
Optimization of Laccase from Ganoderma lucidum Decolorizing Remazol Brilliant Blue R and Glac1 as Main Laccase-Contributing Gene



Abstract
1. Introduction
2. Results and Discussion
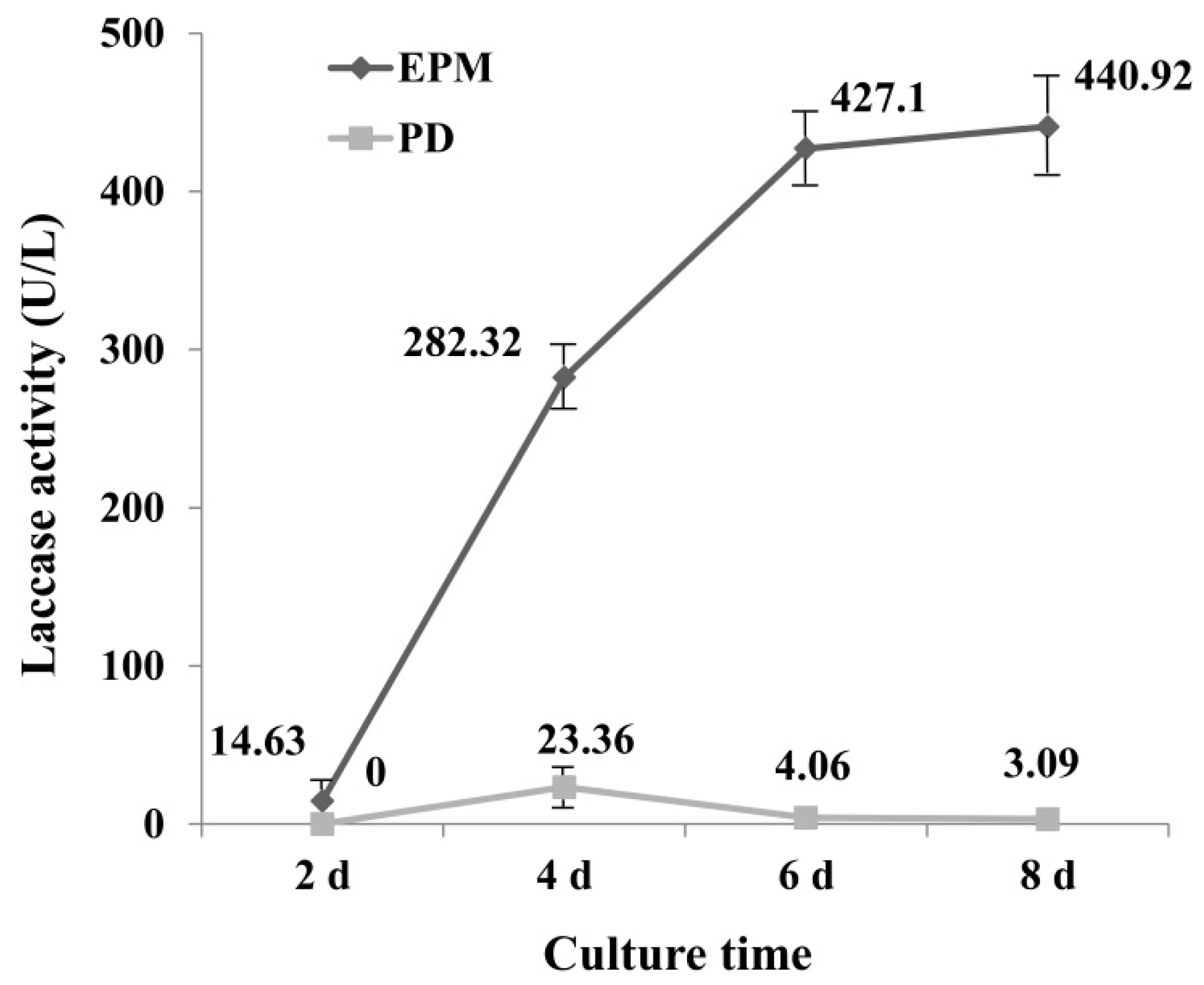
2.1. Laccase Activity in PD and EPM Medium
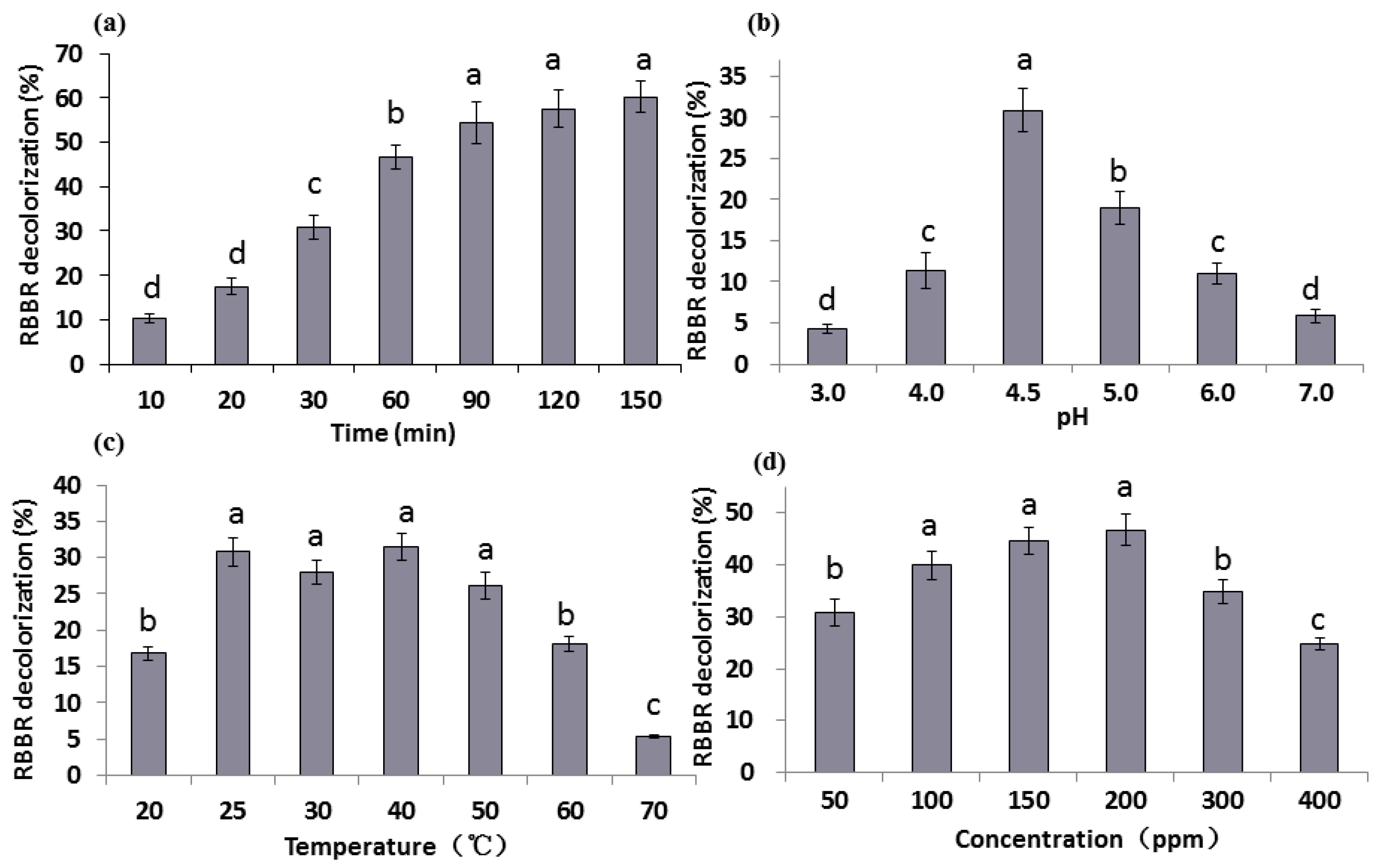
2.2. Effect of Reaction Time, pH, Temperature, and Dye Concentrations on RBBR Decolorization
2.2.1. The Effect of Reaction Time
2.2.2. The Effect of pH
2.2.3. The Effect of Temperature
2.2.4. The Effect of Dye Concentration
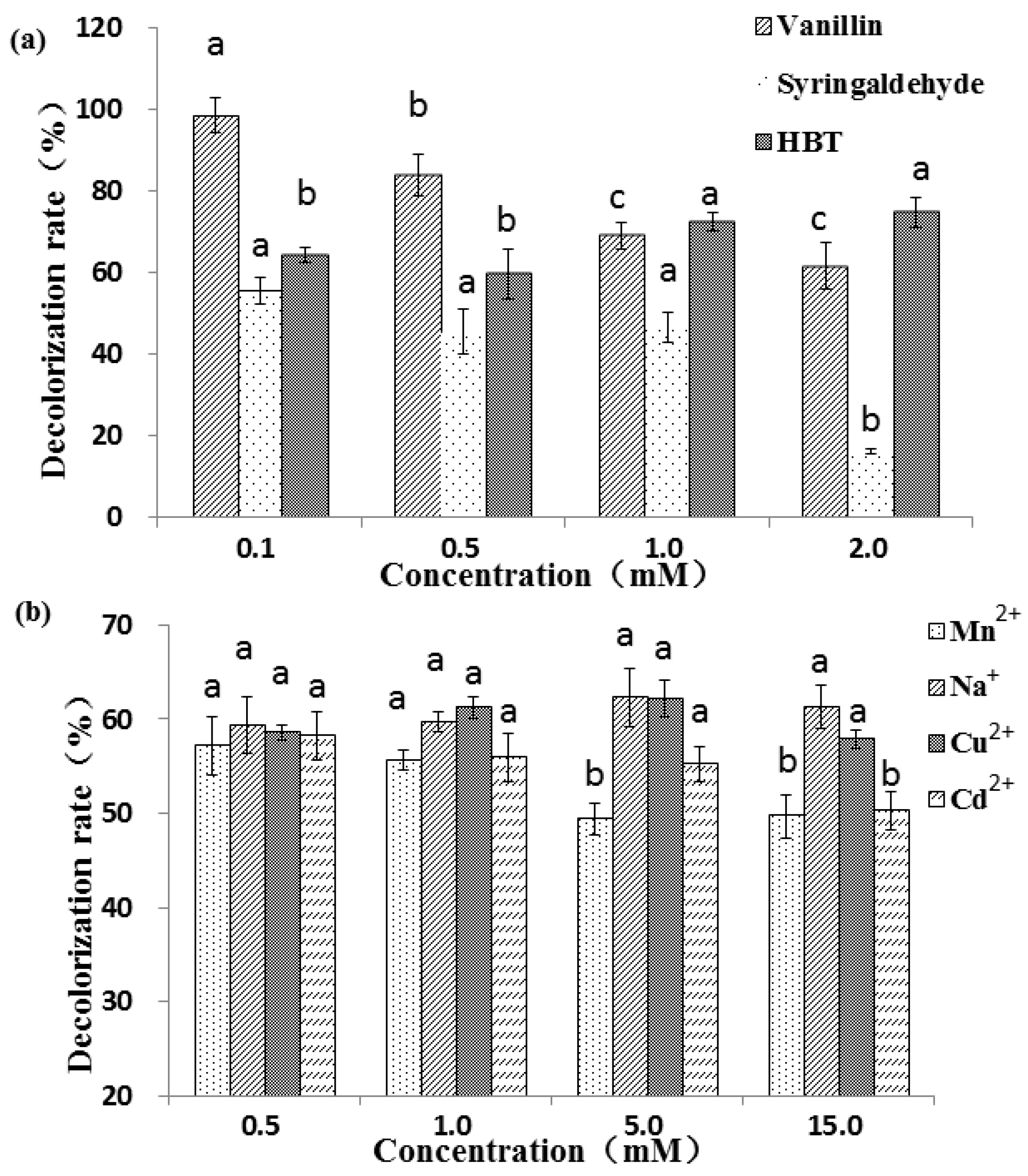
2.3. Effect of Mediators and Metal Ions on RBBR Decolorization
2.4. Laccase Isoenzyme Electrophoresis and Protein Mass Spectrometry Sequencing
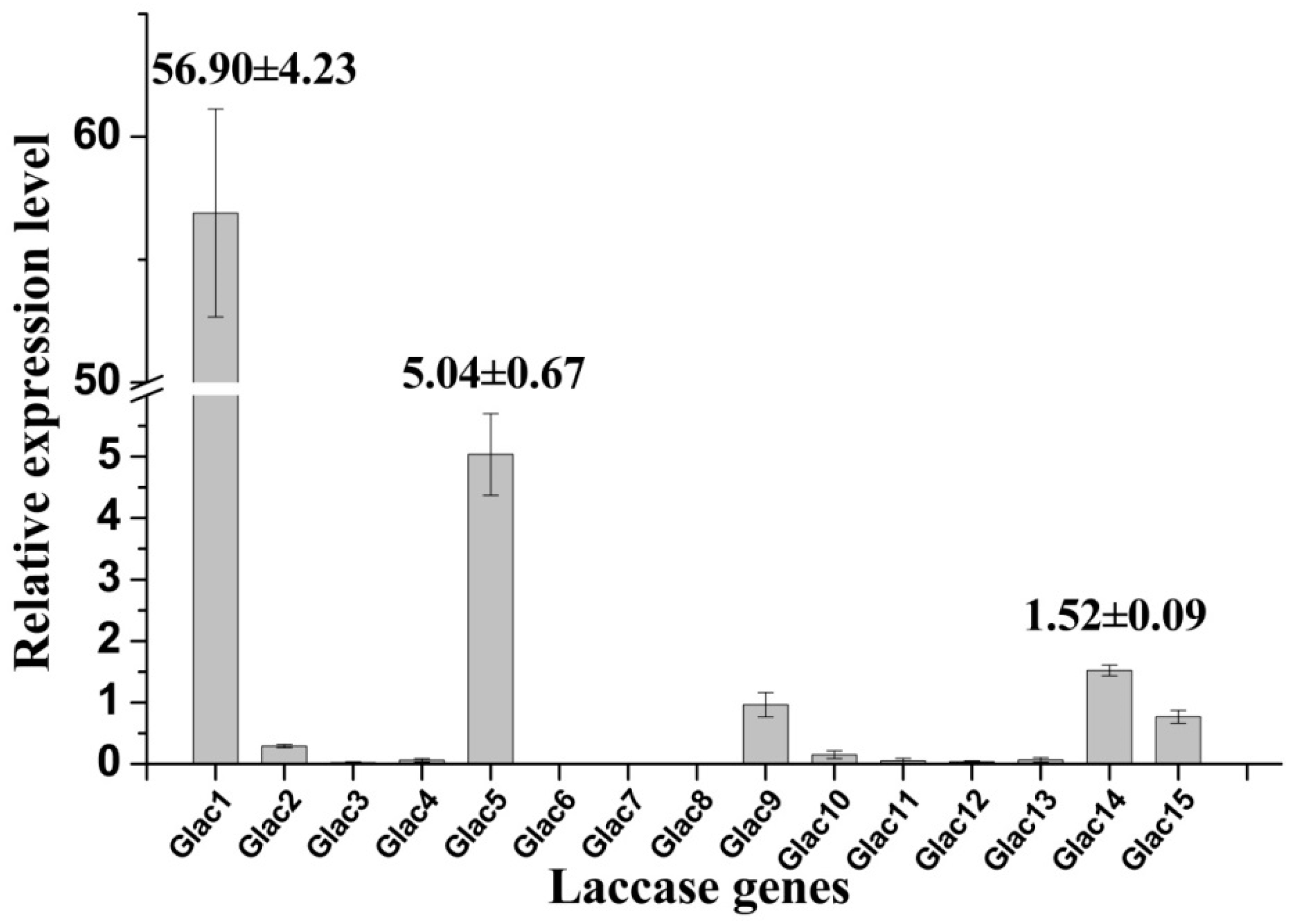
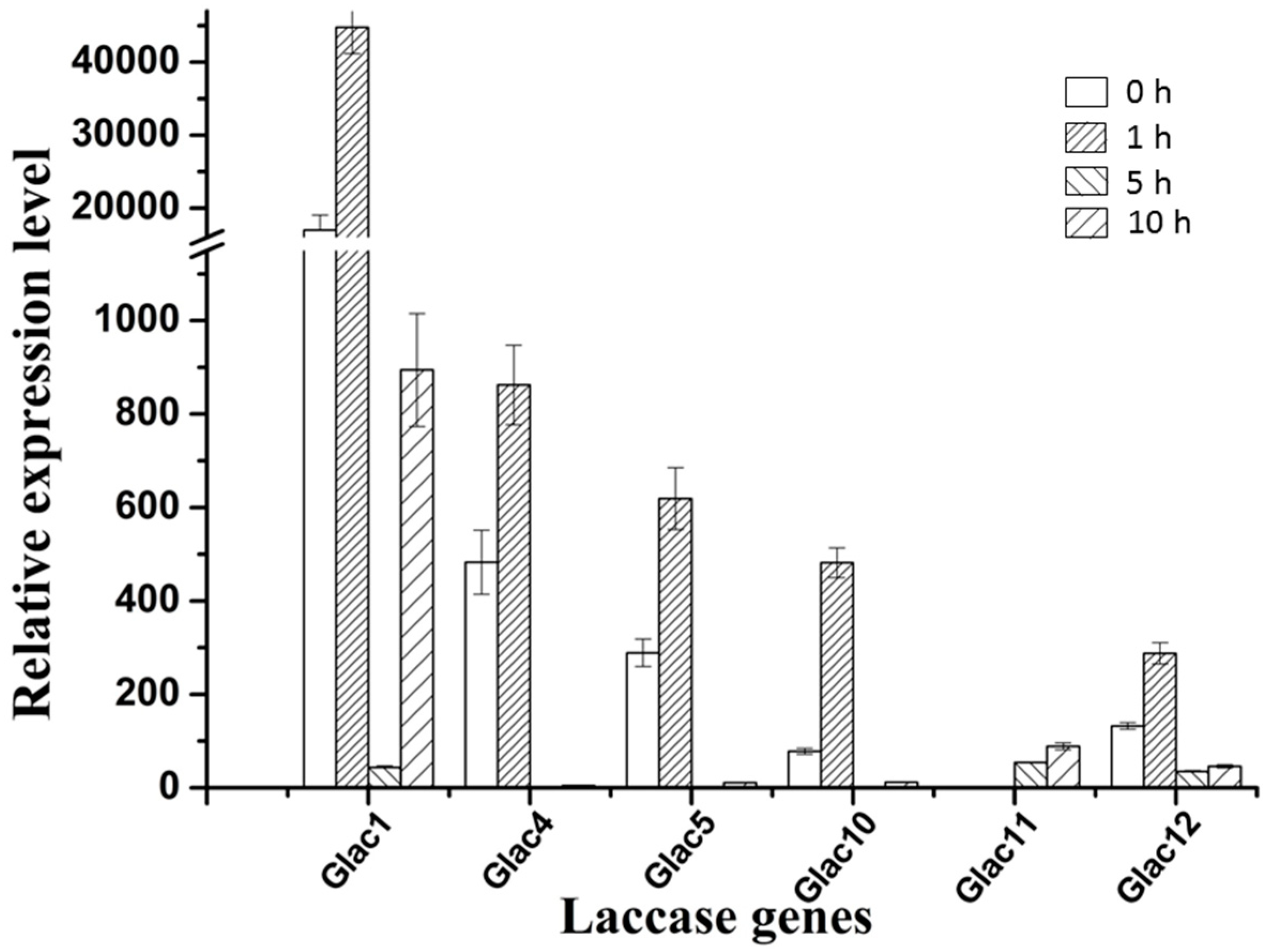
2.5. Transcriptional Analysis of Laccase Genes
3. Material and Methods
3.1. Microorganism and Culture Conditions
3.2. Enzyme Assays
3.3. Dye Decolorization
3.4. Laccase Isoenzyme Electrophoresis and Protein Mass Spectrometry Sequencing
3.5. RNA Extraction and Real-time PCR Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Palmieri, G.; Cennamo, G.; Sannia, G. Remazol brilliant blue r decolourisation by the fungus pleurotus ostreatus and its oxidative enzymatic system. Enzym. Microb. Technol. 2005, 36, 17–24. [Google Scholar] [CrossRef]
- Moreira, M.T.; Feijoo, G.; Lema, J.M. Evaluation of different fungal strains in the decolorisation of synthetic dyes. Biotechnol. Lett. 2000, 22, 1499–1503. [Google Scholar] [CrossRef]
- Li, L.; Dai, W.K.; Yu, P.; Zhao, J.; Qu, Y.B. Decolorisation of synthetic dyes by crude laccase from Rigidoporus lignosus W1. J. Chem. Technol. Biot. 2009, 84, 399–404. [Google Scholar] [CrossRef]
- Eichlerová, I.; Homolka, L.; Benada, O.; Kofroňová, O.; Hubálek, T.; Nerud, F. Decolorization of Orange G and Remazol Brilliant Blue R by the white rot fungus Dichomitus squalens: Toxicological evaluation and morphological study. Chemosphere 2007, 69, 795–802. [Google Scholar] [CrossRef]
- Chander, M.; Arora, D.S. Evaluation of some white-rot fungi for their potential to decolourise industrial dyes. Dye. Pigment. 2007, 72, 192–198. [Google Scholar] [CrossRef]
- Asgher, M.; Bhatti, H.N.; Ashraf, M.; Legge, R.L. Recent developments in biodegradation of industrial pollutants by white-rot fungi and their enzyme system. Biodegradation 2008, 19, 771–783. [Google Scholar] [CrossRef]
- Arora, D.S.; Sharma, R.K. Ligninolytic fungal laccases and their biotechnological applications. Appl. Biochem. Biotechnol. 2010, 160, 1760–1788. [Google Scholar] [CrossRef]
- Rajeeva, S.; Lele, S.S. Three-phase partitioning for concentration and purification of laccase produced by submerged cultures of Ganoderma sp. WR-1. Biochem. Eng. J. 2011, 54, 103–110. [Google Scholar] [CrossRef]
- Shraddha Shekher, R.; Sehgal, S.; Kamthania, M.; Kumar, A. Laccase: Microbial sources, production, purification, and potential biotechnological applications. Enzym. Res. 2011, 1–11. [Google Scholar] [CrossRef]
- Michniewicz, A.; Ledakowicz, S.; Ullrich, R.; Hofrichter, M. Kinetics of the enzymatic decolorization of textile dyes by laccase from Cerrena unicolor. Dye. Pigment. 2008, 77, 295–302. [Google Scholar] [CrossRef]
- Majeau, J.A.; Brar, S.K.; Tyagi, R.D. Laccases for removal of recalcitrant and emerging pollutants. Bioresour. Technol. 2010, 101, 2331–2350. [Google Scholar] [CrossRef] [PubMed]
- Kerovuo, J.S.; Haremza, S.; Koch, O.; Habicher, T.; Robertson, D.E.; Desantis, G.; McCann, R.; Luginbuhl, P. Laccases for Bio-Bleaching. U.S. Patent No. 20160237411, 28 April 2013. [Google Scholar]
- Fang, Q.H.; Zhong, J.J. Submerged fermentation of higher fungus Ganoderma lucidum for production of valuable bioactive metabolites-ganoderic acid and polysaccharide. Biochem. Eng. J. 2002, 10, 61–65. [Google Scholar] [CrossRef]
- Liu, J.; Shimizu, K.; Konishi, F.; Noda, K.; Kumamoto, S.; Kurashiki, K.; Kondo, R. Anti-androgenic activities of the triterpenoids fraction of Ganoderma lucidum. Food Chem. 2007, 100, 1691–1696. [Google Scholar] [CrossRef]
- Yao, Q.; Gong, Z.Y.; Gao, X.G.; Ren, P.F.; Ren, H.X.; Li, J.; Qu, L.; Wang, Z.X.; Liu, Y. Fermentation Process for Improving Yield of Lucid Ganoderma Laccase. China Patent No. 101928699, 20 April 2010. [Google Scholar]
- Murugesan, K.; Yang, I.H.; Kim, Y.M.; Jeon, J.R.; Chang, Y.S. Enhanced transformation of malachite green by laccase of Ganoderma lucidum in the presence of natural phenolic compounds. Appl. Microbiol. Biot. 2009, 82, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Jong-Rok, J.; Kumarasamy, M.; Young-Mo, K.; Eun-Ju, K.; Yoon-Seok, C. Synergistic effect of laccase mediators on pentachlorophenol removal by Ganoderma lucidum laccase. Appl. Microbiol. Biot. 2008, 81, 783–790. [Google Scholar]
- Coelho, J.S.; Souza, C.G.M.; Oliveira, A.L.; Bracht, A.; Costa, M.A.F.; Peralta, R.M. Comparative removal of bentazon by Ganoderma lucidum in liquid and solidstate cultures. Curr. Microbiol. 2010, 60, 350–355. [Google Scholar] [CrossRef]
- Murugesan, K.; Nam, I.H.; Kim, Y.M.; Chang, Y.S. Decolorisation of rective dyes by a thermostable laccase produced by Ganoderma lucidum in solid state culture. Enzym. Microb. Technol. 2007, 40, 1662–1672. [Google Scholar] [CrossRef]
- Songulashvili, G.; Elisashvilli, V.; Wasser, S.P.; Nevo, E.; Hadar, Y. Basidiomycetes laccase and manganese peroxidase activity in submerged fermentation of food industry wastes. Enzym. Microb. Technol. 2007, 41, 57–61. [Google Scholar] [CrossRef]
- Sitarz, A.; Mikkelsen, J.D.; Meyer, A.M.B.S.; Lezyk, M. A Novel lLaccase from Ganoderma Lucidum Capable of Enhancing Enzymatic Degradation of Lignocellulolytic Biomass. Europe Patent No. 2895599, 10 October 2013. [Google Scholar]
- Liao, C.S.; Yuan, S.Y.; Hung, B.H.; Chang, B.V. Removal of organic toxic chemicals using the spent mushroom compost of Ganoderma lucidum. J. Env. Monit. 2012, 14, 1983–1988. [Google Scholar] [CrossRef]
- Pundir, C.S.; Rawal, R.; Chawla, S.; Renuka; Kuhad, R.C. Development of an amperometric polyphenol biosensor based on fungal laccase immobilized on nitrocellulose membrane. Artif. Cell. Blood Sub. 2012, 40, 163–170. [Google Scholar] [CrossRef]
- Fang, Z.; Liu, X.; Chen, L.; Shen, Y.; Zhang, X.; Fang, W.; Wang, X.; Bao, X.; Xiao, Y. Identification of a laccase Glac15 from Ganoderma lucidum 77002 and its application in bioethanol production. Biotechnol. Biofuels 2015, 8, 54. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.B.; Gong, J.; Dai, W.K.; Kang, X.C.; Huang, Z.; Zhang, H.M.; Liu, W.; Liu, L.; Ma, J.P.; Xia, Z.L.; et al. The genome of Ganderma lucidum provide insights into triterpense biosynthesis and wood degradation. Plos ONE 2012, 7, 1932–6203. [Google Scholar]
- Campos, P.A.; Levin, L.N.; Wirth, S.A. Heterologous production, characterization and dye decolorization ability of a novel thermostable laccase isoenzyme from Trametestrogii BAFC 463. Process. Biochem. 2016, 51, 895–903. [Google Scholar] [CrossRef]
- Garrido-Bazán, V.; Téllez-Téllez, M.; Herrera-Estrella, A.; Díaz-Godínez, G.; Nava-Galicia, S.; Villalobos-López, M.; Arroyo-Becerra, A.; Bibbins-Martínez, M. Effect of textile dyes on activity and differential regulation of laccase genes from Pleurotus ostreatus grown in submerged fermentation. Amb. Express 2016, 6, 93–111. [Google Scholar] [CrossRef] [PubMed]
- Murugesan, K.; Kim, Y.M.; Jeon, J.R.; Chang, Y.S. Effect of metal ions on reactive dye decolorization by laccase from Ganoderma lucidum. J. Hazard. Mater. 2009, 168, 523–529. [Google Scholar] [CrossRef]
- Bourbonnais, R.; Paice, M.G. Oxidation of non-phenolic substrates: An expanded role of laccase in lignin biodegradation. FEBS Lett. 1990, 267, 99–102. [Google Scholar] [CrossRef]
- Call, H.P.; Mucke, I. History, overview and applications of mediated lignolytic systems, especially laccase-mediator systems (LignozymR process). J. Biotechnol. 1997, 53, 163–202. [Google Scholar] [CrossRef]
- Ashe, B.; Nguyen, L.N.; Hai, F.I.; Lee, D.J.; Van, D.M.J.P.; Leusch, F.D.L. Impacts of redox-mediator type on trace organic contaminants degradation by laccase: Degradation efficiency, laccase stability and effluent toxicity. Int. Biodeter. Biodegr. 2016, 113, 169–176. [Google Scholar] [CrossRef]
- Boer, C.G.; Obici, L.; Souza, C.G.M.D.; Peralta, R.M. Decolorization of synthetic dyes by solid state cultures of Lentinula (Lentinus) edodes producing manganese peroxidase as the main ligninolytic enzyme. Bioresour. Technol. 2004, 94, 107–112. [Google Scholar] [CrossRef]
- Zilly, A.; Jaqueline, C.M.; Bracht, A.; Giatti, M.D.S.C.; Carvajal, A.E.; Koehnlein, E.A. Influence of NaCl and Na2SO4 on the kinetics and dye decolorization ability of crude laccase from Ganoderma lucidum. Int. Biodeter. Biodegr. 2011, 65, 340–344. [Google Scholar] [CrossRef]
- Moreira, N.S.L.; Matheus, D.R.; Machado, K.M.G. Influence of pH on the growth, laccase activity and RBBR decolorization by tropical basidiomycetes. Braz. Arch. Biol. Technol. 2009, 52, 1075–1082. [Google Scholar] [CrossRef]
- Eichlerová, I.; Homolka, L.; Lisá, L.; Nerud, F. Orange G and Remazol Brilliant Blue R decolorization by white rot fungi Dichomitus squalens, Ischnoderma resinosum and Pleurotus calyptratus. Chemosphere 2005, 60, 398–404. [Google Scholar] [CrossRef] [PubMed]
- Teerapatsakul, C.; Parra, R.; Bucke, C.; Chitradon, L. Improvement of laccase reduction from Ganoderma sp. KU-ALK4 by medium engineering. World J. Microb. Biot. 2007, 23, 1519–1527. [Google Scholar] [CrossRef]
- Manavalan, T.; Manavalan, A.; Thangavelu, K.P.; Heese, K. Characterization of optimized production, purification and application of laccase from Ganoderma lucidum. Biochem. Eng. J. 2013, 70, 106–114. [Google Scholar] [CrossRef]
- Si, J.; Peng, F.; Cui, B. Purification, biochemical characterization and dye decolorization capacity of an alkali-resistant and metal-tolerant laccase from Trametes pubescens. Bioresour. Technol. 2013, 128, 49–57. [Google Scholar] [CrossRef]
- Yan, J.P.; Chen, D.D.; Yang, E.; Niu, J.Z.; Chen, Y.H.; Chagan, I. Purification and characterization of a thermo tolerant laccase isoform in Trametes trogii strain and its potential in dye decolorization. Int. Biodeter. Biodegr. 2014, 93, 186–194. [Google Scholar] [CrossRef]
- Patel, H.; Gupte, S.; Gahlout, M.; Gupte, A. Purification and characterization of an extracellular laccase from solid-state culture of Pleurotus ostreatus HP-1. 3 Biotech. 2014, 4, 77–84. [Google Scholar] [CrossRef]
- Chao, J.; Hou, J.; Kai, W.; Zhang, Y.; Chen, V. Biocatalytic degradation of carbamazepine with immobilized laccase-mediator membrane hybrid reactor. J. Membr. Sci. 2016, 502, 11–20. [Google Scholar]
- Graça, M.B.S.; Amorim, M.T.P.D.; Costa-Ferreira, M. Use of laccase together with redox mediators to decolourize Remazol Brilliant Blue, R. J. Biotechnol. 2001, 89, 123–129. [Google Scholar]
- Cantarella, G.; Galli, C.; Gentili, P. Oxidation of non-phenolic substrates with the laccase/n-hydroxyacetanilide system: Structure of the key intermediate from the mediator and mechanistic insight. NewJ. Chem. 2004, 28, 366–372. [Google Scholar] [CrossRef]
- Galli, C.; Gentili, P. Chemical messengers: Mediated oxidations with the enzyme laccase. J. Phys. Org. Chem. 2010, 17, 12–16. [Google Scholar] [CrossRef]
- Amit, K.; Deepti, S.; Sharma, K.K.; Sakshi, A.; Singh, A.K.; Gill, S.S.; Singhal, B. Gel-based purification and biochemical study of laccase isozymes from Ganoderma sp. and its role in enhanced cotton callogenesis. Front. Microbiol. 2017, 8, 674–682. [Google Scholar]
- Camarero, S.; Ibarra, D.; Martínez, Á.T.; Romero, J.; Gutiérrez, A.; Del, R.J.C. Paper pulp delignification using laccase and natural mediators. Enzym. Microb. Technol. 2007, 40, 1264–1271. [Google Scholar] [CrossRef]
- Vandertol-Vanier, H.A.; Vazquez-Duhalt, R.; Tinoco, R.; Pickard, M.A. Enhanced activity by poly (ethylene glycol) modification of Coriolopsis gallica laccase. J. Ind. Microbiol. Biot. 2002, 29, 214–220. [Google Scholar] [CrossRef]
- Benzina, O.; Daassi, D.; Zouari-Mechichi, H.; Frikha, F.; Woodward, S.; Belbahri, L.; Rodriguez-Couto, S.; Mechichi, T. Decolorization and detoxification of two textile industry effluents by the laccase/1-hydroxybenzotriazole system. Eniviron. Sci. Pollut. R. 2013, 20, 5177–5187. [Google Scholar] [CrossRef]
- Yang, H.; Sun, H.; Zhang, S.; Wu, B.; Pan, B. Potential of acetylacetone as a mediator for Trametes versicolor laccase in enzymatic transformation of organic pollutants. Eniviron. Sci. Pollut. R. 2015, 22, 10882–10889. [Google Scholar] [CrossRef]
- Tychanowicz, G.K.; Souza, D.F.D.; Souza, C.G.M.; Kadowaki, M.K.; Peralta, R.M. copper improves the production of laccase by the white- rot fungus pleurotus pulmonarius in solid state fermentation. Braz. Arch. Biol. Technol. 2006, 49, 699–704. [Google Scholar] [CrossRef]
- Lorenzo, M.; Moldes, D.; Rodríguez Couto, S.; Sanromán, M.A. Inhibition of laccase activity from Trametes versicolor by heavy metals and organic compounds. Chemosphere 2005, 60, 1124–1128. [Google Scholar] [CrossRef]
- Zhuo, R.; Yuan, P.; Yang, Y.; Zhang, S.; Ma, F.Y.; Zhang, X.Y. Induction of laccase by metal ions and aromatic compounds in Pleurotus ostreatus HAUCC 162 and decolorization of different synthetic dyes by the extracellular laccase. Biochem. Eng. J. 2017, 117, 62–72. [Google Scholar] [CrossRef]
- Rodríguez, C.S.; Sanromán, M.A.; Gübitz, G.M. Influence of redox mediators and metal ions on synthetic acid dye decolourization by crude laccase from Trametes hirsuta. Chemosphere 2005, 58, 417–422. [Google Scholar] [CrossRef]
- Das, N.; Sengupta, S.; Mukherjee, M. Importance of laccase in vegetative growth of Pleurotus florida. Appl. Environ. Microbiol. 1997, 63, 4120–4122. [Google Scholar] [PubMed]
- Matcham, S.E.; Jordan, B.R.; Wood, D.A. Estimation of fungal biomass in a soil substrate by three independent methods. Appl. Microbiol. Biot. 1985, 21, 108–112. [Google Scholar]
- Jin, W.S.; Li, J.H.; Feng, H.C.; You, S.; Zhang, L.Y.; Norvienyeku, J.; Hu, K.H.; Sun, S.J.; Wang, Z.H. Importance of a laccase gene (Lcc1) in the development of Ganode rmatsugae. Int. J. Mol. Sci. 2018, 19, 471. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Ng, T.B.; Lin, J.; Ye, X. A novel laccase from basidiomycete Cerrena sp.: Cloning, heterologous expression, and characterization. Int. J. Biol. Macromol. 2015, 77, 344–349. [Google Scholar] [CrossRef]
- Kim, S.I.; Ha, B.S.; Kim, M.S.; Park, M.; Ro, H.S. Evaluation of copper-inducible fungal laccase promoter in foreign gene expression in Pichia pastoris. Biotechnol. Bioproc. Eng. 2016, 21, 53–59. [Google Scholar] [CrossRef]
- Piscitelli, A.; Giardina, P.; Lettera, V.; Pezzella, C.; Sannia, G.; Vincenza, F. Induction and transcriptional regulation of laccases in fungi. Curr. Genom. 2011, 12, 104–112. [Google Scholar] [CrossRef]
- Pezzella, C.; Lettera, V.; Piscitelli, A.; Giardina, P.S.; Sannia, G. Transcriptional analysis of Pleurotus ostreatus laccase genes. Appl. Microbiol. Biotechnol. 2013, 97, 705–717. [Google Scholar] [CrossRef]
- Courty, P.E.; Hoegger, P.J.; Kilaru, S.; Kohler, A.; Buée, M.; Garbaye, J.; Martin, F.; Kües, U. Phylogenetic analysis, genomic organization, and expression analysis of multi-copper oxidases in the ectomycorrhizal basidiomycete Laccaria bicolor. New Phytol. 2009, 182, 736–750. [Google Scholar] [CrossRef]
- Shen, K.Y.; Zhang, X.B.; Qin, P.; Cheng, Q.; Xiang, Q.J. Enzymatic activity and transcription of Ganoderma lucidum laccases following treatment with four heavy metals. Chin. J. Appl. Environ. Biol. 2017, 23, 42–47. [Google Scholar]
Sample Availability: Samples of the compounds are available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treat No. | Temperature (°C) | pH | Dye Concentration (ppm) | Decolorization Percentage (%) |
|---|---|---|---|---|
| 1 | 25 | 4.0 | 100 | 39.20 ± 0.34 d |
| 2 | 25 | 4.5 | 150 | 44.00 ± 0.85 c |
| 3 | 25 | 5.0 | 200 | 35.41 ± 0.08 e |
| 4 | 30 | 4.0 | 150 | 47.65 ± 0.84 b |
| 5 | 30 | 4.5 | 200 | 49.95 ± 0.86 a |
| 6 | 30 | 5.0 | 100 | 26.78 ± 1.00 f |
| 7 | 35 | 4.0 | 200 | 50.34 ± 0.30 a |
| 8 | 35 | 4.5 | 100 | 40.88 ± 0.58 d |
| 9 | 35 | 5.0 | 150 | 45.49 ± 1.23 bc |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, P.; Wu, Y.; Adil, B.; Wang, J.; Gu, Y.; Yu, X.; Zhao, K.; Zhang, X.; Ma, M.; Chen, Q.; et al. Optimization of Laccase from Ganoderma lucidum Decolorizing Remazol Brilliant Blue R and Glac1 as Main Laccase-Contributing Gene. Molecules 2019, 24, 3914. https://doi.org/10.3390/molecules24213914
Qin P, Wu Y, Adil B, Wang J, Gu Y, Yu X, Zhao K, Zhang X, Ma M, Chen Q, et al. Optimization of Laccase from Ganoderma lucidum Decolorizing Remazol Brilliant Blue R and Glac1 as Main Laccase-Contributing Gene. Molecules. 2019; 24(21):3914. https://doi.org/10.3390/molecules24213914
Chicago/Turabian StyleQin, Peng, Yuetong Wu, Bilal Adil, Jie Wang, Yunfu Gu, Xiumei Yu, Ke Zhao, Xiaoping Zhang, Menggen Ma, Qiang Chen, and et al. 2019. "Optimization of Laccase from Ganoderma lucidum Decolorizing Remazol Brilliant Blue R and Glac1 as Main Laccase-Contributing Gene" Molecules 24, no. 21: 3914. https://doi.org/10.3390/molecules24213914
APA StyleQin, P., Wu, Y., Adil, B., Wang, J., Gu, Y., Yu, X., Zhao, K., Zhang, X., Ma, M., Chen, Q., Chen, X., Zhang, Z., & Xiang, Q. (2019). Optimization of Laccase from Ganoderma lucidum Decolorizing Remazol Brilliant Blue R and Glac1 as Main Laccase-Contributing Gene. Molecules, 24(21), 3914. https://doi.org/10.3390/molecules24213914