
Estrogen Receptor Knockout Mice and Their Effects on Fertility



Abstract
1. Introduction
2. Estrogen Receptors
2.1. Overview
2.2. Genes
2.3. Expression Patterns of Esr1 and Esr2
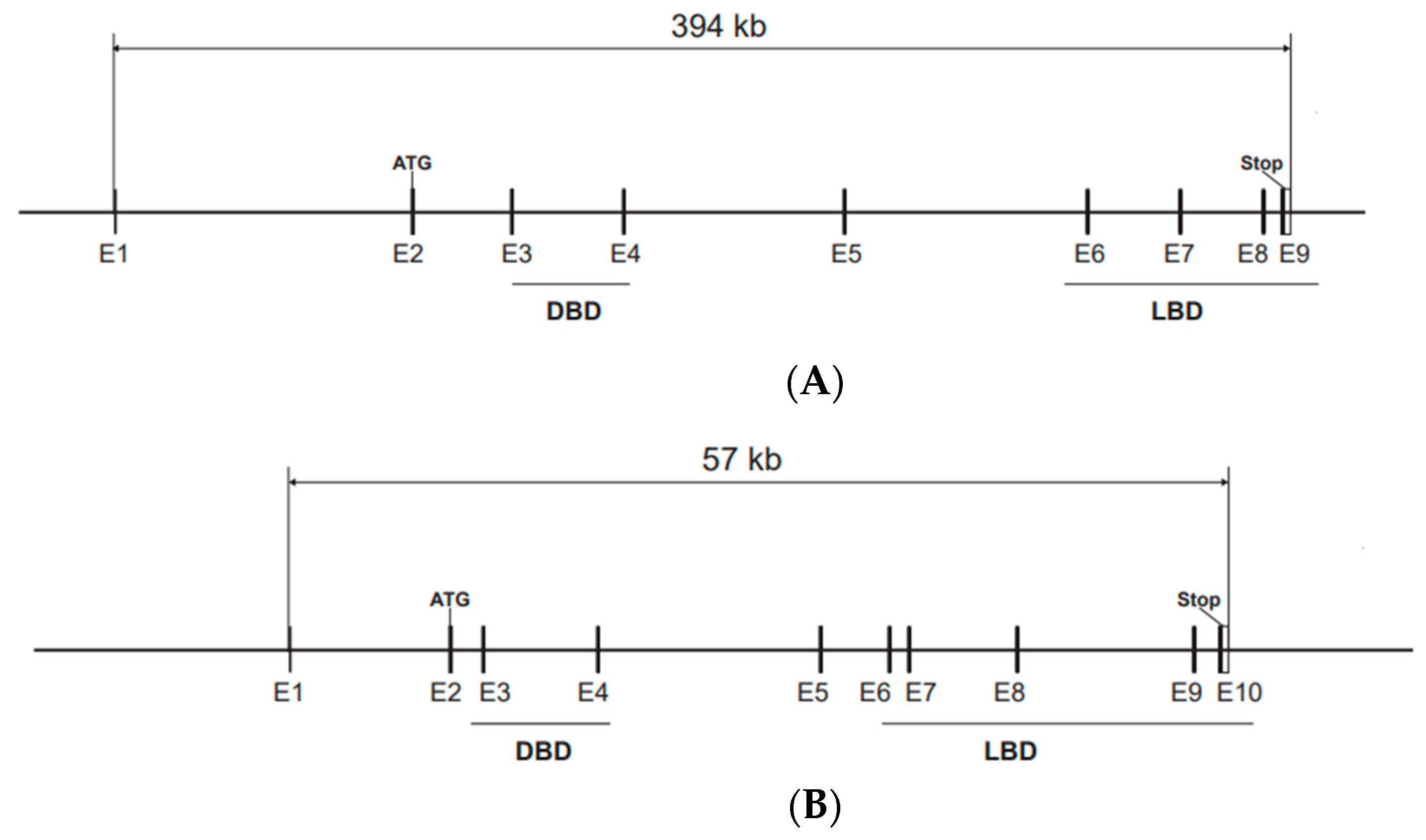
3. Animal Models
3.1. Esr1 Mouse Models

{kind=link}
| Mouse Line | Mutation | Cell Type/Effect | Fertility | References |
|---|---|---|---|---|
| ERKO | Neo insertion into | Global KO | F: infertile | [28] |
| Esr1 exon 3 | M: reduced | |||
| ERαKO, Ex3αERKO | Deletion of Esr1 exon 3 | Global KO | F: infertile | [29,30,31,32] |
| ERα−/− | M: infertile | |||
| ERαfl/fl;CamKIIα-Cre | Deletion of Esr1 exon 3 | Neurons | F: infertile | [37] |
| aP2-CreERαfl/fl | Deletion of Esr1 exon 3 | Adipocytes and | F: infertile | [38] |
| hypothalamus | M: fertile | |||
| ERαflox/flox αGSUcre | Deletion of Esr1 exon 3 | pituitary gonadotroph | F: infertile | [39] |
| UtEpiαERKO | Deletion of Esr1 exon 3 | uterine epithelial cells | F: infertile | [40] |
| WEd/d(Wnt7a-Cre) | Deletion of Esr1 exon 3 | uterine cells | F: infertile | [41] |
| Amhr2Cre/+;Esr1f/− | Deletion of Esr1 exon 3 | uterine stromal cells | F: reduced | [42] |
| NERKI+/− | E207A/G208A in ERα | no DNA binding, DN | F: infertile | [43] |
| M: fertile | ||||
| ERα(EAAE) | Y201E/K210A/K214A/R215E | no DNA binding | F: infertile | [44] |
| in ERα | M: infertile | |||
| ENERKI | G525L in ERα | no E2 binding | F: infertile | [45] |
| M: sub-fertile | ||||
| AF2ERKI | L543A/L544A in ERα | no E2 binding | F: infertile | [46] |
| M: fertile | ||||
| ERα C451A, NOER | C451A in ERα | no membrane ERα | F: infertile | [47,48,49] |
| M: sub-fertile | ||||
| ESR1Y541S/βActin-Cre | Y541S, inducible | constitutively active | F: infertile | [50] |
| M: infertile | ||||
| ERβ−/− | Neo insertion into | Global | F: reduced | [51] |
| Esr2 exon 3 | M: fertile | |||
| ErβKO | Replacement of Esr2 exon 3 | Global | F: reduced | [29] |
| with Neo | M: fertile | |||
| ERβSTL−/L−, Ex3βERKO | Deletion of Esr1 exon 3 | Global | F: reduced * | [52,53,54,55] |
| ERβ-Δex3, CERβKO | M: fertile * | |||
| Esr2ΔE1-10 | Deletion of Esr2 exon 1–10 | Global | F: sub-fertile | [24] |
| M: fertile | ||||
| Esr2Y55F/Y55F | Y55F in ERβ | reduced activity | F: reduced | [56] |
| αβERKO | Esr1/Esr2 double knockout | Global | F: infertile | [57] |
| ERKO × ERβ−/− | M: infertile | |||
| ERαβKO | Esr1/Esr2 double knockout | Global | F: infertile | [29] |
| ERαKO × ERβKO | M: infertile |
3.2. Esr2 Mouse Models
3.3. Esr1/Esr2 Double Knockouts
3.4. Rat Esr1 and Esr2 Knockout Models
4. Human Mutations in ESR1 and ESR2
5. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sauer, B.; Henderson, N. Site-Specific DNA Recombination in Mammalian Cells by the Cre Recombinase of Bacteriophage P1. Proc. Natl. Acad. Sci. USA 1988, 85, 5166–5170. [Google Scholar] [CrossRef] [PubMed]
- Ran, F.A.; Hsu, P.D.; Wright, J.; Agarwala, V.; Scott, D.A.; Zhang, F. Genome Engineering Using the Crispr-Cas9 System. Nat. Protoc. 2013, 8, 2281–2308. [Google Scholar] [CrossRef] [PubMed]
- Eyster, K.M. The Estrogen Receptors: An Overview from Different Perspectives. Methods Mol. Biol. 2016, 1366, 1–10. [Google Scholar] [CrossRef]
- Prossnitz, E.R.; Hathaway, H.J. What Have We Learned About Gper Function in Physiology and Disease from Knockout Mice? J. Steroid Biochem. Mol. Biol. 2015, 153, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Walter, P.; Green, S.; Greene, G.; Krust, A.; Bornert, J.M.; Jeltsch, J.M.; Staub, A.; Jensen, E.; Scrace, G.; Waterfield, M. Cloning of the Human Estrogen Receptor Cdna. Proc. Natl. Acad. Sci. USA 1985, 82, 7889–7893. [Google Scholar] [CrossRef]
- Kuiper, G.G.; Enmark, E.; Pelto-Huikko, M.; Nilsson, S.; Gustafsson, J.A. Cloning of a Novel Receptor Expressed in Rat Prostate and Ovary. Proc. Natl. Acad. Sci. USA 1996, 93, 5925–5930. [Google Scholar] [CrossRef]
- Mangelsdorf, D.J.; Thummel, C.; Beato, M.; Herrlich, P.; Schütz, G.; Umesono, K.; Blumberg, B.; Kastner, P.; Mark, M.; Chambon, P.; et al. The Nuclear Receptor Superfamily: The Second Decade. Cell 1995, 83, 835–839. [Google Scholar] [CrossRef] [PubMed]
- Rosenfeld, M.G.; Lunyak, V.V.; Glass, C.K. Sensors and Signals: A Coactivator/Corepressor/Epigenetic Code for Integrating Signal-Dependent Programs of Transcriptional Response. Genes Dev. 2006, 20, 1405–1428. [Google Scholar] [CrossRef]
- Gronemeyer, H.; Gustafsson, J.; Laudet, V. Principles for Modulation of the Nuclear Receptor Superfamily. Nat. Rev. Drug Discov. 2004, 3, 950–964. [Google Scholar] [CrossRef]
- Heldring, N.; Pike, A.; Andersson, S.; Matthews, J.; Cheng, G.; Hartman, J.; Tujague, M.; Strom, A.; Treuter, E.; Warner, M.; et al. Estrogen Receptors: How Do They Signal and What Are Their Targets. Physiol. Rev. 2007, 87, 905–931. [Google Scholar] [CrossRef] [PubMed]
- Kuiper, G.G.; Carlsson, B.; Grandien, K.; Enmark, E.; Haggblad, J.; Nilsson, S.; Gustafsson, J.A. Comparison of the Ligand Binding Specificity and Transcript Tissue Distribution of Estrogen Receptors Alpha and Beta. Endocrinology 1997, 138, 863–870. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Burch, P.E.; Cooney, A.J.; Lanz, R.B.; Pereira, F.A.; Wu, J.; Gibbs, R.A.; Weinstock, G.; Wheeler, D.A. Genomic Analysis of the Nuclear Receptor Family: New Insights into Structure, Regulation, and Evolution from the Rat Genome. Genome Res. 2004, 14, 580–590. [Google Scholar] [CrossRef] [PubMed]
- Kos, M.; O’Brien, S.; Flouriot, G.; Gannon, F. Tissue-Specific Expression of Multiple Mrna Variants of the Mouse Estrogen Receptor Alpha Gene. FEBS Lett. 2000, 477, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Lewandowski, S.; Kalita, K.; Kaczmarek, L. Estrogen Receptor Beta. Potential Functional Significance of a Variety of Mrna Isoforms. FEBS Lett. 2002, 524, 1–5. [Google Scholar] [CrossRef]
- Ishii, H.; Sakuma, Y. Complex Organization of the 5′-Untranslated Region of the Mouse Estrogen Receptor Alpha Gene: Identification of Numerous Mrna Transcripts with Distinct 5′-Ends. J. Steroid Biochem. Mol. Biol. 2011, 125, 211–218. [Google Scholar] [CrossRef]
- Lu, B.; Leygue, E.; Dotzlaw, H.; Murphy, L.C.; Watson, P. Estrogen Receptor-Beta Mrna Variants in Human and Murine Tissues. Mol Cell. Endocrinol. 1998, 138, 199–203. [Google Scholar] [CrossRef]
- Maruyama, K.; Endoh, H.; Sasaki-Iwaoka, H.; Kanou, H.; Shimaya, E.; Hashimoto, S.; Kato, S.; Kawashima, H. A Novel Isoform of Rat Estrogen Receptor Beta with 18 Amino Acid Insertion in the Ligand Binding Domain as a Putative Dominant Negative Regular of Estrogen Action. Biochem. Biophys. Res. Commun. 1998, 246, 142–147. [Google Scholar] [CrossRef]
- Zhao, C.; Toresson, G.; Xu, L.; Koehler, K.F.; Gustafsson, J.A.; Dahlman-Wright, K. Mouse Estrogen Receptor Beta Isoforms Exhibit Differences in Ligand Selectivity and Coactivator Recruitment. Biochemistry 2005, 44, 7936–7944. [Google Scholar] [CrossRef]
- Ogawa, S.; Inoue, S.; Watanabe, T.; Orimo, A.; Hosoi, T.; Ouchi, Y.; Muramatsu, M. Molecular Cloning and Characterization of Human Estrogen Receptor Betacx: A Potential Inhibitor Ofestrogen Action in Human. Nucleic Acids Res. 1998, 26, 3505–3512. [Google Scholar] [CrossRef] [PubMed]
- Couse, J.F.; Lindzey, J.; Grandien, K.; Gustafsson, J.A.; Korach, K.S. Tissue Distribution and Quantitative Analysis of Estrogen Receptor-Alpha (Eralpha) and Estrogen Receptor-Beta (Erbeta) Messenger Ribonucleic Acid in the Wild-Type and Eralpha-Knockout Mouse. Endocrinology 1997, 138, 4613–4621. [Google Scholar] [CrossRef]
- Irsik, D.L.; Carmines, P.K.; Lane, P.H. Classical Estrogen Receptors and Eralpha Splice Variants in the Mouse. PLoS ONE 2013, 8, e70926. [Google Scholar] [CrossRef]
- Andersson, S.; Sundberg, M.; Pristovsek, N.; Ibrahim, A.; Jonsson, P.; Katona, B.; Clausson, C.M.; Zieba, A.; Ramstrom, M.; Soderberg, O.; et al. Insufficient Antibody Validation Challenges Oestrogen Receptor Beta Research. Nat. Commun. 2017, 8, 15840. [Google Scholar] [CrossRef] [PubMed]
- Ishii, H.; Otsuka, M.; Kanaya, M.; Higo, S.; Hattori, Y.; Ozawa, H. Applicability of Anti-Human Estrogen Receptor Beta Antibody Ppz0506 for the Immunodetection of Rodent Estrogen Receptor Beta Proteins. Int. J. Mol. Sci. 2019, 20, 6312. [Google Scholar] [CrossRef]
- Antonson, P.; Apolinario, L.M.; Shamekh, M.M.; Humire, P.; Poutanen, M.; Ohlsson, C.; Nalvarte, I.; Gustafsson, J.A. Generation of an All-Exon Esr2 Deleted Mouse Line: Effects on Fertility. Biochem. Biophys. Res. Commun. 2020, 529, 231–237. [Google Scholar] [CrossRef]
- Ozawa, M.; Hattori, Y.; Higo, S.; Otsuka, M.; Matsumoto, K.; Ozawa, H.; Ishii, H. Optimized Mouse-on-Mouse Immunohistochemical Detection of Mouse Esr2 Proteins with Ppz0506 Monoclonal Antibody. Acta Histochem. Cytochem. 2022, 55, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Schröder, S.K.; Tag, C.G.; Kessel, J.C.; Antonson, P.; Weiskirchen, R. Immunohistochemical Detection of Estrogen Receptor-Beta (ErΒ) with Ppz0506 Antibody in Murine Tissue: From Pitfalls to Optimization. Biomedicines 2022, 10, 3100. [Google Scholar] [PubMed]
- Maioli, S.; Leander, K.; Nilsson, P.; Nalvarte, I. Estrogen Receptors and the Aging Brain. Essays Biochem. 2021, 65, 913–925. [Google Scholar] [CrossRef]
- Lubahn, D.B.; Moyer, J.S.; Golding, T.S.; Couse, J.F.; Korach, K.S.; Smithies, O. Alteration of Reproductive Function but Not Prenatal Sexual Development after Insertional Disruption of the Mouse Estrogen Receptor Gene. Proc. Natl. Acad. Sci. USA 1993, 90, 11162–11166. [Google Scholar] [CrossRef]
- Dupont, S.; Krust, A.; Gansmuller, A.; Dierich, A.; Chambon, P.; Mark, M. Effect of Single and Compound Knockouts of Estrogen Receptors Alpha (Eralpha) and Beta (Erbeta) on Mouse Reproductive Phenotypes. Development 2000, 127, 4277–4291. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Wolfe, A.; Wang, X.; Chang, C.; Yeh, S.; Radovick, S. Generation and Characterization of a Complete Null Estrogen Receptor Alpha Mouse Using Cre/Loxp Technology. Mol. Cell Biochem. 2009, 321, 145–153. [Google Scholar] [CrossRef]
- Hewitt, S.C.; Kissling, G.E.; Fieselman, K.E.; Jayes, F.L.; Gerrish, K.E.; Korach, K.S. Biological and Biochemical Consequences of Global Deletion of Exon 3 from the Er Alpha Gene. FASEB J. 2010, 24, 4660–4667. [Google Scholar] [CrossRef]
- Antonson, P.; Omoto, Y.; Humire, P.; Gustafsson, J.A. Generation of Eralpha-Floxed and Knockout Mice Using the Cre/Loxp System. Biochem. Biophys. Res. Commun. 2012, 424, 710–716. [Google Scholar] [CrossRef] [PubMed]
- Eddy, E.M.; Washburn, T.F.; Bunch, D.O.; Goulding, E.H.; Gladen, B.C.; Lubahn, D.B.; Korach, K.S. Targeted Disruption of the Estrogen Receptor Gene in Male Mice Causes Alteration of Spermatogenesis and Infertility. Endocrinology 1996, 137, 4796–4805. [Google Scholar] [CrossRef]
- Hess, R.A.; Bunick, D.; Lee, K.H.; Bahr, J.; Taylor, J.A.; Korach, K.S.; Lubahn, D.B. A Role for Oestrogens in the Male Reproductive System. Nature 1997, 390, 509–512. [Google Scholar] [CrossRef]
- Hess, R.A.; Cooke, P.S. Estrogen in the Male: A Historical Perspective. Biol. Reprod. 2018, 99, 27–44. [Google Scholar] [CrossRef] [PubMed]
- Curtis Hewitt, S.; Couse, J.F.; Korach, K.S. Estrogen Receptor Transcription and Transactivation: Estrogen Receptor Knockout Mice: What Their Phenotypes Reveal About Mechanisms of Estrogen Action. Breast Cancer Res. 2000, 2, 345–352. [Google Scholar] [CrossRef]
- Wintermantel, T.M.; Campbell, R.E.; Porteous, R.; Bock, D.; Grone, H.J.; Todman, M.G.; Korach, K.S.; Greiner, E.; Perez, C.A.; Schutz, G.; et al. Definition of Estrogen Receptor Pathway Critical for Estrogen Positive Feedback to Gonadotropin-Releasing Hormone Neurons and Fertility. Neuron 2006, 52, 271–280. [Google Scholar] [CrossRef]
- Antonson, P.; Matic, M.; Portwood, N.; Kuiper, R.V.; Bryzgalova, G.; Gao, H.; Windahl, S.H.; Humire, P.; Ohlsson, C.; Berggren, P.-O.; et al. Ap2-Cre-Mediated Inactivation of Estrogen Receptor Alpha Causes Hydrometra. PLoS ONE 2014, 9, e85581. [Google Scholar] [CrossRef]
- Gieske, M.C.; Kim, H.J.; Legan, S.J.; Koo, Y.; Krust, A.; Chambon, P.; Ko, C. Pituitary Gonadotroph Estrogen Receptor-Alpha Is Necessary for Fertility in Females. Endocrinology 2008, 149, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Winuthayanon, W.; Hewitt, S.C.; Orvis, G.D.; Behringer, R.R.; Korach, K.S. Uterine Epithelial Estrogen Receptor Alpha Is Dispensable for Proliferation but Essential for Complete Biological and Biochemical Responses. Proc. Natl. Acad. Sci. USA 2010, 107, 19272–19277. [Google Scholar] [CrossRef]
- Pawar, S.; Laws, M.J.; Bagchi, I.C.; Bagchi, M.K. Uterine Epithelial Estrogen Receptor-Alpha Controls Decidualization Via a Paracrine Mechanism. Mol. Endocrinol. 2015, 29, 1362–1374. [Google Scholar] [CrossRef]
- Winuthayanon, W.; Lierz, S.L.; Delarosa, K.C.; Sampels, S.R.; Donoghue, L.J.; Hewitt, S.C.; Korach, K.S. Juxtacrine Activity of Estrogen Receptor Alpha in Uterine Stromal Cells Is Necessary for Estrogen-Induced Epithelial Cell Proliferation. Sci. Rep. 2017, 7, 8377. [Google Scholar] [CrossRef] [PubMed]
- Jakacka, M.; Ito, M.; Martinson, F.; Ishikawa, T.; Lee, E.J.; Jameson, J.L. An Estrogen Receptor (Er)Alpha Deoxyribonucleic Acid-Binding Domain Knock-in Mutation Provides Evidence for Nonclassical Er Pathway Signaling in Vivo. Mol. Endocrinol. 2002, 16, 2188–2201. [Google Scholar] [CrossRef] [PubMed]
- Ahlbory-Dieker, D.L.; Stride, B.D.; Leder, G.; Schkoldow, J.; Trolenberg, S.; Seidel, H.; Otto, C.; Sommer, A.; Parker, M.G.; Schutz, G.; et al. DNA Binding by Estrogen Receptor-Alpha Is Essential for the Transcriptional Response to Estrogen in the Liver and the Uterus. Mol. Endocrinol. 2009, 23, 1544–1555. [Google Scholar] [CrossRef]
- Sinkevicius, K.W.; Burdette, J.E.; Woloszyn, K.; Hewitt, S.C.; Hamilton, K.; Sugg, S.L.; Temple, K.A.; Wondisford, F.E.; Korach, K.S.; Woodruff, T.K.; et al. An Estrogen Receptor-Alpha Knock-in Mutation Provides Evidence of Ligand-Independent Signaling and Allows Modulation of Ligand-Induced Pathways in Vivo. Endocrinology 2008, 149, 2970–2979. [Google Scholar] [CrossRef]
- Arao, Y.; Hamilton, K.J.; Ray, M.K.; Scott, G.; Mishina, Y.; Korach, K.S. Estrogen Receptor Alpha Af-2 Mutation Results in Antagonist Reversal and Reveals Tissue Selective Function of Estrogen Receptor Modulators. Proc. Natl. Acad. Sci. USA 2011, 108, 14986–14991. [Google Scholar] [CrossRef]
- Adlanmerini, M.; Solinhac, R.; Abot, A.; Fabre, A.; Raymond-Letron, I.; Guihot, A.-L.; Boudou, F.; Sautier, L.; Vessières, E.; Kim, S.H.; et al. Mutation of the Palmitoylation Site of Estrogen Receptor Alpha in Vivo Reveals Tissue-Specific Roles for Membrane Versus Nuclear Actions. Proc. Natl. Acad. Sci. USA 2014, 111, E283–E290. [Google Scholar] [CrossRef] [PubMed]
- Pedram, A.; Razandi, M.; Lewis, M.; Hammes, S.; Levin, E.R. Membrane-Localized Estrogen Receptor Alpha Is Required for Normal Organ Development and Function. Dev. Cell 2014, 29, 482–490. [Google Scholar] [CrossRef]
- Nanjappa, M.K.; Hess, R.A.; Medrano, T.I.; Locker, S.H.; Levin, E.R.; Cooke, P.S. Membrane-Localized Estrogen Receptor 1 Is Required for Normal Male Reproductive Development and Function in Mice. Endocrinology 2016, 157, 2909–2919. [Google Scholar] [CrossRef]
- Simond, A.M.; Ling, C.; Moore, M.J.; Condotta, S.A.; Richer, M.J.; Muller, W.J. Point-Activated Esr1(Y541s) Has a Dramatic Effect on the Development of Sexually Dimorphic Organs. Genes Dev. 2020, 34, 1304–1309. [Google Scholar] [CrossRef]
- Krege, J.H.; Hodgin, J.B.; Couse, J.F.; Enmark, E.; Warner, M.; Mahler, J.F.; Sar, M.; Korach, K.S.; Gustafsson, J.A.; Smithies, O. Generation and Reproductive Phenotypes of Mice Lacking Estrogen Receptor Beta. Proc. Natl. Acad. Sci. USA 1998, 95, 15677–15682. [Google Scholar] [CrossRef] [PubMed]
- Antal, M.C.; Krust, A.; Chambon, P.; Mark, M. Sterility and Absence of Histopathological Defects in Nonreproductive Organs of a Mouse Erbeta-Null Mutant. Proc. Natl. Acad. Sci. USA 2008, 105, 2433–2438. [Google Scholar] [CrossRef]
- Binder, A.K.; Rodriguez, K.F.; Hamilton, K.J.; Stockton, P.S.; Reed, C.E.; Korach, K.S. The Absence of Er-Beta Results in Altered Gene Expression in Ovarian Granulosa Cells Isolated from in Vivo Preovulatory Follicles. Endocrinology 2013, 154, 2174–2187. [Google Scholar] [CrossRef] [PubMed]
- Maneix, L.; Antonson, P.; Humire, P.; Rochel-Maia, S.; Castaneda, J.; Omoto, Y.; Kim, H.J.; Warner, M.; Gustafsson, J.A. Estrogen Receptor Beta Exon 3-Deleted Mouse: The Importance of Non-Ere Pathways in Erbeta Signaling. Proc. Natl. Acad. Sci. USA 2015, 112, 5135–5140. [Google Scholar] [CrossRef]
- Novaira, H.J.; Negron, A.L.; Graceli, J.B.; Capellino, S.; Schoeffield, A.; Hoffman, G.E.; Levine, J.E.; Wolfe, A.; Wondisford, F.E.; Radovick, S. Impairments in the Reproductive Axis of Female Mice Lacking Estrogen Receptor Beta in Gnrh Neurons. Am. J. Physiol. Endocrinol. Metab. 2018, 315, E1019–E1033. [Google Scholar] [CrossRef] [PubMed]
- Yuan, B.; Yang, J.; Dubeau, L.; Hu, Y.; Li, R. A Phosphotyrosine Switch in Estrogen Receptor Beta Is Required for Mouse Ovarian Function. Front. Cell Dev. Biol. 2021, 9, 649087. [Google Scholar] [CrossRef]
- Couse, J.F.; Hewitt, S.C.; Bunch, D.O.; Sar, M.; Walker, V.R.; Davis, B.J.; Korach, K.S. Postnatal Sex Reversal of the Ovaries in Mice Lacking Estrogen Receptors Alpha and Beta. Science 1999, 286, 2328–2331. [Google Scholar] [CrossRef]
- Antonson, P.; Nalvarte, I.; Varshney, M.; Xu, L.; Windahl, S.H.; Humire, P.; Ohlsson, C.; Gustafsson, J.A.; Dahlman-Wright, K. Identification of Proteins Highly Expressed in Uterine Fluid from Mice with Hydrometra. Biochem. Biophys. Res. Commun. 2015, 466, 650–655. [Google Scholar] [CrossRef]
- Hewitt, S.C.; Korach, K.S. Estrogen Receptors: New Directions in the New Millennium. Endocr. Rev. 2018, 39, 664–675. [Google Scholar] [CrossRef] [PubMed]
- Arao, Y.; Hamilton, K.J.; Goulding, E.H.; Janardhan, K.S.; Eddy, E.M.; Korach, K.S. Transactivating Function (Af) 2-Mediated Af-1 Activity of Estrogen Receptor Alpha Is Crucial to Maintain Male Reproductive Tract Function. Proc. Natl. Acad. Sci. USA 2012, 109, 21140–21145. [Google Scholar] [CrossRef] [PubMed]
- Acconcia, F.; Ascenzi, P.; Bocedi, A.; Spisni, E.; Tomasi, V.; Trentalance, A.; Visca, P.; Marino, M. Palmitoylation-Dependent Estrogen Receptor Alpha Membrane Localization: Regulation by 17beta-Estradiol. Mol. Biol. Cell 2005, 16, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Saito, K.; Dickey, J.E.; Rodeghiero, S.R.; Toth, B.A.; Kelly, M.J.; Deng, Y.; Singh, U.; Deng, G.; Jiang, J.; Cui, H. Hypomorphism of a Novel Long Eralpha Isoform Causes Severe Reproductive Dysfunctions in Female Mice. Endocrinology 2022, 163, bqac160. [Google Scholar] [CrossRef] [PubMed]
- Warner, M.; Wu, W.F.; Montanholi, L.; Nalvarte, I.; Antonson, P.; Gustafsson, J.A. Ventral Prostate and Mammary Gland Phenotype in Mice with Complete Deletion of the Erbeta Gene. Proc. Natl. Acad. Sci. USA 2020, 117, 4902–4909. [Google Scholar] [CrossRef]
- Jayes, F.L.; Burns, K.A.; Rodriguez, K.F.; Kissling, G.E.; Korach, K.S. The Naturally Occurring Luteinizing Hormone Surge Is Diminished in Mice Lacking Estrogen Receptor Beta in the Ovary. Biol. Reprod. 2014, 90, 24. [Google Scholar] [CrossRef]
- Emmen, J.M.; Couse, J.F.; Elmore, S.A.; Yates, M.M.; Kissling, G.E.; Korach, K.S. In Vitro Growth and Ovulation of Follicles from Ovaries of Estrogen Receptor (Er)Alpha and Erbeta Null Mice Indicate a Role for Erbeta in Follicular Maturation. Endocrinology 2005, 146, 2817–2826. [Google Scholar] [CrossRef] [PubMed]
- Couse, J.F.; Yates, M.M.; Deroo, B.J.; Korach, K.S. Estrogen Receptor-Beta Is Critical to Granulosa Cell Differentiation and the Ovulatory Response to Gonadotropins. Endocrinology 2005, 146, 3247–3262. [Google Scholar] [CrossRef] [PubMed]
- Hrabovszky, E.; Shughrue, P.J.; Merchenthaler, I.; Hajszan, T.; Carpenter, C.D.; Liposits, Z.; Petersen, S.L. Detection of Estrogen Receptor-Beta Messenger Ribonucleic Acid and 125i-Estrogen Binding Sites in Luteinizing Hormone-Releasing Hormone Neurons of the Rat Brain. Endocrinology 2000, 141, 3506–3509. [Google Scholar] [CrossRef]
- Rumi, M.A.; Dhakal, P.; Kubota, K.; Chakraborty, D.; Lei, T.; Larson, M.A.; Wolfe, M.W.; Roby, K.F.; Vivian, J.L.; Soares, M.J. Generation of Esr1-Knockout Rats Using Zinc Finger Nuclease-Mediated Genome Editing. Endocrinology 2014, 155, 1991–1999. [Google Scholar] [CrossRef]
- Rumi, M.A.K.; Singh, P.; Roby, K.F.; Zhao, X.; Iqbal, K.; Ratri, A.; Lei, T.; Cui, W.; Borosha, S.; Dhakal, P.; et al. Defining the Role of Estrogen Receptor Beta in the Regulation of Female Fertility. Endocrinology 2017, 158, 2330–2343. [Google Scholar] [CrossRef]
- Smith, E.P.; Boyd, J.; Frank, G.R.; Takahashi, H.; Cohen, R.M.; Specker, B.; Williams, T.C.; Lubahn, D.B.; Korach, K.S. Estrogen Resistance Caused by a Mutation in the Estrogen-Receptor Gene in a Man. N. Engl. J. Med. 1994, 331, 1056–1061. [Google Scholar] [CrossRef]
- Quaynor, S.D.; Stradtman, E.W., Jr.; Kim, H.-G.; Shen, Y.; Chorich, L.P.; Schreihofer, D.A.; Layman, L.C. Delayed Puberty and Estrogen Resistance in a Woman with Estrogen Receptor Alpha Variant. N. Engl. J. Med. 2013, 369, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Bernard, V.; Kherra, S.; Francou, B.; Fagart, J.; Viengchareun, S.; Guéchot, J.; Ladjouze, A.; Guiochon-Mantel, A.; Korach, K.S.; Binart, N.; et al. Familial Multiplicity of Estrogen Insensitivity Associated with a Loss-of-Function Esr1 Mutation. J. Clin. Endocrinol. Metab. 2017, 102, 93–99. [Google Scholar] [CrossRef]
- Delcour, C.; Khawaja, N.; Gonzalez-Duque, S.; Lebon, S.; Talbi, A.; Drira, L.; Chevenne, D.; Ajlouni, K.; de Roux, N. Estrogen Receptor Alpha Inactivation in 2 Sisters: Different Phenotypic Severities for the Same Pathogenic Variant. J. Clin. Endocrinol. Metab. 2022, 107, e2553–e2562. [Google Scholar] [CrossRef] [PubMed]
- Asadi, M.; Ghafouri-Fard, S.; Zare-Abdollahi, D.; Ebrahim-Habibi, A.; Matin, N. Estrogen Receptor Mutation in a Girl with Primary Amenorrhea. Clin. Genet. 2013, 83, 497–498. [Google Scholar] [CrossRef]
- Lang-Muritano, M.; Sproll, P.; Wyss, S.; Kolly, A.; Hurlimann, R.; Konrad, D.; Biason-Lauber, A. Early-Onset Complete Ovarian Failure and Lack of Puberty in a Woman with Mutated Estrogen Receptor Beta (Esr2). J. Clin. Endocrinol. Metab. 2018, 103, 3748–3756. [Google Scholar] [CrossRef] [PubMed]
- Baetens, D.; Güran, T.; Mendonca, B.B.; Gomes, N.L.; De Cauwer, L.; Peelman, F.; Verdin, H.; Vuylsteke, M.; Van der Linden, M.; Stoop, H.; et al. Biallelic and Monoallelic Esr2 Variants Associated with 46,Xy Disorders of Sex Development. Genet. Med. 2018, 20, 717–727. [Google Scholar] [CrossRef] [PubMed]
- McElreavey, K.; Bashamboo, A. Monogenic Forms of Dsd: An Update. Horm. Res. Paediatr. 2021, 1–25. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nalvarte, I.; Antonson, P. Estrogen Receptor Knockout Mice and Their Effects on Fertility. Receptors 2023, 2, 116-126. https://doi.org/10.3390/receptors2010007
Nalvarte I, Antonson P. Estrogen Receptor Knockout Mice and Their Effects on Fertility. Receptors. 2023; 2(1):116-126. https://doi.org/10.3390/receptors2010007
Chicago/Turabian StyleNalvarte, Ivan, and Per Antonson. 2023. "Estrogen Receptor Knockout Mice and Their Effects on Fertility" Receptors 2, no. 1: 116-126. https://doi.org/10.3390/receptors2010007
APA StyleNalvarte, I., & Antonson, P. (2023). Estrogen Receptor Knockout Mice and Their Effects on Fertility. Receptors, 2(1), 116-126. https://doi.org/10.3390/receptors2010007