
Enzymatic Preparation of Bioactive Peptides Exhibiting ACE Inhibitory Activity from Soybean and Velvet Bean: A Systematic Review

 ,
, 
Abstract
1. Introduction
2. Results
2.1. Substrate Preparation as Source of ACEi Peptides from Soybean and Velvet Bean
2.2. Hydrolytic Conditions for Producing ACEi Peptides from Soybean and Velvet Bean Protein Substrates
3. Discussion
3.1. Hydrolytic Conditions of Soybean-Based Bioactive Peptides Preparation
3.2. Hydrolytic Conditions of Velvet Bean-Based Bioactive Peptide Preparation
3.3. Bioactive Peptides Exhibiting ACEi Activity from Soybean Proteins
4. Materials and Methods
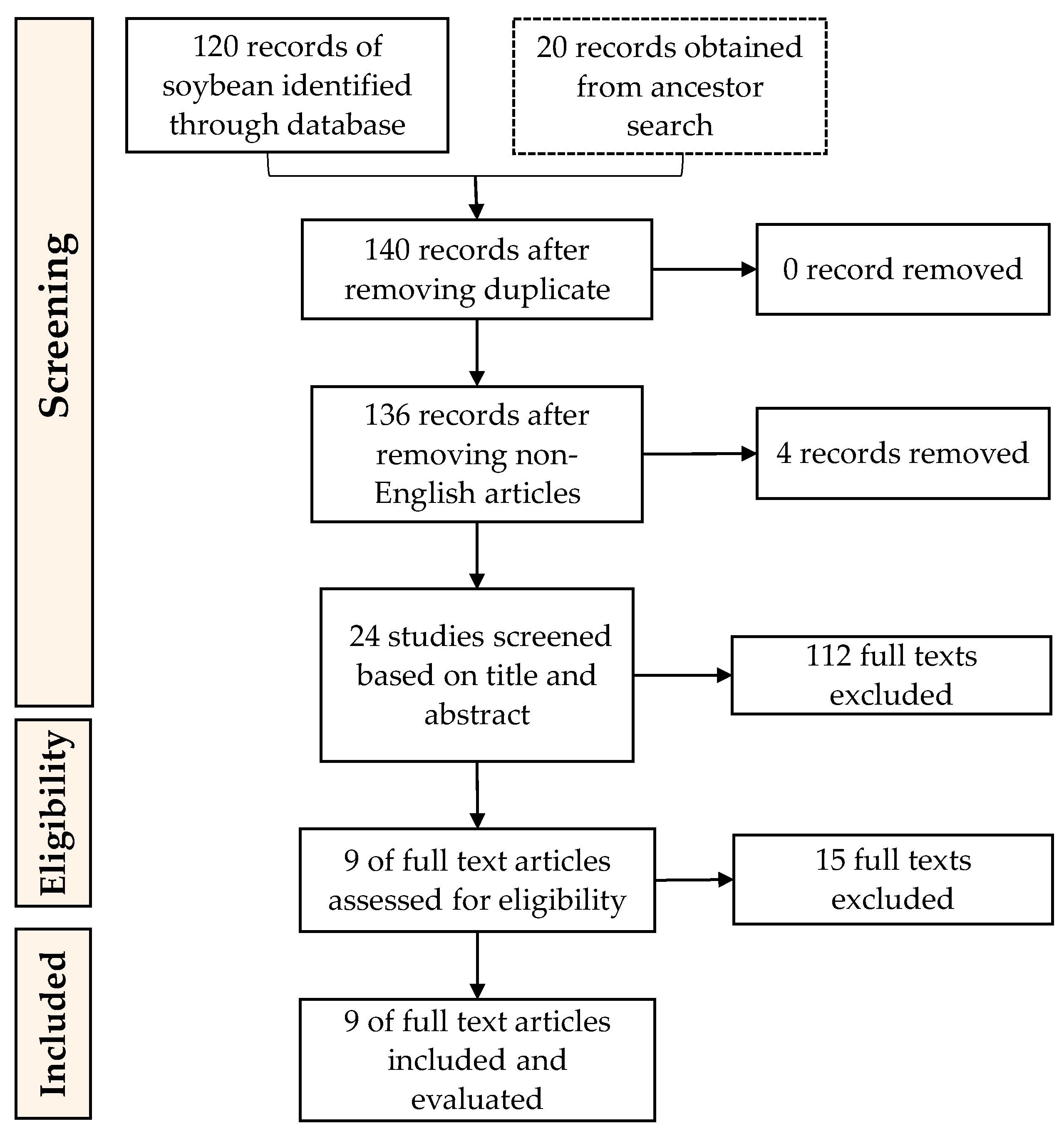
4.1. Defining a Research Question, Inclusion and Exclusion of Articles
4.2. Conducting and Reviewing the Search
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mills, K.T.; Bundy, J.D.; Kelly, T.N.; Reed, J.E.; Kearney, P.M.; Reynolds, K.; Chen, J.; He, J. Global disparities of hypertension prevalence and control. Circulation 2016, 134, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.S.; Vos, T.; Flaxman, A.D.; Danaei, G.; Shibuya, K.; Adair-Rohani, H.; Amann, M.; Anderson, H.R.; Andrews, K.G.; Aryee, M.; et al. A comparative risk assessment of burden of disease and injury attributable to 67 risk factors and risk factor clusters in 21 regions, 1990–2010: A systematic analysis for the global burden of disease study 2010. Lancet 2012, 380, 2224–2260. [Google Scholar] [CrossRef]
- Arnett, D.K.; Blumenthal, R.S.; Albert, M.A.; Buroker, A.B.; Goldberger, Z.D.; Hahn, E.J.; Himmelfarb, C.D.; Khera, A.; Lloyd-Jones, D.; McEvoy, J.W.; et al. 2019 ACC/AHA Guideline on the primary prevention of cardiovascular disease: A Report of the American College of Cardiology/American Heart Association Task Force on Clinical Practice Guidelines. Circulation 2019, 140, e596–e646. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, A.; Robertson, J.I. Captopril in the treatment of clinical hypertension and cardiac failure. Lancet 1979, 314, 836–839. [Google Scholar] [CrossRef]
- Sánchez, A.; Vázquez, A. Bioactive peptides: A review. Food Qual. Saf. 2017, 1, 29–46. [Google Scholar] [CrossRef]
- Sitanggang, A.B.; Sumitra, J.; Budijanto, S. Continuous production of tempe-based bioactive peptides using an automated enzymatic membrane reactor. Innov. Food Sci. Emerg. Technol. 2021, 68, 102639. [Google Scholar] [CrossRef]
- Chalamaiah, M.; Keskin Ulug, S.; Hong, H.; Wu, J. Regulatory requirements of bioactive peptides (protein hydrolysates) from food proteins. J. Funct. Foods 2019, 58, 123–129. [Google Scholar] [CrossRef]
- Chatterjee, C.; Gleddie, S.; Xiao, C.-W. Soybean bioactive peptides and their functional properties. Nutrients 2018, 10, 1211. [Google Scholar] [CrossRef]
- Syah, D.; Sitanggang, A.B.; Faradilla, R.F.; Trisna, V.; Karsono, Y.; Septianita, D.A. The influences of coagulation conditions and storage proteins on the textural properties of soy-curd (tofu). CYTA J. Food 2015, 13. [Google Scholar] [CrossRef]
- Balogun, I.O.; Olatidoye, O.P. Chemical composition and nutritional evaluation of velvet bean seeds (Mucuna utilis) for domestic consumption and industrial utilization in Nigeria. Pak. J. Nutr. 2012, 11, 116–122. [Google Scholar] [CrossRef]
- Mallikarjun Gouda, K.G.; Gowda, L.R.; Rao, A.G.A.; Prakash, V. Angiotensin I-converting enzyme inhibitory peptide derived from glycinin, the 11S globulin of soybean (Glycine max). J. Agric. Food Chem. 2006, 54, 4568–4573. [Google Scholar] [CrossRef]
- Rao, A.G.A.; Narasinga Rao, M.S. A method for isolation of 2S, 7S and 11S proteins of soybean. Prep. Biochem. 1977, 7, 89–101. [Google Scholar] [CrossRef]
- Deng, K.; Huang, Y.; Hua, Y. Isolation of glycinin (11S) from lipid-reduced soybean flour: Effect of processing conditions on yields and purity. Molecules 2012, 17, 2968–2979. [Google Scholar] [CrossRef]
- Deak, N.A.; Murphy, P.A.; Johnson, L.A. Effects of NaCl concentration on salting-in and dilution during salting-out on soy protein fractionation. J. Food Sci. 2006, 71, C247–C254. [Google Scholar] [CrossRef]
- Wu, S.; Murphy, P.A.; Johnson, L.A.; Fratzke, A.R.; Reuber, M.A. Pilot-plant fractionation of soybean glycinin and β-conglycinin. J. Am. Oil Chem. Soc. 1999, 76, 285–293. [Google Scholar] [CrossRef]
- Segura-Campos, M.; CP, E.-A.; Chel-Guerrero, L.; Betancur-Ancona, D. ACE-I inhibitory peptide fractions from enzymatic hydrolysates of velvet bean (Mucuna pruriens). Agric. Sci. 2013, 4, 767–773. [Google Scholar] [CrossRef][Green Version]
- Chalé, F.G.H.; Ruiz, J.C.R.; Fernández, J.J.A.; Ancona, D.A.B.; Campos, M.R.S. ACE inhibitory, hypotensive and antioxidant peptide fractions from Mucuna pruriens proteins. Proc. Biochem. 2014, 39, 1691–1698. [Google Scholar] [CrossRef]
- Tuz, M.A.O.; Campos, M.R.S. Purification of Mucuna pruriens (L) peptide fractions and evaluation of their ACE inhibitory effect. Biocatal. Agric. Biotechnol. 2017, 10, 390–395. [Google Scholar] [CrossRef]
- Abtahi, S.; Aminlari, M. Effect of sodium sulfite, sodium bisulfite, cysteine, and pH on protein solubility and sodium dodecyl sulfate−polyacrylamide gel electrophoresis of soybean milk base. J. Agric. Food Chem. 1997, 45, 4768–4772. [Google Scholar] [CrossRef]
- Mugendi, J.B.W.; Njagi, E.N.M.; Kuria, E.N.; Mwasaru, M.A.; Mureithi, J.G.; Apostolides, Z. Nutritional quality and physicochemical properties of Mucuna bean (Mucuna pruriens L.) protein isolates. Int. Food Res. J. 2010, 17, 357–366. [Google Scholar]
- Mótyán, J.; Tóth, F.; Tőzsér, J. Research applications of proteolytic enzymes in molecular biology. Biomolecules 2013, 3, 923–942. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-R.; Okada, T.; Muramoto, K.; Suetsuna, K.; Yang, S.-C. Identification of angiotensin I-converting enzyme inhibitory peptides derived from the peptic digest of soybean protein. J. Food Biochem. 2002, 26, 543–554. [Google Scholar] [CrossRef]
- Chen, J.R.; Yang, S.C.; Suetsuna, K.; Chao, J.C.J. Soybean protein-derived hydrolysate affects blood pressure in spontaneously hypertensive rats. J. Food Biochem. 2004, 28, 61–73. [Google Scholar] [CrossRef]
- Sitanggang, A.B.; Lesmana, M.; Budijanto, S. Membrane-based preparative methods and bioactivities mapping of tempe-based peptides. Food Chem. 2020, 329, 127193. [Google Scholar] [CrossRef]
- Wu, J.; Ding, X. Hypotensive and physiological effect of angiotensin converting enzyme inhibitory peptides derived from soy protein on spontaneously hypertensive rats. J. Agric. Food Chem. 2001, 49, 501–506. [Google Scholar] [CrossRef]
- Rayaprolu, S.; Hettiarachchy, N.; Horax, R.; Satchithanandam, E.; Chen, P.; Mauromoustakos, A. Amino acid profiles of 44 soybean lines and ACE-I inhibitory activities of peptide fractions from selected lines. J. Am. Oil Chem. Soc. 2015, 92, 1023–1033. [Google Scholar] [CrossRef]
- Li, M.; Xia, S.; Zhang, Y.; Li, X. Optimization of ACE inhibitory peptides from black soybean by microwave-assisted enzymatic method and study on its stability. LWT 2018, 98, 358–365. [Google Scholar] [CrossRef]
- Kuba, M.; Tana, C.; Tawata, S.; Yasuda, M. Production of angiotensin I-converting enzyme inhibitory peptides from soybean protein with Monascus purpureus acid proteinase. Process Biochem. 2005, 40, 2191–2196. [Google Scholar] [CrossRef]
- Kodera, T.; Nio, N. Identification of an angiotensin I-converting enzyme inhibitory peptides from protein hydrolysates by a soybean protease and the antihypertensive effects of hydrolysates in 4 spontaneously hypertensive model rats. J. Food Sci. 2006, 71, C164–C173. [Google Scholar] [CrossRef]
- Martínez-Medina, G.A.; Barragán, A.P.; Ruiz, H.A.; Ilyina, A.; Martínez Hernández, J.L.; Rodríguez-Jasso, R.M.; Hoyos-Concha, J.L.; Aguilar-González, C.N. Fungal proteases and production of bioactive peptides for the food industry. In Enzymes in Food Biotechnology; Elsevier: Amsterdam, The Netherlands, 2019; pp. 221–246. ISBN 9780128132807. [Google Scholar]
- Lo, W.M.Y.; Li-Chan, E.C.Y. Angiotensin I converting enzyme inhibitory peptides from in vitro pepsin–pancreatin digestion of soy protein. J. Agric. Food Chem. 2005, 53, 3369–3376. [Google Scholar] [CrossRef] [PubMed]
- Lafarga, T.; Álvarez, C.; Hayes, M. Bioactive peptides derived from bovine and porcine co-products: A review. J. Food Biochem. 2017, 41, e12418. [Google Scholar] [CrossRef]
- O’Meara, G.M.; Munro, P.A. Effects of reaction variables on the hydrolysis of lean beef tissue by alcalase. Meat Sci. 1984, 11, 227–238. [Google Scholar] [CrossRef]
- Antonov, V.K.; Rumsh, L.D.; Tikhodeeva, A.G. Kinetics of pepsin-catalysed transpeptidation: Evidence for the “amino-enzyme” intermediate. FEBS Lett. 1974, 46, 29–33. [Google Scholar] [CrossRef]
- Wang, R.; Edrington, T.C.; Storrs, S.B.; Crowley, K.S.; Ward, J.M.; Lee, T.C.; Liu, Z.L.; Li, B.; Glenn, K.C. Analyzing pepsin degradation assay conditions used for allergenicity assessments to ensure that pepsin susceptible and pepsin resistant dietary proteins are distinguishable. PLoS ONE 2017, 12, e0171926. [Google Scholar] [CrossRef]
- Awuor, O.L.; Edward Kirwa, M.; Betty, M.; Jackim, M.F. Optimization of alcalase hydrolysis conditions for production of dagaa (Rastrineobola argentea) protein hydrolysate with antioxidative properties. Ind. Chem. 2017, 3. [Google Scholar] [CrossRef]
- Tacias-Pascacio, V.G.; Morellon-Sterling, R.; Siar, E.-H.; Tavano, O.; Berenguer-Murcia, Á.; Fernandez-Lafuente, R. Use of Alcalase in the production of bioactive peptides: A review. Int. J. Biol. Macromol. 2020, 165, 2143–2196. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, M.; Shimabukuro, M.; Kikuchi, S. Production, purification and properties of acid proteinase from genus Monascus. Nippon. Shokuhin Kogyo Gakkaishi 1991, 38, 954–961. [Google Scholar] [CrossRef]
- Cheison, S.C.; Lai, M.-Y.; Leeb, E.; Kulozik, U. Hydrolysis of β-lactoglobulin by trypsin under acidic pH and analysis of the hydrolysates with MALDI–TOF–MS/MS. Food Chem. 2011, 125, 1241–1248. [Google Scholar] [CrossRef]
- Kimball, M.E.; Hsieh, D.S.T.; Rha, C. Chymotrypsin hydrolysis of soybean protein. J. Agric. Food Chem. 1981, 29, 872–874. [Google Scholar] [CrossRef]
- Siala, R.; Frikha, F.; Mhamdi, S.; Nasri, M.; Sellami Kamoun, A. Optimization of acid protease production by Aspergillus niger I1 on shrimp peptone using statistical experimental design. Sci. World J. 2012, 2012, 1–11. [Google Scholar] [CrossRef]
- Nafi’, A.; Foo, H.L.; Jamilah, B.; Ghazali, H.M. Properties of proteolytic enzyme from ginger (Zingiber officinale Roscoe). Int. Food Res. J. 2013, 20, 363–368. [Google Scholar]
- Miwa, T. Characterization of novel cysteine proteases from germinating cotyledons of soybean [Glycine max (L.) Merrill]. J. Biochem. 1999. [Google Scholar] [CrossRef]
- Hoa, N.T.; Huong, T.T.X.; Minh, N.P.; Dao, D.T. Investigation of enzymatic optimization by flavourzyme and celluclast for soy protein hydrolysate powder. Int. J. Adv. Pharm. Biol. Chem. 2014, 3, 563–574. [Google Scholar]
- Nishinari, K.; Fang, Y.; Guo, S.; Phillips, G.O. Soy proteins: A review on composition, aggregation and emulsification. Food Hydrocoll. 2014, 39, 301–318. [Google Scholar] [CrossRef]
- Sharma, S.; Kaur, M.; Goyal, R.; Gill, B.S. Physical characteristics and nutritional composition of some new soybean (Glycine max (L.) Merrill) genotypes. J. Food Sci. Technol. 2014, 51, 551–557. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, N.C. The structure and complexity of the 11S polypeptides in soybeans. J. Am. Oil Chem. Soc. 1985, 62, 1680–1686. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Yamauchi, T.; Katsuda, T.; Yamaji, H.; Katoh, S. Angiotensin-I converting enzyme (ACE) inhibitory mechanism of tripeptides containing aromatic residues. J. Biosci. Bioeng. 2008, 106, 310–312. [Google Scholar] [CrossRef] [PubMed]
- Aluko, R.E. Structure and function of plant protein-derived antihypertensive peptides. Curr. Opin. Food Sci. 2015, 4, 44–50. [Google Scholar] [CrossRef]
- Fan, H.; Liao, W.; Wu, J. Molecular interactions, bioavailability, and cellular mechanisms of angiotensin-converting enzyme inhibitory peptides. J. Food Biochem. 2019, 43, e12572. [Google Scholar] [CrossRef]
- Acharya, K.R.; Sturrock, E.D.; Riordan, J.F.; Ehlers, M.R.W. ACE revisited: A new target for structure-based drug design. Nat. Rev. Drug Discov. 2003, 2, 891–902. [Google Scholar] [CrossRef]
- Jimsheena, V.K.; Gowda, L.R. Angiotensin I-converting enzyme (ACE) inhibitory peptides derived from arachin by simulated gastric digestion. Food Chem. 2011, 125, 561–569. [Google Scholar] [CrossRef]
- Bechaux, J.; Gatellier, P.; Le Page, J.-F.; Drillet, Y.; Sante-Lhoutellier, V. A comprehensive review of bioactive peptides obtained from animal byproducts and their applications. Food Funct. 2019, 10, 6244–6266. [Google Scholar] [CrossRef]
- Jao, C.L.; Huang, S.L.; Hsu, K.C. Angiotensin I-converting enzyme inhibitory peptides: Inhibition mode, bioavailability, and antihypertensive effects. BioMedicine 2012, 2, 130–136. [Google Scholar] [CrossRef]
- Riblett, A.L.; Herald, T.J.; Schmidt, K.A.; Tilley, K.A. Characterization of β-conglycinin and glycinin soy protein fractions from four selected soybean genotypes. J. Agric. Food Chem. 2001, 49, 4983–4989. [Google Scholar] [CrossRef]
- Carey, J.; Schenker, M.; Yannascoli, S.; Ahn, J.; Baldwin, K. How to Write a Systematic Review: A Step-by-Step Guide. Univ. Pa. Orthop. J. 2013, 23, 64–69. [Google Scholar]
- Lekoane, B.K.M.; Mashamba-Thompson, T.P.; Ginindza, T.G. Mapping evidence on the distribution of human papillomavirus-related cancers in sub-Saharan Africa: Scoping review protocol. Syst. Rev. 2017, 6, 229. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G. Preferred Reporting Items for Systematic Reviews and Meta-Analyses: The PRISMA Statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef]
- Ahmed, T.; Javed, S.; Tariq, A.; Budzyńska, B.; D’Onofrio, G.; Daglia, M.; Fazel Nabavi, S.; Mohammad Nabavi, S. Daidzein and its effects on brain. Curr. Med. Chem. 2017, 24, 365–375. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Enzyme | Substrate | Temp. (°C) | Time (h) | pH | Enzyme-to-Substrate Ratio E/S | Peptide Sequence | Ref. |
|---|---|---|---|---|---|---|---|
| Pepsin | Protein concentrate | 37 | 24 | 2 | 6% | IA | Chen et al. [22] |
| TLAGAG | |||||||
| PPL | |||||||
| ITLL | |||||||
| VMALPG | |||||||
| Pepsin | Protein isolate | 39 | 12 | 2 | 3% | - | Chen et al. [23] |
| Acid-precipitated protein | |||||||
| Alcalase | Protein concentrate | 50 | 12 | 9 | 4% | - | Wu & Ding [25] |
| M. purpureus acid proteinase | β-conglycinin | 37 | 10 | 3.3 | - | LAIPVNKP | Kuba et al. [28] |
| LPHF | |||||||
| Glycinin | SPYP | ||||||
| WL | |||||||
| Bovine trypsin | Glycinin | 37 | 18 | 8.2 | 2% | VLIVP | Gouda et al. [11] |
| Bovine chymotrypsin | 37 | 18 | 8.2 | ||||
| Ginger protease | 50 | 16 | 6 | ||||
| Protease P (Amano-P from Aspergillus sp.) | 37 | 18 | 8.2 | ||||
| Protease D3 from E. coli strain JM109 | Protein isolate | 37–40 | 24–48 | 4.5 | 0.2% | YVVFK | Kodera & Nio [29] |
| PNNKPFQ | |||||||
| NWGPLV | |||||||
| IPPGVPYWT | |||||||
| Pepsin | Protein isolate | 37 | 1 | 5.3 | 4% | - | Lo & Li-Chan [31] |
| Pancreatin | 37 | 2 | 7.5 | ||||
| Alcalase | Protein isolate | 55 | 1 | 8 | - | - | Rayaprolu et al. [26] |
| Alcalase | Protein isolate | 30 | 0.25 | 9 | 6% | - | Li et al. [27] |
| Enzyme | Enzyme Type | Hydrolysis Conditions | Ref. | |||
|---|---|---|---|---|---|---|
| Temp. (°C) | Time (h) | pH | Enzyme-to-Substrate Ratio E/S | |||
| Pepsin | Endopeptidase | 37 | 0.75 | 2 | 10% | Herrera-Chale et al. [17] |
| Pancreatin | Exopeptidase | 37 | 0.75 | 7.5 | ||
| Alcalase | Endopeptidase | 50 | 0.75 | 8 | ||
| Flavourzyme | Exopeptidase | 50 | 0.75 | 7 | ||
| Pepsin | Endopeptidase | 37 | 0.75 | 2 | 10% | Tuz & Campos [18] |
| Pancreatin | Exopeptidase | 37 | 0.75 | 7 | ||
| Pepsin | Endopeptidase | 37 | 0.75 | 2 | 10% | Segura-Campos et al. [16] |
| Pancreatin | Exopeptidase | 37 | 0.75 | 7 | ||
| Alcalase | Endopeptidase | 50 | 0.75 | 8 | ||
| Flavourzyme | Exopeptidase | 50 | 0.75 | 7 | ||
| Soybean Protein | Total Molecular Weight/MW (Da) | UniProt Entry | Peptide Strand | Location of Peptide Strand from N-Terminal | Identified ACEi Peptide from Literature | |||
|---|---|---|---|---|---|---|---|---|
| AA Residues | MW (Da) | Ref. | ||||||
| Glycinin | G1 | 55,706 | P04776 | LIAVPTGVAW | 141–150 | IA | 202 | Chen et al. [22] |
| ALSWLRLSAE | 351–360 | WL | 317 | Kuba et al. [28] | ||||
| VLIVPQNFVV | 411–420 | VLIVP | 540 | Gouda et al. [11] | ||||
| G2 | 54,391 | P04405 | ALWLLKLSAQ | 341–350 | WL | 317 | Kuba et al. [28] | |
| TWNPNNKPFQ | 51–60 | PNNKPFQ | 844 | Kodera & Nio [29] | ||||
| G3 | - | |||||||
| G4 | 63,797 | P02858 | HLPSYSPYPR | 81–90 | SPYP | 462 | Kuba et al. [28] | |
| MIIIAQGKGA | 91–100 | IA | 202 | Chen et al. [22] | ||||
| SFNTNEDIAE | 241–250 | IA | 202 | Chen et al. [22] | ||||
| ENIARPSRAD | 391–400 | IA | 202 | Chen et al. [22] | ||||
| YEGNWGPLVN | 541–550 | NWGPLV | 586 | Kodera & Nio [29] | ||||
| G5 | 57,956 | P04347 | GLEYVVFKTH | 461–470 | YVVFK | 655 | Kodera & Nio [29] | |
| β–conglycinin | α | 70,306 | P0DO16 | VSFGIAYWEK | 21–30 | IA | 202 | Chen et al. [22] |
| NENLRLITLA IPVNKPGRFE | 301–320 | LAIPVNKP | 851 | Kuba et al. [28] | ||||
| LLPHFNSKA | 451–460 | LPHF | 513 | Kuba et al. [28] | ||||
| α’ | 72,228 | P11827 | VSFGIAYWEK | 21–30 | IA | 202 | Chen et al. [22] | |
| RMITLAIPVN KPGRFESFFL | 321–340 | LAIPVNKP | 851 | Kuba et al. [28] | ||||
| β | 50,476 | P25974 | QNLKIIKLAI PVNKPGRYDD | 141–160 | LAIPVNKP | 851 | Kuba et al. [28] | |
| EGALLLPHFN | 281–290 | LPHF | 513 | Kuba et al. [28] | ||||
| 2S albumin | 18,460 | P19594 | LLFCIAHTCS | 11–20 | IA | 202 | Chen et al. [22] | |
| No | Sequence | MW (Da) | ACEi IC50 (μM) | Ref. |
|---|---|---|---|---|
| 1 | IA | 200 | 153 | Chen et al. [22] |
| 2 | WL | 317 | 65 | Kuba et al. [28] |
| 3 | PPL | 330 | 37 | Chen et al. [22] |
| 4 | ITLL | 460 | 42 | |
| 5 | SPYP | 462 | 850 | Kuba et al. [28] |
| 6 | TLAGAG | 490 | 14 | Chen et al. [22] |
| 7 | LPHF | 513 | 670 | Kuba et al. [28] |
| 8 | VLIVP | 540 | 1.69 | Gouda et al. [11] |
| 9 | VMALPG | 590 | 39 | Chen et al. [22] |
| 10 | YVVFK | 655.62 | 21 | Kodera & Nio [29] |
| 11 | NWGPLV | 686.56 | 33 | |
| 12 | PNNKPFQ | 844.59 | 44 | |
| 13 | LAIPVNKP | 851 | 70 | Kuba et al. [28] |
| 14 | IPPGVPYWT | 1029.69 | 64 | Kodera & Nio [29] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sitanggang, A.B.; Putri, J.E.; Palupi, N.S.; Hatzakis, E.; Syamsir, E.; Budijanto, S. Enzymatic Preparation of Bioactive Peptides Exhibiting ACE Inhibitory Activity from Soybean and Velvet Bean: A Systematic Review. Molecules 2021, 26, 3822. https://doi.org/10.3390/molecules26133822
Sitanggang AB, Putri JE, Palupi NS, Hatzakis E, Syamsir E, Budijanto S. Enzymatic Preparation of Bioactive Peptides Exhibiting ACE Inhibitory Activity from Soybean and Velvet Bean: A Systematic Review. Molecules. 2021; 26(13):3822. https://doi.org/10.3390/molecules26133822
Chicago/Turabian StyleSitanggang, Azis Boing, Jessica Eka Putri, Nurheni Sri Palupi, Emmanuel Hatzakis, Elvira Syamsir, and Slamet Budijanto. 2021. "Enzymatic Preparation of Bioactive Peptides Exhibiting ACE Inhibitory Activity from Soybean and Velvet Bean: A Systematic Review" Molecules 26, no. 13: 3822. https://doi.org/10.3390/molecules26133822
APA StyleSitanggang, A. B., Putri, J. E., Palupi, N. S., Hatzakis, E., Syamsir, E., & Budijanto, S. (2021). Enzymatic Preparation of Bioactive Peptides Exhibiting ACE Inhibitory Activity from Soybean and Velvet Bean: A Systematic Review. Molecules, 26(13), 3822. https://doi.org/10.3390/molecules26133822