Methods to Discover and Evaluate Proteasome Small Molecule Stimulators

Abstract

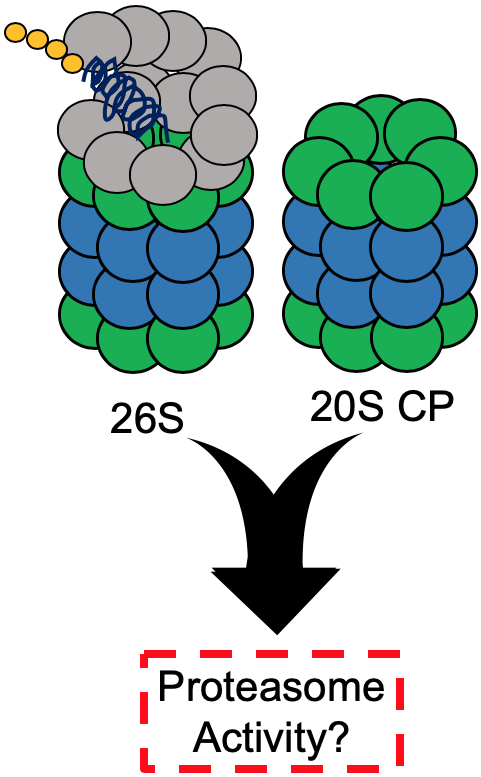
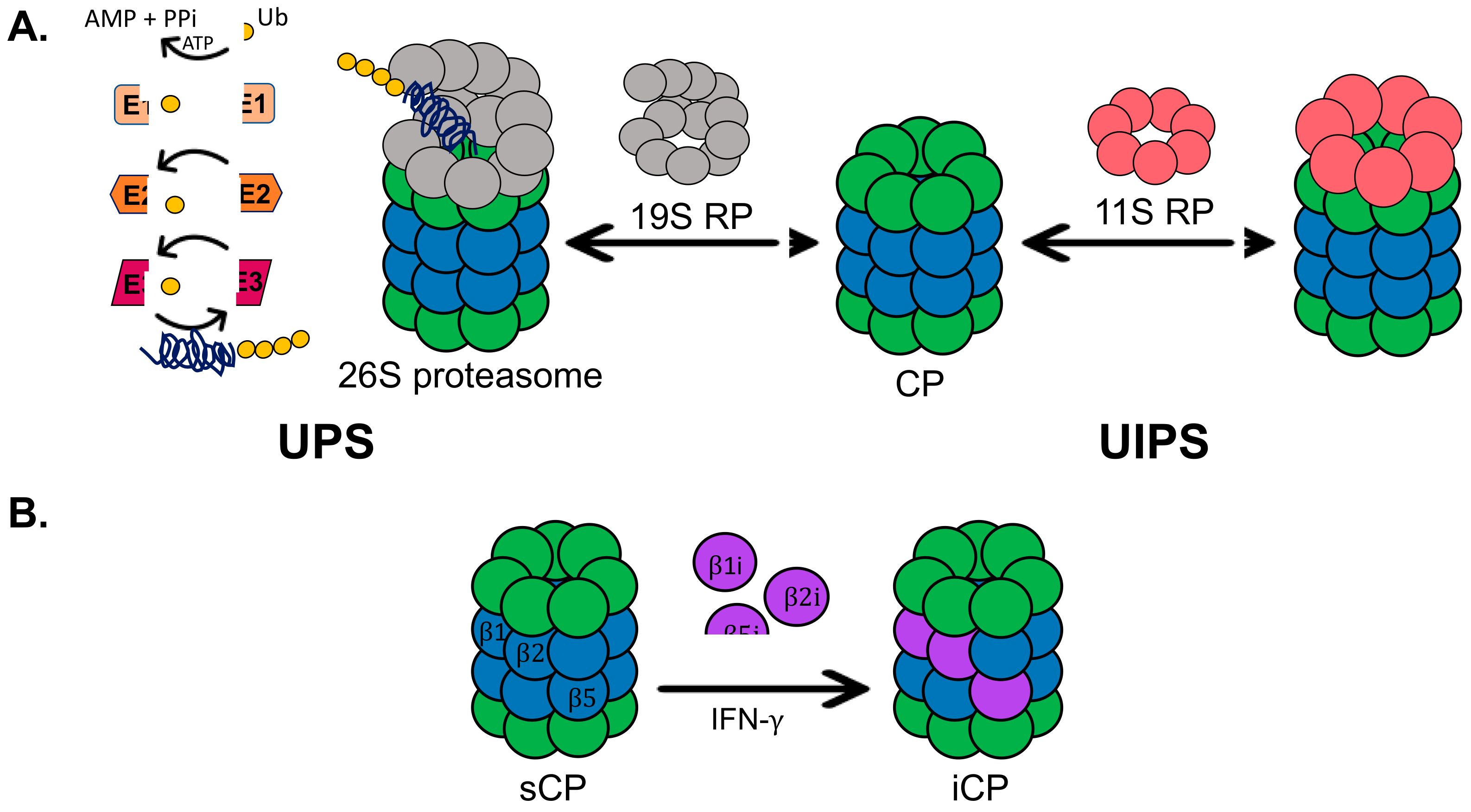
1. Introduction
2. Discovery: Biochemical High-Throughput Screening Assays
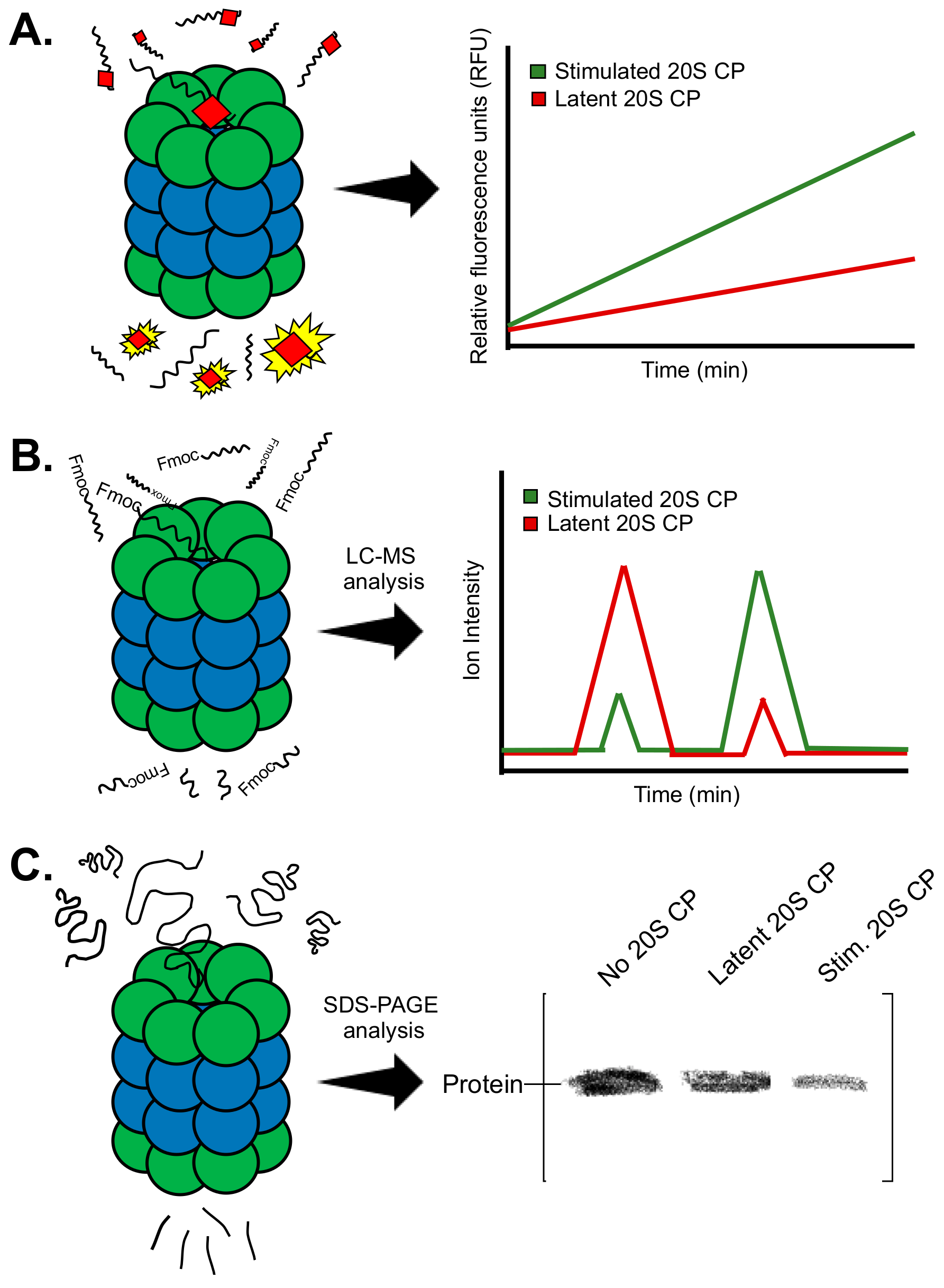
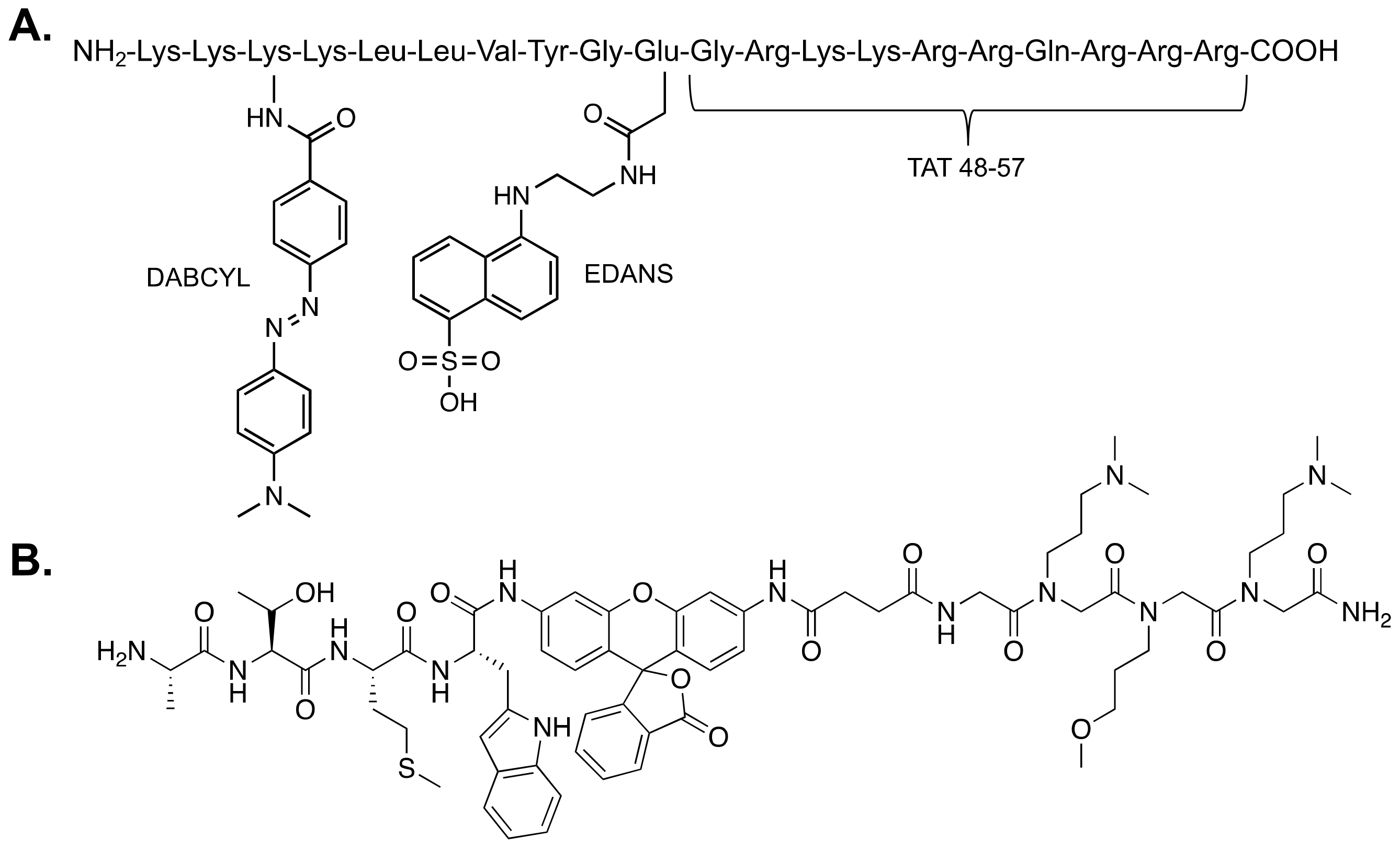
2.1. Fluorogenic Peptide Substrates
2.2. Mass Spectrometry Substrate Analysis
2.3. Gel-Based Assays
2.4. General Mechanism of Small Molecule Stimulators
3. Discovery: In Vitro Assays
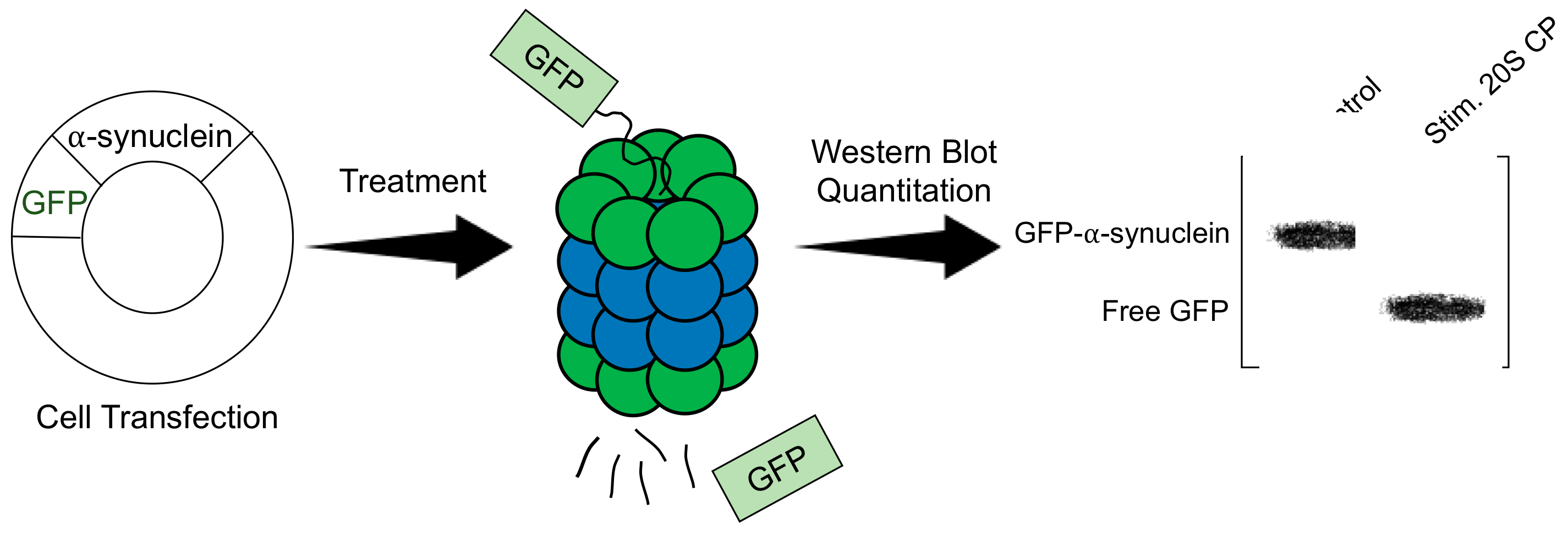
3.1. GFP-Fusion Protein Assay: 20S CP Stimulator
3.2. Analysis of Endogenous Protein Levels
4. Methods to Monitor Proteasome Activity in Cells
4.1. GFP-Fusion Protein Assay: 26S Proteasome Stimulator
4.2. Radioactively Labeled Proteasome Substrates
4.3. In Vitro Fluorogenic and Bioluminescent Reporters
5. Structural Evaluation of Small Molecule Stimulators of the Proteasome
5.1. AFM
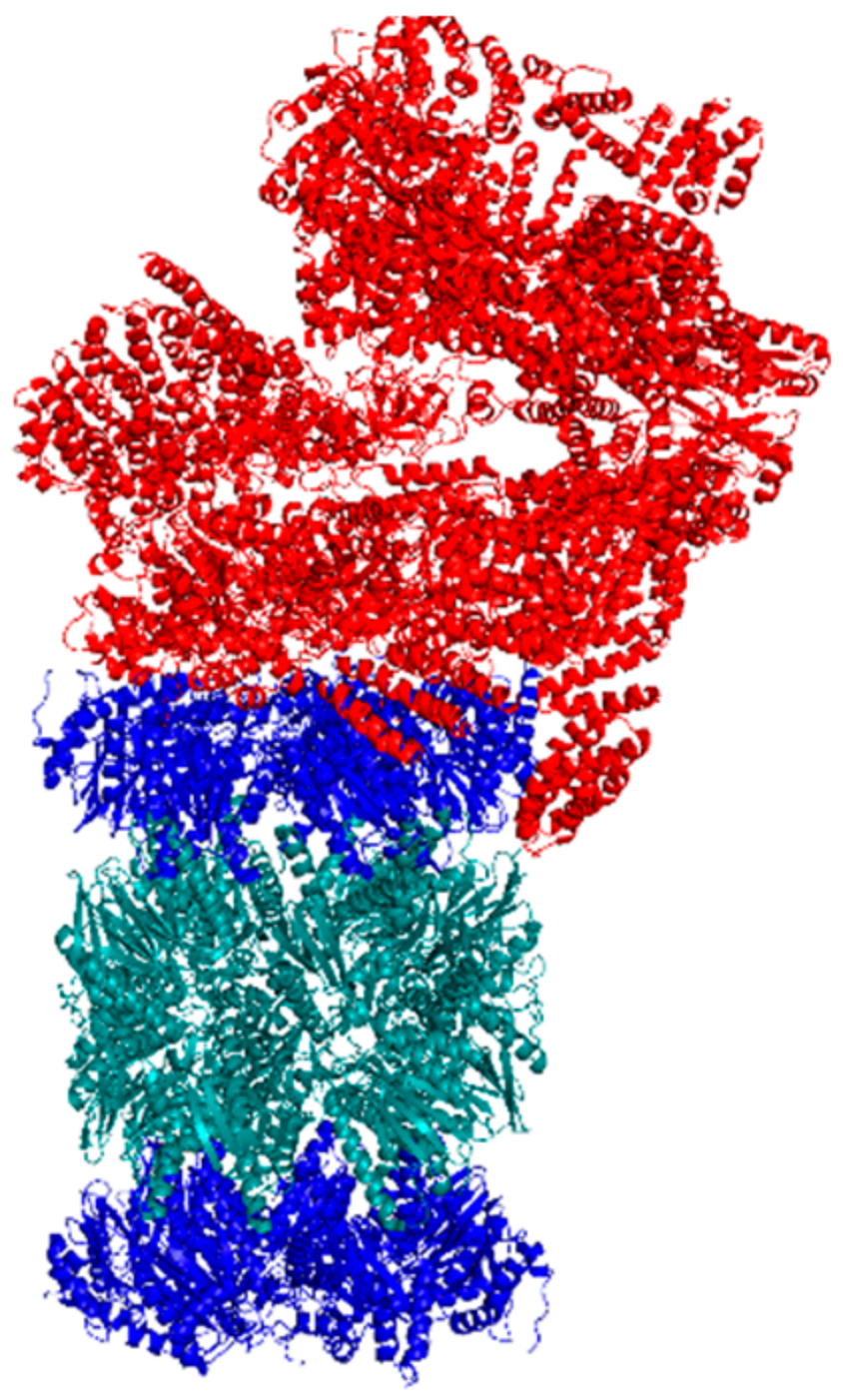
5.2. Cryo-EM
6. Conclusions
Funding
Conflicts of Interest
References
- Glick, D.; Barth, S.; Macleod, K.F. Autophagy: Cellular and Molecular Mechanisms. J. Pathol. 2010, 221, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Sorokin, A.V.; Kim, E.R.; Ovchinnikov, L.P. Proteasome System of Protein Degradation and Processing. Biochem. Biokhimiia 2009, 74, 1411–1442. [Google Scholar] [CrossRef]
- Ciechanover, A. The Ubiquitin-Proteasome Proteolytic Pathway. Cell 1994, 79, 13–21. [Google Scholar] [CrossRef]
- Erales, J.; Coffino, P. Ubiquitin-Independent Proteasomal Degradation. Biochim. Biophys. Acta BBA - Mol. Cell Res. 2014, 1843, 216–221. [Google Scholar] [CrossRef] [PubMed]
- Ehlinger, A.; Walters, K.J. Structural Insights into Proteasome Activation by the 19S Regulatory Particle. Biochemistry 2013, 52. [Google Scholar] [CrossRef] [PubMed]
- Ben-Nissan, G.; Sharon, M. Regulating the 20S Proteasome Ubiquitin-Independent Degradation Pathway. Biomolecules 2014, 4, 862–884. [Google Scholar] [CrossRef]
- Ruggiano, A.; Foresti, O.; Carvalho, P. ER-Associated Degradation: Protein Quality Control and Beyond. J. Cell Biol. 2014, 204, 869–879. [Google Scholar] [CrossRef]
- Werner, E.D.; Brodsky, J.L.; McCracken, A.A. Proteasome-Dependent Endoplasmic Reticulum-Associated Protein Degradation: An Unconventional Route to a Familiar Fate. Proc. Natl. Acad. Sci. USA 1996, 93, 13797–13801. [Google Scholar] [CrossRef]
- Hebert, D.N.; Molinari, M. In and out of the ER: Protein Folding, Quality Control, Degradation, and Related Human Diseases. Physiol. Rev. 2007, 87, 1377–1408. [Google Scholar] [CrossRef]
- Lindholm, D.; Wootz, H.; Korhonen, L. ER Stress and Neurodegenerative Diseases. Cell Death Differ. 2006, 13, 385–392. [Google Scholar] [CrossRef]
- Pickering, A.M.; Davies, K.J.A. Degradation of Damaged Proteins—The Main Function of the 20S Proteasome. Prog. Mol. Biol. Transl. Sci. 2012, 109, 227–248. [Google Scholar] [CrossRef]
- Asher, G.; Reuven, N.; Shaul, Y. 20S Proteasomes and Protein Degradation “by Default”. BioEssays News Rev. Mol. Cell. Dev. Biol. 2006, 28, 844–849. [Google Scholar] [CrossRef] [PubMed]
- Stadtmueller, B.M.; Hill, C.P. Proteasome Activators. Mol. Cell 2011, 41, 8–19. [Google Scholar] [CrossRef] [PubMed]
- Fabre, B.; Lambour, T.; Garrigues, L.; Ducoux-Petit, M.; Amalric, F.; Monsarrat, B.; Burlet-Schiltz, O.; Bousquet-Dubouch, M.-P. Label-Free Quantitative Proteomics Reveals the Dynamics of Proteasome Complexes Composition and Stoichiometry in a Wide Range of Human Cell Lines. J. Proteome Res. 2014, 13, 3027–3037. [Google Scholar] [CrossRef] [PubMed]
- Heink, S.; Ludwig, D.; Kloetzel, P.-M.; Krüger, E. IFN-Gamma-Induced Immune Adaptation of the Proteasome System Is an Accelerated and Transient Response. Proc. Natl. Acad. Sci. USA 2005, 102, 9241–9246. [Google Scholar] [CrossRef] [PubMed]
- Ferrington, D.A.; Gregerson, D.S. Immunoproteasomes: Structure, Function, and Antigen Presentation. Prog. Mol. Biol. Transl. Sci. 2012, 109, 75–112. [Google Scholar] [PubMed]
- McNaught, K.S.P.; Belizaire, R.; Isacson, O.; Jenner, P.; Olanow, C.W. Altered Proteasomal Function in Sporadic Parkinson’s Disease. Exp. Neurol. 2003, 179, 38–46. [Google Scholar] [CrossRef] [PubMed]
- McNaught, K.S.; Jenner, P. Proteasomal Function Is Impaired in Substantia Nigra in Parkinson’s Disease. Neurosci. Lett. 2001, 297, 191–194. [Google Scholar] [CrossRef]
- Palmieri, G.; Cocca, E.; Gogliettino, M.; Valentino, R.; Ruvo, M.; Cristofano, G.; Angiolillo, A.; Balestrieri, M.; Rossi, M.; Di Costanzo, A. Low Erythrocyte Levels of Proteasome and Acyl-Peptide Hydrolase (APEH) Activities in Alzheimer’s Disease: A Sign of Defective Proteostasis? J. Alzheimers Dis. 2017, 60, 1097–1106. [Google Scholar] [CrossRef] [PubMed]
- Gadhave, K.; Bolshette, N.; Ahire, A.; Pardeshi, R.; Thakur, K.; Trandafir, C.; Istrate, A.; Ahmed, S.; Lahkar, M.; Muresanu, D.F.; et al. The Ubiquitin Proteasomal System: A Potential Target for the Management of Alzheimer’s Disease. J. Cell. Mol. Med. 2016, 20, 1392–1407. [Google Scholar] [CrossRef] [PubMed]
- Arrasate, M.; Finkbeiner, S. Protein Aggregates in Huntington’s Disease. Exp. Neurol. 2012, 238, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kabashi, E.; Agar, J.N.; Strong, M.J.; Durham, H.D. Impaired Proteasome Function in Sporadic Amyotrophic Lateral Sclerosis. Amyotroph. Lateral Scler. Off. Publ. World Fed. Neurol. Res. Group Mot. Neuron Dis. 2012, 13, 367–371. [Google Scholar] [CrossRef] [PubMed]
- Hipkiss, A.R. Accumulation of Altered Proteins and Ageing: Causes and Effects. Exp. Gerontol. 2006, 41, 464–473. [Google Scholar] [CrossRef] [PubMed]
- Saez, I.; Vilchez, D. The Mechanistic Links Between Proteasome Activity, Aging and Age-Related Diseases. Curr. Genomics 2014, 15, 38. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, X.; Zhang, T.; Luna, C.; Liton, P.B.; Gonzalez, P. Cytoprotective Effects of Proteasome Beta5 Subunit Overexpression in Lens Epithelial Cells. Mol. Vis. 2007, 13, 31–38. [Google Scholar] [PubMed]
- Chondrogianni, N.; Tzavelas, C.; Pemberton, A.J.; Nezis, I.P.; Rivett, A.J.; Gonos, E.S. Overexpression of Proteasome Beta5 Assembled Subunit Increases the Amount of Proteasome and Confers Ameliorated Response to Oxidative Stress and Higher Survival Rates. J. Biol. Chem. 2005, 280, 11840–11850. [Google Scholar] [CrossRef]
- Vilchez, D.; Saez, I.; Dillin, A. The Role of Protein Clearance Mechanisms in Organismal Ageing and Age-Related Diseases. Nat. Commun. 2014, 5, 5659. [Google Scholar] [CrossRef] [PubMed]
- Chondrogianni, N.; Petropoulos, I.; Franceschi, C.; Friguet, B.; Gonos, E.S. Fibroblast Cultures from Healthy Centenarians Have an Active Proteasome. Exp. Gerontol. 2000, 35, 721–728. [Google Scholar] [CrossRef]
- Vilchez, D.; Boyer, L.; Morantte, I.; Lutz, M.; Merkwirth, C.; Joyce, D.; Spencer, B.; Page, L.; Masliah, E.; Berggren, W.T.; et al. Increased Proteasome Activity in Human Embryonic Stem Cells Is Regulated by PSMD11. Nature 2012, 489, 304–308. [Google Scholar] [CrossRef]
- Lokireddy, S.; Kukushkin, N.V.; Goldberg, A.L. CAMP-Induced Phosphorylation of 26S Proteasomes on Rpn6/PSMD11 Enhances Their Activity and the Degradation of Misfolded Proteins. Proc. Natl. Acad. Sci. USA 2015, 112, E7176–E7185. [Google Scholar] [CrossRef]
- Leestemaker, Y.; de Jong, A.; Witting, K.F.; Penning, R.; Schuurman, K.; Rodenko, B.; Zaal, E.A.; van de Kooij, B.; Laufer, S.; Heck, A.J.R.; et al. Proteasome Activation by Small Molecules. Cell Chem. Biol. 2017, 24, 725–736.e7. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.L.; Njomen, E.; Sjögren, B.; Dexheimer, T.S.; Tepe, J.J. Small Molecule Enhancement of 20S Proteasome Activity Targets Intrinsically Disordered Proteins. ACS Chem. Biol. 2017, 12, 2240–2247. [Google Scholar] [CrossRef] [PubMed]
- Coleman, R.A.; Trader, D.J. Development and Application of a Sensitive Peptide Reporter to Discover 20S Proteasome Stimulators. ACS Comb. Sci. 2018, 20, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Trader, D.J.; Simanski, S.; Dickson, P.; Kodadek, T. Establishment of A Suite of Assays That Support the Discovery of Proteasome Stimulators. Biochim. Biophys. Acta 2017, 1861, 892–899. [Google Scholar] [CrossRef] [PubMed]
- Njomen, E.; Osmulski, P.A.; Jones, C.L.; Gaczynska, M.; Tepe, J.J. Small Molecule Modulation of Proteasome Assembly. Biochemistry 2018, 57, 4214–4224. [Google Scholar] [CrossRef]
- Gillette, T.G.; Kumar, B.; Thompson, D.; Slaughter, C.A.; DeMartino, G.N. Differential Roles of the COOH Termini of AAA Subunits of PA700 (19 S Regulator) in Asymmetric Assembly and Activation of the 26 S Proteasome. J. Biol. Chem. 2008, 283, 31813–31822. [Google Scholar] [CrossRef] [PubMed]
- Kisselev, A.F.; Kaganovich, D.; Goldberg, A.L. Binding of Hydrophobic Peptides to Several Non-Catalytic Sites Promotes Peptide Hydrolysis by All Active Sites of 20 S Proteasomes EVIDENCE FOR PEPTIDE-INDUCED CHANNEL OPENING IN THE α-RINGS. J. Biol. Chem. 2002, 277, 22260–22270. [Google Scholar] [CrossRef]
- Dal Vechio, F.H.; Cerqueira, F.; Augusto, O.; Lopes, R.; Demasi, M. Peptides That Activate the 20S Proteasome by Gate Opening Increased Oxidized Protein Removal and Reduced Protein Aggregation. Free Radic. Biol. Med. 2014, 67, 304–313. [Google Scholar] [CrossRef]
- Tanaka, K.; Yoshimura, T.; Ichihara, A. Role of Substrate in Reversible Activation of Proteasomes (Multi-Protease Complexes) by Sodium Dodecyl Sulfate. J. Biochem. (Tokyo) 1989, 106, 495–500. [Google Scholar] [CrossRef]
- Ruiz de Mena, I.; Mahillo, E.; Arribas, J.; Castaño, J.G. Kinetic Mechanism of Activation by Cardiolipin (Diphosphatidylglycerol) of the Rat Liver Multicatalytic Proteinase. Biochem. J. 1993, 296 Pt 1, 93–97. [Google Scholar] [CrossRef]
- Huang, L.; Ho, P.; Chen, C.-H. Activation and Inhibition of Proteasomes by Betulinic Acid and Its Derivatives. FEBS Lett. 2007, 581, 4955–4959. [Google Scholar] [CrossRef] [PubMed]
- Coleman, R.A.; Muli, C.S.; Zhao, Y.; Bhardwaj, A.; Newhouse, T.R.; Trader, D.J. Analysis of Chain Length, Substitution Patterns, and Unsaturation of AM-404 Derivatives as 20S Proteasome Stimulators. Bioorg. Med. Chem. Lett. 2019, 29, 420–423. [Google Scholar] [CrossRef] [PubMed]
- Giżyńska, M.; Witkowska, J.; Karpowicz, P.; Rostankowski, R.; Chocron, E.S.; Pickering, A.M.; Osmulski, P.; Gaczynska, M.; Jankowska, E. Proline- and Arginine-Rich Peptides as Flexible Allosteric Modulators of Human Proteasome Activity. J. Med. Chem. 2019, 62, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Wilk, S.; Orlowski, M. Evidence That Pituitary Cation-Sensitive Neutral Endopeptidase Is a Multicatalytic Protease Complex. J. Neurochem. 1983, 40, 842–849. [Google Scholar] [CrossRef] [PubMed]
- Liggett, A.; Crawford, L.J.; Walker, B.; Morris, T.C.M.; Irvine, A.E. Methods for Measuring Proteasome Activity: Current Limitations and Future Developments. Leuk. Res. 2010, 34, 1403–1409. [Google Scholar] [CrossRef] [PubMed]
- Kisselev, A.F.; Goldberg, A.L. Monitoring Activity and Inhibition of 26S Proteasomes with Fluorogenic Peptide Substrates. In Methods in Enzymology; Ubiquitin and Protein Degradation, Part A; Academic Press: San Diego, CA, USA, 2005; Volume 398, pp. 364–378. [Google Scholar] [CrossRef]
- Schmidtke, G.; Holzhütter, H.-G.; Bogyo, M.; Kairies, N.; Groll, M.; de Giuli, R.; Emch, S.; Groettrup, M. How an Inhibitor of the HIV-I Protease Modulates Proteasome Activity. J. Biol. Chem. 1999, 274, 35734–35740. [Google Scholar] [CrossRef]
- Schmidtke, G.; Emch, S.; Groettrup, M.; Holzhutter, H.G. Evidence for the Existence of a Non-Catalytic Modifier Site of Peptide Hydrolysis by the 20 S Proteasome. J. Biol. Chem. 2000, 275, 22056–22063. [Google Scholar] [CrossRef]
- Schneider-Poetsch, T.; Ju, J.; Eyler, D.E.; Dang, Y.; Bhat, S.; Merrick, W.C.; Green, R.; Shen, B.; Liu, J.O. Inhibition of Eukaryotic Translation Elongation by Cycloheximide and Lactimidomycin. Nat. Chem. Biol. 2010, 6, 209–217. [Google Scholar] [CrossRef]
- Zhou, P. Determining Protein Half-Lives. In Signal Transduction Protocols; Dickson, R.C., Mendenhall, M.D., Eds.; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2004; pp. 67–77. [Google Scholar] [CrossRef]
- Kastrop, P.M.; Hulshof, S.C.; Bevers, M.M.; Destrée, O.H.; Kruip, T.A. The Effects of Alpha-Amanitin and Cycloheximide on Nuclear Progression, Protein Synthesis, and Phosphorylation during Bovine Oocyte Maturation in Vitro. Mol. Reprod. Dev. 1991, 28, 249–254. [Google Scholar] [CrossRef]
- Hochstrasser, M. Ubiquitin-Dependent Protein Degradation. Annu. Rev. Genet. 1996, 30, 405–439. [Google Scholar] [CrossRef]
- Dantuma, N.P.; Lindsten, K.; Glas, R.; Jellne, M.; Masucci, M.G. Short-Lived Green Fluorescent Proteins for Quantifying Ubiquitin/Proteasome-Dependent Proteolysis in Living Cells. Nat. Biotechnol. 2000, 18, 538. [Google Scholar] [CrossRef] [PubMed]
- Melvin, A.T.; Woss, G.S.; Park, J.H.; Waters, M.L.; Allbritton, N.L. Measuring Activity in the Ubiquitin-Proteasome System: From Large Scale Discoveries to Single Cells Analysis. Cell Biochem. Biophys. 2013, 67, 75–89. [Google Scholar] [CrossRef] [PubMed]
- Greussing, R.; Unterluggauer, H.; Koziel, R.; Maier, A.B.; Jansen-Dürr, P. Monitoring of Ubiquitin-Proteasome Activity in Living Cells Using a Degron (Dgn)-Destabilized Green Fluorescent Protein (GFP)-Based Reporter Protein. J. Vis. Exp. JoVE 2012. [Google Scholar] [CrossRef] [PubMed]
- Menéndez-Benito, V.; Heessen, S.; Dantuma, N.P. Monitoring of Ubiquitin-Dependent Proteolysis with Green Fluorescent Protein Substrates. Methods Enzymol. 2005, 399, 490–511. [Google Scholar] [PubMed]
- Varshavsky, A. The N-End Rule Pathway and Regulation by Proteolysis. Protein Sci. Publ. Protein Soc. 2011, 20, 1298–1345. [Google Scholar] [CrossRef] [PubMed]
- Sha, Z.; Zhao, J.; Goldberg, A.L. Measuring the Overall Rate of Protein Breakdown in Cells and the Contributions of the Ubiquitin-Proteasome and Autophagy-Lysosomal Pathways. Methods Mol. Biol. Clifton NJ 2018, 1844, 261–276. [Google Scholar]
- Anchoori, R.K.; Karanam, B.; Peng, S.; Wang, J.W.; Jiang, R.; Tanno, T.; Orlowski, R.Z.; Matsui, W.; Zhao, M.; Rudek, M.A.; et al. A Bis-Benzylidine Piperidone Targeting Proteasome Ubiquitin Receptor RPN13/ADRM1 as a Therapy for Cancer. Cancer Cell 2013, 24, 791–805. [Google Scholar] [CrossRef]
- Trader, D.J.; Simanski, S.; Kodadek, T. A Reversible and Highly Selective Inhibitor of the Proteasomal Ubiquitin Receptor Rpn13 Is Toxic To Multiple Myeloma Cells. J. Am. Chem. Soc. 2015, 137, 6312–6319. [Google Scholar] [CrossRef]
- Dyson, H.J.; Wright, P.E. Intrinsically Unstructured Proteins and Their Functions. Nat. Rev. Mol. Cell Biol. 2005, 6, 197–208. [Google Scholar] [CrossRef]
- Urru, S.A.M.; Veglianese, P.; De Luigi, A.; Fumagalli, E.; Erba, E.; Gonella Diaza, R.; Carrà, A.; Davoli, E.; Borsello, T.; Forloni, G.; et al. A New Fluorogenic Peptide Determines Proteasome Activity in Single Cells. J. Med. Chem. 2010, 53, 7452–7460. [Google Scholar] [CrossRef]
- Zerfas, B.L.; Trader, D.J. Monitoring the Immunoproteasome in Live Cells Using an Activity-Based Peptide–Peptoid Hybrid Probe. J. Am. Chem. Soc. 2019, 141, 5252–5260. [Google Scholar] [CrossRef] [PubMed]
- Järver, P.; Langel, Ü. Cell-Penetrating Peptides—A Brief Introduction. Biochim. Biophys. Acta BBA - Biomembr. 2006, 1758, 260–263. [Google Scholar] [CrossRef] [PubMed]
- Moravec, R.A.; O’Brien, M.A.; Daily, W.J.; Scurria, M.A.; Bernad, L.; Riss, T.L. Cell-Based Bioluminescent Assays for All Three Proteasome Activities in a Homogeneous Format. Anal. Biochem. 2009, 387, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Osmulski, P.A.; Gaczynska, M. Atomic Force Microscopy Reveals Two Conformations of the 20 S Proteasome from Fission Yeast. J. Biol. Chem. 2000, 275, 13171–13174. [Google Scholar] [CrossRef] [PubMed]
- Gaczynska, M.; Osmulski, P.A. Atomic Force Microscopy of Proteasome Assemblies. Methods Mol. Biol. Clifton NJ 2011, 736, 117–132. [Google Scholar]
- Huang, X.; Luan, B.; Wu, J.; Shi, Y. An Atomic Structure of the Human 26S Proteasome. Nat. Struct. Mol. Biol. 2016, 23, 778–785. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Zhang, S.; Wu, Z.; Li, X.; Wang, W.L.; Zhu, Y.; Stoilova-McPhie, S.; Lu, Y.; Finley, D.; Mao, Y. Cryo-EM Structures and Dynamics of Substrate-Engaged Human 26S Proteasome. Nature 2019, 565, 49. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Commonly Used Fluorogenic Peptide Substrates | ||
|---|---|---|
| Subunit specific cleavage | 1 | Z-LLE-NA Z-LLE-AMC Ac-nLPnLD-AMC |
| 2 | Boc-LRR-AMC Z-ARR-AMC Bz-VGR-AMC | |
| 5 | Suc-LLVY-AMC Z-GGL-AMC Suc-AAF-AMC | |
| Non-specific cleavage | FRET reporter | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coleman, R.A.; Trader, D.J. Methods to Discover and Evaluate Proteasome Small Molecule Stimulators. Molecules 2019, 24, 2341. https://doi.org/10.3390/molecules24122341
Coleman RA, Trader DJ. Methods to Discover and Evaluate Proteasome Small Molecule Stimulators. Molecules. 2019; 24(12):2341. https://doi.org/10.3390/molecules24122341
Chicago/Turabian StyleColeman, Rachel A., and Darci J. Trader. 2019. "Methods to Discover and Evaluate Proteasome Small Molecule Stimulators" Molecules 24, no. 12: 2341. https://doi.org/10.3390/molecules24122341
APA StyleColeman, R. A., & Trader, D. J. (2019). Methods to Discover and Evaluate Proteasome Small Molecule Stimulators. Molecules, 24(12), 2341. https://doi.org/10.3390/molecules24122341