1. Introduction
The scope of provenance experiments is very wide. In addition to the main goal, which is the assessment of the production capacity of populations or individual trees, several detailed studies can be carried out on research trials. They provide valuable information on the variability of morphological, anatomical, and physiological features as well as the resistance of trees to diseases and pests. Previous analyses of data from provenance experiments concerned mainly incremental traits, such as tree height, the volume of trees and stands, as well as qualitative traits based on morphological properties [
1,
2]. Provenance trials are also suitable objects for detailed studies covering a wide range of forest tree properties. The adaptive abilities of each species are related to many features of trees, useful for modeling their future development in the conditions of a changing climate [
3]. Among the climatic factors, temperature and precipitation are believed to be the main determinants of both the occurrence and growth of plant species in forests [
4], and these two factors are changing as an effect of global warming resulting from increased greenhouse gas emissions. Key functional (ecophysiological) attributes include leaf size, which determines water use efficiency and the amount of light captured for photosynthesis [
5,
6], and leaf mass per area (LMA) or its inverse—specific leaf area (SLA, the ratio between leaf area and leaf dry mass), that are used to correlate capture of light and tree growth [
7]. The above characteristics of the needle properties show that these features are modified to a big extent by climatic and genetic factors.
The obtained information on the population variability of various Scots pine features indicates the need for more extensive research in this area, also concerning local origins. Particularly valuable information can be obtained from older experimental sites, where the research can be extended to the characteristics of wood. Such possibilities are provided by the 50-year-old provenance experiment in Rogów (central Poland), which represents 16 national origins of Scots pine. The presented analysis of the variability of needles and shoots is one of the components of the interdisciplinary research carried out in this research facility.
2. Material and Methods
2.1. Research Area
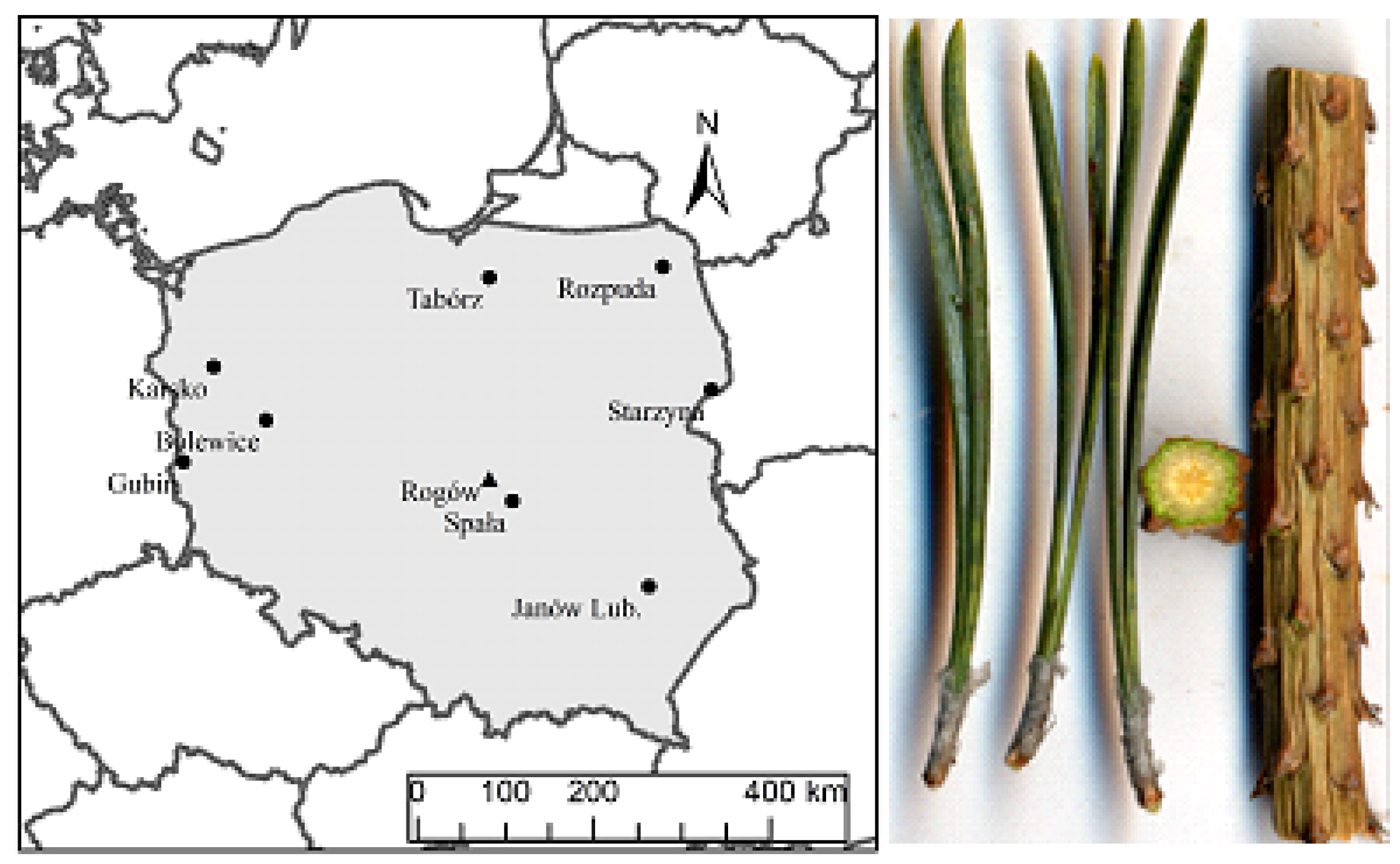
The experimental site is located in central Poland, in the area of Rogów Forest Experimental Station (51°54′29.1″ N 19°54′46.4″ E). It was founded in 1966 in a 5-repetition pattern. The offspring of 16 Polish provenances of Scots pine from all over the country were used. In 2015, samples of shoots and needles from 9 provenances were collected (
Figure 1). The plant material was taken from 50-year-old felled trees. The trees came from the upper layer of the stand and, apart from the basic measurements (incremental features of trees and stands), were used for extensive research on the properties of wood. However, the presented material covers only one feature of needle variability.
2.2. Research Methodology
Nine origins representing the three main areas of Scots pine (Pinus sylvestris L.) in Poland were selected for the measurement of the biometrical features of needles and shoots. Three populations each represented the north-east of the country (Starzyna, Rospuda, and Tabórz), the central (Janów Lubelski, Spała, and Rogów), and the western part of Poland (Bolewice, Karsko, and Gubin). From each population, 13 randomly selected Scots pine trees were tested. Three one-year-old lateral shoots with needles were collected from each selected tree. The research material consisted of 5-cm fragments of shoots with needles, which were cut 3 cm from the bud. The needles were separated from shoots and their pairs were counted. If there were more scarred by the needles on the shoot than the number of separated needles, the gaps were replenished with the nearest needles outside the analyzed section.
Using the Epson Expression12000XL scanner (resolution 12,000 dpi), scans of 5 cm sections and the cross-section of the shoot and samples of about 10 pairs of needles were made (
Figure 1). Using the CooRecorder 9.3.1. software the thickness in half of the length of the taken shoot was measured, and the share of wood, bark, and pith was determined on the cross-sections of the shoots. The actual length (at the edge of the needle) and width of the needles were measured with WinSEEDLE ™ 2017 software.
The shoots and needles were dried at 104 °C in a forced-air dryer by Memmert (type UF110, Producent GmbH + Co.KG). Drying lasted for 24 h. After the drying process was completed, the shoot and the needles were weighed on a laboratory scale (PRECISA BALANCES, type 160M) to determine the dry weight of the shoot and the dry weight of the needles. The accuracy of the dry weight measurement was 0.001 g.
3. Results and Discussion
The trees from which the shoots were collected for detailed measurements were characterized by a similar dbh value (
Table 1). The average thickness of the pines from the central and northern regions was about 220 mm each, and from the western part of Poland they were about 10 mm thicker. The mean values of the tested features of needles and shoots also did not differ much between the three regions. In terms of needle length and total needle length, the needles from the central regions were shorter than those of the other two but had a higher dry weight. The trees originating from the western country part had the thickest shoots, but their volume, dry weight, and density were lower than those of the other regions. One-year-old shoots of trees from central Poland in the cross-section had a slightly higher share of wood (38.6%) than those from the western (37.8%) and northern (38.0%). On the other hand, the lowest share of cortical tissues was found in the pine shoots of central Poland (33.5%). The analysis showed no statistically significant differences between the mean values of all the examined features (
p-value > 0.05).
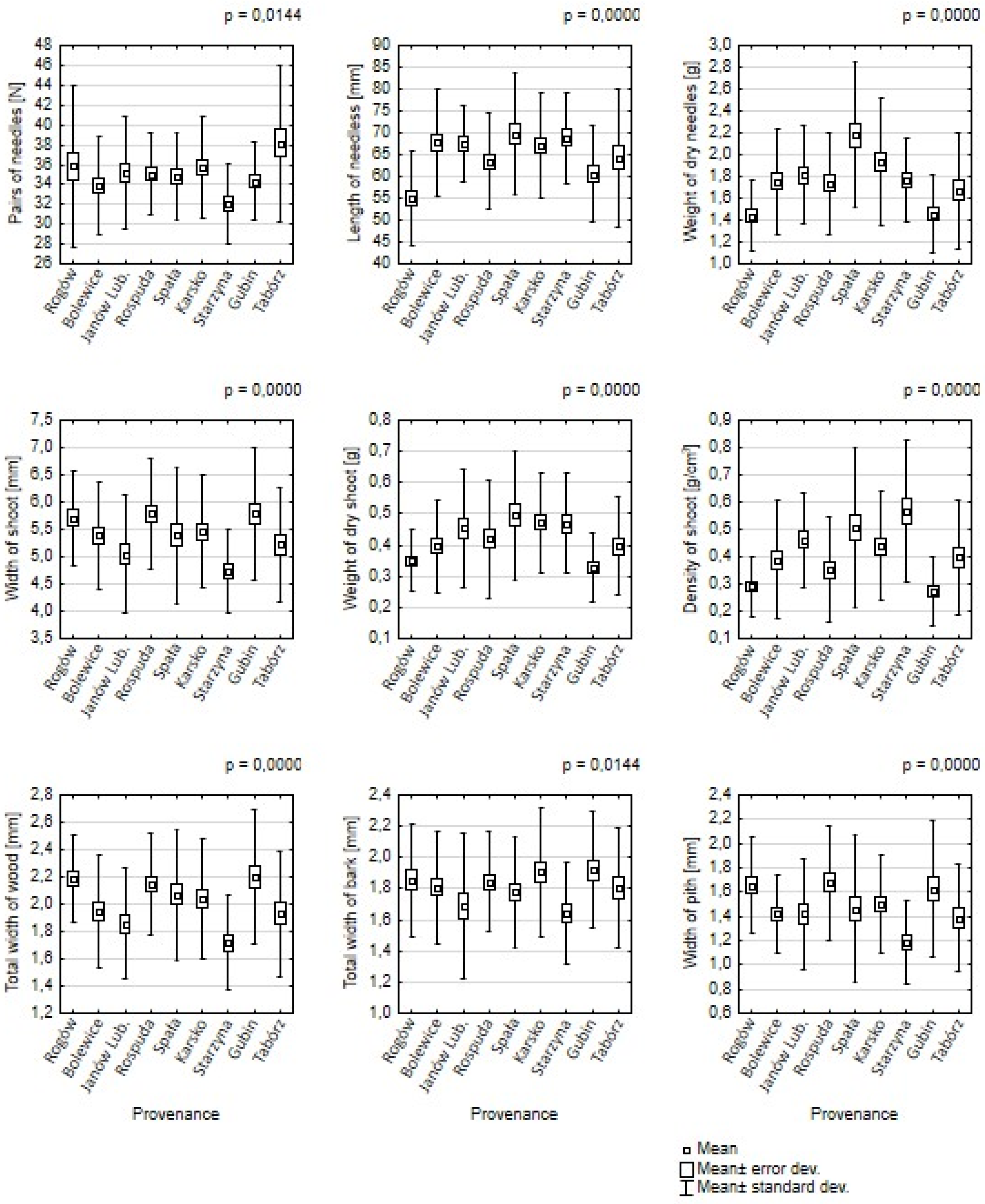
The differences between the average values of all traits for nine populations are statistically significant (α < 0.05) (
Figure 2). The highest density of needles per 5 cm of the shoot was found in the population of Tabórz (32 needles), Rogów and Karsko (36 needles each), and the lowest for Starzyna origin (32 needles) and Bolewice and Gubin (34 needles each). The ranking distribution of the second important feature of needles, which is their length, was significantly different. The pines from Spała and Starzyna (almost 70 mm each) had the longest needles, while the shortest needles were from Rogów (55 mm) and Gubin (approx. 60 mm). The total length of needles was the highest in the populations of Tabórz and Spała, and the lowest was in the populations of Rogów and Gubin. In terms of dry matter, the highest value was recorded in the origin of Spała and Karsko, while the lowest in the local population was from Rogów. The shoots of the Karsko, Janów Lubelski, and Starzyna populations (0.47 g each) had the highest dry weight, while the shoots of Gubin (0.33 g) and Rogów (0.35 g) had the lowest weight. The density of shoots was correlated with the dry matter. Starzyna, Janów Lubelski, and Spała had the highest density of 0.5–0.55 g/cm
3, while the lowest was found in the origins of Gubin (0.28 g) and Rogów (0.29 g).
The condition of the assimilation apparatus determines the efficiency of water use and the amount of light captured and used for the photosynthesis process [
8]. Thus, the differentiation of needle features between provenances may form the processes of photosynthesis and CO
2 binding in a various ways. Leaf modifications most often result from the adaptation of individual origins to specific local environmental conditions [
9]. The greater differentiation of needle traits found in the research between individual origins than between the three regions of Poland may indicate a strong adaptation of pine to local conditions. In addition, our results are consistent with studies carried out in Scotland, where high variability between closely located Scots pine provenances was demonstrated [
10]. These results clearly indicate the need for more detailed studies of individual provenances.
4. Conclusions
There were no significant differences between the studied regions (central, northern, and western Poland) in terms of the variability of the analyzed needle and shoot features.
The variability of the features resulting from the geographical location and climatic conditions in the mother stands, where the analyzed populations originated from, was not found.
The research showed greater variability of Scots pine needles and shoots features between provenances than between regions (central, northern, and western Poland). Within each provenance, different relationships were found between one of the most important incremental traits, which is the tree basal area and the features of needles and shoots. Thus, the results indicate different (specific) properties of needles and shoots in each provenance, which confirms the existence of a strong adaptation of individual provenances to local soil and climatic conditions.
Author Contributions
Conceptualization, W.B.; methodology, W.B.; formal analysis, W.B., M.C., A.K., M.T., H.S. and M.B.; investigation, W.B., M.C., A.K., M.T., H.S. and M.B.; writing—original draft preparation, W.B., M.C., A.K. and M.T.; writing—review and editing, W.B. and A.K.; supervision, W.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Giertych, M. Summary of results on Scotch pine (Pinus sylvestris L.) height growth in IUFRO provenance experiments. Silvae Genet. 1979, 28, 136–152. [Google Scholar]
- Oleksyn, J.; Reich, P.B.; Rachwał, L.; Tjoelker, M.G.; Karolewski, P. Variation in aboveground net primary production of diverse European Pinus sylvestris populations. Trees 2000, 14, 415–421. [Google Scholar] [CrossRef]
- Rehfeldt, G.E.; Worrall, J.J.; Marchetti, S.B.; Crookston, N.L. Adapting forest management to climate change using bioclimate models with topographic drivers. Forestry 2015, 88, 528–539. [Google Scholar] [CrossRef]
- Hansen, A.J.; Neilson, R.P.; Dale, V.H.; Flather, C.H.; Iverson, L.R.; Currie, D.J.; Shafer, S.; Cook, R.; Bartlein, P.J. Global Change in Forests: Responses of Species, Communities, and Biomes: Interactions between climate change and land use are projected to cause large shifts in biodiversity. BioScience 2001, 51, 765–779. [Google Scholar] [CrossRef]
- Xu, F.; Guo, W.; Xu, W.; Wei, Y.; Wang, R. Leaf morphology correlates with water and light availability: What consequences for simple and compound leaves? Prog. Nat. Sci. 2009, 19, 1789–1798. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, Y.; Ma, K. The association of leaf lifespan and background insect herbivory at the interspecific level. Ecology 2017, 98, 425–432. [Google Scholar] [PubMed]
- Poorter, H.; Remkes, C. Leaf area ratio and net assimilation rate of 24 wild species differing in relative growth rate. Oecologia 1990, 83, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Whitman, T.; Aarssen, L.W. The leaf size/number trade-off in herbaceous angiosperms. J. Plant Ecol. 2010, 3, 49–58. [Google Scholar] [CrossRef]
- Poorter, H.; Niinemets, Ü.; Poorter, L.; Wright, I.J.; Villar, R. Causes and consequences of variation in leaf mass per area (LMA): A meta-analysis. New Phytol. 2009, 182, 565–588. [Google Scholar] [CrossRef] [PubMed]
- Provan, J.; Soranzo, N.; Wilson, N.J.; Mcnicol, J.W.; Forrest, G.I.; Cottrell, J.; Powell, W. Gene-pool variation in Caledonian and European Scots pine (Pinus sylvestris L.) revealed by chloroplast simple-sequence repeats. Proc. R. Soc. Lond. Ser. B 1998, 265, 1697–1705. [Google Scholar] [CrossRef] [PubMed]
| Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).




{kind=link}
{kind=link}