
Effects of Lactobacillus curvatus HY7601 and Lactobacillus plantarum KY1032 on Overweight and the Gut Microbiota in Humans: Randomized, Double-Blinded, Placebo-Controlled Clinical Trial

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Test Materials
2.2. Study Subjects and Inclusion/Exclusion Criteria
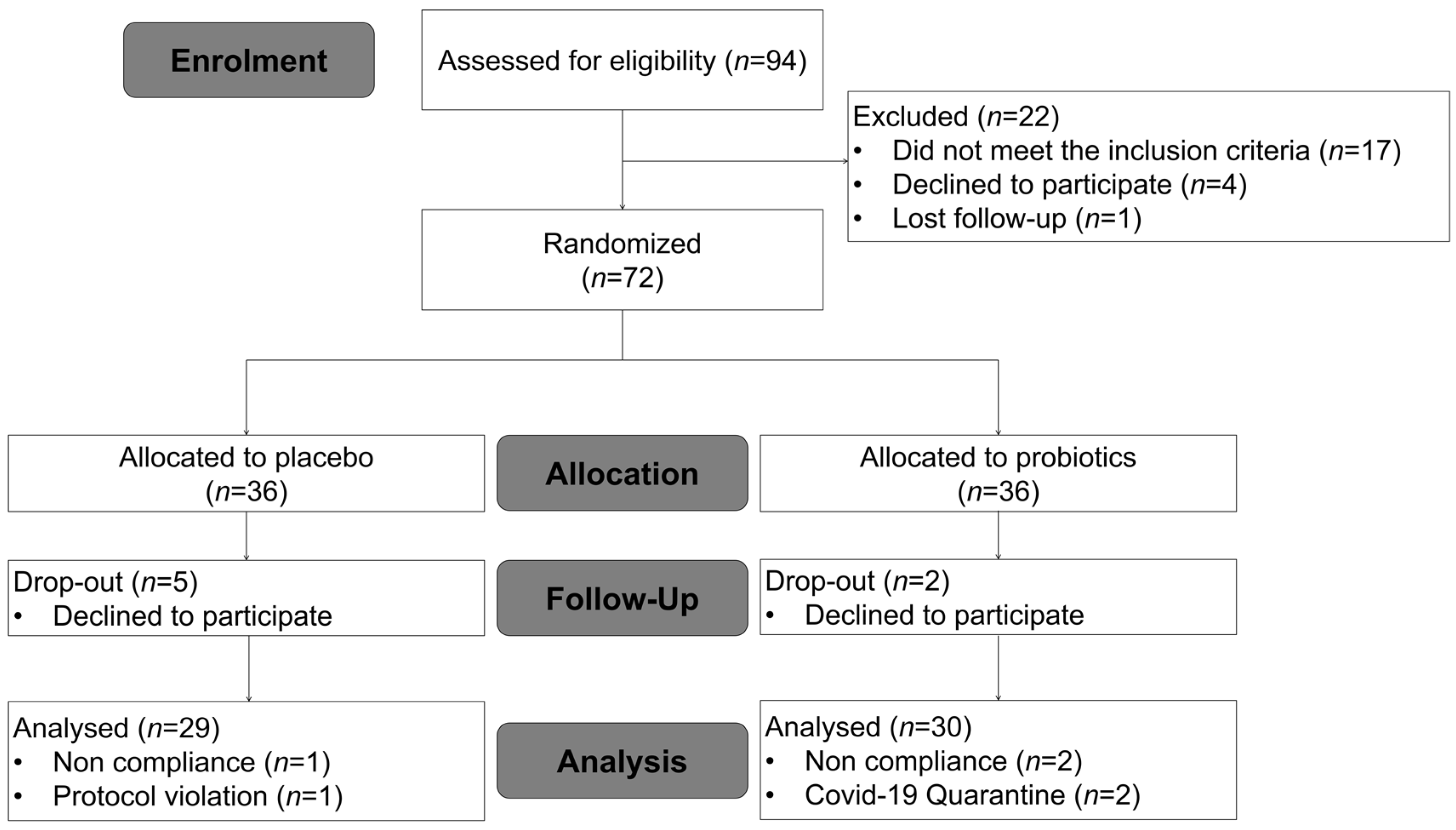
2.3. Study Design
2.4. Outcomes
2.5. Safety
2.6. Fecal Microbiome Analysis
2.6.1. Sample Handling and Collection
2.6.2. 16S rRNA Gene Amplicon Sequencing
2.6.3. Analysis of Operational Taxonomic Units (OTUs)
2.7. Statistics
3. Results
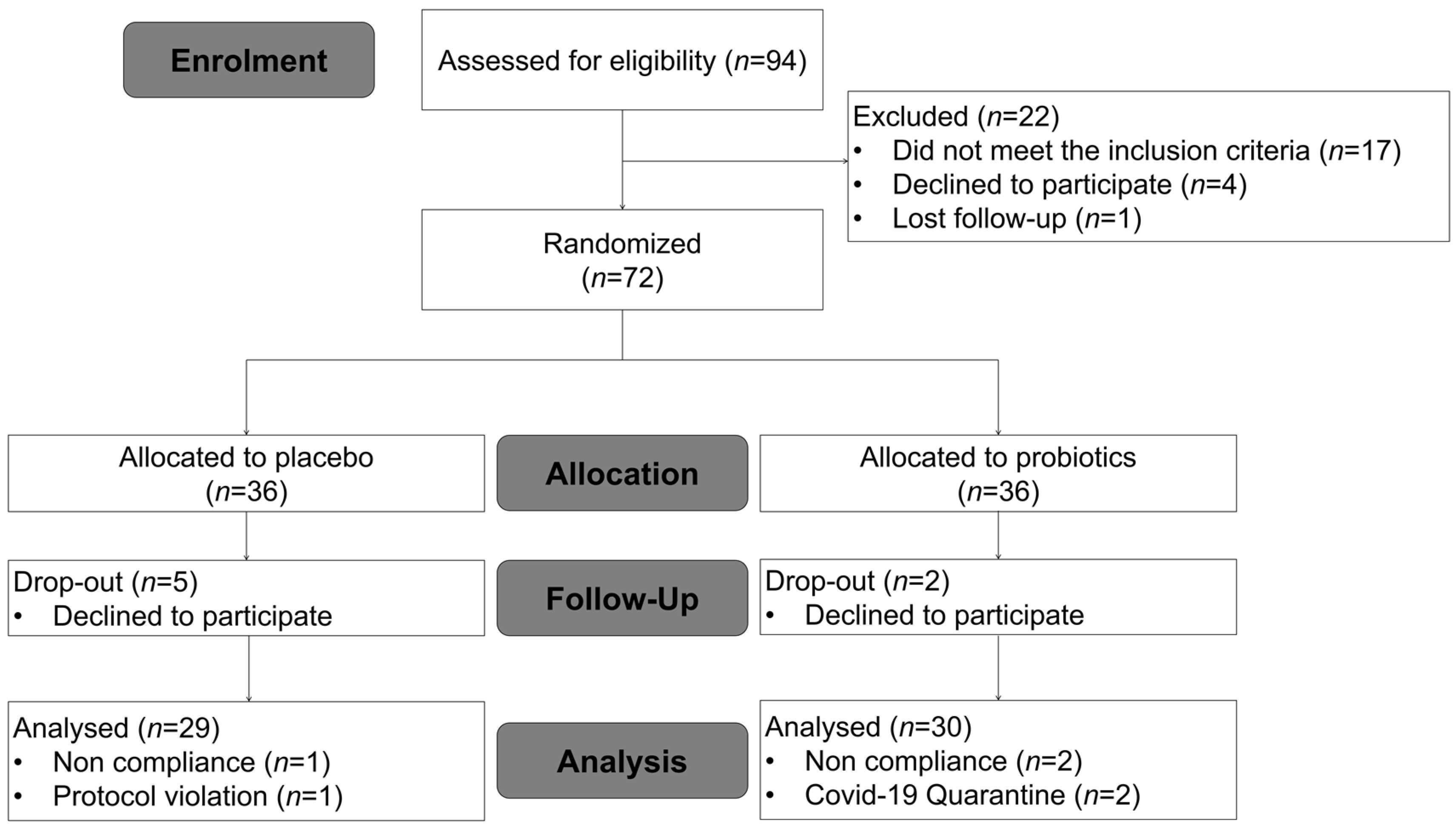
3.1. Baseline Characteristics of Subjects
3.2. Efficacy Analysis
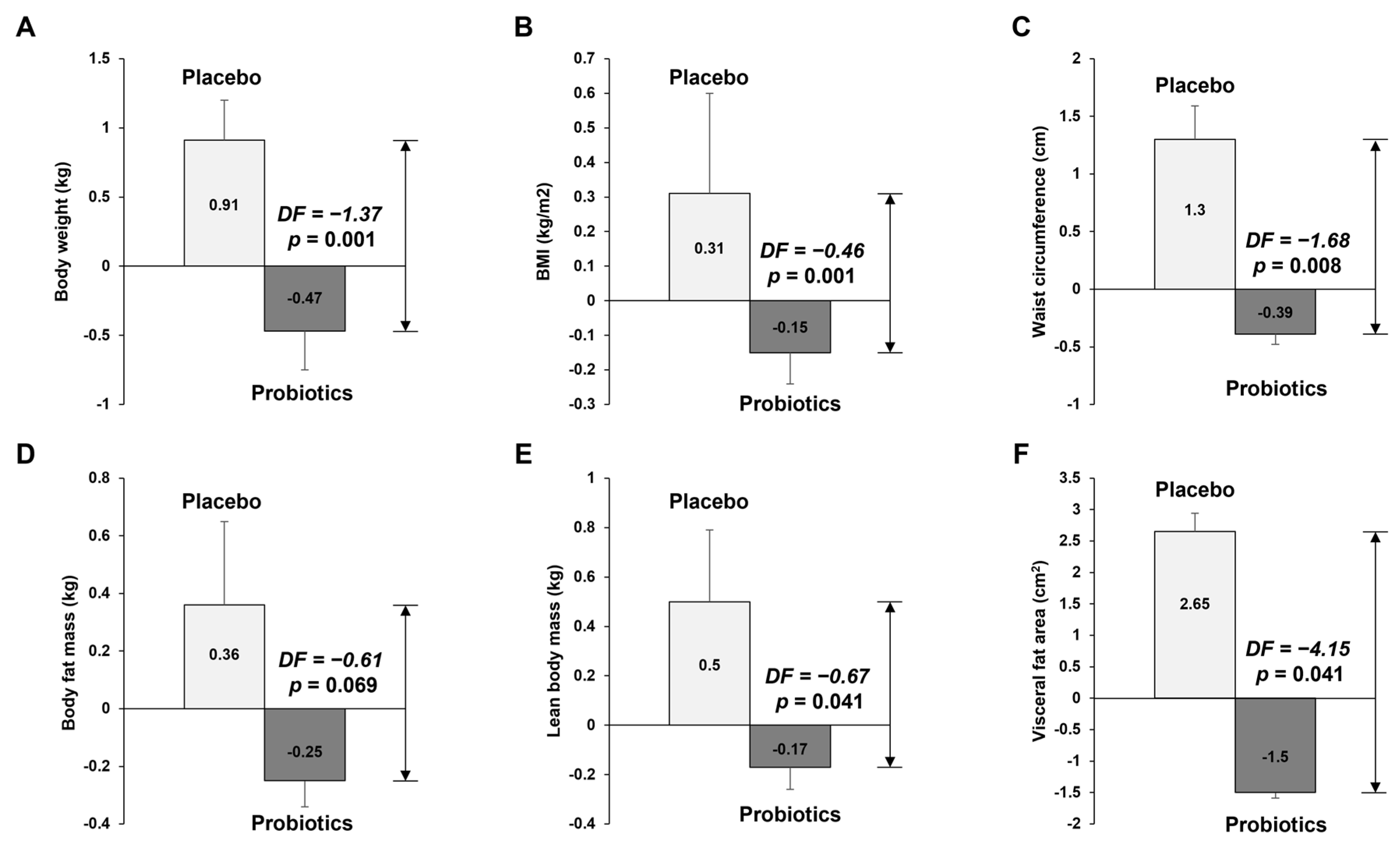
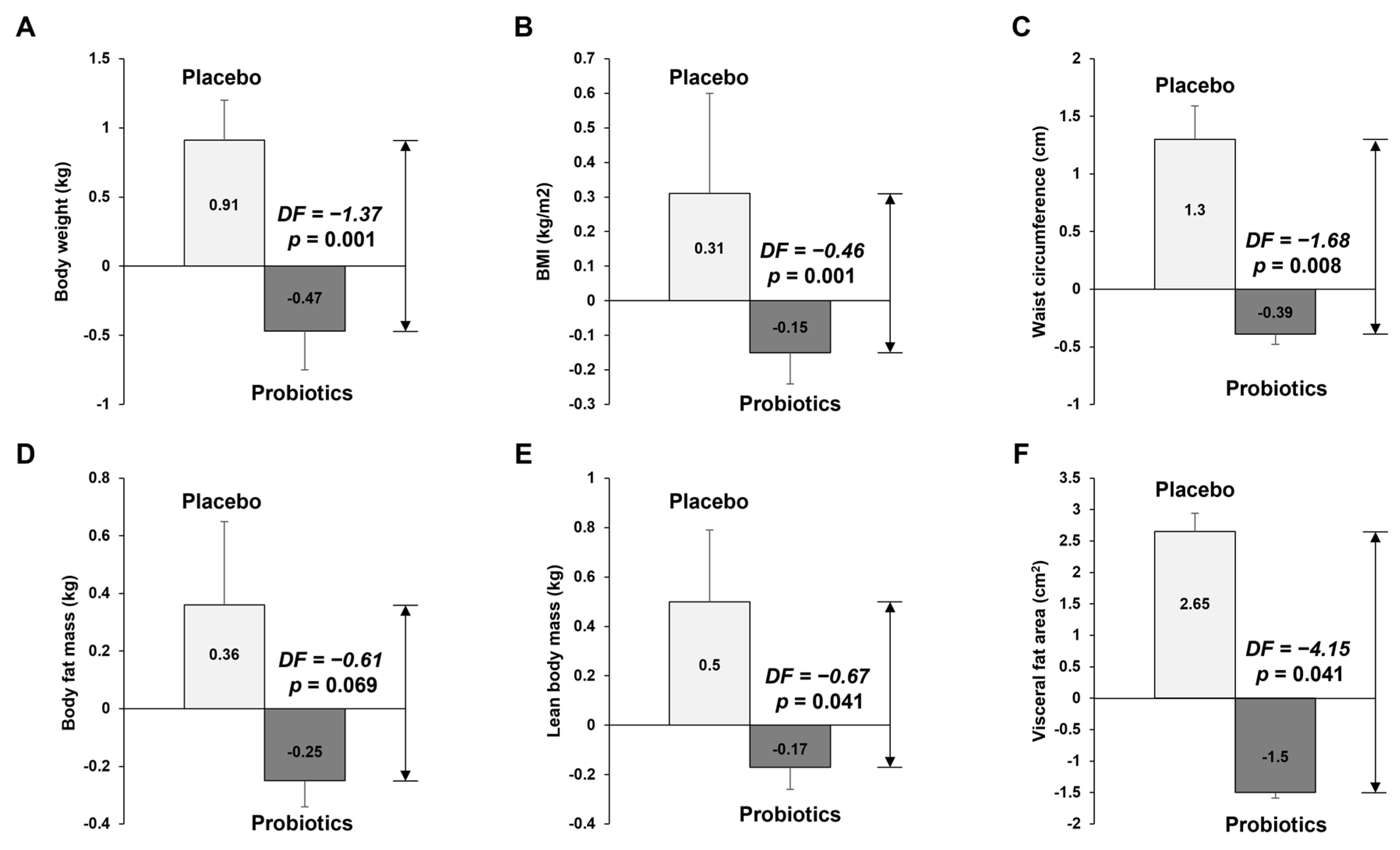
3.2.1. Anthropometric Measurement of Body Composition and Visceral Fat Area
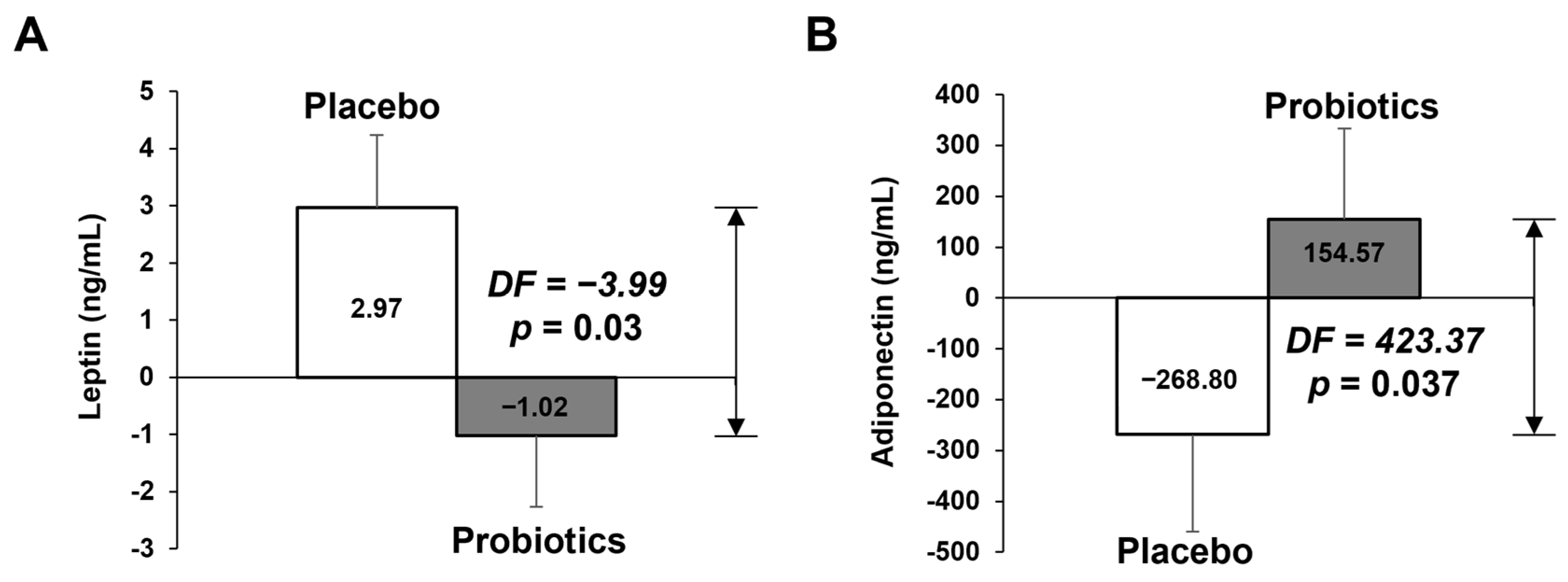
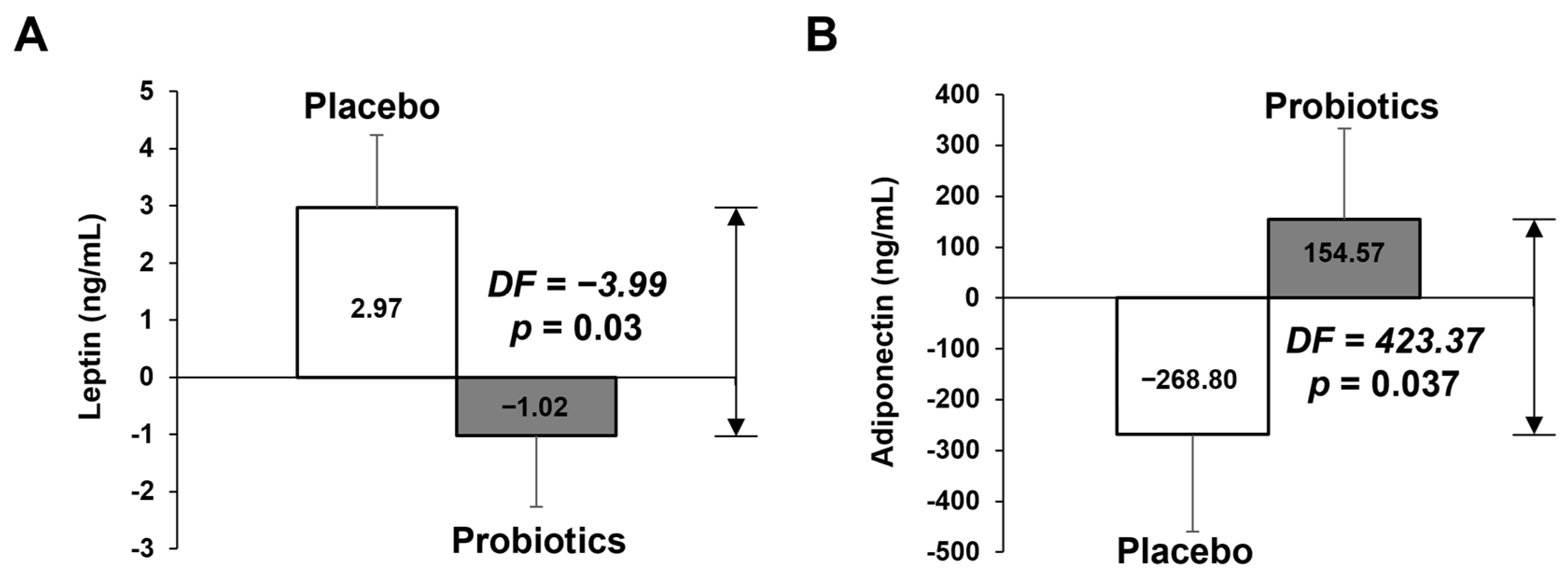
3.2.2. Measurement of Biochemical Variables in Blood
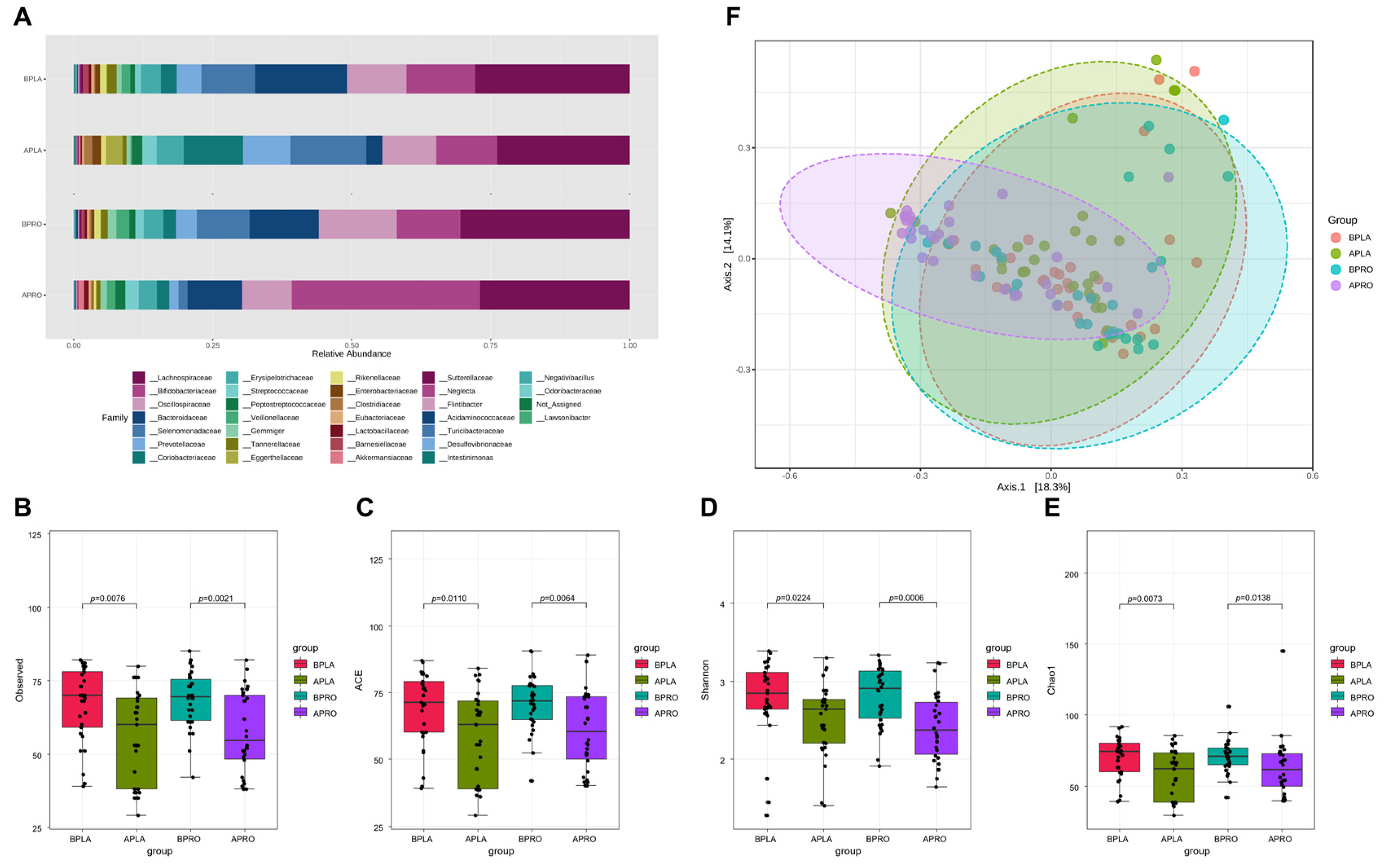
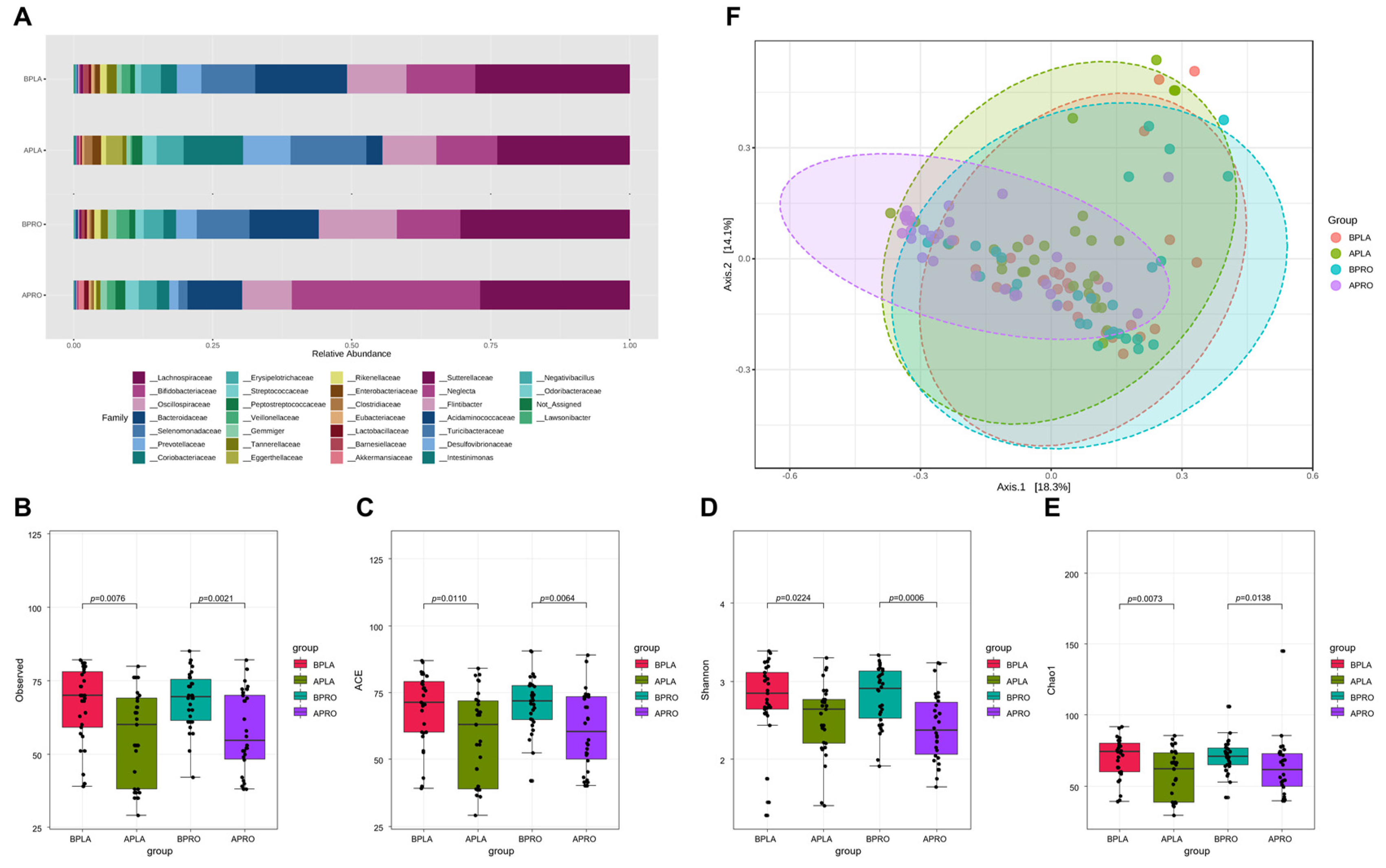
3.3. Microbiome Analyses
3.3.1. Composition of the Gut Microbiota, and Shifts in Bacterial Alpha and Beta Diversity
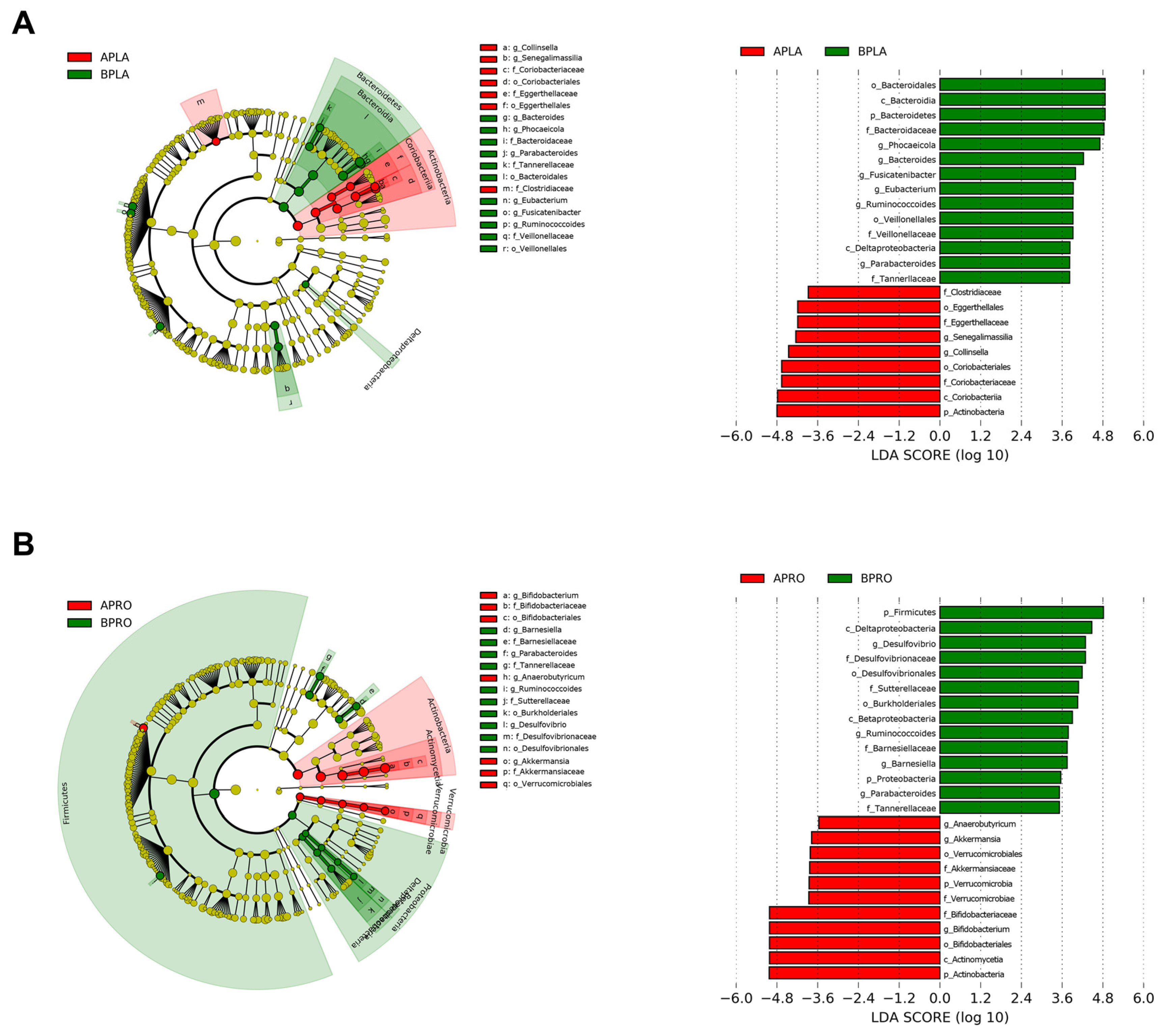
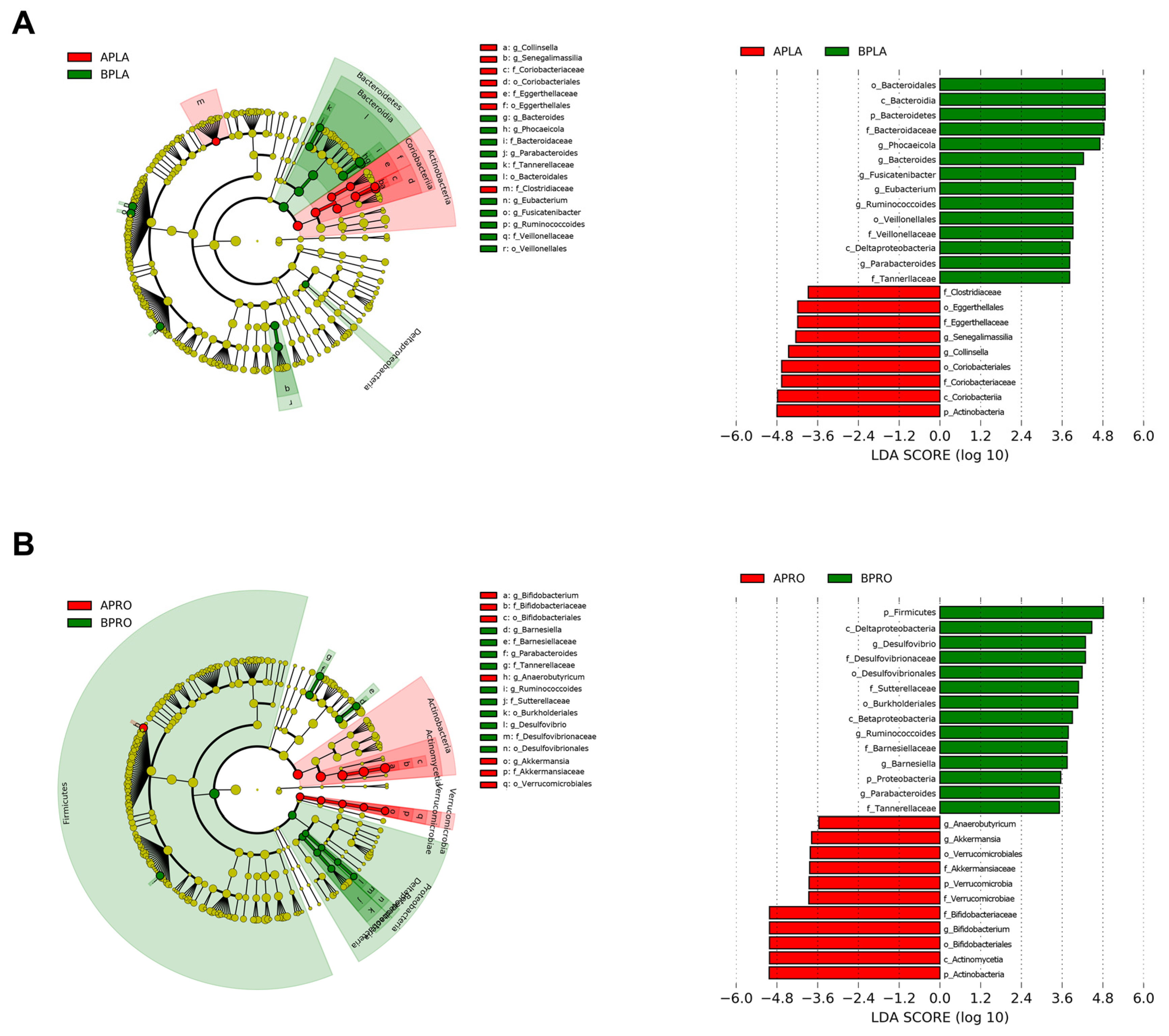
3.3.2. LEfSe
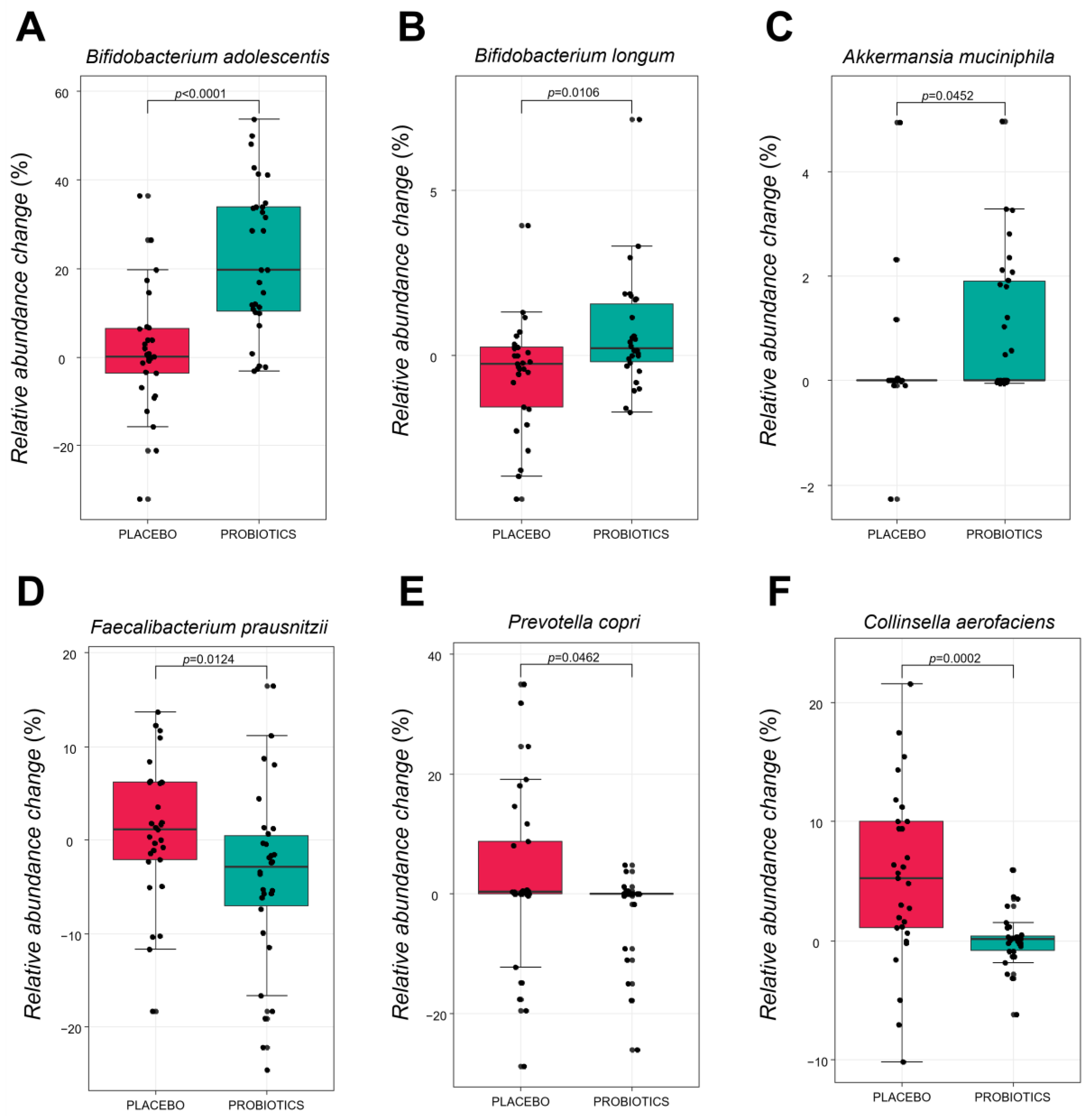
3.3.3. Comparative Analysis of the Gut Microbiota at the Species Level
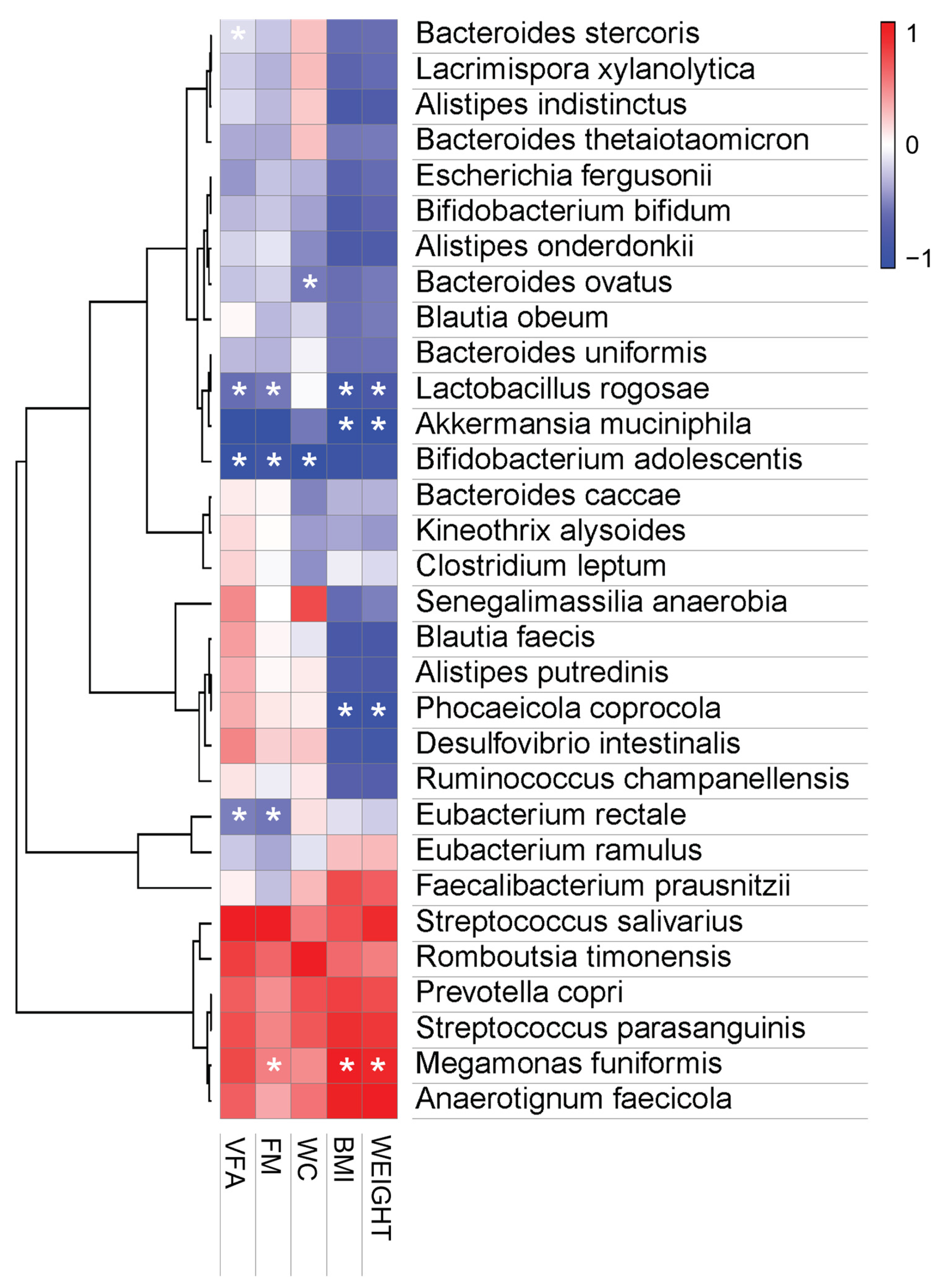
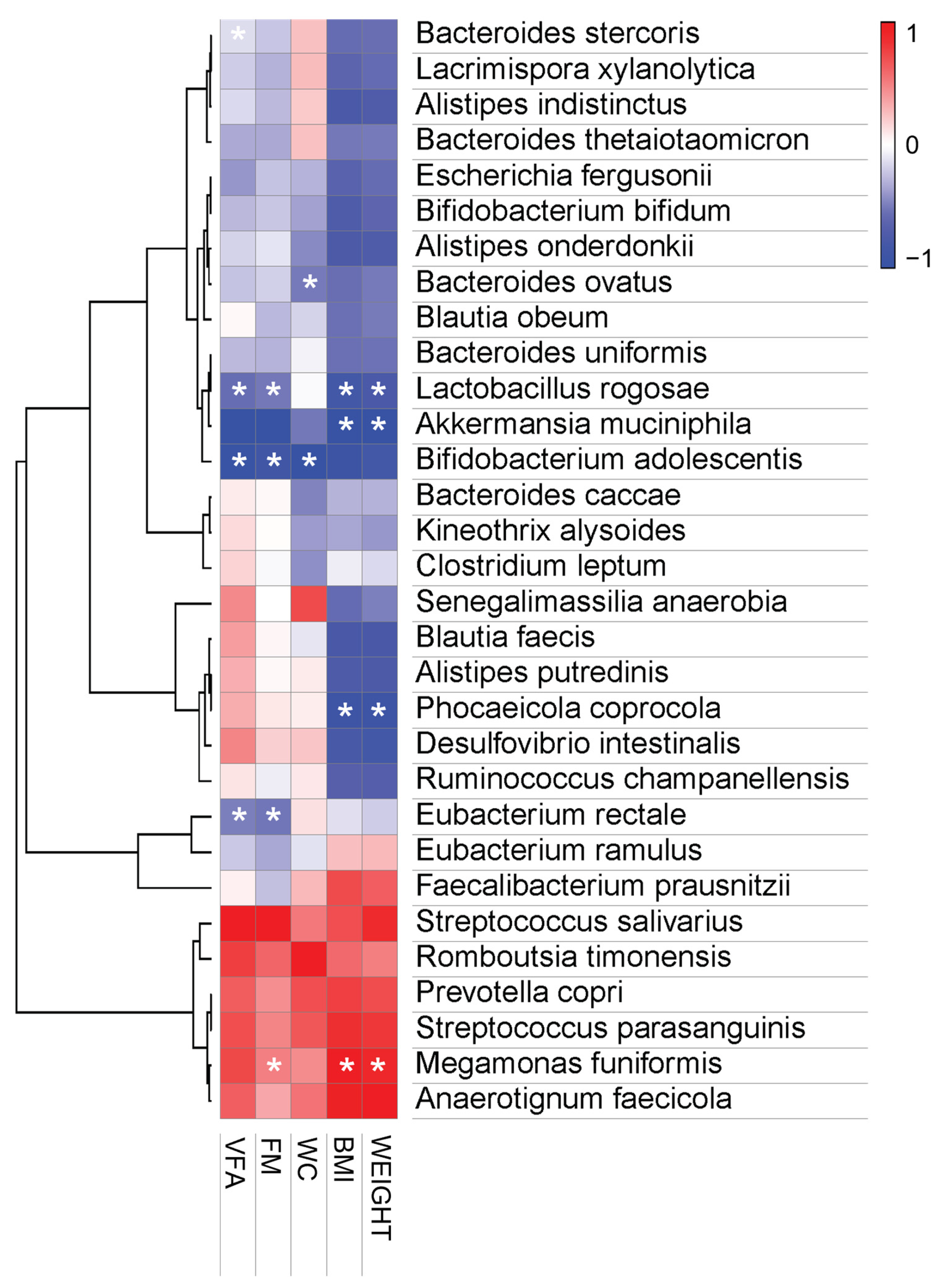
3.3.4. Pearson Correlation Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Camacho, S.; Ruppel, A. Is the calorie concept a real solution to the obesity epidemic? Glob. Health Action 2017, 10, 1289650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hruby, A.; Hu, F.B. The epidemiology of obesity: A big picture. Pharmacoeconomics 2015, 33, 673–689. [Google Scholar] [CrossRef] [PubMed]
- Haththotuwa, R.N.; Wijeyaratne, C.N.; Senarath, U. Worldwide epidemic of obesity. In Obesity and Obstetrics; Elsevier: Amsterdam, The Netherlands, 2020; pp. 3–8. [Google Scholar]
- Kyrou, I.; Randeva, H.S.; Tsigos, C.; Kaltsas, G.; Weickert, M.O. Clinical problems caused by obesity. eBook, Bookshelf ID:NBK278973; 2018; pp. 1–116. Available online: https://pubmed.ncbi.nlm.nih.gov/25905207/ (accessed on 30 April 2022).
- Fruh, S.M. Obesity: Risk factors, complications, and strategies for sustainable long-term weight management. J. Am. Assoc. Nurse Pract. 2017, 29, S3–S14. [Google Scholar] [CrossRef] [PubMed]
- Prentice, A.M.; Jebb, S.A. Beyond body mass index. Obes. Rev. 2001, 2, 141–147. [Google Scholar] [CrossRef]
- Gurunathan, U.; Myles, P. Limitations of Body Mass Index as an Obesity Measure of Perioperative Risk; Oxford University Press: Oxford, UK, 2016; Volume 116, pp. 319–321. [Google Scholar]
- Smith, U. Abdominal obesity: A marker of ectopic fat accumulation. J. Clin. Investig. 2015, 125, 1790–1792. [Google Scholar] [CrossRef] [Green Version]
- Ross, R.; Neeland, I.J.; Yamashita, S.; Shai, I.; Seidell, J.; Magni, P.; Santos, R.D.; Arsenault, B.; Cuevas, A.; Hu, F.B. Waist circumference as a vital sign in clinical practice: A Consensus Statement from the IAS and ICCR Working Group on Visceral Obesity. Nat. Rev. Endocrinol. 2020, 16, 177–189. [Google Scholar] [CrossRef]
- Bozeman, S.R.; Hoaglin, D.C.; Burton, T.M.; Pashos, C.L.; Ben-Joseph, R.H.; Hollenbeak, C.S. Predicting waist circumference from body mass index. BMC Med. Res. Methodol. 2012, 12, 115. [Google Scholar] [CrossRef] [Green Version]
- Aoun, A.; Darwish, F.; Hamod, N. The influence of the gut microbiome on obesity in adults and the role of probiotics, prebiotics, and synbiotics for weight loss. Prev. Nutr. Food Sci. 2020, 25, 113. [Google Scholar] [CrossRef]
- Abenavoli, L.; Scarpellini, E.; Colica, C.; Boccuto, L.; Salehi, B.; Sharifi-Rad, J.; Aiello, V.; Romano, B.; De Lorenzo, A.; Izzo, A.A. Gut microbiota and obesity: A role for probiotics. Nutrients 2019, 11, 2690. [Google Scholar] [CrossRef] [Green Version]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut microbiota functions: Metabolism of nutrients and other food components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Angelakis, E.; Armougom, F.; Million, M.; Raoult, D. The relationship between gut microbiota and weight gain in humans. Future Microbiol. 2012, 7, 91–109. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Shi, B. Gut microbiota as a potential target of metabolic syndrome: The role of probiotics and prebiotics. Cell Biosci. 2017, 7, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiciński, M.; Gębalski, J.; Gołębiewski, J.; Malinowski, B. Probiotics for the treatment of overweight and obesity in humans—A review of clinical trials. Microorganisms 2020, 8, 1148. [Google Scholar] [CrossRef] [PubMed]
- Plaza-Diaz, J.; Ruiz-Ojeda, F.J.; Gil-Campos, M.; Gil, A. Mechanisms of action of probiotics. Adv. Nutr. 2019, 10, S49–S66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pessione, E. Lactic acid bacteria contribution to gut microbiota complexity: Lights and shadows. Front. Cell. Infect. Microbiol. 2012, 2, 86. [Google Scholar] [CrossRef] [Green Version]
- De Filippis, F.; Pasolli, E.; Ercolini, D. The food-gut axis: Lactic acid bacteria and their link to food, the gut microbiome and human health. FEMS Microbiol. Rev. 2020, 44, 454–489. [Google Scholar] [CrossRef]
- Wang, L.-C.; Pan, T.-M.; Tsai, T.-Y. Lactic acid bacteria-fermented product of green tea and Houttuynia cordata leaves exerts anti-adipogenic and anti-obesity effects. J. Food Drug Anal. 2018, 26, 973–984. [Google Scholar] [CrossRef]
- Gan, Y.; Tang, M.W.; Tan, F.; Zhou, X.R.; Fan, L.; Xie, Y.X.; Zhao, X. Anti-obesity effect of Lactobacillus plantarum CQPC01 by modulating lipid metabolism in high-fat diet-induced C57BL/6 mice. J. Food Biochem. 2020, 44, e13491. [Google Scholar] [CrossRef]
- Tsai, Y.-T.; Cheng, P.-C.; Pan, T.-M. Anti-obesity effects of gut microbiota are associated with lactic acid bacteria. Appl. Microbiol. Biotechnol. 2014, 98, 1–10. [Google Scholar] [CrossRef]
- Fabersani, E.; Marquez, A.; Russo, M.; Ross, R.; Torres, S.; Fontana, C.; Puglisi, E.; Medina, R.; Gauffin-Cano, P. Lactic Acid Bacteria Strains Differently Modulate Gut Microbiota and Metabolic and Immunological Parameters in High-Fat Diet-Fed Mice. Front. Nutr. 2021, 8, 718564. [Google Scholar] [CrossRef]
- Choi, I.-D.; Kim, S.-H.; Jeong, J.-W.; Lee, D.E.; Huh, C.-S.; Hong, S.S.; Sim, J.-H.; Ahn, Y.-T. Triglyceride-lowering effects of two probiotics, Lactobacillus plantarum KY1032 and Lactobacillus curvatus HY7601, in a rat model of high-fat diet-induced hypertriglyceridemia. J. Microbiol. Biotechnol. 2016, 26, 483–487. [Google Scholar] [CrossRef] [PubMed]
- Jeung, W.H.; Shim, J.-J.; Woo, S.-W.; Sim, J.-H.; Lee, J.-L. Lactobacillus curvatus HY7601 and Lactobacillus plantarum KY1032 cell extracts inhibit adipogenesis in 3T3-L1 and HepG2 cells. J. Med. Food 2018, 21, 876–886. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.; Lee, Y.J.; Kim, M.; Kim, M.; Kwak, J.H.; Lee, J.-W.; Ahn, Y.-T.; Sim, J.-H.; Lee, J.H. Supplementation with two probiotic strains, Lactobacillus curvatus HY7601 and Lactobacillus plantarum KY1032, reduced body adiposity and Lp-PLA2 activity in overweight subjects. J. Funct. Foods 2015, 19, 744–752. [Google Scholar] [CrossRef]
- Kim, M.; Kim, M.; Kang, M.; Yoo, H.J.; Kim, M.S.; Ahn, Y.-T.; Sim, J.-H.; Jee, S.H.; Lee, J.H. Effects of weight loss using supplementation with Lactobacillus strains on body fat and medium-chain acylcarnitines in overweight individuals. Food Funct. 2017, 8, 250–261. [Google Scholar] [CrossRef] [PubMed]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2–approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fijan, S. Microorganisms with claimed probiotic properties: An overview of recent literature. Int. J. Environ. Res. Public Health 2014, 11, 4745–4767. [Google Scholar] [CrossRef] [PubMed]
- Suvorov, A. Gut microbiota, probiotics, and human health. Biosci. Microbiota Food Health 2013, 32, 81–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, D.C. Probiotics as functional foods. Nutr. Clin. Pract. 2003, 18, 497–506. [Google Scholar] [CrossRef]
- Zubillaga, M.; Weill, R.; Postaire, E.; Goldman, C.; Caro, R.; Boccio, J. Effect of probiotics and functional foods and their use in different diseases. Nutr. Res. 2001, 21, 569–579. [Google Scholar] [CrossRef]
- Gadelha, C.J.M.U.; Bezerra, A.N. Effects of probiotics on the lipid profile: Systematic review. J. Vasc. Bras. 2019, 18, e20180124. [Google Scholar] [CrossRef]
- Park, D.-Y.; Ahn, Y.-T.; Park, S.-H.; Huh, C.-S.; Yoo, S.-R.; Yu, R.; Sung, M.-K.; McGregor, R.A.; Choi, M.-S. Supplementation of Lactobacillus curvatus HY7601 and Lactobacillus plantarum KY1032 in diet-induced obese mice is associated with gut microbial changes and reduction in obesity. PLoS ONE 2013, 8, e59470. [Google Scholar] [CrossRef] [Green Version]
- Jeung, W.H.; Nam, W.; Kim, H.J.; Kim, J.Y.; Nam, B.; Jang, S.S.; Lee, J.-L.; Sim, J.-H.; Park, S.-D. Oral administration of Lactobacillus curvatus HY7601 and Lactobacillus plantarum KY1032 with Cinnamomi ramulus extract reduces diet-induced obesity and modulates gut microbiota. Prev. Nutr. Food Sci. 2019, 24, 136. [Google Scholar] [CrossRef]
- Forny-Germano, L.; De Felice, F.G.; Vieira, M.N.D.N. The role of leptin and adiponectin in obesity-associated cognitive decline and Alzheimer’s disease. Front. Neurosci. 2019, 12, 1027. [Google Scholar] [CrossRef] [Green Version]
- Cambuli, V.M.; Musiu, M.C.; Incani, M.; Paderi, M.; Serpe, R.; Marras, V.; Cossu, E.; Cavallo, M.G.; Mariotti, S.; Loche, S. Assessment of adiponectin and leptin as biomarkers of positive metabolic outcomes after lifestyle intervention in overweight and obese children. J. Clin. Endocrinol. Metab. 2008, 93, 3051–3057. [Google Scholar] [CrossRef] [Green Version]
- Heiss, C.N.; Olofsson, L.E. Gut microbiota-dependent modulation of energy metabolism. J. Innate Immun. 2018, 10, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Al Maskari, M.Y.; Alnaqdy, A.A. Correlation between serum leptin levels, body mass index and obesity in Omanis. Sultan Qaboos Univ. Med. J. 2006, 6, 27. [Google Scholar] [PubMed]
- Zhao, S.; Zhu, Y.; Schultz, R.D.; Li, N.; He, Z.; Zhang, Z.; Caron, A.; Zhu, Q.; Sun, K.; Xiong, W. Partial leptin reduction as an insulin sensitization and weight loss strategy. Cell Metab. 2019, 30, 706–719. [Google Scholar] [CrossRef] [PubMed]
- Yosaee, S.; Khodadost, M.; Esteghamati, A.; Speakman, J.R.; Djafarian, K.; Bitarafan, V.; Shidfar, F. Adiponectin: An Indicator for Metabolic Syndrome. Iran J. Public Health 2019, 48, 1106–1115. [Google Scholar] [CrossRef]
- Achari, A.E.; Jain, S.K. Adiponectin, a therapeutic target for obesity, diabetes, and endothelial dysfunction. Int. J. Mol. Sci. 2017, 18, 1321. [Google Scholar] [CrossRef] [Green Version]
- Balsan, G.A.; Vieira, J.L.d.C.; Oliveira, A.M.d.; Portal, V.L. Relationship between adiponectin, obesity and insulin resistance. Rev. Da Assoc. Médica Bras. 2015, 61, 72–80. [Google Scholar] [CrossRef]
- Nigro, E.; Scudiero, O.; Monaco, M.L.; Palmieri, A.; Mazzarella, G.; Costagliola, C.; Bianco, A.; Daniele, A. New insight into adiponectin role in obesity and obesity-related diseases. BioMed Res. Int. 2014, 2014, 658913. [Google Scholar] [CrossRef]
- De Rosa, A.; Monaco, M.L.; Capasso, M.; Forestieri, P.; Pilone, V.; Nardelli, C.; Buono, P.; Daniele, A. Adiponectin oligomers as potential indicators of adipose tissue improvement in obese subjects. Eur. J. Endocrinol. 2013, 169, 37–43. [Google Scholar] [CrossRef] [Green Version]
- Daniele, A.; Cammarata, R.; Masullo, M.; Nerone, G.; Finamore, F.; D’andrea, M.; Pilla, F.; Oriani, G. Analysis of adiponectin gene and comparison of its expression in two different pig breeds. Obesity 2008, 16, 1869–1874. [Google Scholar] [CrossRef]
- Muscogiuri, G.; Cantone, E.; Cassarano, S.; Tuccinardi, D.; Barrea, L.; Savastano, S.; Colao, A. Gut microbiota: A new path to treat obesity. Int. J. Obes. Suppl. 2019, 9, 10–19. [Google Scholar] [CrossRef]
- Geng, J.; Ni, Q.; Sun, W.; Li, L.; Feng, X. The links between gut microbiota and obesity and obesity related diseases. Biomed. Pharmacother. 2022, 147, 112678. [Google Scholar] [CrossRef] [PubMed]
- Jeelani, A.; Ahmed, S.A.; Momin, F.V. Obesity-Caused by a germ. Int. J. Sci. Res. Publ. 2013, 3, 1–3. [Google Scholar]
- Cuevas-Sierra, A.; Riezu-Boj, J.I.; Guruceaga, E.; Milagro, F.I.; Martínez, J.A. Sex-specific associations between gut Prevotellaceae and host genetics on adiposity. Microorganisms 2020, 8, 938. [Google Scholar] [CrossRef] [PubMed]
- Harakeh, S.M.; Khan, I.; Kumosani, T.; Barbour, E.; Almasaudi, S.B.; Bahijri, S.M.; Alfadul, S.M.; Ajabnoor, G.; Azhar, E.I. Gut microbiota: A contributing factor to obesity. Front. Cell. Infect. Microbiol. 2016, 6, 95. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.-M.; Kim, D.-H. Bifidobacterium adolescentis IM38 ameliorates high-fat diet–induced colitis in mice by inhibiting NF-κB activation and lipopolysaccharide production by gut microbiota. Nutr. Res. 2017, 41, 86–96. [Google Scholar] [CrossRef]
- An, H.M.; Park, S.Y.; Lee, D.K.; Kim, J.R.; Cha, M.K.; Lee, S.W.; Lim, H.T.; Kim, K.J.; Ha, N.J. Antiobesity and lipid-lowering effects of Bifidobacterium spp. in high fat diet-induced obese rats. Lipids Health Dis. 2011, 10, 116. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Wang, N.; Tan, H.-Y.; Li, S.; Zhang, C.; Feng, Y. Function of Akkermansia muciniphila in obesity: Interactions with lipid metabolism, immune response and gut systems. Front. Microbiol. 2020, 11, 219. [Google Scholar] [CrossRef] [Green Version]
- Balamurugan, R.; George, G.; Kabeerdoss, J.; Hepsiba, J.; Chandragunasekaran, A.M.; Ramakrishna, B.S. Quantitative differences in intestinal Faecalibacterium prausnitzii in obese Indian children. Br. J. Nutr. 2010, 103, 335–338. [Google Scholar] [CrossRef] [Green Version]
- Duan, M.; Wang, Y.; Zhang, Q.; Zou, R.; Guo, M.; Zheng, H. Characteristics of gut microbiota in people with obesity. PLoS ONE 2021, 16, e0255446. [Google Scholar] [CrossRef]
- Companys, J.; Gosalbes, M.J.; Pla-Pagà, L.; Calderón-Pérez, L.; Llauradó, E.; Pedret, A.; Valls, R.M.; Jiménez-Hernández, N.; Sandoval-Ramirez, B.A.; Del Bas, J.M. Gut microbiota profile and its association with clinical variables and dietary intake in overweight/obese and lean subjects: A cross-sectional study. Nutrients 2021, 13, 2032. [Google Scholar] [CrossRef]
- Chung, H.; Kim, Y.; Chun, S.; Ji, G.E. Screening and selection of acid and bile resistant bifidobacteria. Int. J. Food Microbiol. 1999, 47, 25–32. [Google Scholar] [CrossRef]
- Jung, S.H.; Hong, D.K.; Bang, S.-J.; Heo, K.; Sim, J.-J.; Lee, J.-L. The functional properties of Lactobacillus casei hy2782 are affected by the fermentation time. Appl. Sci. 2021, 11, 2481. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Placebo (n = 29) | Probiotics (n = 30) | p-Value |
|---|---|---|---|
| Gender (n, female/male) | 8/21 | 5/25 | 0.486 |
| Smoker (Y/N) | 9/20 | 7/23 | 0.710 |
| Drinker (Y/N) | 21/8 | 25/5 | 0.486 |
| Age (years) | 39.34 ± 1.61 | 35.7 ± 1.44 | 0.096 |
| Height (m) | 171.62 ± 1.69 | 171.32 ± 1.34 | 0.891 |
| Weight (kg) | 79.07 ± 1.85 | 79.21 ± 2.19 | 0.961 |
| BMI (kg/m2) | 26.81 ± 0.47 | 26.87 ± 0.52 | 0.880 |
| Waist circumference (cm) | 94.92 ± 1.32 | 93.99 ± 1.88 | 0.217 |
| Hip circumference (cm) | 101.97 ± 0.83 | 101.58 ± 0.93 | 0.752 |
| Percent body fat (%) | 30.17 ± 1.22 | 27.53 ± 1.34 | 0.151 |
| Body fat mass (kg) | 23.66 ± 0.95 | 21.86 ± 1.3 | 0.268 |
| Lean body mass (kg) | 55.41 ± 1.78 | 57.35 ± 1.77 | 0.443 |
| Visceral fat area (cm2) | 107.98 ± 5.49 | 97 ± 6.79 | 0.085 |
| SBP (mmHg) | 135.93 ± 1.66 | 132.47 ± 2.16 | 0.210 |
| DBP (mmHg) | 132.66 ± 2.09 | 131.27 ± 2.28 | 0.656 |
| HR (beats/min) | 83.41 ± 1.98 | 80.1 ± 1.61 | 0.079 |
| Total cholesterol (mg/dL) | 200 ± 5.28 | 215.93 ± 6.47 | 0.161 |
| HDL-cholesterol (mg/dL) | 53.45 ± 2.15 | 52.5 ± 2.59 | 0.462 |
| LDL-cholesterol (mg/dL) | 132.31 ± 6.79 | 148.8 ± 6.47 | 0.084 |
| Triglyceride (mg/dL) | 137.34 ± 11.02 | 140.87 ± 16.48 | 0.688 |
| Insulin (μIU/mL) | 7.08 ± 0.83 | 6.16 ± 0.6 | 0.400 |
| Leptin (ng/mL) | 19.13 ± 12.38 | 15.31 ± 2.08 | 0.084 |
| Adiponectin (ng/mL) | 2741.52 ± 306.7 | 2222.47 ± 214.29 | 0.240 |
| hs-CRP (mg/dL) | 0.99 ± 0.28 | 1.62 ± 0.59 | 0.103 |
| Variables | Placebo (n = 29) | Probiotics (n = 30) | p-Value | ||||
|---|---|---|---|---|---|---|---|
| Baseline | 12 Weeks | Change | Baseline | 12 Weeks | Change | ||
| Body weight (kg) | 79.07 ± 1.85 | 79.99 ± 1.95 | 0.93 ± 0.28 | 79.21 ± 2.19 | 78.74 ± 2.12 | −0.47 ± 0.29 | 0.001 T |
| BMI (kg/m2) | 26.81 ± 0.47 | 27.13 ± 0.53 | 0.32 ± 0.1 | 26.87 ± 0.52 | 26.73 ± 0.51 | −0.15 ± 0.1 | <0.001 T |
| Waist circumference (cm) | 94.92 ± 1.32 | 96.23 ± 1.52 | 1.31 ± 0.5 | 93.99 ± 1.88 | 93.58 ± 1.93 | −0.41 ± 0.37 | 0.007 T |
| Hip circumference (cm) | 101.97 ± 0.83 | 101.59 ± 0.89 | −0.38 ± 0.44 | 101.58 ± 1.93 | 101.08 ± 0.89 | −0.5 ± 0.21 | 0.062 M |
| Percent body fat (%) | 30.17 ± 1.22 | 30.24 ± 1.3 | 0.08 ± 0.2 | 27.53 ± 1.34 | 27.29 ± 1.35 | −0.23 ± 0.28 | 0.366 T |
| Body fat mass (kg) | 23.66 ± 0.95 | 24.05 ± 1.11 | 0.39 ± 0.22 | 21.86 ± 1.3 | 21.58 ± 1.31 | −0.28 ± 0.24 | 0.043 T |
| Lean body mass (kg) | 55.41 ± 1.78 | 55.92 ± 1.81 | 0.51 ± 0.2 | 57.35 ± 1.77 | 57.16 ± 1.72 | −0.18 ± 0.25 | 0.032 T |
| Visceral fat area (cm2) | 107.98 ± 5.49 | 110.81 ± 6.1 | 2.82 ± 1.33 | 97 ± 6.8 | 95.34 ± 7 | −1.67 ± 1.44 | 0.025 T |
| Variables | Placebo (n = 29) | Probiotics (n = 30) | p-Value | ||||
|---|---|---|---|---|---|---|---|
| Baseline | 12 Weeks | Change | Baseline | 12 Weeks | Change | ||
| Total cholesterol (mg/dL) | 200 ± 5.28 | 203.69 ± 6.07 | 3.69 ± 3.83 | 215.93 ± 6.47 | 215.33 ± 7.81 | −0.6 ± 5.04 | 0.501 M |
| HDL-cholesterol (mg/dL) | 53.45 ± 2.15 | 52 ± 1.85 | −1.45 ± 1.15 | 52.5 ± 2.59 | 52.1 ± 2.93 | −0.4 ± 1.18 | 0.527 T |
| LDL-cholesterol (mg/dL) | 132.31 ± 6.79 | 146.1 ± 7.13 | 13.79 ± 4.14 | 148.8 ± 6.47 | 153.3 ± 8.03 | 4.5 ± 4.59 | 0.139 T |
| Triglyceride (mg/dL) | 137.34 ± 11.02 | 145.59 ± 12.25 | 8.24 ± 9.99 | 140.87 ± 16.48 | 157.93 ± 17.66 | 17.07 ± 11.4 | 0.563 M |
| Insulin (μIU/mL) | 7.08 ± 0.83 | 6.32 ± 0.52 | −0.77 ± 0.79 | 6.16 ± 0.6 | 8.9 ± 3.66 | 2.74 ± 3.55 | 0.934 M |
| Leptin (ng/mL) | 19.13 ± 2.38 | 21.83 ± 2.7 | 2.7 ± 1.59 | 15.31 ± 20.8 | 14.55 ± 1.82 | −0.76 ± 0.88 | 0.117 M |
| Adiponectin (ng/mL) | 2741.52 ± 306.7 | 2494.69 ± 265.53 | −246.83 ± 178.68 | 2222.47 ± 214.29 | 2355.8 ± 190.75 | 133.33 ± 150.47 | 0.046 M |
| hs-CRP | 0.99 ± 0.28 | 0.78 ± 0.18 | −0.21 ± 0.2 | 1.62 ± 0.59 | 4.74 ± 3.96 | 3.13 ± 3.63 | 0.619 M |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mo, S.-J.; Lee, K.; Hong, H.-J.; Hong, D.-K.; Jung, S.-H.; Park, S.-D.; Shim, J.-J.; Lee, J.-L. Effects of Lactobacillus curvatus HY7601 and Lactobacillus plantarum KY1032 on Overweight and the Gut Microbiota in Humans: Randomized, Double-Blinded, Placebo-Controlled Clinical Trial. Nutrients 2022, 14, 2484. https://doi.org/10.3390/nu14122484
Mo S-J, Lee K, Hong H-J, Hong D-K, Jung S-H, Park S-D, Shim J-J, Lee J-L. Effects of Lactobacillus curvatus HY7601 and Lactobacillus plantarum KY1032 on Overweight and the Gut Microbiota in Humans: Randomized, Double-Blinded, Placebo-Controlled Clinical Trial. Nutrients. 2022; 14(12):2484. https://doi.org/10.3390/nu14122484
Chicago/Turabian StyleMo, Sung-Joon, Kippeum Lee, Hyoung-Ju Hong, Dong-Ki Hong, Seung-Hee Jung, Soo-Dong Park, Jae-Jung Shim, and Jung-Lyoul Lee. 2022. "Effects of Lactobacillus curvatus HY7601 and Lactobacillus plantarum KY1032 on Overweight and the Gut Microbiota in Humans: Randomized, Double-Blinded, Placebo-Controlled Clinical Trial" Nutrients 14, no. 12: 2484. https://doi.org/10.3390/nu14122484
APA StyleMo, S.-J., Lee, K., Hong, H.-J., Hong, D.-K., Jung, S.-H., Park, S.-D., Shim, J.-J., & Lee, J.-L. (2022). Effects of Lactobacillus curvatus HY7601 and Lactobacillus plantarum KY1032 on Overweight and the Gut Microbiota in Humans: Randomized, Double-Blinded, Placebo-Controlled Clinical Trial. Nutrients, 14(12), 2484. https://doi.org/10.3390/nu14122484