
The Role of Lactylation in Mental Illness: Emphasis on Microglia

 , and
, and
Abstract
1. Introduction
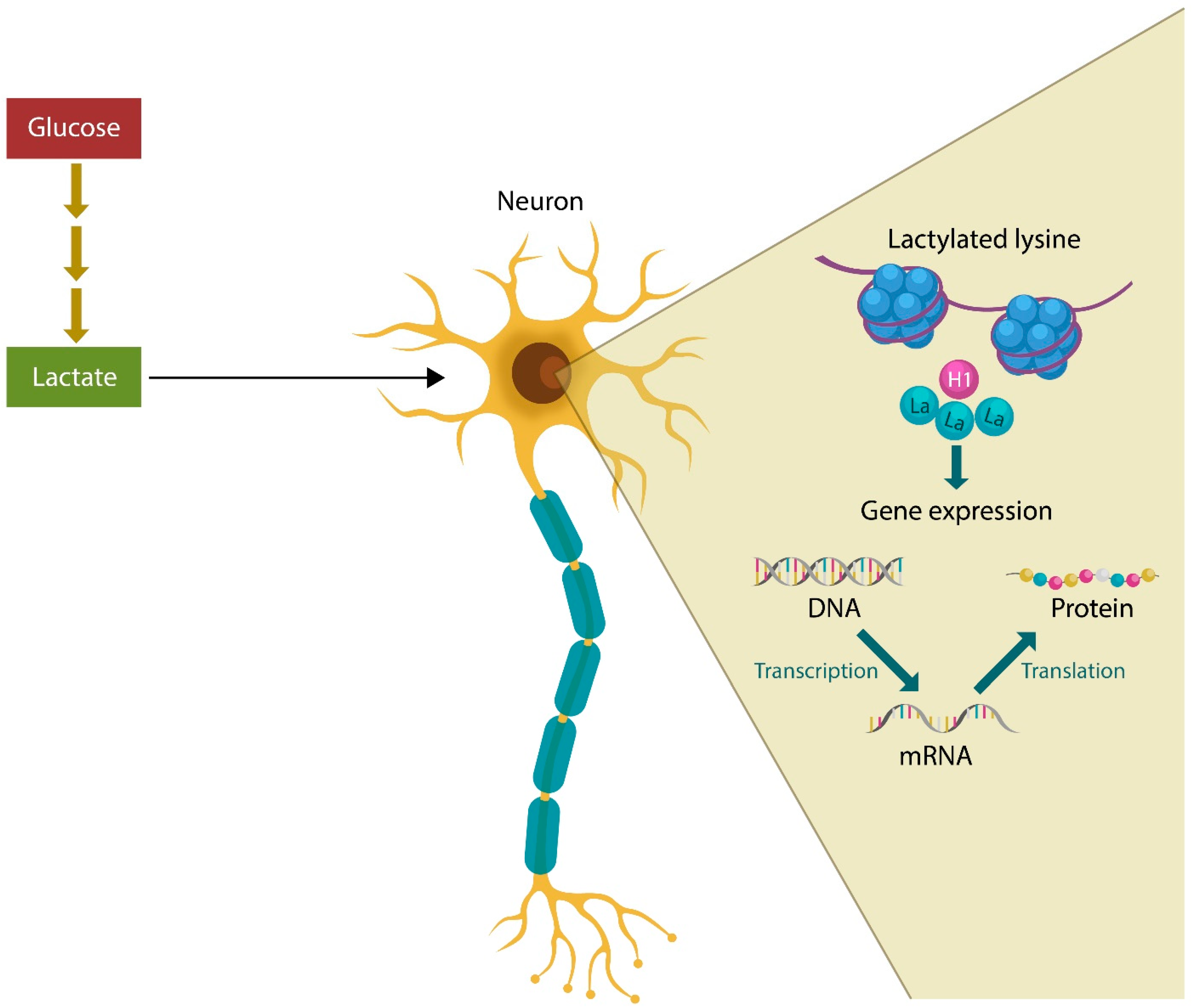
2. Lactate and Mental Illness
Mental Illness, Lactylation, and Microbes
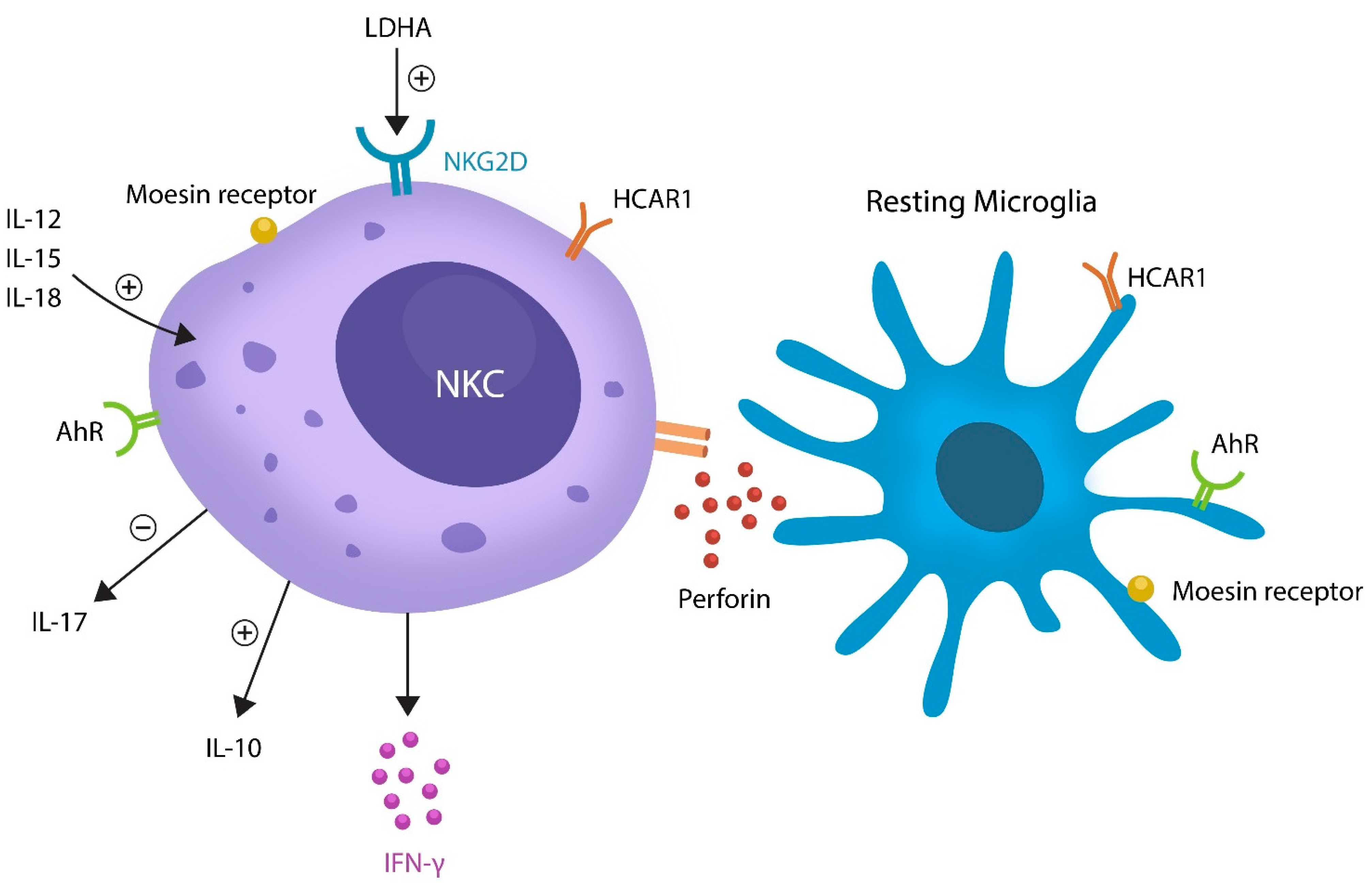
3. Oxytocin and Microglia in Mental Illness

{kind=link}
{kind=link}
| Lactate | OXT | AhR | References |
|---|---|---|---|
| Deactivates microglia by NF-kB inhibition | Deactivates microglia via eIF-2α–ATF4 pathway | Deactivates microglia via NF-kB | [4,15,50] |
| Excess lactate promotes Kla | No known effect on lactylation | Increases LDHA and lactate | [24,132] |
| Lowers ER stress | Lowers ER stress | Lowers ER stress | [16,33,133] |
| Augments gut barrier function | Augments gut barrier | Augments gut barrier | [90,124,134] |
| Upregulated in schizophrenia | Downregulated in schizophrenia | Implicated in schizophrenia | [1,135,136] |
4. Aryl Hydrocarbon Receptor and Mental Illness
5. Microbial Translocation Outside the GI Tract
6. Potential Therapeutic Strategies
7. Lactate
- 1.
- Lactylation inhibitors are compounds belonging to several classes of therapeutics, such as LDHA inhibitors, demethylzeylasteral (DML), and phytotherapeutics, including polyphenols. Here, we review only the agents that can cross the BBB, lower ER stress, and strengthen the intestinal barrier [210,211]. These are as follows.
- 2.
- Sirtuins
- 3.
- LDH inhibitorsLactate-HCAR1 signaling is regulated by LDHA or LDH-5, glycolytic enzymes upregulated by psychological stress, suggesting that inhibiting these enzymes could be therapeutic for neuropsychiatric illness [216]. LDH inhibitors, include the following:
- 3.1.
- 3.2.
- 3.3.
- Galloflavin or NSC 107022 is a polyphenol found in berries which upregulates SIRT6, a key regulator of neuronal mitochondria [183,219]. Dysfunctional SIRT6 was associated with depression-like behavior in mice, suggesting potential benefits for humans with MDD [220]. Galloflavin has been patented as a modulator of protein kinases that can treat a variety of conditions, ranging from cancer to neuropsychiatric disorders (US patent 20030187007A1).
8. OXT
8.1. Angiotensin IV (ANG IV) as an OXT Agonist
8.2. OXT Fragments
9. AhR
- -
- Laquinimod, a quinolone-3-carboxamide developed for the treatment of MS, is a selective AhR ligand that demonstrated encouraging results in animal models of MDD and anxiety [176].
- -
- CH223191 is an antagonistic selective AhR ligand with a favorable profile for neuropsychiatric disorders [179]. This compound was patented in the US for cancer treatment but has still to enter clinical trials for neuropsychiatric disorders (US patent 20160175278A1).
- -
- -
- Dimethyl fumarate, indicated for MS, is not an AhR ligand; however, it upregulates the protective AhR-expressing CD56bright NKCs which eliminate the detrimental autoreactive immune cells to avert autoimmunity [232,233]. For this reason, CD56bright NKCs expansion via AhR or transplantation may comprise a potential therapeutic strategy for neuropsychiatric illness. For example, CD56bright NK cells allogeneic or stem cell transplantation, a treatment used for acute myeloid leukemia, could be therapeutic in both MS and schizophrenia [185,234].
10. Microglia, Lactylation, and Mental Illness
11. Limitations
12. Conclusions
- The aberrant elimination of anti-inflammatory resting microglia by pathologically activated NKCs.
- The reactivation of the neuronal cell cycle with resultant aneuploidy or apoptosis as these cells lack the molecular machinery to complete mitosis.
- Several interventions may restore the homeostasis of anti-inflammatory microglia, including 1. lowering lactylation, 2. enhancing OXT signaling, 3. modulating AhR, and 4. expanding CD56NKCs. These strategies lower the ER stress and strengthen the gut barrier, limiting microbial migration outside of the GI tract, thus decreasing the risk of microglial activation by translocated bacterial antigens.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Rowland, L.M.; Pradhan, S.; Korenic, S.; Wijtenburg, S.A.; Hong, L.E.; Edden, R.A.; Barker, P.B. Elevated brain lactate in schizophrenia: A 7 T magnetic resonance spectroscopy study. Transl. Psychiatry 2016, 6, e967. [Google Scholar] [CrossRef] [PubMed]
- Dogan, A.E.; Yuksel, C.; Du, F.; Chouinard, V.-A.; Öngür, D. Brain lactate and pH in schizophrenia and bipolar disorder: A systematic review of findings from magnetic resonance studies. Neuropsychopharmacology 2018, 43, 1681–1690. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, C.R.; Mielnik, C.A.; Funk, A.; O’Donovan, S.M.; Bentea, E.; Pletnikov, M.; Ramsey, A.J.; Wen, Z.; Rowland, L.M.; McCullumsmith, R.E. Measurement of lactate levels in postmortem brain, iPSCs, and animal models of schizophrenia. Sci. Rep. 2019, 9, 5087. [Google Scholar] [CrossRef]
- Pruett, B.S.; Meador-Woodruff, J.H. Evidence for altered energy metabolism, increased lactate, and decreased pH in schizophrenia brain: A focused review and meta-analysis of human postmortem and magnetic resonance spectroscopy studies. Schizophr. Res. 2020, 223, 29–42. [Google Scholar] [CrossRef]
- Vallée, A.; Vallée, J.-N. Warburg effect hypothesis in autism Spectrum disorders. Mol. Brain 2018, 11, 1. [Google Scholar] [CrossRef]
- Park, H.-J.; Choi, I.; Leem, K.-H. Decreased Brain pH and Pathophysiology in Schizophrenia. Int. J. Mol. Sci. 2021, 22, 8358. [Google Scholar] [CrossRef]
- Li, J.; Chen, L.; Qin, Q.; Wang, D.; Zhao, J.; Gao, H.; Yuan, X.; Zhang, J.; Zou, Y.; Mao, Z.; et al. Upregulated hexokinase 2 expression induces the apoptosis of dopaminergic neurons by promoting lactate production in Parkinson’s disease. Neurobiol. Dis. 2022, 163, 105605. [Google Scholar] [CrossRef]
- Hollnagel, J.-O.; Cesetti, T.; Schneider, J.; Vazetdinova, A.; Valiullina-Rakhmatullina, F.; Lewen, A.; Rozov, A.; Kann, O. Lactate Attenuates Synaptic Transmission and Affects Brain Rhythms Featuring High Energy Expenditure. iScience 2020, 23, 101316. [Google Scholar] [CrossRef]
- Krzyściak, W.; Karcz, P.; Bystrowska, B.; Szwajca, M.; Bryll, A.; Śmierciak, N.; Ligęzka, A.; Turek, A.; Kozicz, T.; Skalniak, A.E.; et al. The Association of the Oral Microbiota with the Effects of Acid Stress Induced by an Increase of Brain Lactate in Schizophrenia Patients. Biomedicines 2023, 11, 240. [Google Scholar] [CrossRef]
- Ghosh, S.; Castillo, E.; Frias, E.S.; Swanson, R.A. Bioenergetic regulation of microglia. Glia 2017, 66, 1200–1212. [Google Scholar] [CrossRef]
- Nimmerjahn, A.; Kirchhoff, F.; Helmchen, F. Resting Microglial Cells Are Highly Dynamic Surveillants of Brain Parenchyma in Vivo. Science 2005, 308, 1314–1318. [Google Scholar] [CrossRef] [PubMed]
- Block, M.L.; Zecca, L.; Hong, J.-S. Microglia-mediated neurotoxicity: Uncovering the molecular mechanisms. Nat. Rev. Neurosci. 2007, 8, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Tauffenberger, A.; Fiumelli, H.; Almustafa, S.; Magistretti, P.J. Lactate and pyruvate promote oxidative stress resistance through hormetic ROS signaling. Cell Death Dis. 2019, 10, 653. [Google Scholar] [CrossRef] [PubMed]
- Monsorno, K.; Buckinx, A.; Paolicelli, R.C. Microglial metabolic flexibility: Emerging roles for lactate. Trends Endocrinol. Metab. 2022, 33, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Le, W. Differential Roles of M1 and M2 Microglia in Neurodegenerative Diseases. Mol. Neurobiol. 2016, 53, 1181–1194. [Google Scholar] [CrossRef]
- Karnib, N.; El-Ghandour, R.; El Hayek, L.; Nasrallah, P.; Khalifeh, M.; Barmo, N.; Jabre, V.; Ibrahim, P.; Bilen, M.; Stephan, J.S.; et al. Lactate is an antidepressant that mediates resilience to stress by modulating the hippocampal levels and activity of histone deacetylases. Neuropsychopharmacology 2019, 44, 1152–1162. [Google Scholar] [CrossRef] [PubMed]
- Carrard, A.; Elsayed, M.; Margineanu, M.; Boury-Jamot, B.; Fragnière, L.; Meylan, E.M.; Petit, J.-M.; Fiumelli, H.; Magistretti, P.J.; Martin, J.-L. Peripheral administration of lactate produces antidepressant-like effects. Mol. Psychiatry 2016, 23, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Fan, M.; Wang, X.; Xu, J.; Wang, Y.; Tu, F.; Gill, P.S.; Ha, T.; Liu, L.; Williams, D.L.; et al. Lactate promotes macrophage HMGB1 lactylation, acetylation, and exosomal release in polymicrobial sepsis. Cell Death Differ. 2021, 29, 133–146. [Google Scholar] [CrossRef]
- Gao, Y.; Zhou, H.; Liu, G.; Wu, J.; Yuan, Y.; Shang, A. Tumor Microenvironment: Lactic Acid Promotes Tumor Development. J. Immunol. Res. 2022, 2022, 3119375. [Google Scholar] [CrossRef]
- Ishihara, S.; Hata, K.; Hirose, K.; Okui, T.; Toyosawa, S.; Uzawa, N.; Nishimura, R.; Yoneda, T. The lactate sensor GPR81 regulates glycolysis and tumor growth of breast cancer. Sci. Rep. 2022, 12, 6261. [Google Scholar] [CrossRef]
- Ichihara, Y.; Doi, T.; Ryu, Y.; Nagao, M.; Sawada, Y.; Ogata, T. Oligodendrocyte Progenitor Cells Directly Utilize Lactate for Promoting Cell Cycling and Differentiation. J. Cell. Physiol. 2016, 232, 986–995. [Google Scholar] [CrossRef] [PubMed]
- Cerexhe, L.; Easton, C.; Macdonald, E.; Renfrew, L.; Sculthorpe, N. Blood lactate concentrations during rest and exercise in people with Multiple Sclerosis: A systematic review and meta-analysis. Mult. Scler. Relat. Disord. 2021, 57, 103454. [Google Scholar] [CrossRef] [PubMed]
- Keytsman, C.; Hansen, D.; Wens, I.; Eijnde, B.O. Exercise-induced lactate responses in Multiple Sclerosis: A retrospective analysis. Neurorehabilitation 2019, 45, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Pan, R.-Y.; He, L.; Zhang, J.; Liu, X.; Liao, Y.; Gao, J.; Liao, Y.; Yan, Y.; Li, Q.; Zhou, X.; et al. Positive feedback regulation of microglial glucose metabolism by histone H4 lysine 12 lactylation in Alzheimer’s disease. Cell Metab. 2022, 34, 634–648.e6. [Google Scholar] [CrossRef]
- Xie, Y.; Hu, H.; Liu, M.; Zhou, T.; Cheng, X.; Huang, W.; Cao, L. The role and mechanism of histone lactylation in health and diseases. Front. Genet. 2022, 13, 949252. [Google Scholar] [CrossRef]
- Liberti, M.V.; Locasale, J.W. The Warburg Effect: How Does it Benefit Cancer Cells? Trends Biochem. Sci. 2016, 41, 211–218, Erratum in Trends Biochem. Sci. 2016, 41, 287. [Google Scholar] [CrossRef]
- Zhao, Y.; Xu, H. Microglial lactate metabolism as a potential therapeutic target for Alzheimer’s disease. Mol. Neurodegener. 2022, 17, 36. [Google Scholar] [CrossRef]
- Sabari, B.R.; Zhang, D.; Allis, C.D.; Zhao, Y. Metabolic regulation of gene expression through histone acylations. Nat. Rev. Mol. Cell Biol. 2017, 18, 90–101. [Google Scholar] [CrossRef]
- Zhang, D.; Tang, Z.; Huang, H.; Zhou, G.; Cui, C.; Weng, Y.; Liu, W.; Kim, S.; Lee, S.; Perez-Neut, M.; et al. Metabolic regulation of gene expression by histone lactylation. Nature 2019, 574, 575–580. [Google Scholar] [CrossRef]
- Okazaki, S.; Boku, S.; Otsuka, I.; Mouri, K.; Aoyama, S.; Shiroiwa, K.; Sora, I.; Fujita, A.; Shirai, Y.; Shirakawa, O.; et al. The cell cycle-related genes as biomarkers for schizophrenia. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2016, 70, 85–91. [Google Scholar] [CrossRef]
- Luo, Y.; Yang, Z.; Yu, Y.; Zhang, P. HIF1α lactylation enhances KIAA1199 transcription to promote angiogenesis and vasculogenic mimicry in prostate cancer. Int. J. Biol. Macromol. 2022, 222, 2225–2243. [Google Scholar] [CrossRef] [PubMed]
- Andersson, U.; Yang, H. HMGB1 is a critical molecule in the pathogenesis of Gram-negative sepsis. J. Intensive Med. 2022, 2, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Klein, B.Y.; Tamir, H.; Ludwig, R.J.; Glickstein, S.B.; Welch, M.G. Colostrum oxytocin modulates cellular stress response, inflammation, and autophagy markers in newborn rat gut villi. Biochem. Biophys. Res. Commun. 2017, 487, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Maejima, Y.; Yokota, S.; Ono, T.; Yu, Z.; Yamachi, M.; Hidema, S.; Nollet, K.E.; Nishimori, K.; Tomita, H.; Yaginuma, H.; et al. Identification of oxytocin expression in human and murine microglia. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2022, 119, 110600. [Google Scholar] [CrossRef]
- Pedersen, C.A.; Gibson, C.M.; Rau, S.W.; Salimi, K.; Smedley, K.L.; Casey, R.L.; Leserman, J.; Jarskog, L.F.; Penn, D.L. Intranasal oxytocin reduces psychotic symptoms and improves Theory of Mind and social perception in schizophrenia. Schizophr. Res. 2011, 132, 50–53. [Google Scholar] [CrossRef]
- Macdonald, K.; Feifel, D. Oxytocin in schizophrenia: A review of evidence for its therapeutic effects. Acta Neuropsychiatr. 2012, 24, 130–146. [Google Scholar] [CrossRef]
- Caldwell, H.K.; Stephens, S.L.; Young, W.S. Oxytocin as a natural antipsychotic: A study using oxytocin knockout mice. Mol. Psychiatry 2009, 14, 190–196. [Google Scholar] [CrossRef]
- Jahangard, L.; Shayganfard, M.; Ghiasi, F.; Salehi, I.; Haghighi, M.; Ahmadpanah, M.; Bahmani, D.S.; Brand, S. Serum oxytocin concentrations in current and recent suicide survivors are lower than in healthy controls. J. Psychiatr. Res. 2020, 128, 75–82. [Google Scholar] [CrossRef]
- Panaro, M.A.; Benameur, T.; Porro, C. Hypothalamic Neuropeptide Brain Protection: Focus on Oxytocin. J. Clin. Med. 2020, 9, 1534. [Google Scholar] [CrossRef]
- Parris, M.S.; Grunebaum, M.F.; Galfalvy, H.C.; Andronikashvili, A.; Burke, A.K.; Yin, H.; Min, E.; Huang, Y.-Y.; Mann, J.J. Attempted suicide and oxytocin-related gene polymorphisms. J. Affect. Disord. 2018, 238, 62–68. [Google Scholar] [CrossRef]
- Baudon, A.; Creusot, E.C.; Althammer, F.; Schaaf, C.P.; Charlet, A. Emerging role of astrocytes in oxytocin-mediated control of neural circuits and brain functions. Prog. Neurobiol. 2022, 217, 102328. [Google Scholar] [CrossRef] [PubMed]
- Jokinen, J.; Chatzittofis, A.; Hellström, C.; Nordström, P.; Uvnäs-Moberg, K.; Åsberg, M. Low CSF oxytocin reflects high intent in suicide attempters. Psychoneuroendocrinology 2012, 37, 482–490. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Liu, S.; Bai, X.; Gao, Y.; Liu, G.; Wang, X.; Liu, D.; Li, T.; Hao, A.; Wang, Z. Oxytocin inhibits lipopolysaccharide-induced inflammation in microglial cells and attenuates microglial activation in lipopolysaccharide-treated mice. J. Neuroinflammation 2016, 13, 77. [Google Scholar] [CrossRef] [PubMed]
- Inoue, Y.U.; Miwa, H.; Hori, K.; Kaneko, R.; Morimoto, Y.; Koike, E.; Asami, J.; Kamijo, S.; Yamada, M.; Hoshino, M.; et al. Targeting Neurons with Functional Oxytocin Receptors: A Novel Set of Simple Knock-In Mouse Lines for Oxytocin Receptor Visualization and Manipulation. Eneuro 2022, 9, ENEURO.0423-21.2022. [Google Scholar] [CrossRef] [PubMed]
- Mairesse, J.; Zinni, M.; Pansiot, J.; Hassan-Abdi, R.; Demene, C.; Colella, M.; Charriaut-Marlangue, C.; Novais, A.R.B.; Tanter, M.; Maccari, S.; et al. Oxytocin receptor agonist reduces perinatal brain damage by targeting microglia. Glia 2018, 67, 345–359. [Google Scholar] [CrossRef] [PubMed]
- De Cagna, F.; Fusar-Poli, L.; Damiani, S.; Rocchetti, M.; Giovanna, G.; Mori, A.; Politi, P.; Brondino, N. The Role of Intranasal Oxytocin in Anxiety and Depressive Disorders: A Systematic Review of Randomized Controlled Trials. Clin. Psychopharmacol. Neurosci. 2019, 17, 1–11. [Google Scholar] [CrossRef]
- Sorg, O. AhR signalling and dioxin toxicity. Toxicol. Lett. 2014, 230, 225–233. [Google Scholar] [CrossRef]
- Climaco-Arvizu, S.; Domínguez-Acosta, O.; Cabañas-Cortés, M.A.; Rodríguez-Sosa, M.; Gonzalez, F.J.; Vega, L.; Elizondo, G. Aryl hydrocarbon receptor influences nitric oxide and arginine production and alters M1/M2 macrophage polarization. Life Sci. 2016, 155, 76–84. [Google Scholar] [CrossRef]
- Sahebnasagh, A.; Hashemi, J.; Khoshi, A.; Saghafi, F.; Avan, R.; Faramarzi, F.; Azimi, S.; Habtemariam, S.; Sureda, A.; Khayatkashani, M.; et al. Aromatic hydrocarbon receptors in mitochondrial biogenesis and function. Mitochondrion 2021, 61, 85–101. [Google Scholar] [CrossRef]
- Di Giaimo, R.; Durovic, T.; Barquin, P.; Kociaj, A.; Lepko, T.; Aschenbroich, S.; Breunig, C.T.; Irmler, M.; Cernilogar, F.M.; Schotta, G.; et al. The Aryl Hydrocarbon Receptor Pathway Defines the Time Frame for Restorative Neurogenesis. Cell Rep. 2018, 25, 3241–3251.e5. [Google Scholar] [CrossRef]
- Tanaka, M.; Fujikawa, M.; Oguro, A.; Itoh, K.; Vogel, C.F.A.; Ishihara, Y. Involvement of the Microglial Aryl Hydrocarbon Receptor in Neuroinflammation and Vasogenic Edema after Ischemic Stroke. Cells 2021, 10, 718. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Wang, S.; Hu, S.; Yang, B.; Zhang, H. Host-microbiota interactions: The aryl hydrocarbon receptor in the acute and chronic phases of cerebral ischemia. Front. Immunol. 2022, 13, 967300. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Nieves, U.Y.; Mundy, D.C.; Shin, J.H.; Tam, K.; Sunwoo, J.B. The aryl hydrocarbon receptor modulates the function of human CD56bright NK cells. Eur. J. Immunol. 2018, 48, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Lünemann, A.; Lünemann, J.D.; Roberts, S.; Messmer, B.; da Silva, R.B.; Raine, C.S.; Münz, C. Human NK Cells Kill Resting but Not Activated Microglia via NKG2D- and NKp46-Mediated Recognition. J. Immunol. 2008, 181, 6170–6177. [Google Scholar] [CrossRef]
- Peter, K.; Rehli, M.; Singer, K.; Renner-Sattler, K.; Kreutz, M. Lactic acid delays the inflammatory response of human monocytes. Biochem. Biophys. Res. Commun. 2015, 457, 412–418. [Google Scholar] [CrossRef]
- Hagihara, H.; Shoji, H.; Otabi, H.; Toyoda, A.; Katoh, K.; Namihira, M.; Miyakawa, T. Protein lactylation induced by neural excitation. Cell Rep. 2021, 37, 109820. [Google Scholar] [CrossRef]
- Jiang, J.; Huang, D.; Jiang, Y.; Hou, J.; Tian, M.; Li, J.; Sun, L.; Zhang, Y.; Zhang, T.; Li, Z.; et al. Lactate Modulates Cellular Metabolism Through Histone Lactylation-Mediated Gene Expression in Non-Small Cell Lung Cancer. Front. Oncol. 2021, 11, 647559. [Google Scholar] [CrossRef]
- Mertens, J.; Herdy, J.R.; Traxler, L.; Schafer, S.T.; Schlachetzki, J.C.M.; Böhnke, L.; Reid, D.A.; Lee, H.; Zangwill, D.; Fernandes, D.P.; et al. Age-dependent instability of mature neuronal fate in induced neurons from Alzheimer’s patients. Cell Stem Cell 2021, 28, 1533–1548.e6. [Google Scholar] [CrossRef]
- Yurov, Y.B.; Vorsanova, S.G.; Demidova, I.; Kolotii, A.D.; Soloviev, I.V.; Iourov, I.Y. Mosaic Brain Aneuploidy in Mental Illnesses: An Association of Low-level post-zygotic Aneuploidy with Schizophrenia and Comorbid Psychiatric Disorders. Curr. Genom. 2018, 19, 163–172. [Google Scholar] [CrossRef]
- Shegay, P.V.; Zabolotneva, A.A.; Shatova, O.P.; Shestopalov, A.V.; Kaprin, A.D. Evolutionary View on Lactate-Dependent Mechanisms of Maintaining Cancer Cell Stemness and Reprimitivization. Cancers 2022, 14, 4552. [Google Scholar] [CrossRef]
- Fan, Y.; Abrahamsen, G.; McGrath, J.J.; Mackay-Sim, A. Altered Cell Cycle Dynamics in Schizophrenia. Biol. Psychiatry 2012, 71, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Katsel, P.; Davis, K.L.; Li, C.; Tan, W.; Greenstein, E.; Hoffman, L.B.K.; Haroutunian, V. Abnormal Indices of Cell Cycle Activity in Schizophrenia and their Potential Association with Oligodendrocytes. Neuropsychopharmacology 2008, 33, 2993–3009. [Google Scholar] [CrossRef] [PubMed]
- Lineweaver, C.H.; Bussey, K.J.; Blackburn, A.C.; Davies, P.C.W. Cancer progression as a sequence of atavistic reversions. Bioessays 2021, 43, e2000305. [Google Scholar] [CrossRef]
- Alvarez, Z.; Hyroššová, P.; Perales, J.C.; Alcantara, S.; Ova, P.H. Neuronal Progenitor Maintenance Requires Lactate Metabolism and PEPCK-M-Directed Cataplerosis. Cereb. Cortex 2014, 26, 1046–1058. [Google Scholar] [CrossRef] [PubMed]
- Odenwelder, D.C.; Lu, X.; Harcum, S.W. Induced pluripotent stem cells can utilize lactate as a metabolic substrate to support proliferation. Biotechnol. Prog. 2020, 37, e3090. [Google Scholar] [CrossRef]
- Goldman, S.; Pulsinelli, W.A.; Clarke, W.Y.; Kraig, R.P.; Plum, F. The Effects of Extracellular Acidosis on Neurons and Glia in vitro. J. Cereb. Blood Flow Metab. 1989, 9, 471–477. [Google Scholar] [CrossRef]
- Tan, Z.; Chu, D.Z.V.; Chan, Y.J.A.; Lu, Y.E.; Rancati, G. Mammalian Cells Undergo Endoreduplication in Response to Lactic Acidosis. Sci. Rep. 2018, 8, 2890. [Google Scholar] [CrossRef]
- Lev-Vachnish, Y.; Cadury, S.; Rotter-Maskowitz, A.; Feldman, N.; Roichman, A.; Illouz, T.; Varvak, A.; Nicola, R.; Madar, R.; Okun, E. L-Lactate Promotes Adult Hippocampal Neurogenesis. Front. Neurosci. 2019, 13, 403. [Google Scholar] [CrossRef]
- Ordoño, J.; Pérez-Amodio, S.; Ball, K.; Aguirre, A.; Engel, E. The generation of a lactate-rich environment stimulates cell cycle progression and modulates gene expression on neonatal and hiPSC-derived cardiomyocytes. Biomater. Adv. 2022, 139, 213035. [Google Scholar] [CrossRef]
- Rosenblatt, J. Mitosis: Moesin and the Importance of Being Round. Curr. Biol. 2008, 18, R292–R293. [Google Scholar] [CrossRef]
- Congdon, E.E.; Sigurdsson, E.M. Tau-targeting therapies for Alzheimer disease. Nat. Rev. Neurol. 2018, 14, 399–415. [Google Scholar] [CrossRef] [PubMed]
- Beckmann, A.; Ramirez, P.; Gamez, M.; Gonzalez, E.; De Mange, J.; Bieniek, K.F.; Ray, W.J.; Frost, B. Moesin is an effector of tau-induced actin overstabilization, cell cycle activation, and neurotoxicity in Alzheimer’s disease. iScience 2023, 26, 106152. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Zhou, J.; Chen, Q.; Xu, X.; Gao, J.; Li, X.; Shao, Q.; Zhou, B.; Zhou, H.; Wei, S.; et al. Tumor metabolite lactate promotes tumorigenesis by modulating MOESIN lactylation and enhancing TGF-β signaling in regulatory T cells. Cell Rep. 2022, 40, 111122. [Google Scholar] [CrossRef] [PubMed]
- Satooka, H.; Matsui, M.; Ichioka, S.; Nakamura, Y.; Hirata, T. The ERM protein moesin regulates natural killer cell homeostasis in vivo. Cell. Immunol. 2022, 371, 104456. [Google Scholar] [CrossRef] [PubMed]
- Ben-Shmuel, A.; Sabag, B.; Biber, G.; Barda-Saad, M. The Role of the Cytoskeleton in Regulating the Natural Killer Cell Immune Response in Health and Disease: From Signaling Dynamics to Function. Front. Cell Dev. Biol. 2021, 9, 609532. [Google Scholar] [CrossRef] [PubMed]
- Bowden, N.A.; Weidenhofer, J.; Scott, R.J.; Schall, U.; Todd, J.; Michie, P.T.; Tooney, P.A. Preliminary investigation of gene expression profiles in peripheral blood lymphocytes in schizophrenia. Schizophr. Res. 2006, 82, 175–183. [Google Scholar] [CrossRef]
- Moreno-Yruela, C.; Zhang, D.; Wei, W.; Bæk, M.; Liu, W.; Gao, J.; Danková, D.; Nielsen, A.L.; Bolding, J.E.; Yang, L.; et al. Class I histone deacetylases (HDAC1–3) are histone lysine delactylases. Sci. Adv. 2022, 8, eabi6696. [Google Scholar] [CrossRef]
- Yu, J.; de Belle, I.; Liang, H.; Adamson, E.D. Coactivating Factors p300 and CBP Are Transcriptionally Crossregulated by Egr1 in Prostate Cells, Leading to Divergent Responses. Mol. Cell 2004, 15, 83–94. [Google Scholar] [CrossRef]
- Gilbert, T.; Zürcher, N.R.; Wu, C.J.; Bhanot, A.; Hightower, B.G.; Kim, M.; Albrecht, D.S.; Wey, H.-Y.; Schroeder, F.A.; Rodriguez-Thompson, A.; et al. PET neuroimaging reveals histone deacetylase dysregulation in schizophrenia. J. Clin. Investig. 2018, 129, 364–372. [Google Scholar] [CrossRef]
- Martínez-Pinteño, A.; Gassó, P.; Prohens, L.; Segura, A.G.; Parellada, M.; Saiz-Ruiz, J.; Cuesta, M.J.; Bernardo, M.; Lafuente, A.; Mas, S.; et al. Identification of EP300 as a Key Gene Involved in Antipsychotic-Induced Metabolic Dysregulation Based on Integrative Bioinformatics Analysis of Multi-Tissue Gene Expression Data. Front. Pharmacol. 2021, 12, 729474. [Google Scholar] [CrossRef]
- Tan, M.; Shen, L.; Hou, Y. Epigenetic modification of BDNF mediates neuropathic pain via miR-30a-3p/EP300 axis in CCI rats. Biosci. Rep. 2020, 40, BSR20194442. [Google Scholar] [CrossRef] [PubMed]
- Gören, J.L. Brain-derived neurotrophic factor and schizophrenia. Ment. Health Clin. 2016, 6, 285–288. [Google Scholar] [CrossRef] [PubMed]
- Bredy, T.W.; Wu, H.; Crego, C.; Zellhoefer, J.; Sun, Y.E.; Barad, M. Histone modifications around individual BDNF gene promoters in prefrontal cortex are associated with extinction of conditioned fear. Learn. Mem. 2007, 14, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.; Yang, F.; Xiao, Z.; Luo, H.; Chen, H.; Chen, Z.; Liu, Q.; Xiao, Y. Lactylation: Novel epigenetic regulatory and therapeutic opportunities. Am J Physiol Endocrinol Metab. 2023, 324, E330–E338. [Google Scholar] [CrossRef] [PubMed]
- Boutahar, N.; Reynaud, E.; Lassabliere, F.; Borg, J. Brain-derived neurotrophic factor inhibits cell cycle reentry but not endoplasmic reticulum stress in cultured neurons following oxidative or excitotoxic stress. J. Neurosci. Res. 2010, 88, 2263–2271. [Google Scholar] [CrossRef] [PubMed]
- Sedgwick, A.J.; Ghazanfari, N.; Constantinescu, P.; Mantamadiotis, T.; Barrow, A.D. The Role of NK Cells and Innate Lymphoid Cells in Brain Cancer. Front. Immunol. 2020, 11, 1549. [Google Scholar] [CrossRef]
- Marzan, D.E.; Brügger-Verdon, V.; West, B.L.; Liddelow, S.; Samanta, J.; Salzer, J.L. Activated microglia drive demyelination via CSF1R signaling. Glia 2021, 69, 1583–1604. [Google Scholar] [CrossRef]
- Gaffney, D.O.; Jennings, E.Q.; Anderson, C.C.; Marentette, J.O.; Shi, T.; Oxvig, A.-M.S.; Streeter, M.D.; Johannsen, M.; Spiegel, D.A.; Chapman, E.; et al. Non-enzymatic Lysine Lactoylation of Glycolytic Enzymes. Cell Chem. Biol. 2020, 27, 206–213.e6. [Google Scholar] [CrossRef]
- Tarantino, N.; Leboyer, M.; Bouleau, A.; Hamdani, N.; Richard, J.R.; Boukouaci, W.; Ching-Lien, W.; Godin, O.; Bengoufa, D.; Le Corvoisier, P.; et al. Natural killer cells in first-episode psychosis: An innate immune signature? Mol. Psychiatry 2021, 26, 5297–5306. [Google Scholar] [CrossRef]
- Lee, Y.-S.; Kim, T.-Y.; Kim, Y.; Lee, S.-H.; Kim, S.; Kang, S.W.; Yang, J.-Y.; Baek, I.-J.; Sung, Y.H.; Park, Y.-Y.; et al. Microbiota-Derived Lactate Accelerates Intestinal Stem-Cell-Mediated Epithelial Development. Cell Host Microbe 2018, 24, 833–846.e6. [Google Scholar] [CrossRef]
- Wang, S.P.; Rubio, L.A.; Duncan, S.; Donachie, G.E.; Holtrop, G.; Lo, G.; Farquharson, F.M.; Wagner, J.; Parkhill, J.; Louis, P.; et al. Pivotal Roles for pH, Lactate, and Lactate-Utilizing Bacteria in the Stability of a Human Colonic Microbial Ecosystem. Msystems 2020, 5, e00645-20. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.-F.; Wang, C.-Y.; Wang, J.-H.; Wang, Q.-N.; Li, S.-J.; Wang, H.-O.; Zhou, F.; Li, J.-M. Short-Chain Fatty Acids Ameliorate Depressive-like Behaviors of High Fructose-Fed Mice by Rescuing Hippocampal Neurogenesis Decline and Blood–Brain Barrier Damage. Nutrients 2022, 14, 1882. [Google Scholar] [CrossRef] [PubMed]
- Caslin, H.L.; Abebayehu, D.; Pinette, J.A.; Ryan, J.J. Lactate Is a Metabolic Mediator That Shapes Immune Cell Fate and Function. Front. Physiol. 2021, 12, 688485. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, L.; Glesaaen, E.R.; Palibrk, V.; Pannone, M.; Wang, W.; Al-Jabri, A.; Suganthan, R.; Meyer, N.; Austbø, M.L.; Lin, X.; et al. Lactate receptor HCAR1 regulates neurogenesis and microglia activation after neonatal hypoxia-ischemia. Elife 2022, 11, e76451. [Google Scholar] [CrossRef] [PubMed]
- Errea, A.; Cayet, D.; Marchetti, P.; Tang, C.; Kluza, J.; Offermanns, S.; Sirard, J.-C.; Rumbo, M. Lactate Inhibits the Pro-Inflammatory Response and Metabolic Reprogramming in Murine Macrophages in a GPR81-Independent Manner. PLoS ONE 2016, 11, e0163694. [Google Scholar] [CrossRef]
- Nicola, R.; Madar, R.; Okun, E. HCAR1-Mediated l-Lactate Signaling Suppresses Microglial Phagocytosis. NeuroMol. Med. 2022, 24, 399–404. [Google Scholar] [CrossRef]
- Dong, H.; Zhang, J.; Zhang, H.; Han, Y.; Lu, C.; Chen, C.; Tan, X.; Wang, S.; Bai, X.; Zhai, G.; et al. YiaC and CobB regulate lysine lactylation in Escherichia coli. Nat. Commun. 2022, 13, 6628. [Google Scholar] [CrossRef]
- Wiwanitkit, V. Psychosis and E. coli Infection: A Forgotten Issue. Indian J. Psychol. Med. 2012, 34, 407–408. [Google Scholar] [CrossRef]
- Kleimann, A.; Toto, S.; Eberlein, C.K.; Kielstein, J.T.; Bleich, S.; Frieling, H.; Sieberer, M. Psychiatric Symptoms in Patients with Shiga Toxin-Producing E. coli O104:H4 Induced Haemolytic-Uraemic Syndrome. PLoS ONE 2014, 9, e101839. [Google Scholar] [CrossRef]
- Fouladkhah, A.; Geornaras, I.; Yang, H.; Sofos, J.N. Lactic acid resistance of Shiga toxin-producing Escherichia coli and multidrug-resistant and susceptible Salmonella Typhimurium and Salmonella Newport in meat homogenate. Food Microbiol. 2013, 36, 260–266. [Google Scholar] [CrossRef]
- Graham, K.L.; Carson, C.M.; Ezeoke, A.; Buckley, P.F.; Miller, B.J. Urinary Tract Infections in Acute Psychosis. J. Clin. Psychiatry 2014, 75, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.; McEvoy, J.P.; Miller, B.J. Urinary tract infection, inflammation, and cognition in phase 1 of the Clinical Antipsychotic Trials of Intervention Effectiveness Study. Ann. Clin. Psychiatry 2019, 31, 242–248. [Google Scholar] [PubMed]
- Tadié, J.-M.; Bae, H.-B.; Banerjee, S.; Zmijewski, J.W.; Abraham, E. Differential activation of RAGE by HMGB1 modulates neutrophil-associated NADPH oxidase activity and bacterial killing. Am. J. Physiol. -Cell Physiol. 2012, 302, C249–C256. [Google Scholar] [CrossRef]
- Lin, A.E.; Beasley, F.C.; Olson, J.; Keller, N.; Shalwitz, R.A.; Hannan, T.; Hultgren, S.J.; Nizet, V. Role of Hypoxia Inducible Factor-1α (HIF-1α) in Innate Defense against Uropathogenic Escherichia coli Infection. PLoS Pathog. 2015, 11, e1004818. [Google Scholar] [CrossRef]
- Manosalva, C.; Quiroga, J.; Hidalgo, A.I.; Alarcón, P.; Anseoleaga, N.; Hidalgo, M.A.; Burgos, R.A. Role of Lactate in Inflammatory Processes: Friend or Foe. Front Immunol. 2022, 12, 808799. [Google Scholar] [CrossRef]
- Al-Dujaili, A.H.; Mousa, R.F.; Al-Hakeim, H.K.; Maes, M. High Mobility Group Protein 1 and Dickkopf-Related Protein 1 in Schizophrenia and Treatment-Resistant Schizophrenia: Associations With Interleukin-6, Symptom Domains, and Neurocognitive Impairments. Schizophr. Bull. 2020, 47, 530–541. [Google Scholar] [CrossRef]
- Torrey, E.F.; Yolken, R.H. Toxoplasma gondii and Schizophrenia. Emerg. Infect. Dis. 2003, 9, 1375–1380. [Google Scholar] [CrossRef]
- Zhao, W.; Yu, H.; Liu, X.; Wang, T.; Yao, Y.; Zhou, Q.; Zheng, X.; Tan, F. Systematic identification of the lysine lactylation in the protozoan parasite Toxoplasma gondii. Parasites Vectors 2022, 15, 180. [Google Scholar] [CrossRef]
- Carrillo, G.L.; Ballard, V.A.; Glausen, T.; Boone, Z.; Teamer, J.; Hinkson, C.L.; Wohlfert, E.A.; Blader, I.J.; Fox, M.A. Toxoplasma infection induces microglia-neuron contact and the loss of perisomatic inhibitory synapses. Glia 2020, 68, 1968–1986. [Google Scholar] [CrossRef]
- Obed, C.; Wu, M.; Chen, Y.; An, R.; Cai, H.; Luo, Q.; Yu, L.; Wang, J.; Liu, F.; Shen, J.; et al. Toxoplasma gondii dense granule protein 3 promotes endoplasmic reticulum stress-induced apoptosis by activating the PERK pathway. Parasites Vectors 2022, 15, 276. [Google Scholar] [CrossRef]
- Briceño, M.P.; Nascimento, L.A.C.; Nogueira, N.P.; Barenco, P.V.C.; Ferro, E.A.V.; Rezende-Oliveira, K.; Goulart, L.R.; Alves, P.T.; Barbosa, B.D.F.; Lima, W.R.; et al. Toxoplasma gondii Infection Promotes Epithelial Barrier Dysfunction of Caco-2 Cells. J. Histochem. Cytochem. 2016, 64, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Walter, M.H.; Abele, H.; Plappert, C.F. The Role of Oxytocin and the Effect of Stress During Childbirth: Neurobiological Basics and Implications for Mother and Child. Front. Endocrinol. 2021, 12, 742236. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, M.; Leng, G. Dendritic peptide release and peptide-dependent behaviours. Nat. Rev. Neurosci. 2006, 7, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Sabe, M.; Zhao, N.; Crippa, A.; Strauss, G.P.; Kaiser, S. Intranasal Oxytocin for Negative Symptoms of Schizophrenia: Systematic Review, Meta-Analysis, and Dose-Response Meta-Analysis of Randomized Controlled Trials. Int. J. Neuropsychopharmacol. 2021, 24, 601–614. [Google Scholar] [CrossRef] [PubMed]
- Kéri, S.; Kiss, I.; Kelemen, O. Sharing secrets: Oxytocin and trust in schizophrenia. Soc. Neurosci. 2009, 4, 287–293. [Google Scholar] [CrossRef]
- Matsuzaki, M.; Matsushita, H.; Tomizawa, K.; Matsui, H. Oxytocin: A therapeutic target for mental disorders. J. Physiol. Sci. 2012, 62, 441–444. [Google Scholar] [CrossRef]
- Lin, Y.-T.; Chen, C.-C.; Huang, C.-C.; Nishimori, K.; Hsu, K.-S. Oxytocin stimulates hippocampal neurogenesis via oxytocin receptor expressed in CA3 pyramidal neurons. Nat. Commun. 2017, 8, 537. [Google Scholar] [CrossRef]
- Marlin, B.J.; Froemke, R.C. Oxytocin modulation of neural circuits for social behavior. Dev. Neurobiol. 2016, 77, 169–189. [Google Scholar] [CrossRef]
- Costa-Mattioli, M.; Gobert, D.; Stern, E.; Gamache, K.; Colina, R.; Cuello, C.; Sossin, W.; Kaufman, R.; Pelletier, J.; Rosenblum, K.; et al. eIF2α Phosphorylation Bidirectionally Regulates the Switch from Short- to Long-Term Synaptic Plasticity and Memory. Cell 2007, 129, 195–206. [Google Scholar] [CrossRef]
- Trinh, M.A.; Kaphzan, H.; Wek, R.C.; Pierre, P.; Cavener, D.R.; Klann, E. Brain-Specific Disruption of the eIF2α Kinase PERK Decreases ATF4 Expression and Impairs Behavioral Flexibility. Cell Rep. 2012, 1, 676–688. [Google Scholar] [CrossRef]
- Timberlake, M., II; Dwivedi, Y. Linking unfolded protein response to inflammation and depression: Potential pathologic and therapeutic implications. Mol. Psychiatry 2019, 24, 987–994. [Google Scholar] [CrossRef] [PubMed]
- Mitsuda, T.; Omi, T.; Tanimukai, H.; Sakagami, Y.; Tagami, S.; Okochi, M.; Kudo, T.; Takeda, M. Sigma-1Rs are upregulated via PERK/eIF2α/ATF4 pathway and execute protective function in ER stress. Biochem. Biophys. Res. Commun. 2011, 415, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Cobos, E.J.; Entrena, J.M.; Nieto, F.R.; Cendán, C.M.; Del Pozo, E. Pharmacology and Therapeutic Potential of Sigma1 Receptor Ligands. Curr. Neuropharmacol. 2008, 6, 344–366. [Google Scholar] [CrossRef]
- Almási, N.; Török, S.; Dvorácskó, S.; Tömböly, C.; Csonka, Á.; Baráth, Z.; Murlasits, Z.; Valkusz, Z.; Pósa, A.; Varga, C.; et al. Lessons on the Sigma-1 Receptor in TNBS-Induced Rat Colitis: Modulation of the UCHL-1, IL-6 Pathway. Int. J. Mol. Sci. 2020, 21, 4046. [Google Scholar] [CrossRef]
- Liu, D.; Yang, L.; Liu, P.; Ji, X.; Qi, X.; Wang, Z.; Chi, T.; Zou, L. Sigma–1 receptor activation alleviates blood–brain barrier disruption post cerebral ischemia stroke by stimulating the GDNF–GFRα1–RET pathway. Exp. Neurol. 2021, 347, 113867. [Google Scholar] [CrossRef] [PubMed]
- Kowalczyk, M.; Kowalczyk, E.; Kwiatkowski, P.; Łopusiewicz, Ł.; Talarowska, M.; Sienkiewicz, M. Cellular Response to Unfolded Proteins in Depression. Life 2021, 11, 1376. [Google Scholar] [CrossRef]
- Kamarudin, M.N.A.; Parhar, I. Emerging therapeutic potential of anti-psychotic drugs in the management of human glioma: A comprehensive review. Oncotarget 2019, 10, 3952–3977. [Google Scholar] [CrossRef]
- Liśkiewicz, P.; Kaczmarczyk, M.; Misiak, B.; Wroński, M.; Bąba-Kubiś, A.; Skonieczna-Żydecka, K.; Marlicz, W.; Bieńkowski, P.; Misera, A.; Pełka-Wysiecka, J.; et al. Analysis of gut microbiota and intestinal integrity markers of inpatients with major depressive disorder. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2020, 106, 110076. [Google Scholar] [CrossRef]
- Mao, J.; Hu, Y.; Ruan, L.; Ji, Y.; Lou, Z. Role of endoplasmic reticulum stress in depression (Review). Mol. Med. Rep. 2019, 20, 4774–4780. [Google Scholar] [CrossRef]
- Patel, S.; Sharma, D.; Kalia, K.; Tiwari, V. Crosstalk between endoplasmic reticulum stress and oxidative stress in schizophrenia: The dawn of new therapeutic approaches. Neurosci. Biobehav. Rev. 2017, 83, 589–603. [Google Scholar] [CrossRef]
- Hayashi, T. Conversion of psychological stress into cellular stress response: Roles of the sigma-1 receptor in the process. Psychiatry Clin. Neurosci. 2015, 69, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Safe, S.; Jin, U.-H.; Park, H.; Chapkin, R.S.; Jayaraman, A. Aryl Hydrocarbon Receptor (AHR) Ligands as Selective AHR Modulators (SAhRMs). Int. J. Mol. Sci. 2020, 21, 6654. [Google Scholar] [CrossRef] [PubMed]
- Shackleford, G.; Sampathkumar, N.K.; Hichor, M.; Weill, L.; Meffre, D.; Juricek, L.; Laurendeau, I.; Chevallier, A.; Ortonne, N.; Larousserie, F.; et al. Involvement of Aryl hydrocarbon receptor in myelination and in human nerve sheath tumorigenesis. Proc. Natl. Acad. Sci. USA 2018, 115, E1319–E1328. [Google Scholar] [CrossRef] [PubMed]
- Saraste, M.; Irjala, H.; Airas, L. Expansion of CD56Bright natural killer cells in the peripheral blood of multiple sclerosis patients treated with interferon-beta. Neurol. Sci. 2007, 28, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Feifel, D.; Macdonald, K.; Nguyen, A.; Cobb, P.; Warlan, H.; Galangue, B.; Minassian, A.; Becker, O.; Cooper, J.; Perry, W.; et al. Adjunctive Intranasal Oxytocin Reduces Symptoms in Schizophrenia Patients. Biol. Psychiatry 2010, 68, 678–680. [Google Scholar] [CrossRef] [PubMed]
- Schubert, K.O.; Föcking, M.; Cotter, D.R. Proteomic pathway analysis of the hippocampus in schizophrenia and bipolar affective disorder implicates 14-3-3 signaling, aryl hydrocarbon receptor signaling, and glucose metabolism: Potential roles in GABAergic interneuron pathology. Schizophr. Res. 2015, 167, 64–72. [Google Scholar] [CrossRef]
- Uvnäs-Moberg, K.; Björkstrand, E.; Hillegaart, V.; Ahlenius, S. Oxytocin as a possible mediator of SSRI-induced antidepressant effects. Psychopharmacology 1999, 142, 95–101. [Google Scholar] [CrossRef]
- Galbally, M.; Watson, S.J.; Keelan, J.A.; Spigset, O.; Lewis, A. The relationship between oxytocin blood concentrations and antidepressants over pregnancy and the postpartum. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2020, 109, 110218. [Google Scholar] [CrossRef]
- Humble, M.B.; Bejerot, S. Orgasm, Serotonin Reuptake Inhibition, and Plasma Oxytocin in Obsessive-Compulsive Disorder. Gleaning From a Distant Randomized Clinical Trial. Sex Med. 2016, 4, e145–e155. [Google Scholar] [CrossRef]
- Abbasinazari, M.; Heidari-Kord, M.; Mazaheri-Meybodi, A.; Eshraghi, A.; Bayati, N. Plasma Oxytocin Level and Sexual Dysfunction in Depressed Women Treated by Either Fluoxetine or Citalopram: A Pilot Clinical Trial. Iran. J. Pharm. Res. 2018, 17, 408–414. [Google Scholar]
- Bujanow, W. Is Oxytocin an Anti-Schizophrenic Hormone? Can. Psychiatr. Assoc. J. 1974, 19, 323. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Zhu, J. Innate Lymphoid Cells and Intestinal Inflammatory Disorders. Int. J. Mol. Sci. 2022, 23, 1856. [Google Scholar] [CrossRef] [PubMed]
- Robinette, M.L.; Fuchs, A.; Cortez, V.S.; Lee, J.S.; Wang, Y.; Durum, S.K.; Gilfillan, S.; Colonna, M.; Immunological Genome Consortium. Transcriptional programs define molecular characteristics of innate lymphoid cell classes and subsets. Nat. Immunol. 2015, 16, 306–317. [Google Scholar] [CrossRef]
- Crane, C.A.; Austgen, K.; Haberthur, K.; Hofmann, C.; Moyes, K.W.; Avanesyan, L.; Fong, L.; Campbell, M.J.; Cooper, S.; Oakes, S.A.; et al. Immune evasion mediated by tumor-derived lactate dehydrogenase induction of NKG2D ligands on myeloid cells in glioblastoma patients. Proc. Natl. Acad. Sci. USA 2014, 111, 12823–12828. [Google Scholar] [CrossRef]
- Wensveen, F.M.; Jelenčić, V.; Polić, B. NKG2D: A Master Regulator of Immune Cell Responsiveness. Front. Immunol. 2018, 9, 441. [Google Scholar] [CrossRef] [PubMed]
- Borovcanin, M.M.; Minic Janicijevic, S.; Jovanovic, I.P.; Gajovic, N.M.; Jurisevic, M.M.; Arsenijevic, N.N. Type 17 Immune Response Facilitates Progression of Inflammation and Correlates with Cognition in Stable Schizophrenia. Diagnostics 2020, 10, 926. [Google Scholar] [CrossRef] [PubMed]
- Matusevicius, D.; Kivisäkk, P.; He, B.; Kostulas, N.; Özenci, V.; Fredrikson, S.; Link, H. Interleukin-17 mRNA expression in blood and CSF mononuclear cells is augmented in multiple sclerosis. Mult. Scler. J. 1999, 5, 101–104. [Google Scholar] [CrossRef] [PubMed]
- Dawidowski, B.; Górniak, A.; Podwalski, P.; Lebiecka, Z.; Misiak, B.; Samochowiec, J. The Role of Cytokines in the Pathogenesis of Schizophrenia. J. Clin. Med. 2021, 10, 3849. [Google Scholar] [CrossRef]
- Al-Asmary, S.M.; Kadasah, S.; Arfin, M.; Tariq, M.; Al-Asmari, A. Genetic variants of interleukin-10 gene promoter are associated with schizophrenia in Saudi patients: A case-control study. North Am. J. Med. Sci. 2014, 6, 558–565. [Google Scholar] [CrossRef]
- Warre-Cornish, K.; Perfect, L.; Nagy, R.; Duarte, R.R.R.; Reid, M.J.; Raval, P.; Mueller, A.; Evans, A.L.; Couch, A.; Ghevaert, C.; et al. Interferon-γ signaling in human iPSC–derived neurons recapitulates neurodevelopmental disorder phenotypes. Sci. Adv. 2020, 6, eaay9506. [Google Scholar] [CrossRef]
- Ramirez, J.-M.; Brembilla, N.C.; Sorg, O.; Chicheportiche, R.; Matthes, T.; Dayer, J.-M.; Saurat, J.-H.; Roosnek, E.; Chizzolini, C. Activation of the aryl hydrocarbon receptor reveals distinct requirements for IL-22 and IL-17 production by human T helper cells. Eur. J. Immunol. 2010, 40, 2450–2459. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.C.; Chang, L.H.; Huang, S.S.; Huang, Y.J.; Chih, C.L.; Kuo, H.C.; Lee, Y.H.; Lee, I.H. Aryl hydrocarbon receptor modulates stroke-induced astrogliosis and neurogenesis in the adult mouse brain. J. Neuroinflam. 2019, 16, 187. [Google Scholar] [CrossRef]
- van Venrooij, J.A.E.M.; Fluitman, S.B.A.H.A.; Lijmer, J.G.; Kavelaars, A.; Heijnen, C.J.; Westenberg, H.G.M.; Kahn, R.S.; Wied, C.C.G.-D. Impaired Neuroendocrine and Immune Response to Acute Stress in Medication-Naive Patients With a First Episode of Psychosis. Schizophr. Bull. 2012, 38, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Gross, C.C.; Ahmetspahic, D.; Ruck, T.; Schulte-Mecklenbeck, A.; Schwarte, K.; Jörgens, S.; Scheu, S.; Windhagen, S.; Graefe, B.; Melzer, N.; et al. Alemtuzumab treatment alters circulating innate immune cells in multiple sclerosis. Neurol. -Neuroimmunol. Neuroinflamm. 2016, 3, e289. [Google Scholar] [CrossRef]
- Chu, C.-S.; Li, D.-J.; Chu, C.-L.; Wu, C.-C.; Lu, T. Decreased IL-1ra and NCAM-1/CD56 Serum Levels in Unmedicated Patients with Schizophrenia Before and After Antipsychotic Treatment. Psychiatry Investig. 2018, 15, 727–732. [Google Scholar] [CrossRef] [PubMed]
- Postal, B.G.; Ghezzal, S.; Aguanno, D.; André, S.; Garbin, K.; Genser, L.; Brot-Laroche, E.; Poitou, C.; Soula, H.; Leturque, A.; et al. AhR activation defends gut barrier integrity against damage occurring in obesity. Mol. Metab. 2020, 39, 101007. [Google Scholar] [CrossRef]
- Ghiboub, M.; Verburgt, C.M.; Sovran, B.; Benninga, M.A.; de Jonge, W.J.; Van Limbergen, J.E. Nutritional Therapy to Modulate Tryptophan Metabolism and Aryl Hydrocarbon-Receptor Signaling Activation in Human Diseases. Nutrients 2020, 12, 2846. [Google Scholar] [CrossRef]
- Marinelli, L.; Martin-Gallausiaux, C.; Bourhis, J.-M.; Béguet-Crespel, F.; Blottière, H.M.; Lapaque, N. Identification of the novel role of butyrate as AhR ligand in human intestinal epithelial cells. Sci. Rep. 2019, 9, 643. [Google Scholar] [CrossRef]
- Tagliabue, S.G.; Faber, S.C.; Motta, S.; Denison, M.S.; Bonati, L. Modeling the binding of diverse ligands within the Ah receptor ligand binding domain. Sci. Rep. 2019, 9, 10693. [Google Scholar] [CrossRef]
- Wu, K.K. Cytoguardin: A Tryptophan Metabolite against Cancer Growth and Metastasis. Int. J. Mol. Sci. 2021, 22, 4490. [Google Scholar] [CrossRef]
- Peyraud, F.; Guegan, J.-P.; Bodet, D.; Cousin, S.; Bessede, A.; Italiano, A. Targeting Tryptophan Catabolism in Cancer Immunotherapy Era: Challenges and Perspectives. Front. Immunol. 2022, 13, 807271. [Google Scholar] [CrossRef] [PubMed]
- Juhász, C.; Nahleh, Z.; Zitron, I.; Chugani, D.C.; Janabi, M.Z.; Bandyopadhyay, S.; Ali-Fehmi, R.; Mangner, T.J.; Chakraborty, P.K.; Mittal, S.; et al. Tryptophan metabolism in breast cancers: Molecular imaging and immunohistochemistry studies. Nucl. Med. Biol. 2012, 39, 926–932. [Google Scholar] [CrossRef] [PubMed]
- Papadimitriou, N.; Gunter, M.J.; Murphy, N.; Gicquiau, A.; Achaintre, D.; Brezina, S.; Gumpenberger, T.; Baierl, A.; Ose, J.; Geijsen, A.J.M.R.; et al. Circulating tryptophan metabolites and risk of colon cancer: Results from case-control and prospective cohort studies. Int. J. Cancer 2021, 149, 1659–1669. [Google Scholar] [CrossRef] [PubMed]
- Pessione, E. Lactic acid bacteria contribution to gut microbiota complexity: Lights and shadows. Front Cell Infect Microbiol. 2012, 2, 86. [Google Scholar] [CrossRef]
- Takeda, T.; Komiya, Y.; Koga, T.; Ishida, T.; Ishii, Y.; Kikuta, Y.; Nakaya, M.; Kurose, H.; Yokomizo, T.; Shimizu, T.; et al. Dioxin-induced increase in leukotriene B4 biosynthesis through the aryl hydrocarbon receptor and its relevance to hepatotoxicity owing to neutrophil infiltration. J. Biol. Chem. 2017, 292, 10586–10599. [Google Scholar] [CrossRef]
- Yu, M.; Wang, Q.; Ma, Y.; Li, L.; Yu, K.; Zhang, Z.; Chen, G.; Li, X.; Xiao, W.; Xu, P.; et al. Aryl Hydrocarbon Receptor Activation Modulates Intestinal Epithelial Barrier Function by Maintaining Tight Junction Integrity. Int. J. Biol. Sci. 2018, 14, 69–77. [Google Scholar] [CrossRef]
- Goya-Jorge, E.; Gonza, I.; Bondue, P.; Douny, C.; Taminiau, B.; Daube, G.; Scippo, M.-L.; Delcenserie, V. Human Adult Microbiota in a Static Colon Model: AhR Transcriptional Activity at the Crossroads of Host–Microbe Interaction. Foods 2022, 11, 1946. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, Y.; Wang, H.; Liu, L.; Li, W.; Xie, P. The regulatory effects of lactic acid on neuropsychiatric disorders. Discov. Ment. Health 2022, 2, 8. [Google Scholar] [CrossRef]
- Guo, N.; Jiang, Y.W.; Song, X.R.; Li, Y.Y.; Liu, Z.M.; Fu, Y.J. Effect of Bacillus natto solid-state fermentation on the functional constituents and properties of Ginkgo seeds. J. Food Biochem. 2019, 43, e12820. [Google Scholar] [CrossRef]
- Zhang, Y.; Anoopkumar-Dukie, S.; Davey, A.K. SIRT1 and SIRT2 Modulators: Potential Anti-Inflammatory Treatment for Depression? Biomolecules 2021, 11, 353. [Google Scholar] [CrossRef]
- Vasconcelos, G.S.; Dos Santos Júnior, M.A.; Monte, A.S.; da Silva, F.E.R.; Lima, C.N.C.; Moreira Lima Neto, A.B.; Medeiros, I.D.S.; Teixeira, A.L.; de Lucena, D.F.; Vasconcelos, S.M.M.; et al. Low-dose candesartan prevents schizophrenia-like behavioral alterations in a neurodevelopmental two-hit model of schizophrenia. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2021, 111, 110348. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Feng, F.; Wu, J.; Fan, S.; Han, J.; Wang, S.; Yang, L.; Liu, W.; Wang, C.; Xu, K. Demethylzeylasteral targets lactate by inhibiting histone lactylation to suppress the tumorigenicity of liver cancer stem cells. Pharmacol. Res. 2022, 181, 106270. [Google Scholar] [CrossRef] [PubMed]
- Sidrauski, C.; McGeachy, A.M.; Ingolia, N.T.; Walter, P. The small molecule ISRIB reverses the effects of eIF2α phosphorylation on translation and stress granule assembly. Elife 2015, 4, e05033. [Google Scholar] [CrossRef] [PubMed]
- Hritcu, L.; Ionita, R.; Postu, P.A.; Gupta, G.K.; Turkez, H.; Lima, T.C.; Carvalho, C.U.S.; de Sousa, D.P. Antidepressant Flavonoids and Their Relationship with Oxidative Stress. Oxidative Med. Cell. Longev. 2017, 2017, 5762172. [Google Scholar] [CrossRef] [PubMed]
- Hayashi-Takagi, A.; Araki, Y.; Nakamura, M.; Vollrath, B.; Duron, S.G.; Yan, Z.; Kasai, H.; Huganir, R.L.; Campbell, D.A.; Sawa, A. PAKs inhibitors ameliorate schizophrenia-associated dendritic spine deterioration in vitro and in vivo during late adolescence. Proc. Natl. Acad. Sci. USA 2014, 111, 6461–6466. [Google Scholar] [CrossRef]
- Gil-Ad, I.; Amit, B.H.; Hayardeni, L.; Tarasenko, I.; Taler, M.; Gueta, R.U.; Weizman, A. Effects of the Anti-Multiple Sclerosis Immunomodulator Laquinimod on Anxiety and Depression in Rodent Behavioral Models. J. Mol. Neurosci. 2014, 55, 552–560. [Google Scholar] [CrossRef]
- Mao, N.; Fan, Y.; Liu, W.; Yang, H.; Yang, Y.; Li, Y.; Jin, F.; Li, T.; Yang, X.; Gao, X.; et al. Oxamate Attenuates Glycolysis and ER Stress in Silicotic Mice. Int. J. Mol. Sci. 2022, 23, 3013. [Google Scholar] [CrossRef]
- Khan, R.S.; Yu, C.; Kastin, A.J.; He, Y.; Ehrensing, R.H.; Hsuchou, H.; Stone, K.; Pan, W. Brain Activation by Peptide Pro-Leu-Gly-NH2 (MIF-1). Int. J. Pept. 2010, 2010, 537639. [Google Scholar] [CrossRef]
- Choi, E.-Y.; Lee, H.; Dingle, R.W.C.; Kim, K.B.; Swanson, H.I. Development of Novel CH223191-Based Antagonists of the Aryl Hydrocarbon Receptor. Mol. Pharmacol. 2011, 81, 3–11. [Google Scholar] [CrossRef]
- Deiab, S.; Mazzio, E.; Eyunni, S.; McTier, O.; Mateeva, N.; Elshami, F.; Soliman, K.F.A. 1,2,3,4,6-Penta-O-galloylglucose within Galla Chinensis Inhibits Human LDH-A and Attenuates Cell Proliferation in MDA-MB-231 Breast Cancer Cells. Evid. -Based Complement. Altern. Med. 2015, 2015, 276946. [Google Scholar] [CrossRef]
- Cid-Jofré, V.; Moreno, M.; Reyes-Parada, M.; Renard, G.M. Role of Oxytocin and Vasopressin in Neuropsychiatric Disorders: Therapeutic Potential of Agonists and Antagonists. Int. J. Mol. Sci. 2021, 22, 12077. [Google Scholar] [CrossRef] [PubMed]
- Ala, M.; Ghasemi, M.; Mohammad Jafari, R.; Dehpour, A.R. Beyond its anti-migraine properties, sumatriptan is an anti-inflammatory agent: A systematic review. Drug Dev. Res. 2021, 82, 896–906. [Google Scholar] [CrossRef] [PubMed]
- Manerba, M.; Vettraino, M.; Fiume, L.; Di Stefano, G.; Sartini, A.; Giacomini, E.; Buonfiglio, R.; Roberti, M.; Recanatini, M. Galloflavin (CAS 568-80-9): A Novel Inhibitor of Lactate Dehydrogenase. Chemmedchem 2011, 7, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Uba, A.I.; Radicella, C.; Readmond, C.; Scorese, N.; Liao, S.; Liu, H.; Wu, C. Binding of agonist WAY-267,464 and antagonist WAY-methylated to oxytocin receptor probed by all-atom molecular dynamics simulations. Life Sci. 2020, 252, 117643. [Google Scholar] [CrossRef] [PubMed]
- Vukicevic, M.; Chalandon, Y.; Helg, C.; Matthes, T.; Dantin, C.; Huard, B.; Chizzolini, C.; Passweg, J.; Roosnek, E. CD56bright NK cells after hematopoietic stem cell transplantation are activated mature NK cells that expand in patients with low numbers of T cells. Eur. J. Immunol. 2010, 40, 3246–3254. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, J.; Zhu, K.; Wang, D.; Zhao, X.; Zhang, H.; Wu, S.; Wang, Y.; Wang, J. Microglial aryl hydrocarbon receptor enhances phagocytic function via SYK and promotes remyelination in the cuprizone mouse model of demyelination. J. Neuroinflamm. 2023, 20, 83. [Google Scholar] [CrossRef]
- Fehsel, K.; Schwanke, K.; Kappel, B.; Fahimi, E.; Meisenzahl-Lechner, E.; Esser, C.; Hemmrich, K.; Haarmann-Stemmann, T.; Kojda, G.; Lange-Asschenfeldt, C. Activation of the aryl hydrocarbon receptor by clozapine induces preadipocyte differentiation and contributes to endothelial dysfunction. J. Psychopharmacol. 2022, 36, 191–201. [Google Scholar] [CrossRef]
- Korac, K.; Rajasekaran, D.; Sniegowski, T.; Schniers, B.; Ibrahim, A.; Bhutia, Y.D. Carbidopa, an activator of aryl hydrocarbon receptor, suppresses IDO1 expression in pancreatic cancer and decreases tumor growth. Biochem. J. 2022, 479, 1807–1824. [Google Scholar] [CrossRef]
- Juricek, L.; Coumoul, X. The Aryl Hydrocarbon Receptor and the Nervous System. Int. J. Mol. Sci. 2018, 19, 2504. [Google Scholar] [CrossRef]
- Robichon, K.; Sondhauss, S.; Jordan, T.W.; Keyzers, R.A.; Connor, B.; La Flamme, A.C. Localisation of clozapine during experimental autoimmune encephalomyelitis and its impact on dopamine and its receptors. Sci. Rep. 2021, 11, 2966. [Google Scholar] [CrossRef]
- Mosca, L.; Mantero, V.; Penco, S.; La Mantia, L.; De Benedetti, S.; Marazzi, M.R.; Spreafico, C.; Erminio, C.; Grassi, L.; Lando, G.; et al. HLA-DRB1*15 association with multiple sclerosis is confirmed in a multigenerational Italian family. Funct. Neurol. 2017, 32, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, J.I.; Jarskog, L.F.; Hilliard, C.; Alfirevic, A.; Duncan, L.; Fourches, D.; Huang, H.; Lek, M.; Neale, B.M.; Ripke, S.; et al. Clozapine-induced agranulocytosis is associated with rare HLA-DQB1 and HLA-B alleles. Nat. Commun. 2014, 5, 4757. [Google Scholar] [CrossRef] [PubMed]
- Sfera, A.; Hazan, S.; Klein, C.; del Campo, C.M.Z.-M.; Sasannia, S.; Anton, J.J.; Rahman, L.; Andronescu, C.V.; Sfera, D.O.; Kozlakidis, Z.; et al. Microbial Translocation Disorders: Assigning an Etiology to Idiopathic Illnesses. Appl. Microbiol. 2023, 3, 212–240. [Google Scholar] [CrossRef]
- Johansson, V.; Jakobsson, J.; Fortgang, R.G.; Zetterberg, H.; Blennow, K.; Cannon, T.D.; Hultman, C.M.; Wetterberg, L.; Landén, M. Cerebrospinal fluid microglia and neurodegenerative markers in twins concordant and discordant for psychotic disorders. Eur. Arch. Psychiatry Clin. Neurosci. 2016, 267, 391–402. [Google Scholar] [CrossRef] [PubMed]
- Weber, N.S.; Gressitt, K.L.; Cowan, D.N.; Niebuhr, D.W.; Yolken, R.H.; Severance, E.G. Monocyte activation detected prior to a diagnosis of schizophrenia in the US Military New Onset Psychosis Project (MNOPP). Schizophr. Res. 2018, 197, 465–469. [Google Scholar] [CrossRef] [PubMed]
- Severance, E.G.; Gressitt, K.L.; Stallings, C.R.; Origoni, A.E.; Khushalani, S.; Leweke, F.M.; Dickerson, F.B.; Yolken, R.H. Discordant patterns of bacterial translocation markers and implications for innate immune imbalances in schizophrenia. Schizophr. Res. 2013, 148, 130–137. [Google Scholar] [CrossRef]
- Kamaeva, D.A.; Smirnova, L.P.; Vasilieva, S.N.; Kazantseva, D.V.; Vasilieva, A.R.; Ivanova, S.A. Catalytic Antibodies in Bipolar Disorder: Serum IgGs Hydrolyze Myelin Basic Protein. Int. J. Mol. Sci. 2022, 23, 7397. [Google Scholar] [CrossRef]
- von Zedtwitz, K.; Matteit, I.; Michel, M.; Feige, B.; Runge, K.; Denzel, D.; Schlump, A.; Nickel, K.; Schiele, M.A.; Berger, B.; et al. Anti-MOG autoantibody-associated schizophreniform psychosis. Acta Neuropsychiatr. 2021, 34, 47–54. [Google Scholar] [CrossRef]
- Ponomarenko, N.; Durova, O.M.; Vorobiev, I.I.; Belogurov, A.A.; Telegin, G.B.; Suchkov, S.V.; Misikov, V.K.; Morse, H.C., 3rd; Gabibov, A.G. Catalytic activity of autoantibodies toward myelin basic protein correlates with the scores on the multiple sclerosis expanded disability status scale. Immunol. Lett. 2006, 103, 45–50. [Google Scholar] [CrossRef]
- Schott, K.; Schaefer, J.-E.; Richartz, E.; Batra, A.; Eusterschulte, B.; Klein, R.; Berg, P.A.; Bartels, M.; Mann, K.; Buchkremer, G. Autoantibodies to serotonin in serum of patients with psychiatric disorders. Psychiatry Res. 2003, 121, 51–57. [Google Scholar] [CrossRef]
- Zong, S.; Hoffmann, C.; Mané-Damas, M.; Molenaar, P.; Losen, M.; Martinez-Martinez, P. Neuronal Surface Autoantibodies in Neuropsychiatric Disorders: Are There Implications for Depression? Front. Immunol. 2017, 8, 752. [Google Scholar] [CrossRef] [PubMed]
- Westall, F.C. Molecular Mimicry Revisited: Gut Bacteria and Multiple Sclerosis. J. Clin. Microbiol. 2006, 44, 2099–2104. [Google Scholar] [CrossRef] [PubMed]
- Brundin, L.; Erhardt, S.; Bryleva, E.Y.; Achtyes, E.D.; Postolache, T.T. The role of inflammation in suicidal behaviour. Acta Psychiatr. Scand. 2015, 132, 192–203. [Google Scholar] [CrossRef] [PubMed]
- Ohlsson, L.; Gustafsson, A.; Lavant, E.; Suneson, K.; Brundin, L.; Westrin, Å.; Ljunggren, L.; Lindqvist, D. Leaky gut biomarkers in depression and suicidal behavior. Acta Psychiatr. Scand. 2018, 139, 185–193, Erratum in Acta Psychiatr. Scand. 2020, 142, 423. [Google Scholar] [CrossRef] [PubMed]
- Ahrens, A.P.; Sanchez-Padilla, D.E.; Drew, J.C.; Oli, M.W.; Roesch, L.F.W.; Triplett, E.W. Saliva microbiome, dietary, and genetic markers are associated with suicidal ideation in university students. Sci. Rep. 2022, 12, 14306. [Google Scholar] [CrossRef]
- Eyzaguirre-Velásquez, J.; González-Toro, M.P.; González-Arancibia, C.; Escobar-Luna, J.; Beltrán, C.J.; Bravo, J.A.; Julio-Pieper, M. Sertraline and Citalopram Actions on Gut Barrier Function. Dig. Dis. Sci. 2020, 66, 3792–3802. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, T.; He, L.; Fu, J.-Y.; Deng, H.-X.; Xue, X.-L.; Chen, B.-T. Bacterial Translocation Associates With Aggression in Schizophrenia Inpatients. Front. Syst. Neurosci. 2021, 15, 704069. [Google Scholar] [CrossRef]
- Bulgart, H.R.; Neczypor, E.W.; Wold, L.E.; Mackos, A.R. Microbial involvement in Alzheimer disease development and progression. Mol. Neurodegener. 2020, 15, 42. [Google Scholar] [CrossRef]
- Liang, L.; Liu, P.; Deng, Y.; Li, J.; Zhao, S. L-lactate inhibits lipopolysaccharide-induced inflammation of microglia in the hippocampus. Int. J. Neurosci. 2022, 23, 1–8. [Google Scholar] [CrossRef]
- Guglielmetti, S.; Bernardi, S.; Del Bo’, C.; Cherubini, A.; Porrini, M.; Gargari, G.; Hidalgo-Liberona, N.; Gonzalez-Dominguez, R.; Peron, G.; Zamora-Ros, R.; et al. Effect of a polyphenol-rich dietary pattern on intestinal permeability and gut and blood microbiomics in older subjects: Study protocol of the MaPLE randomised controlled trial. BMC Geriatr. 2020, 20, 77. [Google Scholar] [CrossRef]
- Figueira, I.; Garcia, G.; Pimpão, R.C.; Terrasso, A.P.; Costa, I.; Almeida, A.F.; Tavares, L.; Pais, T.F.; Pinto, P.; Ventura, M.R.; et al. Polyphenols journey through blood-brain barrier towards neuronal protection. Sci. Rep. 2017, 7, 11456. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Tang, W.; Zhao, J.; Fan, W.; Zhang, Y.; Zhang, C. A Comprehensive Analysis of the Effect of SIRT1 Variation on the Risk of Schizophrenia and Depressive Symptoms. Front. Genet. 2020, 11, 832. [Google Scholar] [CrossRef] [PubMed]
- Iacono, L.L.; Visco-Comandini, F.; Valzania, A.; Viscomi, M.T.; Coviello, M.; Giampà, A.; Roscini, L.; Bisicchia, E.; Siracusano, A.; Troisi, A.; et al. Adversity in childhood and depression: Linked through SIRT1. Transl. Psychiatry 2015, 5, e629. [Google Scholar] [CrossRef] [PubMed]
- Zu, H.; Li, C.; Dai, C.; Pan, Y.; Ding, C.; Sun, H.; Zhang, X.; Yao, X.; Zang, J.; Mo, X. SIRT2 functions as a histone delactylase and inhibits the proliferation and migration of neuroblastoma cells. Cell Discov. 2022, 8, 54. [Google Scholar] [CrossRef] [PubMed]
- Peña, C.J.; Nestler, E.J. Progress in Epigenetics of Depression. Prog. Mol. Biol. Transl. Sci. 2018, 157, 41–66. [Google Scholar] [CrossRef]
- Cui, B.; Luo, Y.; Tian, P.; Peng, F.; Lu, J.; Yang, Y.; Su, Q.; Liu, B.; Yu, J.; Luo, X.; et al. Stress-induced epinephrine enhances lactate dehydrogenase A and promotes breast cancer stem-like cells. J. Clin. Investig. 2019, 129, 1030–1046. [Google Scholar] [CrossRef]
- Zhao, Z.; Han, F.; Yang, S.; Wu, J.; Zhan, W. Oxamate-mediated inhibition of lactate dehydrogenase induces protective autophagy in gastric cancer cells: Involvement of the Akt–mTOR signaling pathway. Cancer Lett. 2015, 358, 17–26. [Google Scholar] [CrossRef]
- Ren, Y.-Y.; Zhang, X.-R.; Li, T.-N.; Zeng, Y.-J.; Wang, J.; Huang, Q.-W. Galla Chinensis, a Traditional Chinese Medicine: Comprehensive review of botany, traditional uses, chemical composition, pharmacology and toxicology. J. Ethnopharmacol. 2021, 278, 114247. [Google Scholar] [CrossRef]
- Rahnasto-Rilla, M.; Järvenpää, J.; Huovinen, M.; Schroderus, A.-M.; Ihantola, E.-L.; Küblbeck, J.; Khadeer, M.; Moaddel, R.; Lahtela-Kakkonen, M. Effects of galloflavin and ellagic acid on sirtuin 6 and its anti-tumorigenic activities. Biomed. Pharmacother. 2020, 131, 110701. [Google Scholar] [CrossRef]
- Mao, Q.; Gong, X.; Zhou, C.; Tu, Z.; Zhao, L.; Wang, L.; Wang, X.; Sun, L.; Xia, J.; Lian, B.; et al. Up-regulation of SIRT6 in the hippocampus induced rats with depression-like behavior via the block Akt/GSK3β signaling pathway. Behav. Brain Res. 2017, 323, 38–46. [Google Scholar] [CrossRef]
- Sever, I.H.; Ozkul, B.; Erisik Tanriover, D.; Ozkul, O.; Elgormus, C.S.; Gur, S.G.; Sogut, I.; Uyanikgil, Y.; Cetin, E.O.; Erbas, O. Protective effect of oxytocin through its anti-inflammatory and antioxidant role in a model of sepsis-induced acute lung injury: Demonstrated by CT and histological findings. Exp. Lung Res. 2021, 47, 426–435. [Google Scholar] [CrossRef] [PubMed]
- Erbaş, O.; Altuntaş, İ. Oxytocin and Neuroprotective Effects; IntechOpen: London, UK, 2021. [Google Scholar] [CrossRef]
- Gonzalez, A.; Hammock, E.A.D. Oxytocin and microglia in the development of social behaviour. Philos. Trans. R. Soc. B Biol. Sci. 2022, 377, 20210059. [Google Scholar] [CrossRef] [PubMed]
- Labandeira-García, J.L.; Garrido-Gil, P.; Rodriguez-Pallares, J.; Valenzuela, R.; Borrajo, A.; Rodríguez-Perez, A.I. Brain renin-angiotensin system and dopaminergic cell vulnerability. Front. Neuroanat. 2014, 8, 67. [Google Scholar] [CrossRef] [PubMed]
- Srinivasa, S.; Aulinas, A.; O’malley, T.; Maehler, P.; Adler, G.K.; Grinspoon, S.K.; Lawson, E.A. Oxytocin response to controlled dietary sodium and angiotensin II among healthy individuals. Am. J. Physiol. Endocrinol. Metab. 2018, 315, E671–E675. [Google Scholar] [CrossRef] [PubMed]
- Gard, P.R.; Daw, P.; Mashhour, Z.S.; Tran, P. Interactions of angiotensin IV and oxytocin on behaviour in mice. J. Renin-Angiotensin-Aldosterone Syst. 2007, 8, 133–138. [Google Scholar] [CrossRef]
- Kokare, D.M.; Singru, P.S.; Dandekar, M.P.; Chopde, C.T.; Subhedar, N.K. Involvement of alpha-melanocyte stimulating hormone (α-MSH) in differential ethanol exposure and withdrawal related depression in rat: Neuroanatomical–behavioral correlates. Brain Res. 2008, 1216, 53–67. [Google Scholar] [CrossRef]
- Bialesova, L.; Novotna, A.; Macejova, D.; Brtko, J.; Dvorak, Z. Agonistic effect of selected isoflavones on arylhydrocarbon receptor in a novel AZ-AhR transgenic gene reporter human cell line. Gen. Physiol. Biophys. 2015, 34, 331–334. [Google Scholar] [CrossRef][Green Version]
- Venkataramaiah, C.; Payani, S.; Priya, B.L.; Pradeepkiran, J.A. Therapeutic potentiality of a new flavonoid against ketamine induced glutamatergic dysregulation in schizophrenia: In vivo and in silico approach. Biomed. Pharmacother. 2021, 138, 111453. [Google Scholar] [CrossRef]
- Křížová, L.; Dadáková, K.; Kašparovská, J.; Kašparovský, T. Isoflavones. Molecules 2019, 24, 1076. [Google Scholar] [CrossRef]
- Long, T.; He, W.; Pan, Q.; Zhang, S.; Zhang, D.; Qin, G.; Chen, L.; Zhou, J. Microglia P2X4R-BDNF signalling contributes to central sensitization in a recurrent nitroglycerin-induced chronic migraine model. J. Headache Pain 2020, 21, 4. [Google Scholar] [CrossRef]
- Smith, M.; Calabresi, P.; Bhargava, P. Dimethyl fumarate treatment alters NK cell function in multiple sclerosis. Eur. J. Immunol. 2017, 48, 380–383. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, N.; Ødum, N.; Ursø, B.; Lanier, L.L.; Spee, P. Cytotoxicity of CD56bright NK Cells towards Autologous Activated CD4+ T Cells Is Mediated through NKG2D, LFA-1 and TRAIL and Dampened via CD94/NKG2A. PLoS ONE 2012, 7, e31959. [Google Scholar] [CrossRef] [PubMed]
- Ruggeri, L.; Capanni, M.; Urbani, E.; Perruccio, K.; Shlomchik, W.D.; Tosti, A.; Posati, S.; Rogaia, D.; Frassoni, F.; Aversa, F.; et al. Effectiveness of Donor Natural Killer Cell Alloreactivity in Mismatched Hematopoietic Transplants. Science 2002, 295, 2097–2100. [Google Scholar] [CrossRef] [PubMed]
- Monji, A.; Kato, T.; Kanba, S. Cytokines and schizophrenia: Microglia hypothesis of schizophrenia. Psychiatry Clin. Neurosci. 2009, 63, 257–265. [Google Scholar] [CrossRef]
- Wake, H.; Moorhouse, A.J.; Jinno, S.; Kohsaka, S.; Nabekura, J. Resting Microglia Directly Monitor the Functional State of Synapses In Vivo and Determine the Fate of Ischemic Terminals. J. Neurosci. 2009, 29, 3974–3980. [Google Scholar] [CrossRef]
- Jha, M.K.; Jo, M.; Kim, J.-H.; Suk, K. Microglia-Astrocyte Crosstalk: An Intimate Molecular Conversation. Neurosci. 2018, 25, 227–240. [Google Scholar] [CrossRef]
- Correa, F.G.; Hernangómez, M.; Guaza, C. Understanding Microglia–Neuron Cross Talk: Relevance of the Microglia–Neuron Cocultures. Methods Mol. Biol. 2013, 1041, 215–229. [Google Scholar] [CrossRef]
- Mills, J.; Ladner, L.; Soliman, E.; Leonard, J.; Morton, P.D.; Theus, M.H. Cross-Talk and Subset Control of Microglia and Associated Myeloid Cells in Neurological Disorders. Cells 2022, 11, 3364. [Google Scholar] [CrossRef]
- Yang, S.; Qin, C.; Hu, Z.W.; Zhou, L.Q.; Yu, H.H.; Chen, M.; Bosco, D.B.; Wang, W.; Wu, L.J.; Tian, D.S. Microglia reprogram metabolic profiles for phenotype and function changes in central nervous system. Neurobiol. Dis. 2021, 152, 105290. [Google Scholar] [CrossRef]
- Nijland, P.G.; Michailidou, I.; Witte, M.E.; Mizee, M.R.; van der Pol, S.M.; van Het Hof, B.; Reijerkerk, A.; Pellerin, L.; van der Valk, P.; de Vries, H.E.; et al. Cellular distribution of glucose and monocarboxylate transporters in human brain white matter and multiple sclerosis lesions. Glia 2014, 62, 1125–1141. [Google Scholar] [CrossRef]
- Goyal, M.S.; Vlassenko, A.G.; Blazey, T.M.; Su, Y.; Couture, L.E.; Durbin, T.J.; Bateman, R.J.; Benzinger, T.L.-S.; Morris, J.C.; Raichle, M.E. Loss of Brain Aerobic Glycolysis in Normal Human Aging. Cell Metab. 2017, 26, 353–360.e3. [Google Scholar] [CrossRef] [PubMed]
- Hipkiss, A.R. Aging, Alzheimer’s Disease and Dysfunctional Glycolysis; Similar Effects of Too Much and Too Little. Aging Dis. 2019, 10, 1328–1331. [Google Scholar] [CrossRef] [PubMed]
- Zuccoli, G.S.; Guest, P.C.; Martins-De-Souza, D. Effects on Glial Cell Glycolysis in Schizophrenia: An Advanced Aging Phenotype? Adv. Exp. Med. Biol. 2019, 1178, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Lv, X.; Thompson, E.W.; Ostrikov, K.K. Histone lactylation: Epigenetic mark of glycolytic switch. Trends Genet. 2021, 38, 124–127. [Google Scholar] [CrossRef]
- Szeto, A.; Sun-Suslow, N.; Mendez, A.J.; Hernandez, R.I.; Wagner, K.V.; McCabe, P.M. Regulation of the macrophage oxytocin receptor in response to inflammation. Am. J. Physiol. Endocrinol. Metab. 2017, 312, E183–E189. [Google Scholar] [CrossRef]
- Kaneko, Y.; Pappas, C.; Tajiri, N.; Borlongan, C.V. Oxytocin modulates GABAAR subunits to confer neuroprotection in stroke in vitro. Sci. Rep. 2016, 6, 35659. [Google Scholar] [CrossRef]
- Buonfiglioli, A.; Hambardzumyan, D. Macrophages and microglia: The cerberus of glioblastoma. Acta Neuropathol. Commun. 2021, 9, 54. [Google Scholar] [CrossRef]
- Genuis, S.J. Toxic causes of mental illness are overlooked. Neurotoxicology 2008, 29, 1147–1149. [Google Scholar] [CrossRef]
- Krüger, T.; Long, M.; Bonefeld-Jørgensen, E.C. Plastic components affect the activation of the aryl hydrocarbon and the androgen receptor. Toxicology 2008, 246, 112–123. [Google Scholar] [CrossRef]
- Scott, S.A.; Fu, J.; Chang, P.V. Microbial tryptophan metabolites regulate gut barrier function via the aryl hydrocarbon receptor. Proc. Natl. Acad. Sci. USA 2020, 117, 19376–19387. [Google Scholar] [CrossRef]
- Zhang, J.; Velmeshev, D.; Hashimoto, K.; Huang, Y.; Hofmann, J.W.; Shi, X.; Chen, J.; Leidal, A.M.; Dishart, J.G.; Cahill, M.K.; et al. Neurotoxic microglia promote TDP-43 proteinopathy in progranulin deficiency. Nature 2020, 588, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Piccioni, G.; Mango, D.; Saidi, A.; Corbo, M.; Nisticò, R. Targeting Microglia-Synapse Interactions in Alzheimer’s Disease. Int. J. Mol. Sci. 2021, 22, 2342. [Google Scholar] [CrossRef] [PubMed]
- Gober, R.; Ardalan, M.; Shiadeh, S.M.J.; Duque, L.; Garamszegi, S.P.; Ascona, M.; Barreda, A.; Sun, X.; Mallard, C.; Vontell, R.T. Microglia activation in postmortem brains with schizophrenia demonstrates distinct morphological changes between brain regions. Brain Pathol. 2021, 32, e13003. [Google Scholar] [CrossRef] [PubMed]


| Lactate | OXT | AhR | References |
|---|---|---|---|
| L-lactate | Intranasal OXT | Flavonoids | [114,168,169] |
| Lactylation inhibitors | ARBs | Nattokinase | [169,170,171] |
| Demethylzeylasteral (DML) | ISRIB | Isoflavones | [172,173,174] |
| Sirtuins | PAK inhibitors | Laquinimod | [170,175,176] |
| Sodium oxamate | MIF-1 | CH 22391 | [177,178,179] |
| Gallnut extract CN102836354A | OT-1 (TC OT 39) | Triptans (Sumatriptan) | [180,181,182] |
| Galloflavin (NSC 107022) | WAY 267464 | CD56bright NKCs | [183,184,185] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sfera, A.; Klein, C.; Anton, J.J.; Kozlakidis, Z.; Andronescu, C.V. The Role of Lactylation in Mental Illness: Emphasis on Microglia. Neuroglia 2023, 4, 119-140. https://doi.org/10.3390/neuroglia4020009
Sfera A, Klein C, Anton JJ, Kozlakidis Z, Andronescu CV. The Role of Lactylation in Mental Illness: Emphasis on Microglia. Neuroglia. 2023; 4(2):119-140. https://doi.org/10.3390/neuroglia4020009
Chicago/Turabian StyleSfera, Adonis, Carolina Klein, Johnathan J. Anton, Zisis Kozlakidis, and Christina V. Andronescu. 2023. "The Role of Lactylation in Mental Illness: Emphasis on Microglia" Neuroglia 4, no. 2: 119-140. https://doi.org/10.3390/neuroglia4020009
APA StyleSfera, A., Klein, C., Anton, J. J., Kozlakidis, Z., & Andronescu, C. V. (2023). The Role of Lactylation in Mental Illness: Emphasis on Microglia. Neuroglia, 4(2), 119-140. https://doi.org/10.3390/neuroglia4020009