1. Introduction
Diabetes mellitus (DM) is a chronic metabolic disorder that is becoming increasingly prevalent in many developed countries including Malaysia. Diabetic patients with badly controlled glucose levels are highly likely to develop diabetic foot ulcer (DFU) with an incidence rate of 19–34% and DFUs have a tendency to get infected due to the ulcers’ position under the foot [
1]. It has been estimated that the global DFU treatment market alone would increase from 7.03 billion USD to 11.05 billion USD by 2027. The most typical reasons are insufficient glycemic management, latent neuropathy, peripheral vascular disease and neglected foot care. In addition, it is a common cause of foot osteomyelitis and amputation of the lower extremities. Patients are admitted in larger numbers attributable to chronic wounds compared to any other diabetic disease. In the United States, diabetes is currently the most common cause of non-traumatic amputations. Approximately five percent of diabetics develop foot ulcers, and one percent require amputation [
2]. Hyperglycaemia-induced wounds persist in the inflammatory phase and impair wound closure through the disruption of protein synthesis, migration and proliferation of keratinocytes and fibroblasts [
3], thereby impeding the formation of granulation tissue and reducing wound tensile strength.
Oxidative stress greatly contributes to the development of chronic wounds as excessive reactive oxygen species (ROS) generated from the activation of several biochemical pathways including the AGE/RAGE, polyol and hexosamine pathways which induce high oxidative stress that increases advanced glycation end products (AGEs). AGEs have been reported to impair wound contraction and prolong inflammation that hampers ECM proliferation [
4]. High level of MMPs, especially MMP-9 leads to the development of non-healing chronic diabetic wounds. Therefore, antioxidants have been widely studied and applied as an effective treatment strategy in diabetic wound treatment and management.
Wound healing is a complex yet organized biological process that restores the anatomic integrity of the skin upon any form of injury. The healing process involves the interaction of various intracellular and extracellular processes which are mostly regulated by a group of enzymes known as the matrix metalloproteinases (MMPs). MMPs are a family of endopeptidases that are involved in releasing growth factors from the ECM, cleavage of growth factor receptors from the cell surface, and ectodomain shedding of adhesion molecules from membranal proteins of the cell surface [
5].
The hallmark of the proliferative stage in the wound healing process is the rebuilding of new granulation tissue and ECM synthesis. The organization and maintenance of the ECM are highly dependent on various intracellular and extracellular processes which are mostly regulated by a group of enzymes known as the matrix metalloproteinases (MMPs). Four distinct classes of enzymes are associated with wound healing: collagenases, gelatinases, stromelysins and membrane-type metalloproteinases. Although these enzymes play a pivotal role in cell migration and tissue remodelling, an imbalance of these MMPs leads to excessive degradation which has been linked with the nonhealing nature of diabetic ulcers [
6]. As such, MMP inhibitors (MMPIs) are essential in ensuring that the MMP activity is balanced throughout the wound healing process. Many naturally occurring and synthetic MMPIs have been explored for wound healing. Some examples of natural MMP inhibitors include curcumin, resveratrol, theaflavin and catechin derivatives [
7].
However, with the evolution of technology such as artificial intelligence, molecular docking and in silico studies, the search for effective natural compounds has become more productive and efficient.
Marantodes pumilum var. alata, popularly known as Kacip Fatimah, is a Malaysian herb that has been widely used in traditional medicine by women to facilitate post-partum recovery, relieve menstrual problems and treat dysentery. Previous studies have reported
M. pumilum to possess antibacterial, anti-inflammatory, antioxidant and xanthine oxidase inhibitory properties [
8,
9,
10].
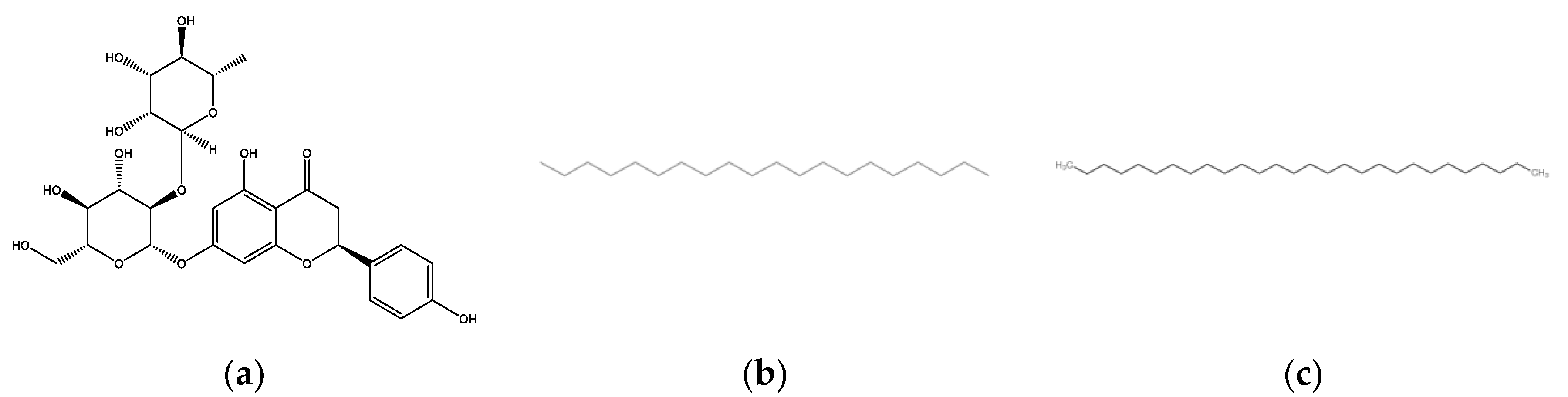
In this study, naringin (NAR), eicosane (EIC) and octacosane (OCT) identified and isolated from the DCM extract of M. pumilum, were evaluated for their molecular wound healing and antioxidant activity using in vitro, in vivo and in silico approaches.
3. Discussion
The prevalence of diabetes mellitus (DM) and its consequences are increasing worldwide, placing a significant burden on individuals and healthcare systems [
12]. The effects of diabetic wounds on disability, morbidity and mortality are considerable. The treatment of diabetic wounds remains a significant concern for the medical system. Reports indicate that minimal amounts of reactive oxygen species (ROS) are essential to prevent external damage, but its overproduction leads to the formation of diabetic wounds [
13]. Redox imbalance is a leading cause of non-healing diabetic wounds [
14] due to an excess of reactive oxygen species (ROS) in tissues and a loss in antioxidant capacity. Long-term type 2 diabetes is associated with severe decreases in antioxidant enzyme activity [
15]. Glutathione (GSH) plays an important role in maintaining the redox status of the cell. Reduced GSH levels have been noticed in diabetic wounds due to the disturbance in the redox state of the cell that causes metabolic disturbances as seen in diabetic patients [
16]. Some researchers have hypothesized that a hyperglycemic state directly impacts the function of keratinocytes and fibroblasts, causing alterations in protein synthesis, proliferation, and migration, decreased antimicrobial peptide production and increased oxidative stress [
17,
18,
19,
20].
Matrix metalloproteinases (MMPs) are zinc-dependent proteolytic enzymes implicated in collagen degradation, cellular interactions and cell-matrix interactions via modulating the levels of cytokines, growth hormones and various biological fragments concealed in ECM [
21,
22,
23,
24,
25,
26]. Imbalance and dysregulation of these enzymes lead to excessive degradation, which relates to non-healing wound processes and the conversion of acute wounds into chronic wounds. Elevated MMPs cause decreased expression of MMP tissue inhibitors (TIMPs). TIMPs are found to be reduced in diabetic chronic wounds which aggravates the problem [
27,
28,
29,
30]. The imbalance between MMP and TIMP levels is a major cause of hyperglycemia, hyperlipidemia and hypertension in diabetic ulcers and chronic wounds [
31,
32]. The overexpression of MMP-13 and MMP-9 is linked to a high concentration of glucose at the wound site [
33].
Elevated levels of matrix metalloproteinases (MMPs) have been identified in these wounds because of oxidative stress and glycation end products, which may lead to diabetic peripheral artery disease [
34,
35]. MMPs are primarily responsible for proteolytic degradation, which has a significant impact on the chronic wound healing process. The deterioration of extracellular matrix (ECM) induced by MMPs, specifically MMP-1, -2, and -9, renders these diabetic wounds worse [
36]. In chronic wounds, the mismatch between the level of ECM breakdown and repair is a crucial component causing a delay in the wound healing process. Disproportionate availability of cytokines and other growth factors are needed for wound healing [
37,
38]. In the absence of TIMP, MMP-1, -8 and -9 are reported to be upregulated in venous wounds [
39,
40]. To ensure the balance of MMPs, it would require suppressing the effect of MMP-2 and MMP-9 along with upregulating the expression of matrix metalloproteinase inhibitors such as TIMP-1 and TIMP-2 [
41]. A second target would be to boost MMP-8 while inhibiting MMP-9. MMP-8 has been demonstrated to accelerate the healing of diabetic wounds in the absence of MMP-9 and vice versa, thus necessitating the inhibition of MMP-9 without affecting MMP-8. Another target is to cleave the MMP-3 zymogen, which is required for MMP-9 activation [
42,
43,
44,
45].
In a diabetic rat wound model, the administration of naringin, eicosane and octacosane promoted faster wound healing. Naringin, eicosane and octacosane may promote wound healing due to their potent free radical scavenging, hydroxyproline and glutathione action (antioxidant) as shown in
Table 1 and
Table 3. Multiple research projects have reported a link between ROS and the development of diabetic wounds. Antioxidants prevent the evolution of diabetic neuropathy by scavenging reactive oxygen species (ROS), regenerating endogenous and exogenous antioxidants, renovating oxidised proteins, inhibiting NF-kB and regulating gene transcription. Antioxidants are commonly employed in clinical settings to treat diabetic microangiopathy because a balanced redox state is likely essential for rapid repair [
46,
47,
48].
Naringin is a water-soluble flavonoid that is present in many citrus fruits whereas eicosane and octacosane are long-chain hydrocarbons that are naturally found in plants such as
Taraxacum officinale,
Hypericum hircinum and
Acacia nilotica [
49]. Flavonoids are secondary metabolites that are abundant in various plants with over thousands of individual flavonoid compounds identified up to date. The antioxidant action of flavonoids is frequently studied due to their capability as potent free radical scavengers. Naringin is a flavanone glycoside containing naringenin, a well-studied flavonoid for its antioxidant activity. Flavonoids generally have a molecular structure of a C6-C3-C6 carbon skeleton that comprises two 6-carbon benzene rings linked by a 3-carbon heterocyclic ring [
50]. The free radical scavenging ability of naringin is strong due to the location and number of -OH groups and the conjugation and resonance effects between these groups [
51]. Naringin has been reported to be effective in reducing oxidative stress in both in vitro and in vivo models [
52,
53,
54,
55].
Hydrocarbons, on the other hand, have not been extensively studied for their antioxidant activity. The findings of this study showed that eicosane and octacosane exhibited moderate antioxidant activity as compared to naringin. The reduced antioxidant activity of hydrocarbons is due to the lack of hydroxyl groups as they are mainly composed of carbons and hydrogens. However, a study by Farzaliyev (2012) showed that the antioxidant activity of hydrocarbons can be greatly improved when combined with functional groups that react with peroxide radicals such as phenols and aromatic amines and additives that decompose hydroperoxides such as sulphides [
56].
Epidermal growth factor (EGF) administration improves wound healing because of its alleviation of oxidative stress [
57]. Biogenic AgNPs synthesized from
Brevibacillus brevis KN8 could inhibit the overexpression of MMP-2 and MMP-9 in granulation tissues and accelerate wound healing in diabetic mice beyond the antimicrobial activity [
58]. Ferulic acid (FA) is a natural antioxidant derived from fruits and vegetables that inhibited lipid peroxidation and increased the expression of catalase, superoxide dismutase, glutathione, nitric oxide and serum zinc and copper, which probably improves the healing process in diabetic ulcers [
59]. Syringic acid treatment also promotes migration and proliferation to improve wound healing [
60]. The fusion protein decreased serum proinflammatory cytokines such as IL-6, TNF-α, expression of cyclooxygenase-2, and increased activities of antioxidant enzymes including superoxide dismutase, glutathione peroxidase and catalase, and it also increased proangiogenic cytokine levels including VEGF, intercellular adhesion molecule, and expression of VEGF, FGF-2, p-ERK and p-Akt protein in granulation in diabetic rats, which significantly accelerated the diabetic wound healing [
61]. Chlorogenic acid, a dietary antioxidant that could accelerate wound healing, enhances hydroxyproline content, decreases malondialdehyde/nitric oxide and elevates the level of reduced glutathione in the wound bed [
62].
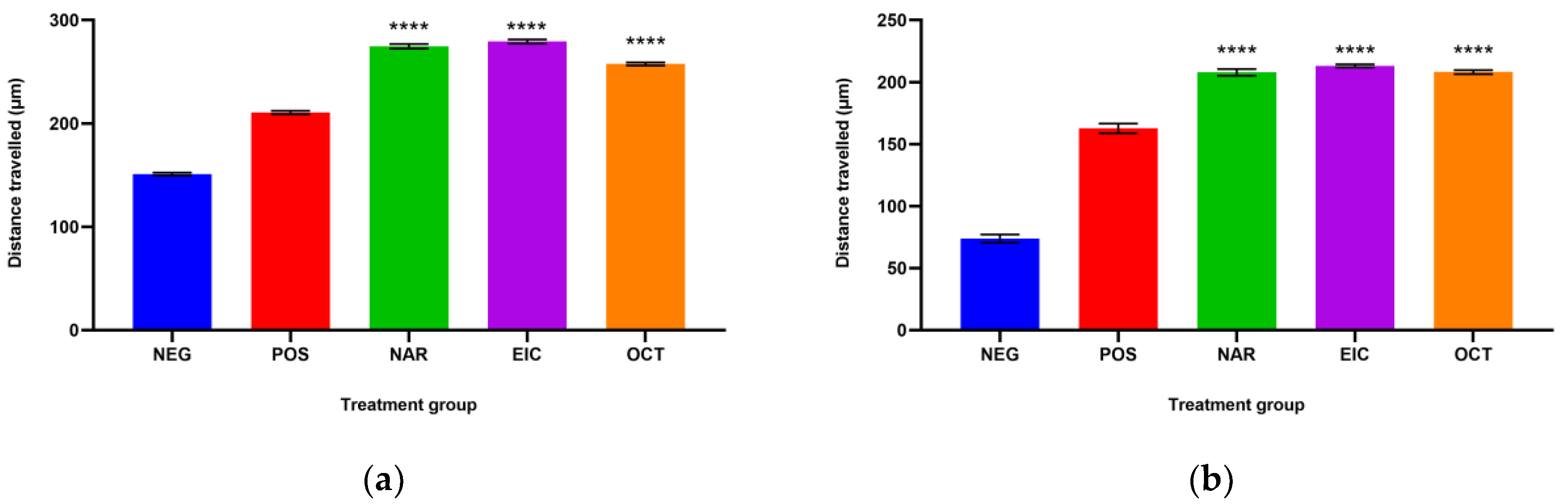
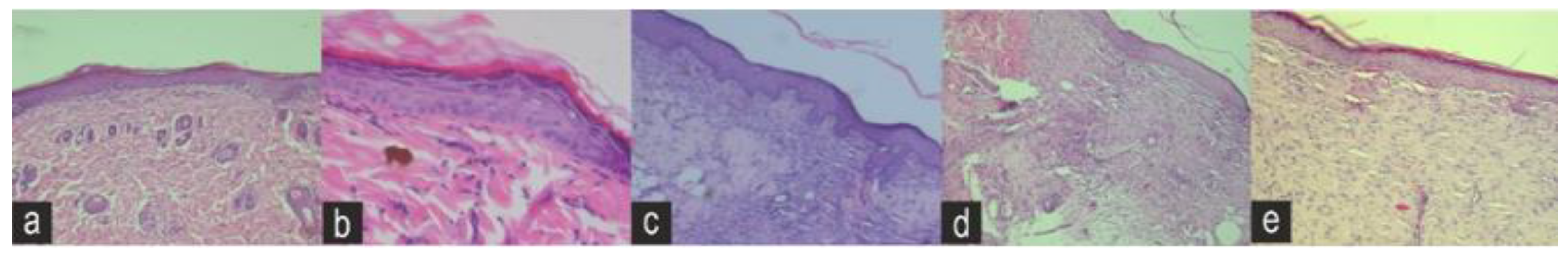
The fibroblast migration and re-epithelialization activities of naringin, eicosane and octacosane were similar to each other as observed in
Figure 1 and
Figure 2 (in vitro) and
Figure 3,
Figure 6,
Figure 7 and
Figure 8 (in vivo). Fibroblasts play an important role in the proliferative phase of the wound healing process and are involved in ECM synthesis along with other growth factors such as transforming growth factor (TGF-β1) and basic fibroblast growth factor (b-FGF). The multiplication of skin fibroblasts is essential for the earliest stages of wound healing [
63]. The effects of naringin, eicosane and octacosane on epithelialization, fibroblast migration, and polymorphonuclear leukocyte migration (PMNL) are depicted in
Figure 8.
Figure 1 and
Figure 2 show that insulin-resistant HDF cells exhibited enhanced fibroblast migration. Epithelialization, fibroblast movement and polymorphonuclear leukocyte migration (PNML) were detected in the naringin, eicosane and octacosane-treated group, possibly because of an increase in collagen synthesis, fibroblast proliferation and tumour necrosis factor-alpha (TNF-) upregulation or by activating the collagen VI expressions and transforming growth factor-beta 1 (TGF-1) as well as its antioxidant capacity [
64,
65].
These substances caused the cohesion of collagen fibres in the dermis and made connective tissue cells visible in the fibres’ labia. Cellular events for wound repair [
66] include angiogenesis, cell migration and proliferation, creation of granulation tissue, collagen synthesis, and re-epithelialization. Consistent with previous studies, treatment of animals with naringin, eicosane and octacosane resulted in increased wound capillary density, re-epithelialization, fibroblast proliferation, and collagen synthesis [
67,
68]. The interleukins IL-1, IL-6 and TNF- enhance the inflammatory response; collagen production is mediated by FGF-b, IGF and TGF-; and angiogenesis is stimulated by FGF-B, VEGF components A, TGF- and HIF-1 [
69,
70].
Wound healing is a multicellular process that includes fibroblasts, keratinocytes, endothelial cells and inflammatory cells. Haemostasis, the inflammatory phase, the proliferation phase and the remodelling phase comprise four different yet overlapping phases of the healing process. Crosstalk between different groups of molecules, including extracellular matrix (ECM), integrins, growth factors and MMPs, regulates the phases of wound healing. The migration of cells on extracellular matrix (ECM) and modification and destruction of ECM by matrix metalloproteinases (MMPs) are essential components in wound repair [
71]. MMPs can be categorised into ‘Clans’ and ‘Families’ based on fold similarity and evolutionary ties, respectively. The MMP class is comprised of eight clans and about forty families. In accordance with the substrate specificity and homology organisation: (1) Collagenases (MMP-1, MMP-8, MMP-13), (2) Gelatinases (MMP-2 and MMP-9), (3) Stromelysin (MMP-3, MMP-10, MMP-12), (4) Matrilysin (MMP-7, MMP-26), (5) Membrane Type (MT) MMPs (MT-MMP-14, -15, -16, -17, -24, -25) and (6) Additional MMPs (MMP-19, -20, -21, -22, -23, -27, -28). In accordance with the MMPs’ organisational structure: (1) the prototypical MMP (type-1 collagenases), (2) Matrilysins are missing the hemopexin domain, (3) Gelatinases have three type II fibronectin domains and (4) MT-MMPs are localised at the cell membrane surface [
72,
73].
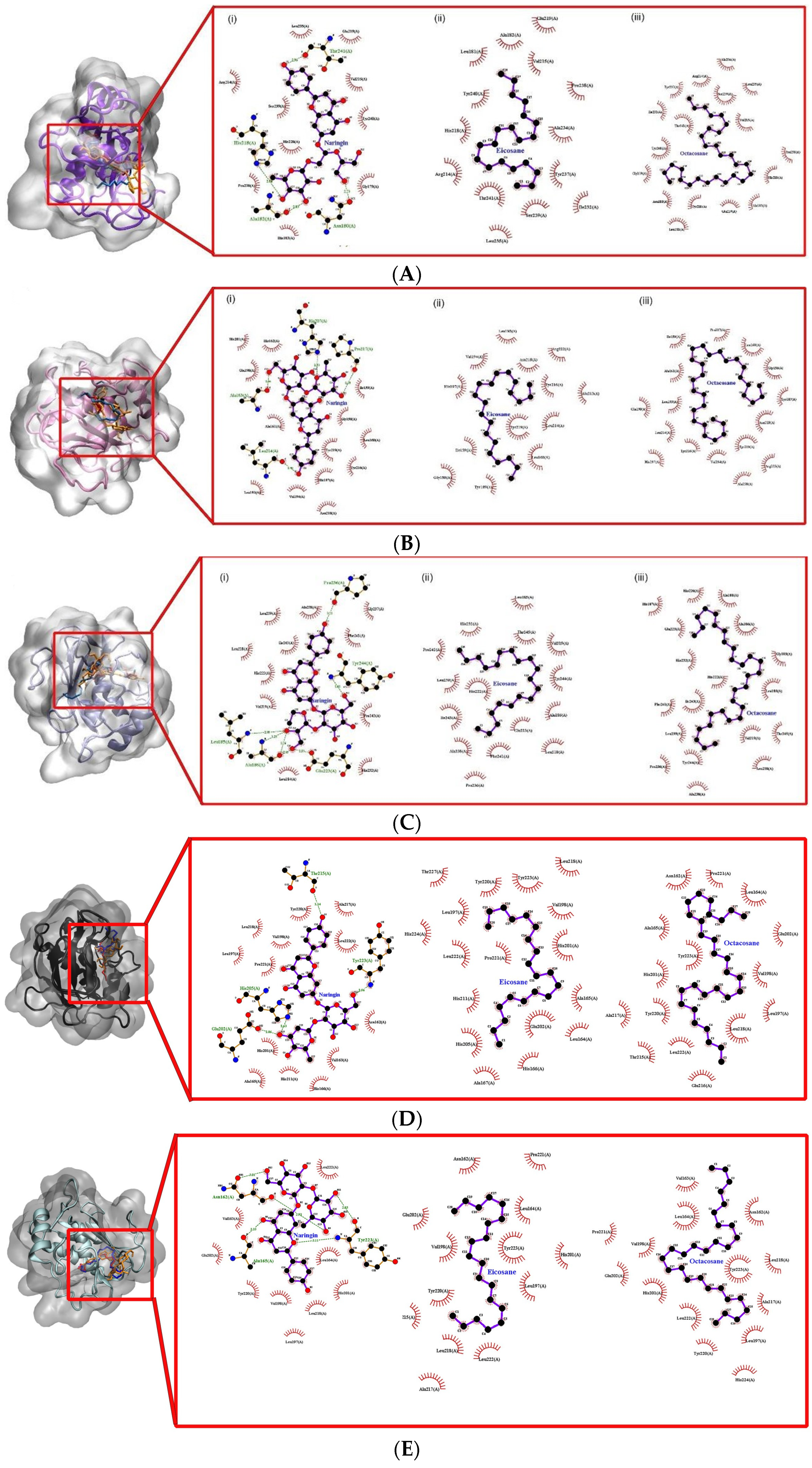
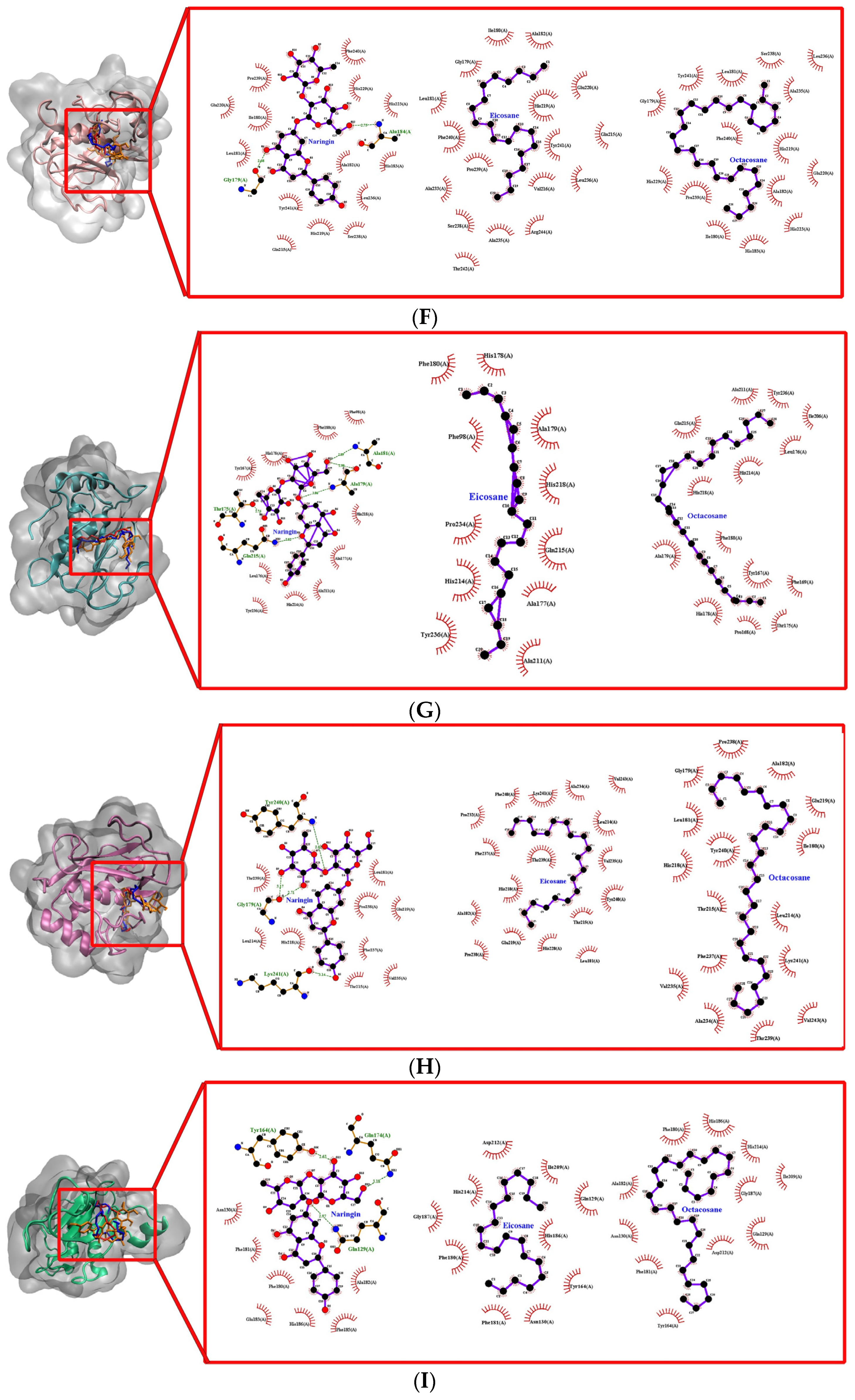
The molecular docking approach assists to model the interaction between a small molecule and a protein at the atomic level, which allows one to characterize the behaviour of small molecules in the binding site of target proteins as well as to elucidate fundamental biochemical processes [
27]. The docking process involves two basic steps: prediction of the ligand conformation as well as its position and orientation within these sites (usually referred to as pose) and assessment of the binding affinity. These two steps are related to sampling methods and scoring schemes, respectively, which will be discussed in the theory section. It provides a prediction of the ligand-receptor complex structure, binding energy and affinity using computation methods. The molecular docking result reveals a strong interaction and affinity between the compounds and MMPs. It shows the compounds’ high chance of modulation of the MMPs. Pharmacologically, the higher the binding affinity, the higher the target occupancy and the higher the chance of activation. This result is in consonance with the observed result. There was evidence of collagen synthesis, fibroblast migration and re-epithelization which are evidence of MMP activation.
The molecular docking performed in this study showed the binding energy of naringin was the highest for collagenases I, II and III against the control, followed by octacosane and eicosane. All three drug compounds showed the highest binding energy with collagenase III compared to collagenases I and II. According to Armstrong and Jude (2002), MMP-13 has a unique ability to cleave type I, II and III collagen [
74]. A study by Toriseva et al. (2012) reported that MMP-13 is vital in wound healing as it coordinates the growth of wound granulation tissue and modulates the expression of genes involved in inflammation, proteolysis and cell viability [
34]. Collagenase-1, 2 and 3 along with stromelysin 1 and gelatinase B play a significant part in the proliferation and re-epithelization phases of wound healing. It also aids in the granulation of tissue to cover the wound with a dense network of blood vessels. These arteries stimulate the release of platelet-derived growth factors, which aid in wound debridement and reorganisation of damaged type I collagen. It affects the release of inflammatory cytokines, including as IGN and TGF-, as well as the synthesis of collagen and fibroblast synapses [
74,
75,
76,
77,
78,
79,
80,
81]. The vascular endothelial growth factor (VEGF) secreted by macrophages stimulates the migration and proliferation of keratinocytes and endothelial cells, which comprise MMP-1, 2, 9 and 13, and, therefore, play a significant role in wound healing [
82,
83].
Gelatinases, such as gelatinase A (MMP-2) and gelatinase B (MMP-9), promote the migration of keratinocytes and the healing process. MMP-9 maintains the wound-healing factors such as vascular endothelial growth factor and dermatopontin [
84,
85,
86]. Gelatinases (MMP-2 and MMP-9) function in keratinocyte migration and angiogenesis. Gelatinase A (MMP-2) is secreted by fibroblasts while gelatinase B (MMP-9) and is produced by leukocytes and possibly, keratinocytes. The binding energies of naringin, eicosane and octacosane were relatively stronger with gelatinase B than with gelatinase A. A study by Hingorani et al. (2018) showed that wound healing was delayed in MMP-2/-9 double knockout (DKO) mice as compared to the wild-type group which suggests that this poor response observed in DKO mice could be attributed to the loss of MMP-9 as it is usually expressed at the leading edge of migrating cells that allows these cells to reepithelialise the wounded site [
87]. While several studies support the positive role of MMP-9 in wound healing, it is interesting to note the findings from a study by Reiss et al. (2010) that postulated MMP-9 could be a causal factor of chronic wound development as active MMP-9 expression showed a significant decrease in epithelial migration and type IV collagen [
88].
Stromelysins are involved in the degradation of the extracellular matrix. The binding energies of all three drug compounds showed the strongest for stromelysin 1 (MMP-3) and stromelysin 2 (MMP-10). Stromelysins 1 and 2 can cleave several types of collagens, proteoglycans, laminin and fibronectin [
89]. Among the tested drug compounds, naringin showed the strongest binding energies with stromelysins 1 and 2, followed by eicosane and octacosane. Stromelysin 1 is responsible for the initiation of wound contraction through the migration of fibroblasts, and downregulation of stromelysin 1 is clearly attributed to delayed wound healing as observed in chronic wounds [
90]. Stromelysin 2 is primarily involved in epithelial cell migration and its activity is influenced by cytokines, depending on the cell type. Salmela et al. (2004) reported an increase in stromelysin 2-expressing epithelial cells when cytokines TNF-α or IL-1β was added to the culture media [
91].
MMP therapy targets a variety of synthetic and natural substances. Numerous broad-spectrum MMP inhibitors (MMPI) have been studied for this purpose, but we require more specific treatments than those currently available [
92]. Compounds such as (R, S)-ND-336, antibodies GS-5745, SSDS-3 and REGA-3G12 and atelocollagen type I, when combined with 4-vinyl-benzyl chloride -CD (D3)7/MMP-9-siRNA, reduce MMP-9 activity in diabetic wounds [
84,
93,
94,
95,
96]. Withaferin A (3-azido Withaferin A) is a naturally occurring steroidal lactone derived from the plant
Withania somnifera, while Cantharidin is a naturally occurring chemical derived from the plant
Mylabris phalerata. Celastrol is an additional phytoconstituent of
Tripterygium wilfordii with anti-proliferative activity. Ginsenoside Rd is derived from
Panax ginseng leaf, and lycorine is a naturally occurring substance that is widely dispersed in the
Amaryllidaceae plant family. All these naturally occurring plant extracts modulate the activity of the MMPI [
97,
98,
99,
100,
101,
102]. Naringin is a flavonoid found naturally in citrus fruits [
103]. Treatment with naringin decreases the expression of p-ERK and p-JNK, which are molecular markers implicated in the MAPK signalling pathway, hence decreasing the expression of MMP-2 and MMP-9. Gallic acid has also been reported to activate the natural MMP-2 regulator TIMP-1 [
100].
The figure below is a schematic diagram of the mechanism of action of naringin, eicosane and octacosane in the in vitro and in vivo models, and the in silico assessment conducted on the MMPs involved in wound healing (
Figure 12).


 ,
, 

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}