Abstract
The Angiotensin-I-converting enzyme (ACE) is a peptidase with a significant role in the regulation of blood pressure. Within this work, a systematic review on the enzymatic preparation of Angiotensin-I-Converting Enzyme inhibitory (ACEi) peptides is presented. The systematic review is conducted by following PRISMA guidelines. Soybeans and velvet beans are known to have high protein contents that make them suitable as sources of parent proteins for the production of ACEi peptides. Endopeptidase is commonly used in the preparation of soybean-based ACEi peptides, whereas for velvet bean, a combination of both endo- and exopeptidase is frequently used. Soybean glycinin is the preferred substrate for the preparation of ACEi peptides. It contains proline as one of its major amino acids, which exhibits a potent significance in inhibiting ACE. The best enzymatic treatments for producing ACEi peptides from soybean are as follows: proteolytic activity by Protease P (Amano-P from Aspergillus sp.), a temperature of 37 °C, a reaction time of 18 h, pH 8.2, and an E/S ratio of 2%. On the other hand, the best enzymatic conditions for producing peptide hydrolysates with high ACEi activity are through sequential hydrolytic activity by the combination of pepsin-pancreatic, an E/S ratio for each enzyme is 10%, the temperature and reaction time for each proteolysis are 37 °C and 0.74 h, respectively, pH for pepsin is 2.0, whereas for pancreatin it is 7.0. As an underutilized pulse, the studies on the enzymatic hydrolysis of velvet bean proteins in producing ACEi peptides are limited. Conclusively, the activity of soybean-based ACEi peptides is found to depend on their molecular sizes, the amino acid residues, and positions. Hydrophobic amino acids with nonpolar side chains, positively charged, branched, and cyclic or aromatic residues are generally preferred for ACEi peptides.
1. Introduction
Hypertension is a high prevalence disease and is considered one of the major health problems globally [1]. Lim et al. [2] reported that cardiovascular diseases due to complications of hypertension account for 9.4 million deaths every year. It is therefore of importance to take the appropriate mitigations to reduce the mortality rate due to hypertension. Otherwise known as high blood pressure, hypertension is a medical condition where the arterial blood pressure (BP) is abnormally high. According to the 2019 ACC/AHA Guideline on the Primary Prevention of Cardiovascular Disease [3], a normal BP is described as having a systolic and diastolic pressure of less than 120 and 80 mmHg, respectively (BP < 120/80 mmHg). There are two stages of hypertension. Stage 1 is defined with BP 130–139/80–89 mmHg, while stage 2 hypertension is for BP ≥ 140/90 mm Hg [3]. As mentioned above, hypertension could lead to cardiovascular diseases and stroke. Hypertension is usually treated with blood pressure regulating drugs such as angiotensin-I-converting enzyme (ACE) inhibitors (e.g., lisinopril, captopril), vasodilator, etc. Given the side effects of synthetic ACE inhibitors (e.g., taste disturbances, cough, and swelling of the lower layer of human skin or angioneurotic edema) [4], various investigations have been afforded to find potent ACE inhibitors from natural products, especially from food proteins. As a result of the increasing interest regarding functional foods in the past few years, it has been reported that food proteins-derived bioactive peptides have several benevolent effects on human health, including inhibitory activity against ACE [5,6]. Therefore, bioactive peptides can be considered as an alternative for managing hypertension.
A bioactive peptide is defined as an organic compound with a positive impact on human health (e.g., inhibitory activity against ACE, antioxidant capacity, antimicrobial activity, anti-thrombotic, immunomodulatory, etc.) which consists of 2–20 amino acids joined by covalent bonds called peptide bonds [7]. In the digestive system of the human body, bioactive peptides are liberated by digestive proteases, such as pepsin or microbial enzymes. Additionally, processing food and ripening can release bioactive peptides.
Nature is an abundant source of bioactive peptides produced by organisms such as plants and animals. Although animal products remain the greatest source of bioactive peptides, this work will mainly discuss plant source bioactive peptides derived specifically from soybean and velvet beans. Soybean nutritional content consists of 35–40% protein, 20% lipids, and 9% dietary fiber based on dry-weight soybean [8,9]. Because of its high protein content, soybean is mostly utilized as a source of bioactive peptides among other plants. Meanwhile, a less well-known type of legumes called velvet bean has a nutritional content of approximately 25% protein and 14% crude fat based on its dry weight [10]. As both beans are considered as potent protein sources in the human diet, their utilization as sources of parent proteins for producing bioactive peptides is promising. However, in the case of velvet bean, studies related to its utilization as a parent protein source are scarce. Thus, it is important to elucidate the technological approach of producing velvet bean-derived peptides especially for inhibiting ACE activity.
Highlighting the elevating interest in the bioactive peptides as part of the diet and the higher prevalence of the non-communicable disease of hypertension, this systematic review discusses the advances in the enzymatic preparation of bioactive peptides from soybean and velvet beans exhibiting Angiotensin-I-Converting Enzyme inhibitory (ACEi) activity. This study focuses its discussion on the optimum hydrolytic conditions required to produce ACEi peptides and the influence of peptides’ intrinsic properties (i.e., amino acid residues and their arrangement in the sequence, molecular weight, hydrophobicity) on the ACEi activity.
2. Results
2.1. Substrate Preparation as Source of ACEi Peptides from Soybean and Velvet Bean
The preparation of substrates from soybeans is rarely discussed in the literature. Substrates from soybeans as sources of parent proteins can be soy protein concentrate or isolate, soybean flour-rich in protein, and principal soybean storage proteins (i.e., glycinin or β-conglycinin). Gouda et al. [11] prepared the soy protein substrate, glycinin. This method follows a previously described method developed in a study by Rao and Rao [12] with the use of (NH4)2SO4 precipitation and centrifugation. Water containing β-mercaptoethanol (0.1% v/v) is used to extract defatted soybean flour for 4–6 h under constant agitation. The solution is then centrifuged at 6000–8000 rpm for 45 min at 25 °C, followed by the addition of dry MgCl2 until the final MgCl2 concentration in the solution reaches 5 mM. Glycinin is collected by centrifugation, and the precipitate is dried with a freeze drier. Freeze drying is used as a preferred water removal method because it has the advantage to cause less damage to the structure of the protein substrate. Nevertheless, the fractionation of glycinin in most studies involves the precipitation of the alkaline soy protein extract at pH 6.3–7.0 [13,14,15].
For the preparation of the velvet bean substrate, wet fractionation is the method that is commonly used [16,17,18]. Initially, velvet bean flour is prepared by grounding the grains with a disk mill followed by sieving. The prepared bean flour then undergoes suspension in 3% sodium bisulfite with a 1:6 ratio (w:v) and left to soak for an hour with a constant agitation under alkaline pH (pH = 8). The role of sodium bisulfite is to increase the solubility of the velvet bean protein. Abtahi and Aminlari [19] stated that the modification of protein with a chemical treatment, such as sodium bisulfite, increases the protein dispersibility index (PDI). After fiber solid separation and washing with 3% sodium bisulfite, the protein-starch suspension is then left to sediment for 30 min. The purpose of sedimentation is to recover starch. The pH of protein solution pH is adjusted to an isoelectric point (i.e., pH 4.2) using 1.0 M HCl solution. The precipitate is obtained by centrifuging the solution at 1317× g for 20 min and further dried using a freeze-drier at −47 °C and pressure of 13 × 10−3 mbar [16,17,18]. In another study by Mugendi et al. [20] who characterized the nutritional properties of velvet bean protein isolate, the extraction was conducted with distilled water at pH 9 followed by centrifugation. The pH of the extract was then adjusted to 4.5 to precipitate the protein.
2.2. Hydrolytic Conditions for Producing ACEi Peptides from Soybean and Velvet Bean Protein Substrates
Enzymes for proteolysis are classified as endopeptidases and exopeptidases, based on the site of action on the substrate. Exopeptidases hydrolyze at the N- or C-terminal ends of the peptide, while endopeptidases cleave peptide bonds within and distant from the ends of a polypeptide chain or at the non-terminals of the sequence [21]. The most common enzymes used for producing soybean-based bioactive peptides are pepsin [22,23], papain [6,24], alcalase [25,26,27], proteinase from M. purpureus [28], trypsin, chymotrypsin, ginger protease, and Amano Protease from Aspergillus sp. [11], and protease D3 from E. coli strain JM109 [29]. All of these enzymes are endopeptidases. Endopeptidases, such as alcalase and proteinase K produce short-chain hydrophobic amino acids which are preferred in enhancing ACEi activity [21]. Additionally, prolyl endopeptidases such as Protease P from Aspergillus niger are often used as it can yield in proline-containing bioactive peptides which are favored for their strong affinity to ACE [30]. Hydrolytic conditions of soybean proteins for producing ACEi peptides are shown in Table 1.

Table 1.
Enzymatic hydrolysis conditions of soybean proteins using endopeptidases to produce ACEi peptides.
For velvet bean, the proteolytic enzymes reported limitedly in the literature are a combination of pepsin-pancreatin [16,17,18] and alcalase–flavourzyme [16,17]. In contrast to soybean-derived peptides, for velvet bean sourced peptides, the hydrolysis is conducted with a combination of both endopeptidase and exopeptidase. The application of both endo- and exo-peptidase allows it to have a broad cleavage action and produce a shorter chain of peptides. Table 2 shows the enzymatic hydrolysis conditions of velvet bean-derived proteins.
Table 2.
Enzymatic hydrolysis conditions of velvet bean protein concentrate to produce ACEi peptides.
3. Discussion
3.1. Hydrolytic Conditions of Soybean-Based Bioactive Peptides Preparation
Besides fermentation, bioactive peptides can also be produced by the hydrolytic activity of proteases on soybean parent proteins [24]. Important factors to consider in producing soybean-derived bioactive peptides through proteolysis are the type of enzyme, reaction temperature, time of hydrolysis, pH, and enzyme-substrate ratio (E/S) [32]. At a low enzyme-to-substrate (E/S) ratio, the enzyme will continuously cut the most susceptible peptide bonds during the hydrolysis period. Meanwhile, with the increase of E/S ratio, cleavage action is faster during the initial stage of hydrolysis and becomes slower at a later stage. This is because at the initial stage, the reaction is spent by rapid cleavage of the susceptible peptide bonds and at a later stage, enzymes degrade the less susceptible peptide bonds [33]. The reported E/S ratios for producing ACEi peptides from soybean proteins are mostly less than 10%, with majority at 4 and 6%.
Among others, common proteolytic enzymes used for the production of bioactive peptides from soybeans include pepsin, alcalase, and protease D3. Pepsin is a protease that hydrolyzes peptide bonds between the aromatic amino acids such as phenylalanine, tryptophan, and tyrosine [34]. It is classified as an endopeptidase. Wang et al. [35] stated that pepsin effectively degrades proteins between pH 1.2–2.5 and the optimal pH for pepsin’s proteolytic activity is 1.6 with a temperature of 37 °C, and an E/S ratio of 10 U:1 μg. According to Chen et al. [22,23], the optimum pH used for pepsin hydrolysis was 2, while for Lo and Li-Chan [31], the optimum pH used was 5. This discrepancy is likely influenced by the unique physiochemical properties of each of the substrate proteins in different pH environments. Chen et al. [22] selected a hydrolysis temperature of 37 °C for 24 h while in the other study by the same authors [23], 39 °C was used with half the time from the prior study.
Alcalase is known to produce peptides with hydrophobic domains at the C-terminal with optimum conditions of hydrolysis as follows: 56 °C in temperature, pH 7, E/S ratio of 2% (v/w) for 6 h [36]. Wu and Ding [25], Rayaprolu et al. [26], Li et al. [27] used alcalase in producing bioactive peptides from soybean with different conditions. Both Wu & Ding [25] and Rayaprolu et al. [26] used 50–55 °C hydrolysis temperatures, which complies with the optimal hydrolyzing temperature for alcalase for producing other types of bioactive peptides [37]. Meanwhile, Li et al. [27] used reaction temperature of 37 °C. As for the hydrolysis time, it varies in all three studies with Wu & Ding [25] reporting 12 h, Rayaprolu et al. [26] 1 h and Li et al. [27] 0.25 h. All three used alkaline hydrolytic conditions as alcalase might have optimum pH up to 10 [37].
The conditions for hydrolyzing soybean proteins for producing ACEi peptides may be different from the conditions where the optimum catalytic activity of the enzyme appears using standard substrate (casein, albumin, etc.). A study conducted by Yasuda et al. [38] suggested that protease derived from Monascus prureus has an optimum temperature of 50 °C and a pH of 3.2 to achieve optimum activity. However, utilizing the same enzyme source, Kuba et al. [28] conducted the hydrolysis at 37 °C with a pH of 3.3 for producing ACEi peptides.
Gouda et al. [11] compared four different enzymes for the production of bioactive peptides derived from soybeans, namely trypsin, chymotrypsin, protease P, and ginger protease. Trypsin and chymotrypsin used in that study were bovine-sourced and in both cases, the hydrolysis conditions were 37 °C at a pH of 8.2 for 18 h. These enzymes are digestive enzymes found in the small intestine which has a pH of 8–9. Hydrolysis of β-Lactoglobulin with trypsin that was reported in another study [39], carried out at pH 7.8, 37 °C, and a hydrolysis time of 2.42 h. Meanwhile, Kimball et al. [40] hydrolysed soybean proteins using chymotrypsin with optimum conditions as follows: E/S ratio of 2/100 (w/w) at 37 °C using a reaction time of 20–30 min. Reaction with protease P has also been used with the same hydrolysis conditions as trypsin and chymotrypsin. Siala et al. [41] reported that for protease derived from Aspergillus niger, the optimum pH was 4.0, while the enzyme was highly active at a temperature range of 30–60 °C, with an optimum activity at 50 °C. Meanwhile, ginger sourced protease, a cysteine protease indicated by the presence of cysteine residual at the active site of the enzyme, is best utilized at a pH range of 6–8 with a temperature of 60 °C [42]. Protease D3 is another novel cysteine peptidase, purified from germinating soybean cotyledon. According to Miwa [43], protease D3 worked at an optimum temperature of 40 °C with a pH above 4. Kodera and Nio [29] confirmed the former statement by utilizing protease D3 at 37–40 °C, at a pH of 4.5.
The optimum pH, hydrolysis time, E/S ratio, and reacting temperature used to produce ACEi peptides from soybean-derived proteins vary depending on the enzyme utilized. However, the optimum pH and temperature are relatively consistent, at least for pepsin and alcalase. Pepsin is optimum at pHs 1.2–2.5 and temperatures of 37–39 °C, whereas for alcalase pHs 7–9 and temperatures of 50–60 °C are the used conditions for producing ACEi peptides from soybean proteins.
3.2. Hydrolytic Conditions of Velvet Bean-Based Bioactive Peptide Preparation
Hydrolysis of parent proteins from velvet bean adopts the combination of endopeptidase and exopeptidase enzymes. Chalé et al. [17] and Segura-Campos et al. [16] both conducted hydrolysis of velvet bean concentrate using a pepsin-pancreatin combination and an alcalase-flavourzyme combination, while Tuz and Campos [18] used a combination of pepsin–pancreatin. The hydrolytic conditions for pepsin and pancreatin hydrolysis were 37 °C for 15 min at pH 2 for pepsin and at pH 7 for pancreatin. This complies with the previous study by Wang et al. [35] that utilizes pepsin for degrading both pepsin susceptible and resistant proteins. Meanwhile, flavourzyme, a mixture of endo and exopeptidases found in Aspergillus oryzae, works actively in the temperature of 50–55 °C and pH 5–7. Chalé et al. [17] and Segura-Campos et al. [16] used flavourzyme in combination with alcalase to hydrolyze velvet bean-derived proteins with the same hydrolysis condition. The optimum conditions used were 50 °C, pH 7 for 15 min. These conditions match Nguyen et al. [44] optimum conditions, however, in hydrolysing soybean protein. The range of the ACE inhibition activities (half maximal inhibitory concentration/IC50) from velvet bean peptide fraction is 0.0009–10.2 μg/mL [17,18]. The best enzymatic treatments having ACE IC50 of 0.0009 μg/mL as follows: the sequential hydrolytic activity by the combination of pepsin–pancreatin, E/S ratio for each is 10%, the temperature and reaction time for each is 37 °C and 0.74 h, respectively, and pH for pepsin and pancreatin is 2 and 7, respectively. Unfortunately, there is no study reported to identify the structures of ACEi peptides from velvet bean proteins at the time this work is caried out. Therefore, the following discussion focuses on ACEi peptides derived from soybean proteins.
3.3. Bioactive Peptides Exhibiting ACEi Activity from Soybean Proteins
The soybean protein isolate consists of various proteins which are grouped into four main protein classes, namely 2S (albumin), 7S (β-conglycinin), 11S (glycinin), and 15S. These storage proteins are grouped based on the sedimentation coefficients when the protein solution is subjected to a centrifugal field [45]. Although there are other minor proteins in soybeans, such as hemagglutinins, trypsin inhibitor, and intrinsic enzymes, 7S and 11S proteins are the most abundant and account for 75% of total storage protein content [9,46].
For 11S, there are five identified protein subunits which are divided into two groups based on homology. Group 1 comprises of G1 (53.6 kDa), G2 (52.4 kDa), G3 (52.2 kDa), and Group 2 with G4 (61.2 kDa) and G5 (55.4 kDa) [47]. β-conglycinin is the major glycoprotein for 7S storage protein. It consists of three major subunits, namely α (ca. 67 kDa), α′ (ca. 71 kDa), and β (ca. 50 kDa), in which all three have different physiochemical properties. The 7S fraction of globulins also comprises two more proteins, in addition to β-conglycinin, namely γ-conglycinin and Bo-conglycinin. In total, these soybean storage proteins are considered as potent parent proteins for producing ACEi peptides. In Table 3, the identified ACEi peptides from published literature are presented. Additionally, these identified ACEi peptides are matched with amino acid sequences from the typical soybean parent proteins, such as glycinin, β-conglycinin, and 2S albumin, to elucidate the source of those identified ACEi peptides.
Table 3.
Identified ACEi peptides and their corresponding specific soybean proteins as sources.
Chen et al. [22] identified five peptides (i.e., IA, TLAGAG, PPL, ITLL, and VMALPG) exhibiting ACEi activity produced from soybean proteins. Four other ACEi peptides were isolated from β-conglycinin and glycinin, namely LAIPVNKP, LPHF, SPYP, and WL by Kuba et al. [28]. Moreover, Gouda et al. [11] isolated VLIVP from glycinin. Other ACEi peptides isolated from soybeans were YVVFK, PNNKPFQ, NWGPLV, and IPPGVPYWT [29]. A comparison of those identified ACEi peptides with the SWISS-PROT database of soybean-derived parent proteins shows that most of the identified ACEi peptides are found in soybean protein, G1-G2, G4, α, α′, β, and 2S albumin. Identified ACEi peptides that are consistent with the database are IA, NWGPLV, SPYP, WL, LPHF, LAIPVNKP, VLIVP, and PNNKPFQ (refer to Table 3).
The molecular weight (MW) of an ACEi peptide can determine its affinity with ACE as the binding site might be too narrow for large MW peptides. ACEi activities from the corresponding identified peptides in Table 3 are evaluated and VLIVP, a peptide with a MW of 540 Da, is found to have the highest ACEi activity with an IC50 of 1.69 µM (Table 4). On the contrary, the lowest ACEi activity belongs to SPYP with an IC50 value of 850 µM. Despite having a considerably low MW (i.e., 462 Da), SPYP has the lowest ACEi activity. A correlation between MWs of ACEi peptides and their inhibition values is shown in Figure 1. The coefficient determination is 0.036, which indicates a very weak correlation. This suggests that ACEi activity does not entirely depend on peptide’s MW. Especially when MW of peptide is lower than 1 kDa, the molecular structure of ACEi peptide remarkably dictates the interaction between an ACEi peptide and ACE’s active side. These structural factors, such as peptide hydrophobicity and the types of amino acid residues encrypted within peptide strand are reported to influence the inhibition activity on ACE [24,48,49].
Table 4.
Molecular weights and inhibition values of ACEi bioactive peptides.
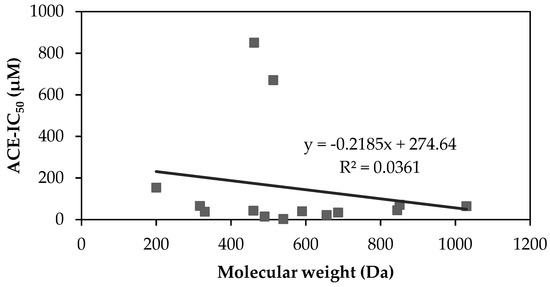
Figure 1.
Correlation between ACEi activity and peptide’s molecular weight. MWs and IC50 values are taken from Table 4.
Peptides with strong ACEi activities are mostly composed of hydrophobic amino acids with nonpolar side chain, positively charged, branched, and cyclic or aromatic residues, and proline at C terminus [24,49,50]. ACE contains HEXXH (i.e., histidine, glutamic acid, unknown, unknown, and histidine) as the active site, where two histidines (His383 and His387), together with the glutamate (Glu411), form zinc (Zn2+) binding ligands [51]. According to Jimsheena and Gowda [52], the presence of proline at the C-terminus in QRP and its short coordination distance, especially the carbonyl oxygen of the peptide bond between Q and R (3.2 Å) led to an increase in ACE inhibition. Bechaux et al. [53] and Aluko [49] also stated that the preferred ACEi peptide has an N-terminal branched-chain amino acid (aliphatic side chain with a branch), and of the C-terminal proline, aromatic, branched, or basic amino acids. Thus, peptides with N terminal of V or I, and C terminal of W, Y, P, and F are more preferred. The presence of proline in peptide sequences exhibiting ACEi activity is also reported by Sitanggang et al. [24] on fermented soybean (tempeh)-derived peptides (i.e., NEGDVLVIPPGVP, APIDVVVPPGNT, VAPTPNVPPYAG, FLVPPQ, FLVPPQE). It is known that the existence of proline and hydroxyproline in peptides is unaffected by the action of digestive proteases especially tripeptides with C-terminal proline-proline [54]. The resistance of bioactive peptides from the gastrointestinal proteases might be beneficial in maintaining the activity of ACEi peptides.
The fact that VLIVP has the highest ACEi activity (see Table 4) is consistent with the former reasoning statements [24,49,52,53]. VLIVP has a branched amino acid, namely valine at the N-terminal site and proline at C-terminal. Another factor that also contributes to the lowest IC50 value is the use of glycinin as a substrate for proteolysis [11]. According to Riblett et al. [55], glycinin contained proline as one of its major amino acids, which was a preferred amino acid in producing ACEi peptides, compared to β-conglycinin. Considering that VLIVP has the highest ACEi activity, the concluded best enzymatic treatments for producing ACEi peptides from soybean parent proteins as follows: proteolytic activity by protease P (Amano-P from Aspergillus sp.), the temperature of 37 °C, a reaction time of 18 h, pH 8.2 and E/S ratio of 2% [11].
4. Materials and Methods
The approach in constructing this review followed Carey et al. [56] which is a step-by-step guideline in conducting a systematic review. General views of the procedures consist of initial planning, conducting searches, data extraction, and quality analysis.
4.1. Defining a Research Question, Inclusion and Exclusion of Articles
This work carried out a literature review to search for relevant references. As a start, a review question was made. The review question chosen to conduct the research was “What are the best enzymatic treatments for producing potent bioactive peptides exhibiting ACE inhibitory activities from soybean and velvet bean proteins?” This scientific question was chosen to better clarify the purposes of this review, which were to demonstrate the optimum hydrolytic conditions that produce a high ACEi activity of a bioactive peptide and to evaluate the influence of structural factors of the identified bioactive peptide(s) on the ACEi activity. Based on that, the review question was categorized into a search tool, namely PEO, which was used to organize framework of main concepts [57]. PEO stands for population, exposure, and outcome. Within this study, the parent proteins from soybean and velvet bean, the enzymatic preparation conditions, and bioactive peptides exhibiting ACEi activity were considered as population, exposure, and outcome, respectively.
Furthermore, the studies or articles included were selected based on several criteria that were considered important for reference selection. Firstly, only studies published in English were included. There were no limitations regarding the publication dates. In addition, studies unrelated to the enzymatic preparations of velvet bean or soybean bioactive peptides exhibiting the ACEi effect were excluded.
4.2. Conducting and Reviewing the Search
References corresponding to this work were selected using “Preferred Reporting Items for Systematic reviews and Meta-Analysis” (PRISMA) guidelines (Figure 2 and Figure 3) as it is considered as the common guideline for conducting systematic reviews [58]. The following steps used in conducting this research were (1) Data collection (conducting searches of articles relevant to this work in database); (2) Data screening (selecting the articles based on criteria or classification determined beforehand); (3) Data integration (integrating selected references found and making the selection); (4) Data analysis (analysing the integrated data); (5) Data conclusion (providing results of the review).
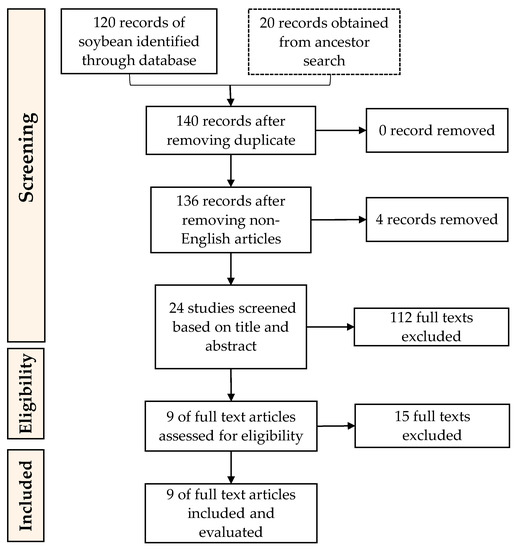
Figure 2.
PRISMA guidelines for the inclusion of articles on the enzymatic preparation of ACEi peptides from soybean proteins.
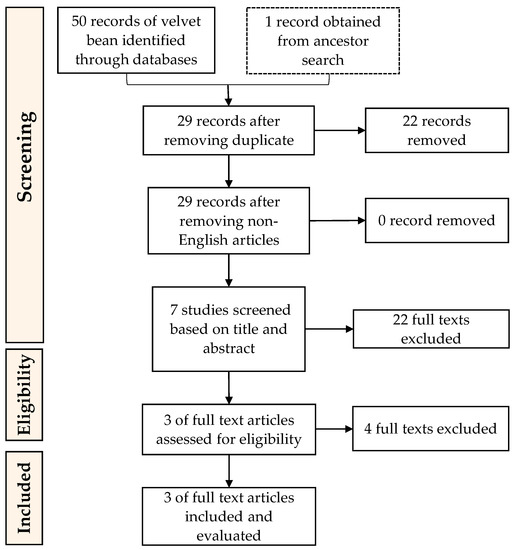
Figure 3.
PRISMA guidelines for the inclusion of articles on the enzymatic preparation of ACEi peptides from velvet bean proteins.
Studies relevant to the review were searched using databases such as Google Scholar (https://scholar.google.co.id; accessed on 30 May 2020), and Wiley (https://onlinelibrary.wiley.com; accessed on 30 May 2020). These databases were chosen as they are considered as common database used that support the Boolean search. Boolean search is a structured search where users can include several operations (AND, OR, NOT) to specify or broaden the search results. In searching data or references related to the ACEi peptides from soybeans, the Boolean used was “enzymatic preparation” AND “bioactive peptide” AND soybean AND (“blood pressure” OR hypertension). For velvet bean, the Boolean used was “bioactive peptide” AND (Mucuna OR “velvet bean”) AND (“blood pressure” OR hypertension).
Records of references related to the research topic of soybean ACEi peptides resulted in a total of 120 records from database searches and 20 records from ancient searches. Ancestor searches were also used to add more corresponding references to this study. Ancestor searches are conducted by going through the references from the selected articles to obtain more data sources [59]. Management and tracking of the records were by Mendeley (Mendeley Ltd., Elsevier, Amsterdam, The Netherlands), a reference manager that helps to collect references and organize citations. Duplicates were not found, although there were four non-English articles. References from both databases and ancestor searches were screened based on the inclusion and exclusion described above. After a further screening process, nine soybean records were used and evaluated in this research. The last screening process was to remove texts that are not specifically about soybean or not specifically discussed ACEi activity. As for velvet beans, a total of 50 records from database searches and one record from ancient searches were found. After merging duplicates and removing the non-English articles, 56 records were left to be screened based on the conformity of the title and abstract, and the research topic. The texts excluded here were studies that are not specifically about velvet bean or did not specifically discuss ACEi activity. This resulted in three velvet bean ACEi peptide-related articles to be included in this review.
5. Conclusions
The preparation of ACEi peptides from soybean in most literature is performed by the proteolytic activity of endopeptidases. The best enzymatic treatments for producing ACEi peptides from soybean parent proteins as follows: proteolytic activity by Protease P (Amano-P from Aspergillus sp.), a temperature of 37 °C, a reaction time of 18 h, pH 8.2 and an E/S ratio of 2%. The identified ACEi peptide having an IC50 of 1.69 µM is VLIVP. This peptide has a relatively low MW of 450 Da, which is presumably important to have it buried in the active site of ACE. Most importantly, this peptide has V and P at N- and C-terminal, respectively, which is a preferred configuration for enhancing ACEi activity. It is worth mentioning that besides MW of an ACEi peptide, other structural factors, such as peptide hydrophobicity and the types of amino acid residues encrypted within peptide strand, are also influential to enhance the ACEi activity.
As for velvet bean, the enzymes used for the hydrolytic actions are a combination of exo- and endo-peptidase. The best enzymatic treatments for producing peptide hydrolysates with a high ACEi activity are as follows: sequential hydrolytic activity by the combination of pepsin–pancreatic, an E/S ratio for each is 10%, the temperature and reaction time for each are 37 °C and 0.74 h, respectively, pH for pepsin is 2, whereas for pancreatin it is 7. Studies on the enzymatic hydrolysis of velvet bean proteins for producing ACEi peptides are limited. Additionally, there are no studies related to the identification of the molecular structures of ACEi peptides. Since velvet bean also has a high protein content, it is thus considered as a potent source of ACEi peptides. Therefore, the research interest should be directed in this area in the future.
Author Contributions
Conceptualization, A.B.S., J.E.P., N.S.P. and E.H.; methodology, A.B.S., J.E.P., N.S.P. and E.H.; formal analysis, A.B.S., J.E.P. and N.S.P.; investigation, A.B.S. and J.E.P.; data curation, J.E.P.; resources, A.B.S., E.S. and S.B.; writing—original draft preparation, A.B.S., J.E.P. and N.S.P.; writing—review and editing, A.B.S., N.S.P., E.H., E.S. and S.B.; supervision, A.B.S. and N.S.P.; funding acquisition, A.B.S., E.S. and S.B. All authors have read and agreed to the published version of the manuscript.
Funding
WCR Program 2325/IT3.L1/PN/2021.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing not applicable.
Acknowledgments
The authors acknowledge the support from the Ministry of Education, Culture, Research and Technology, Indonesia for supporting this work.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mills, K.T.; Bundy, J.D.; Kelly, T.N.; Reed, J.E.; Kearney, P.M.; Reynolds, K.; Chen, J.; He, J. Global disparities of hypertension prevalence and control. Circulation 2016, 134, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.S.; Vos, T.; Flaxman, A.D.; Danaei, G.; Shibuya, K.; Adair-Rohani, H.; Amann, M.; Anderson, H.R.; Andrews, K.G.; Aryee, M.; et al. A comparative risk assessment of burden of disease and injury attributable to 67 risk factors and risk factor clusters in 21 regions, 1990–2010: A systematic analysis for the global burden of disease study 2010. Lancet 2012, 380, 2224–2260. [Google Scholar] [CrossRef]
- Arnett, D.K.; Blumenthal, R.S.; Albert, M.A.; Buroker, A.B.; Goldberger, Z.D.; Hahn, E.J.; Himmelfarb, C.D.; Khera, A.; Lloyd-Jones, D.; McEvoy, J.W.; et al. 2019 ACC/AHA Guideline on the primary prevention of cardiovascular disease: A Report of the American College of Cardiology/American Heart Association Task Force on Clinical Practice Guidelines. Circulation 2019, 140, e596–e646. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, A.; Robertson, J.I. Captopril in the treatment of clinical hypertension and cardiac failure. Lancet 1979, 314, 836–839. [Google Scholar] [CrossRef]
- Sánchez, A.; Vázquez, A. Bioactive peptides: A review. Food Qual. Saf. 2017, 1, 29–46. [Google Scholar] [CrossRef]
- Sitanggang, A.B.; Sumitra, J.; Budijanto, S. Continuous production of tempe-based bioactive peptides using an automated enzymatic membrane reactor. Innov. Food Sci. Emerg. Technol. 2021, 68, 102639. [Google Scholar] [CrossRef]
- Chalamaiah, M.; Keskin Ulug, S.; Hong, H.; Wu, J. Regulatory requirements of bioactive peptides (protein hydrolysates) from food proteins. J. Funct. Foods 2019, 58, 123–129. [Google Scholar] [CrossRef]
- Chatterjee, C.; Gleddie, S.; Xiao, C.-W. Soybean bioactive peptides and their functional properties. Nutrients 2018, 10, 1211. [Google Scholar] [CrossRef]
- Syah, D.; Sitanggang, A.B.; Faradilla, R.F.; Trisna, V.; Karsono, Y.; Septianita, D.A. The influences of coagulation conditions and storage proteins on the textural properties of soy-curd (tofu). CYTA J. Food 2015, 13. [Google Scholar] [CrossRef]
- Balogun, I.O.; Olatidoye, O.P. Chemical composition and nutritional evaluation of velvet bean seeds (Mucuna utilis) for domestic consumption and industrial utilization in Nigeria. Pak. J. Nutr. 2012, 11, 116–122. [Google Scholar] [CrossRef]
- Mallikarjun Gouda, K.G.; Gowda, L.R.; Rao, A.G.A.; Prakash, V. Angiotensin I-converting enzyme inhibitory peptide derived from glycinin, the 11S globulin of soybean (Glycine max). J. Agric. Food Chem. 2006, 54, 4568–4573. [Google Scholar] [CrossRef]
- Rao, A.G.A.; Narasinga Rao, M.S. A method for isolation of 2S, 7S and 11S proteins of soybean. Prep. Biochem. 1977, 7, 89–101. [Google Scholar] [CrossRef]
- Deng, K.; Huang, Y.; Hua, Y. Isolation of glycinin (11S) from lipid-reduced soybean flour: Effect of processing conditions on yields and purity. Molecules 2012, 17, 2968–2979. [Google Scholar] [CrossRef]
- Deak, N.A.; Murphy, P.A.; Johnson, L.A. Effects of NaCl concentration on salting-in and dilution during salting-out on soy protein fractionation. J. Food Sci. 2006, 71, C247–C254. [Google Scholar] [CrossRef]
- Wu, S.; Murphy, P.A.; Johnson, L.A.; Fratzke, A.R.; Reuber, M.A. Pilot-plant fractionation of soybean glycinin and β-conglycinin. J. Am. Oil Chem. Soc. 1999, 76, 285–293. [Google Scholar] [CrossRef]
- Segura-Campos, M.; CP, E.-A.; Chel-Guerrero, L.; Betancur-Ancona, D. ACE-I inhibitory peptide fractions from enzymatic hydrolysates of velvet bean (Mucuna pruriens). Agric. Sci. 2013, 4, 767–773. [Google Scholar] [CrossRef][Green Version]
- Chalé, F.G.H.; Ruiz, J.C.R.; Fernández, J.J.A.; Ancona, D.A.B.; Campos, M.R.S. ACE inhibitory, hypotensive and antioxidant peptide fractions from Mucuna pruriens proteins. Proc. Biochem. 2014, 39, 1691–1698. [Google Scholar] [CrossRef]
- Tuz, M.A.O.; Campos, M.R.S. Purification of Mucuna pruriens (L) peptide fractions and evaluation of their ACE inhibitory effect. Biocatal. Agric. Biotechnol. 2017, 10, 390–395. [Google Scholar] [CrossRef]
- Abtahi, S.; Aminlari, M. Effect of sodium sulfite, sodium bisulfite, cysteine, and pH on protein solubility and sodium dodecyl sulfate−polyacrylamide gel electrophoresis of soybean milk base. J. Agric. Food Chem. 1997, 45, 4768–4772. [Google Scholar] [CrossRef]
- Mugendi, J.B.W.; Njagi, E.N.M.; Kuria, E.N.; Mwasaru, M.A.; Mureithi, J.G.; Apostolides, Z. Nutritional quality and physicochemical properties of Mucuna bean (Mucuna pruriens L.) protein isolates. Int. Food Res. J. 2010, 17, 357–366. [Google Scholar]
- Mótyán, J.; Tóth, F.; Tőzsér, J. Research applications of proteolytic enzymes in molecular biology. Biomolecules 2013, 3, 923–942. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-R.; Okada, T.; Muramoto, K.; Suetsuna, K.; Yang, S.-C. Identification of angiotensin I-converting enzyme inhibitory peptides derived from the peptic digest of soybean protein. J. Food Biochem. 2002, 26, 543–554. [Google Scholar] [CrossRef]
- Chen, J.R.; Yang, S.C.; Suetsuna, K.; Chao, J.C.J. Soybean protein-derived hydrolysate affects blood pressure in spontaneously hypertensive rats. J. Food Biochem. 2004, 28, 61–73. [Google Scholar] [CrossRef]
- Sitanggang, A.B.; Lesmana, M.; Budijanto, S. Membrane-based preparative methods and bioactivities mapping of tempe-based peptides. Food Chem. 2020, 329, 127193. [Google Scholar] [CrossRef]
- Wu, J.; Ding, X. Hypotensive and physiological effect of angiotensin converting enzyme inhibitory peptides derived from soy protein on spontaneously hypertensive rats. J. Agric. Food Chem. 2001, 49, 501–506. [Google Scholar] [CrossRef]
- Rayaprolu, S.; Hettiarachchy, N.; Horax, R.; Satchithanandam, E.; Chen, P.; Mauromoustakos, A. Amino acid profiles of 44 soybean lines and ACE-I inhibitory activities of peptide fractions from selected lines. J. Am. Oil Chem. Soc. 2015, 92, 1023–1033. [Google Scholar] [CrossRef]
- Li, M.; Xia, S.; Zhang, Y.; Li, X. Optimization of ACE inhibitory peptides from black soybean by microwave-assisted enzymatic method and study on its stability. LWT 2018, 98, 358–365. [Google Scholar] [CrossRef]
- Kuba, M.; Tana, C.; Tawata, S.; Yasuda, M. Production of angiotensin I-converting enzyme inhibitory peptides from soybean protein with Monascus purpureus acid proteinase. Process Biochem. 2005, 40, 2191–2196. [Google Scholar] [CrossRef]
- Kodera, T.; Nio, N. Identification of an angiotensin I-converting enzyme inhibitory peptides from protein hydrolysates by a soybean protease and the antihypertensive effects of hydrolysates in 4 spontaneously hypertensive model rats. J. Food Sci. 2006, 71, C164–C173. [Google Scholar] [CrossRef]
- Martínez-Medina, G.A.; Barragán, A.P.; Ruiz, H.A.; Ilyina, A.; Martínez Hernández, J.L.; Rodríguez-Jasso, R.M.; Hoyos-Concha, J.L.; Aguilar-González, C.N. Fungal proteases and production of bioactive peptides for the food industry. In Enzymes in Food Biotechnology; Elsevier: Amsterdam, The Netherlands, 2019; pp. 221–246. ISBN 9780128132807. [Google Scholar]
- Lo, W.M.Y.; Li-Chan, E.C.Y. Angiotensin I converting enzyme inhibitory peptides from in vitro pepsin–pancreatin digestion of soy protein. J. Agric. Food Chem. 2005, 53, 3369–3376. [Google Scholar] [CrossRef] [PubMed]
- Lafarga, T.; Álvarez, C.; Hayes, M. Bioactive peptides derived from bovine and porcine co-products: A review. J. Food Biochem. 2017, 41, e12418. [Google Scholar] [CrossRef]
- O’Meara, G.M.; Munro, P.A. Effects of reaction variables on the hydrolysis of lean beef tissue by alcalase. Meat Sci. 1984, 11, 227–238. [Google Scholar] [CrossRef]
- Antonov, V.K.; Rumsh, L.D.; Tikhodeeva, A.G. Kinetics of pepsin-catalysed transpeptidation: Evidence for the “amino-enzyme” intermediate. FEBS Lett. 1974, 46, 29–33. [Google Scholar] [CrossRef]
- Wang, R.; Edrington, T.C.; Storrs, S.B.; Crowley, K.S.; Ward, J.M.; Lee, T.C.; Liu, Z.L.; Li, B.; Glenn, K.C. Analyzing pepsin degradation assay conditions used for allergenicity assessments to ensure that pepsin susceptible and pepsin resistant dietary proteins are distinguishable. PLoS ONE 2017, 12, e0171926. [Google Scholar] [CrossRef]
- Awuor, O.L.; Edward Kirwa, M.; Betty, M.; Jackim, M.F. Optimization of alcalase hydrolysis conditions for production of dagaa (Rastrineobola argentea) protein hydrolysate with antioxidative properties. Ind. Chem. 2017, 3. [Google Scholar] [CrossRef]
- Tacias-Pascacio, V.G.; Morellon-Sterling, R.; Siar, E.-H.; Tavano, O.; Berenguer-Murcia, Á.; Fernandez-Lafuente, R. Use of Alcalase in the production of bioactive peptides: A review. Int. J. Biol. Macromol. 2020, 165, 2143–2196. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, M.; Shimabukuro, M.; Kikuchi, S. Production, purification and properties of acid proteinase from genus Monascus. Nippon. Shokuhin Kogyo Gakkaishi 1991, 38, 954–961. [Google Scholar] [CrossRef]
- Cheison, S.C.; Lai, M.-Y.; Leeb, E.; Kulozik, U. Hydrolysis of β-lactoglobulin by trypsin under acidic pH and analysis of the hydrolysates with MALDI–TOF–MS/MS. Food Chem. 2011, 125, 1241–1248. [Google Scholar] [CrossRef]
- Kimball, M.E.; Hsieh, D.S.T.; Rha, C. Chymotrypsin hydrolysis of soybean protein. J. Agric. Food Chem. 1981, 29, 872–874. [Google Scholar] [CrossRef]
- Siala, R.; Frikha, F.; Mhamdi, S.; Nasri, M.; Sellami Kamoun, A. Optimization of acid protease production by Aspergillus niger I1 on shrimp peptone using statistical experimental design. Sci. World J. 2012, 2012, 1–11. [Google Scholar] [CrossRef]
- Nafi’, A.; Foo, H.L.; Jamilah, B.; Ghazali, H.M. Properties of proteolytic enzyme from ginger (Zingiber officinale Roscoe). Int. Food Res. J. 2013, 20, 363–368. [Google Scholar]
- Miwa, T. Characterization of novel cysteine proteases from germinating cotyledons of soybean [Glycine max (L.) Merrill]. J. Biochem. 1999. [Google Scholar] [CrossRef]
- Hoa, N.T.; Huong, T.T.X.; Minh, N.P.; Dao, D.T. Investigation of enzymatic optimization by flavourzyme and celluclast for soy protein hydrolysate powder. Int. J. Adv. Pharm. Biol. Chem. 2014, 3, 563–574. [Google Scholar]
- Nishinari, K.; Fang, Y.; Guo, S.; Phillips, G.O. Soy proteins: A review on composition, aggregation and emulsification. Food Hydrocoll. 2014, 39, 301–318. [Google Scholar] [CrossRef]
- Sharma, S.; Kaur, M.; Goyal, R.; Gill, B.S. Physical characteristics and nutritional composition of some new soybean (Glycine max (L.) Merrill) genotypes. J. Food Sci. Technol. 2014, 51, 551–557. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, N.C. The structure and complexity of the 11S polypeptides in soybeans. J. Am. Oil Chem. Soc. 1985, 62, 1680–1686. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Yamauchi, T.; Katsuda, T.; Yamaji, H.; Katoh, S. Angiotensin-I converting enzyme (ACE) inhibitory mechanism of tripeptides containing aromatic residues. J. Biosci. Bioeng. 2008, 106, 310–312. [Google Scholar] [CrossRef] [PubMed]
- Aluko, R.E. Structure and function of plant protein-derived antihypertensive peptides. Curr. Opin. Food Sci. 2015, 4, 44–50. [Google Scholar] [CrossRef]
- Fan, H.; Liao, W.; Wu, J. Molecular interactions, bioavailability, and cellular mechanisms of angiotensin-converting enzyme inhibitory peptides. J. Food Biochem. 2019, 43, e12572. [Google Scholar] [CrossRef]
- Acharya, K.R.; Sturrock, E.D.; Riordan, J.F.; Ehlers, M.R.W. ACE revisited: A new target for structure-based drug design. Nat. Rev. Drug Discov. 2003, 2, 891–902. [Google Scholar] [CrossRef]
- Jimsheena, V.K.; Gowda, L.R. Angiotensin I-converting enzyme (ACE) inhibitory peptides derived from arachin by simulated gastric digestion. Food Chem. 2011, 125, 561–569. [Google Scholar] [CrossRef]
- Bechaux, J.; Gatellier, P.; Le Page, J.-F.; Drillet, Y.; Sante-Lhoutellier, V. A comprehensive review of bioactive peptides obtained from animal byproducts and their applications. Food Funct. 2019, 10, 6244–6266. [Google Scholar] [CrossRef]
- Jao, C.L.; Huang, S.L.; Hsu, K.C. Angiotensin I-converting enzyme inhibitory peptides: Inhibition mode, bioavailability, and antihypertensive effects. BioMedicine 2012, 2, 130–136. [Google Scholar] [CrossRef]
- Riblett, A.L.; Herald, T.J.; Schmidt, K.A.; Tilley, K.A. Characterization of β-conglycinin and glycinin soy protein fractions from four selected soybean genotypes. J. Agric. Food Chem. 2001, 49, 4983–4989. [Google Scholar] [CrossRef]
- Carey, J.; Schenker, M.; Yannascoli, S.; Ahn, J.; Baldwin, K. How to Write a Systematic Review: A Step-by-Step Guide. Univ. Pa. Orthop. J. 2013, 23, 64–69. [Google Scholar]
- Lekoane, B.K.M.; Mashamba-Thompson, T.P.; Ginindza, T.G. Mapping evidence on the distribution of human papillomavirus-related cancers in sub-Saharan Africa: Scoping review protocol. Syst. Rev. 2017, 6, 229. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G. Preferred Reporting Items for Systematic Reviews and Meta-Analyses: The PRISMA Statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef]
- Ahmed, T.; Javed, S.; Tariq, A.; Budzyńska, B.; D’Onofrio, G.; Daglia, M.; Fazel Nabavi, S.; Mohammad Nabavi, S. Daidzein and its effects on brain. Curr. Med. Chem. 2017, 24, 365–375. [Google Scholar]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).