilized pharmacological supplementation of n-3 PUFAs in form of capsules, but not functional food.
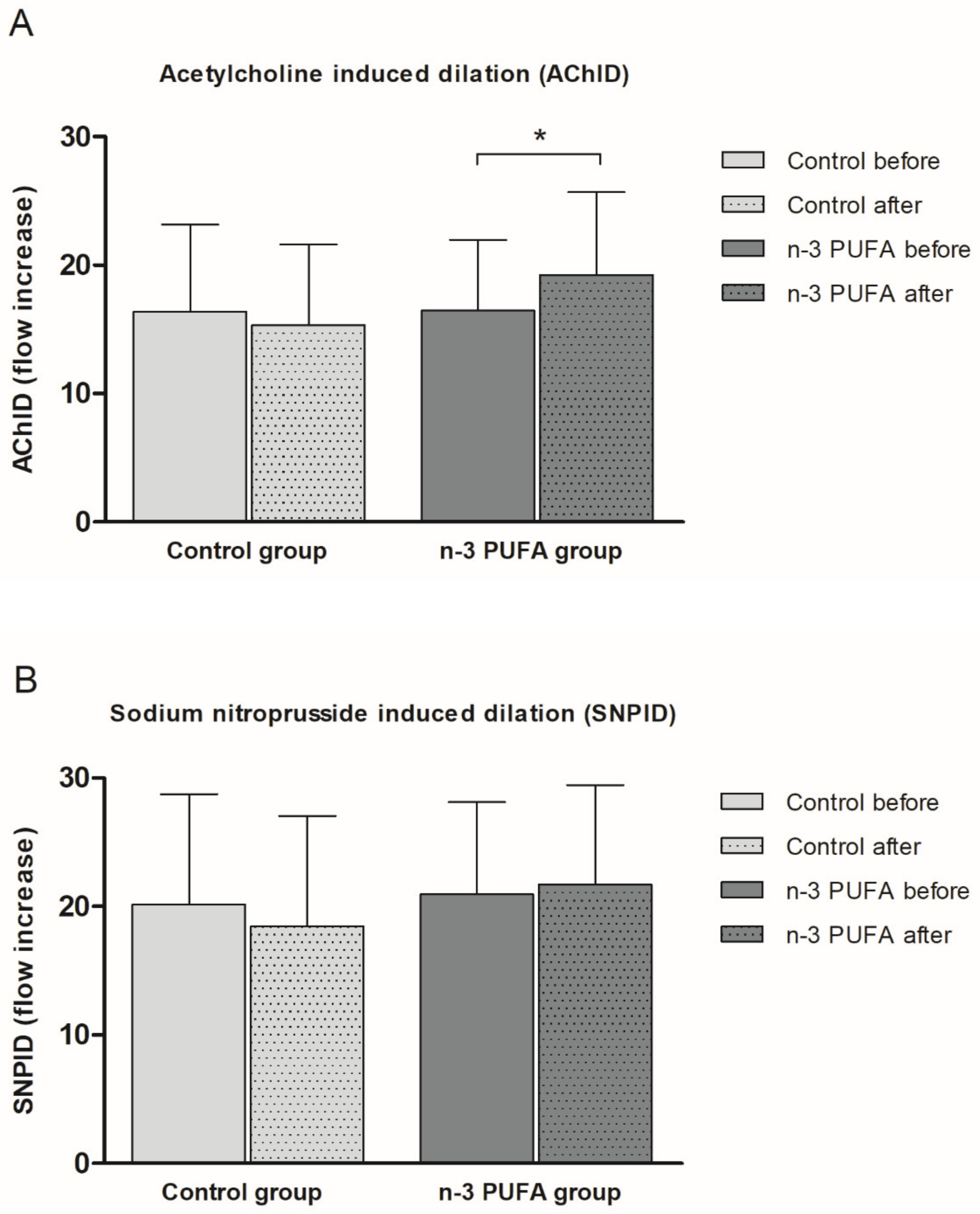
The present study demonstrated, for the first time, that peripheral microvascular endothelial-dependent vasodilation, but not endothelial-independent is improved in response to n-3 PUFAs enriched hen eggs consumption in young healthy individuals. Our results are in agreement with improved endothelium-dependent vasodilation following iontophoretic applications of ACh and SNP in healthy individuals who took fish oil supplementation (EPA + DHA) for 8 months [
50]. In contrast, a randomized cross-over designed study of three diets, each lasting for 4 weeks reported that postprandial forearm skin post-occlusive reactive hyperemia was significantly increased only after monounsaturated fatty acids-rich Mediterranean diet, but not low-fat diet enriched in ALA in 20 healthy men [
51]. On the other hand, previously we reported that young healthy subjects who consumed 777 mg of n-3 PUFAs/day in enriched eggs for 3 weeks had improved skin microvascular reactivity in response to PORH [
19]. All together, these results suggest that a threshold dose of n-3 PUFAs in food/serum needs to be achieved to observe effects in microcirculatory vasodilator responses.
3.4. n-3 PUFAs Enriched Hen Eggs and Pro- and Anti-inflammatory Cytokines, Chemokines, Growth Factors, and Soluble Cell Adhesion Molecules
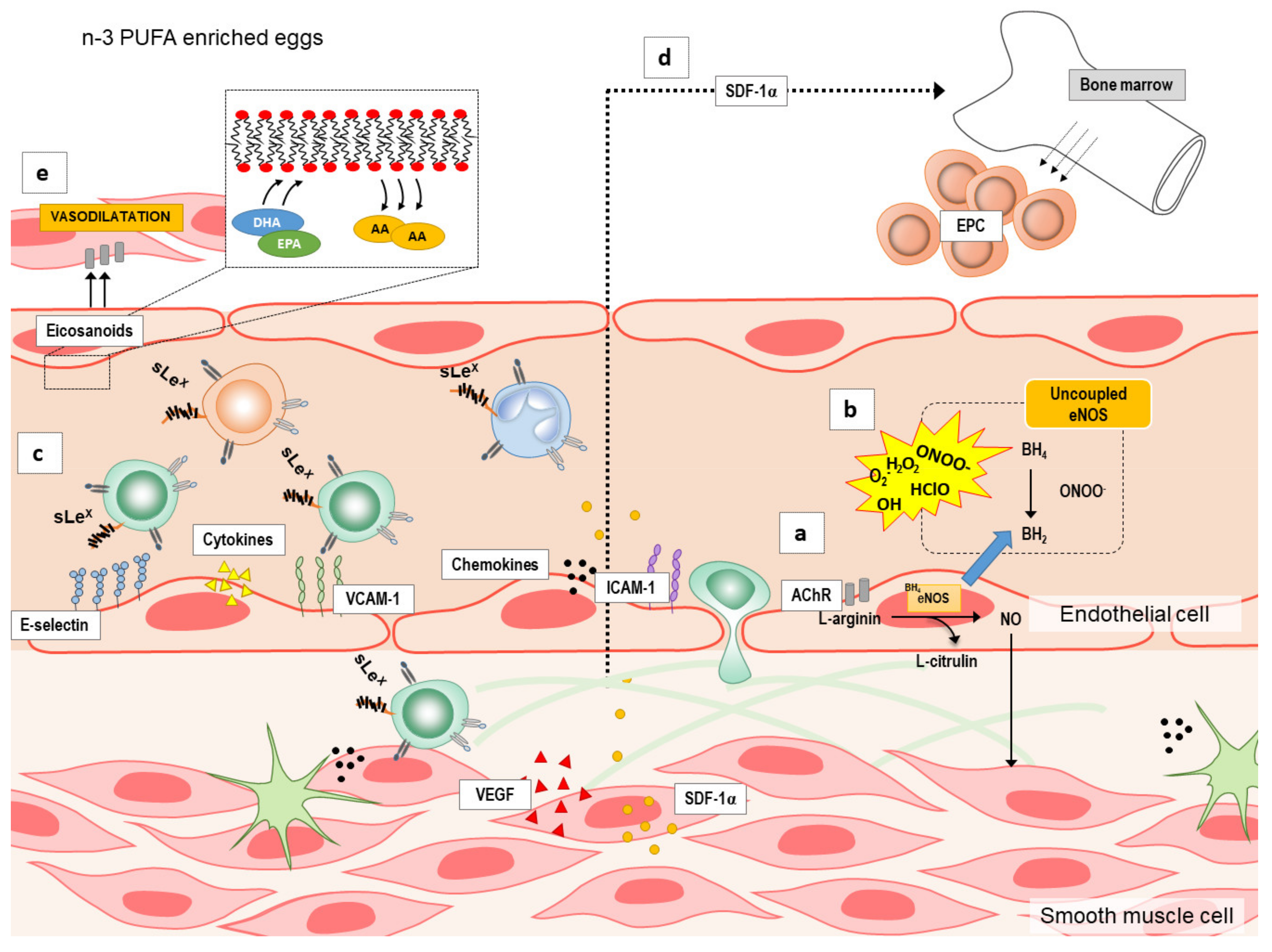
Still, mechanisms mediating endothelium-dependent vasodilation in both micro- and macrovasculature, both in patients and healthy individuals are not known. It has been suggested that n-3 PUFAs may improve endothelial function by increasing the bioavailability of main endothelial vasodilator NO (increasing NO production and/or decreasing oxidative stress level) (
Figure 1a), and/or by changing the expression/activity of other endothelial-derived vasoactive mediators, in particular metabolites of arachidonic acid (e.g., COX metabolites PGI2, PGH2, TXA2; CYP450 metabolites epoxyeicosatrienoic acids, EETs; and 20-hydroxyeicosatetraenoic acid, 20-HETE) (
Figure 1e) [
12,
22,
23,
25,
52,
53].
Additionally, inflammatory stimuli, such as cytokines and chemokines can activate endothelium, as an early event in the initiation of adhesion of monocytes and other leukocytes to the endothelium, preceding their infiltration and extravasation to the site of injury [
54]. Activated endothelium interacts with blood leukocytes, which is characterized by increased expression of cell adhesion molecules (e.g., ICAM-1, and VCAM-1), and increased secretion of chemokines (e.g., LAP, SDF-1α) and pro-inflammatory cytokines (e.g., IL-6 and TNF-α), and with the progression of inflammation with a release of variety of vascular growth factors (VEGF-A and VEGF-D), which under certain circumstances may act as angiogenic mediators that promote atherogenic plaque progression and instability (
Figure 1c,d). On the other hand, VEGF is a potent growth factor for endothelial cells and inducer of angiogenesis, important for endothelial integrity and thus for vascular function [
55]. A previous study by Wu and colleagues showed that fish-oil supplementation enhanced mobilization of bone-marrow derived EPCs independently of eNOS genotype in patients with moderate risk for CV diseases [
21]. In the present study, SDF-1 serum levels were unaffected by diets, suggesting there were no alterations in EPCs numbers in the blood, possibly due to non-existent or low risk for CV diseases in our cohort.
There is a good evidence that n-3 PUFAs can decrease (or prevent) endothelium–leukocyte interactions by decreasing the expression of cell adhesion molecules (ICAM, VCAM, and/or E-selectin) [
54,
56]. The few animal feeding studies in mice and rats, all reported reduced VCAM-1 expression after feeding with EPA and/or DHA. In healthy individuals, n-3 PUFAs may reduce levels of soluble adhesion molecules (e.g., reduced sICAM-1 after supplementation of 2–6.6 g of EPA + DHA daily for 8–12 weeks; reduced sVCAM-1 after supplementation of 1 g EPA + DHA for 12 weeks) [
57,
58,
59]. However, there are also reports on no change (e.g., no change in sICAM-1 and sVCAM-1 after 1.35, 2.7, or 4.05 g EPA daily for 12 weeks in healthy young and older men) [
60], or even increase in sCAMs following n-3 PUFAs intake (e.g., increased sVCAM-1 after 1.3 g DHA and 700 mg EPA daily for 8 weeks in healthy men and women [
61]; increase in sICAM-1 after 1.37 g EPA and 240 mg DHA daily for 8 weeks in healthy individuals) [
62]. The present study demonstrated that consumption of n-3 PUFAs enriched hen eggs (but also of regular eggs) did not induce significant change in sICAM-1 and sVCAM-1 levels in healthy individuals. Thus, we may exclude endothelial activation as a mechanism of action.
Additionally, n-3 PUFAs may be able to influence the levels of circulatory pro- and anti-inflammatory molecules (cytokines, chemokines, and growth factors) directly. n-3 PUFAs intake is associated with reduced concentrations of acute phase protein reactants (CRP), pro-inflammatory eicosanoids, cytokines, chemokines, and other inflammation biomarkers [
56]. Moreover, in addition to inhibiting pro-inflammatory mediators, some animal studies also report that n-3 PUFAs may reciprocally increase the concentration of the anti-inflammatory cytokine IL-10 [
63,
64]. While the results on the effect of n-3 PUFAs on inflammatory molecules in population with increased CV risk remain inconclusive, majority of studies in healthy individuals demonstrated that n-3 PUFAs did not significantly affect the serum concentration of any of the cytokines (e.g., IL-1α, IL-1β, IL-2, IL-4, IL-5, IL-6, IL-8, IL-10, IL-12p70, IL-13, TNF-α, and INFγ) and chemokines (e.g., CCL2, CCL3, CCL5, and CCl11) measured in these studies [
56]. On the other hand, fish oil supplements to healthy individuals decreased the production of TNF-α, IL-1β, and IL-6 by endotoxin-stimulated monocytes or mononuclear cells [
65]. Potential reason may be an insufficient dose of provided n-3 PUFAs (<2 g EPA + DHA), together with basally low levels of cytokines and chemokines in serum of healthy subjects. The latter is supported by the fact that anti-inflammatory effects of n-3 PUFAs have been repeatedly confirmed in studies employing individuals suffering from autoimmune or inflammatory disorders [
66]. To our knowledge, the present study is the first study to report that functional food rich in n-3 PUFAs decreased INFγ and increased IL-10 serum concentration in healthy individuals. Furthermore, consumption of regular hen eggs for three weeks increased IL-10 (less than observed in n-3 PUFA group), but also increased IL-17A and VEGF-A serum concentration in healthy individuals. These results may indicate that unlike the uptake of n-6 PUFAs which leads to an increase in the concentration of pro-inflammatory cytokines (such as IL-17A) and angiogenic growth factors (VEGF-A), uptake of n-3 PUFAs may contribute to maintaining a favorable anti-inflammatory milieu in healthy individuals.


 ,
, 





{kind=link}
{kind=link}