Abstract
The article attests the presence of the genus Rucervus in the paleontological record of Europe and presents the description of new species of large-sized deer Rucervus radulescui sp. nov. from the Early Pleistocene of Valea Grăunceanului (Southern Romania) and Rucervus gigans sp. nov. from the late Early Pleistocene of Apollonia-1 (Greece). The described cervid species represent two different evolutionary radiations of Rucervus that are grouped into the extinct subgenus Arvernoceros that represents the northern evolutionary radiation and the nominotypical subgenus that is regarded as the southern evolutionary radiation and represented today by only one species Rucervus duvaucelii. The evolutionary radiation and dispersals of Rucervus are regarded in the paleobiogeographic context of faunal exchanges between southeastern Europe, Caucasus, and Near East during the Early Pleistocene and the westward dispersal of early hominins in Eurasia.
Keywords:
Villafranchian; deer; Rucervus; Arvernoceros; taxonomy; systematics; morphology; phylogeny; paleobiogeography 1. Introduction
Fossil deer represent a peculiar element of Eurasian Early Pleistocene faunas met by early hominins during their out-of-Africa journey. Deer are opportunistic ruminant herbivores that carry physiologically expensive antlers shed every year and dominated in the broad variety of Eurasian Early Pleistocene ecosystems. Cervids flourished in young renewed ecosystems emerged after each climate shift or geological cataclysm, but they failed to enter the old Tertiary African ecosystems that are composed by specialized and tightly co-adapted faunal and floral species [1]. The meeting of hominins with cervids during their colonization of Eurasia is meaningful, since these two systematic groups are remarkable by their ecological opportunism and high tolerance to Early Pleistocene climate fluctuations [1,2,3,4]. The ecological context of cervid and human coexistence is still poorly studied and only a few studies are focused upon the implication of cervid ecomorphology in early human dispersal into Eurasia [5,6].
In the light of early human colonization of the Mediterranean part of Europe, any additional information yielded by the paleontological record of Southeastern Europe is important and helps to understand the factors that triggered and shaped the early human dispersals in this European region and in the whole west Eurasian area [7,8,9].
Nonetheless, the confused and poorly understood taxonomy and systematics of Early Pleistocene Eurasian deer represent the main difficulty for the implication of the fossil cervid record in the paleoecologic and paleobiogeographic studies. This difficulty objectively is caused by the generally low craniodental and postcranial specialization of deer due to their generally opportunistic and more or less flexible ecological habits. Since the taxonomy of cervids is traditionally based on antler morphology, in some cases it is difficult to associate them with dental and postcranial material and the body size remains the only morphological characteristic that is available for distinguishing the postcranial and dental elements of the cervid skeleton.
The above mentioned general difficulty of cervid taxonomy and systematics becomes more acute in the case of Early Pleistocene deer from Southeastern Europe. The great variety of fossil deer from this region is still poorly understood. Partly, this difficulty is caused by the complicated paleobiogeographic position of Southeastern Europe that brings in contact the immense paleobiogeographic zones of Central, Eastern, and Southern Asia, and the large mountain systems that contributed to evolutionary radiation and paleobiogeographic zonation. On the other hand, according to the historical tradition, the views on systematics and taxonomy of east European fossil deer were adapted to the knowledge on fossil deer from Western Europe, where most of the described fossil species came from and which represents the western paleobiogeographic cul-de-sac of Eurasia.
This work provides a detailed study of interesting and abundant—but still poorly understood—cervid remains from the Early Pleistocene of Valea Grăunceanului (Romania).
Valea Grăunceanului (=Bugiulești) from Southern Romania is an exceptional Early Pleistocene paleontological site that yielded a rich and peculiar fauna of large mammals [10,11]. The biostratigraphic position of the deposits of Tetoiu-1 Formation that crop out in the site of Valea Grăunceanului corresponds to MN 17/MQ1 and it is generally correlated with Olivola F. U. [11,12,13]. The abundant remains of Nyctereutes and the absence of Canis etruscus define the peculiar character of the fauna from Valea Grăunceanului [14] and place it before the so-called “wolf event” [15] or “Pachycrocuta event” [16]. The fauna contains Villafranchian elements, such as Mammuthus meridionalis (Nesti, 1825), Stephanorhinus sp., Metacervocerus rhenanus (Dubois, 1904), Pliotragus ardeus (Depéret, 1884), Gazellospira torticornis (Aymard, 1854), Nyctereutes megamastoides (Pomel, 1843), Ursus etruscus Cuvier, 1823, Pliocrocuta perrieri (Croizet & Jobert, 1828), Homotherium crenatidens (Fabrini, 1890), Megantereon cultridens (Cuvier, 1824), and Lynx issiodorensis (Croizet & Jobert, 1828) [10,11,13,14]. It is considered unique due to the presence of some specific mammalian forms with restricted geographic distribution in Europe: Hystrix sp.; Manis cf. hungarica Kormos, 1934; Mitilanotherium inexpectatum Samson & Radulescu, 1966; and, Paradolichopithecus arvernensis geticus Necrasov et al., 1961 [11,13,14]. Valea Grăunceanului is the type locality for the comparatively large stenonid horse Equus athanasiui (Samson, 1975) [17] and the large terrestrial cercopithecine primate Paradolichopithecus arvernensis geticus. The pebble tools discovered in Tetoiu deposits (Bugiulesti) and the specific composition of the fauna aroused an interest to paleoenvironmental studies of this site in the context of “out of Africa” hominine dispersal [5,14,18]. Despite the great interest of the fauna, some faunal elements are still poorly known. This is the case of the large-sized deer that is one of the most common and well-represented elements of the fauna.
The large-sized deer from Valea Grăunceanului was often arbitrarily reported as Eucladoceros sp. [5,6,11,13]. In a previous publication, I proposed the reconstruction of an antler (Figure 3D, [19]), which allows for the allocation of the deer from Valea Grăunceanului to the genus Arvernoceros. This conclusion was also supported by the brief description of cranial and dental morphology that I gave later [20]. The rich postcranial material of the deer from Valea Grăunceanului has been a subject of ecomorphological study [6]. However, as Curran [6] noticed, the uncertainty allocation of the deer complicates the comparison of the cervid sample from Valea Grăunceanului with other deer forms. In this work, I present a description and analysis of skeleton remains of large-sized cervid from Valea Graunceanului and review their systematic position, phylogenetic relationships, and paleobiogeographic affinities. Furthermore, the obtained results allowed for establishing the phylogenetic and paleozoogeographic relationships of European Early Pleistocene deer with Asian fossil forms and to record the presence of the genus Rucervus in the European paleontological record.
This study also provides a new look on the systematic position of large-sized deer from the Early Pleistocene of southeastern Europe, namely from Liventsovka and Apollonia-1. The large-sized deer from the late Early Pleistocene of Apollonia-1 (MNQ 20: [21,22,23,24]) is particularly interesting from a paleozoogeographic point of view, and it is regarded here as a new species of South Asian origin.
2. Materials and Methods
The studied material of large-sized deer from Valea Grăunceanului is stored in the Institute of Speleology “Emil Racoviță” of the Romanian Academy of Sciences, Bucharest (collection numbers marked with GR) and in the Museum of Oltenia, Craiova, Romania (collection numbers are marked here with BUG). This study also involves the fossil material of large-sized deer from: Apollonia-1 (Greece) stored in the Department of Geology of the University of Thessaloniki, Liventsovka (Russia) stored in the State University of Rostov-upon-Don, and Salcia (Republic of Moldova) stored in the Institute of Zoology of the Academy of Sciences of Moldova. The comparative material includes the sample of Eucladoceros ctenoides from Sénèze that is stored in the Paleontological Museum of Claude Bernard University of Lyon 1; the mixed material of Arvernoceros ardei/Praeelaphus perrieri from Perrier-Etouaires stored in the National Museum of Natural History in Paris; and, the sample of giant deer Megaloceros giganteus from Ireland stored in several European paleontological collections (the Natural History Museum of London, the National Museum of Natural History of Paris, the Paleontological Museum of Munich, the National Museum of Natural History of Bucharest “Grigore Antipa”).
The terminology of dental morphology is applied from Heintz [25]. The measurements are taken according to the recommendations of von den Driesch [26]. The height of first antler ramification is measured from the medial side. The cheek tooth row length or length of tooth series is measured as a maximum length at tooth crowns. The tooth crown length is measured at grinding surface; the tooth crown breadth is taken as a maximum linguolabial measurement. In the case when we are dealing with a poorly known species known mostly from dental remains, an estimated body mass is provided based on dental variables according to the method that was proposed by Janis [27].
Abbreviations used in the article: DLM, mediolateral diameter; DAP, anteroposterior diameter; L, length; D, breadth; H, height; dist, distal; prox, proximal; ap., apical part of antler beam; bs., basal tine of antler; cr., crown tine of antler; M, mean; n, sample size; sin, left; dx, right; ISER, the Institute of Speleology “Emil Racoviță”; MNHB; MO, the Museum of Oltenia, Craiova; the National Museum of Natural History of Bucharest “Grigore Antipa”; SGUT, the School of Geology of the University of Thessaloniki; NHML, the Natural History Museum of London, MNHN, the National Museum of Natural History of Paris; PMM, the Paleontological Museum of Munich; UL, the Paleontological Museum of Claude Bernard University of Lyon 1; and, UR, the University of Rostov-upon-Don.
3. Systematic Paleontology
3.1. Taxonomic Context
- Family: Cervidae Goldfuss, 1820.
- Subfamily: Cervinae Goldfuss, 1820.
- Genus: Rucervus Hodgson, 1838.
Emended diagnosis of Hodgson [28]. Body size varies from medium to very large; antlers of moderate size, with a basal forward tine and distal crown tines inserted on the posterior side of the beam; the crown tines are frequently dichotomously branched; the basal tine is often supplemented with an accessory prong or is branched; no middle tine. The braincase is moderately flexed; parietal bones are not convex. Basioccipitale is bell-shaped, broadened in the pharyngeal tubercles. Small upper canines are present in both sexes.
Type species. Cervus elaphoides Hodgson, 1835 [29], a junior synonym of Cervus duvaucelii Cuvier, 1823 [30].
Comments. Hodgson [28] indicated in the diagnosis of Cervus (Rucervus) that upper canines are only present in males. Pocock [31] confirmed Hodgson’s statement, but he recognized that he was able to check this character only in a single female skull of Rucervus duvaucelii that was studied by Hodgson. According to the craniological material stored in the MNHN, the upper canines are present in both sexes of R. duvaucelii.
The definition of the genus Arvernoceros Heintz, 1970 [25] based on Cervus ardei Croizet et Jobert, 1828 from the Late Pliocene (Early Villafranchian) of Perrier-Etouaires (France) generally corresponds to the diagnosis of Rucervus and therefore is a junior synonym of the latter, or, at the most, could be regarded as a subgenus of Rucervus, as proposed in the present study.
3.2. The New Deer Species from Valea Grăunceanului
3.2.1. Taxonomic Description
Subgenus: Arvernoceros Heintz, 1970.
Diagnosis. The first tine is situated comparatively higher above the burr; the height of the first ramification is no less than twice the basal diameter of antler; the crown part of antler is relatively shorter (about 1/3 of the antler beam length or less) and often is palmated.
Type species. Cervus ardei Croizet & Jobert, 1828.
Composition. Rucervus ardei (Croizet & Jobert, 1928), Rucervus vereschagini (David, 1992), Rucervus giulii (Kahlke, 1997), Rucervus radulescui sp. nov.
Distribution. Early Pliocene–Early Pleistocene, Western Eurasia.
Rucervus radulescui sp. nov.
Synonymy. 1977 Cervus philisi Schaub, 1941—Alekseyeva [32], p. 129, Figure 25. 1990 Eucladoceros sp.—Radulesco & Samson [10], p. 227. 2005 Eucladoceros ctenoides (Nesti, 1841)—Kostopoulos & Athanassiou [33], p. 182, Figure 3. 2008 Arvernoceros sp.—Titov [34], p. 104, Table 46. 2009 Arvernoceros sp.—Croitor [19], p. 382, Figure 3D. 2015 Eucladoceros sp.—Curran [6], p. 299.
Derivato nominis. The species is named in honour of the Romanian palaeontologist Prof. Constantin Radulescu.
Diagnosis. The body size is similar to that of modern European red deer. Antlers are thin and large, with distal crown transformed into a small palmation divided into two lobes terminated with tines. The antler beam cross-section is oval. The basal tine is situated at a long distance from the burr: the basal ramification height exceeds by 2–3 times the antler base diameter. Lower fourth premolar (P4) is simple.
Differential diagnosis. The new species is distinguished from E. ctenoides and E. dicranios by a different antler bauplan: the antler crown in R. radulescui is formed by up to three tines—which may be bifurcated—inserted on the posterior side of the distal portion of the beam. The antler crown in Eucladoceros is formed by three to five crown tines that are inserted on the anterior side of the antler beam. Unlike Eucladoceros, R. radulescui does not possess the accessory prong situated somewhat medially in the area of the first ramification.
R. radulescui is distinguished from Rucervus simplicidens/colberti from the Siwalik fauna by the relatively high position of the first ramification, the relatively shorter crown segment of the antler (the part of antler beam that bears the crown tines), and the palmated distal portion of the antler. The new species is distinguished from R. ardei (Croizet & Jobert, 1828) from the Late Pliocene of Perrier-Etouaires (France) by the absence of a small additional prong on the basal tine, the somewhat divergent pedicles, and the practically complete reduction of the lingual cingulum in the upper molars. Unlike both R. ardei and R. simplicidens/colberti, the new species is characterized by the pronouncedly oval cross-section of the beam.
R. radulescui is distinguished from R. verestchagini (David, 1992) from the Early Pleistocene of Salcia (Moldova) by its significantly smaller body size, the simple cylinder-shaped first tine (not flattened and trifurcated as in R. verestchagini), and the well-developed distal palmation.
Locus typicus. Valea Grăunceanului (=Grăunceanu, =Bugiulești, =Roșiile), western slope of the Dealul Viilor Hill, Tetoiu Area, Vâlcea County, Southern Romania (Figure 1, [13]).
Other sites. Salcia (Moldova), Morskaya Station, Liventsovka (Russia), Gerakarou (Greece).
Stratum typicum. Lower faunal horizon (T-1), Early Pleistocene, MN17-MQ1; correlated with the Tiglian C5 warm stage of the Netherlands and the fauna from Volakas in Greece [10,11,13,14,35].
Holotype. The left shed antler (Figure 1) with collection number “GR-965, c. 17, v. 690” (ISER), which rather belongs to a young adult individual. The first (basal) tine is broken off, the distal palmation is damaged, and its anterior lobe, which corresponds to the apical portion of antler, is destroyed. The posterior lobe of palmation terminates into two tines. The apical parts of those tines are broken off. The total length of the preserved antler part is ca. 540 mm. The antler base is oval-shaped: its DAP is 43.6 mm and DLM is 36.6 mm. The antler circumference above the burr is 125 mm. The height of the first ramification is 106.2 mm. There is a specific sharp rib connecting the beam and the first tine in the area of ramification. The distance between the first ramification and the distal palmation measures to ca. 290 mm. The beam is S-shaped: it is gently curved backward above the first ramification and then it is moderately bowed towards the anterior, gradually expanding into the distal palmation, which is slightly concave from the medial side. The breadth of the posterior lobe of palmation is 89.0 mm. The antler surface is smooth, without pearling; the longitudinal ribs and furrows are moderately expressed. The beam cross-section is oval.
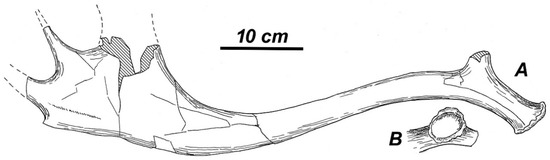
Figure 1.
Rucervus radulescui sp. nov. from Valea Grăunceanului: holotype, the left shed antler “GR-965, c. 17, v. 690” (Institute of Speleology “Emil Racoviță”; ISER): (A) medial view; and, (B) basal view.
3.2.2. Additional Material from the Type Locality
Antlers. The distal portion of right antler “GR-975, c. 17, 720” (ISER) is a somewhat larger specimen that represents a well-preserved palmation (Figure 2). The total length of the antler fragment amounts to 412 mm. The preserved part of the antler beam is strongly compressed laterally: the dimensions at the level of fracture are 44.8 mm (vertical) by 21.0 mm (horizontal). The palmated part of the antler is gently bowed and consists of two lobes separated from each other by a deep rounded incision. The anterior lobe of palmation is interpreted here as a continuation of the main beam. The anterior lobe is supplemented with two prongs, which apparently should be regarded as posterior crown tines. The destroyed distal part of the anterior lobe corresponds to the apical part of the antler beam. The posterior lobe of palmation is regarded here as a lowermost flattened crown tine terminated with a bifurcation.
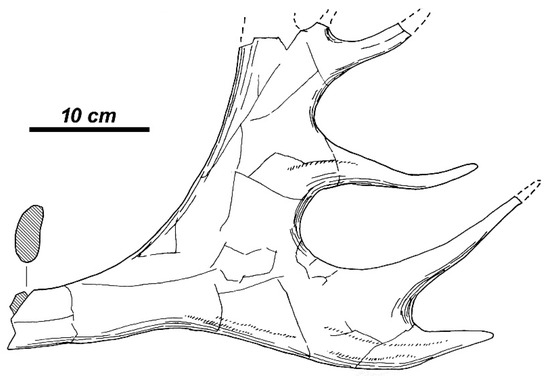
Figure 2.
Rucervus radulescui sp. nov. from Valea Grăunceanului: the medial view of distal portion of right antler “GR-975, c. 17, 720” (ISER) with shape of cross-section at the breakage point (shaded).
The left shed antler BUG-692 (Figure 3) is characterized by a regular circular base (DLM, 42.7 mm; DAP, 43.3 mm). The antler burr is well expressed (DLM, 56.6 mm; DAP, 55.6 mm). The measurements of antler beam above the burr attain 42.0 mm (DLM) and 43.5 mm (DAP). The antler beam that is above the burr slightly deviates sidewards and towards the posterior. In the area of the first ramification, the antler becomes strongly compressed. The first (basal) tine is situated very high from the burr: height of ramification amounts to 119.0 mm if measured from the lateral side and 110.0 mm if measured from the median side. The first tine forms with the antler beam an angle of 90°. The tine is simple, not branched, slightly bent. The cross-section shape of the tine is oval in its proximal part (36.6 × 28.3 mm) and is almost circular near the top. The antler beam above the first ramification is bent towards the posterior and gently curved towards the outer (lateral) side. A well-expressed rib starting from the first ramification extends along the anterior side of the beam and it gradually disappears at ca. 13 cm above the ramification. The beam cross-section is irregular with the sharpened anterior side and the rounded posterior side. The distal portion of the beam is compressed from the sides and has an oval cross-section. The distal part of left shed antler is destroyed; however, one can notice that the antler beam becomes extended into a palmation. The plane of palmation forms an angle of 45° with the plane of the first ramification.
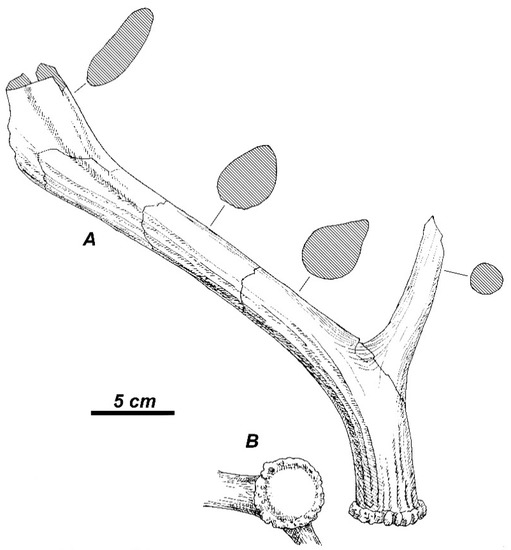
Figure 3.
Rucervus radulescui sp. nov. from Valea Grăunceanului: the left shed antler BUG-692 (Museum of Oltenia; MO); (A) medial view with cross-sections (shaded); and, (B) basal view.
The shed antler BUG-776 preserves only its proximal part with basal ramification and it shows the same morphology and size as in the previous specimen (Table 1).

Table 1.
Antler measurements of Rucervus radulescui sp. nov. from Valea Grăunceanului (GR, BUG), Liventsovka (RGU), and Salcia (52-403), and Rucervus gigans sp. nov. from Apollonia-1 (APL).
Skull. The left (BUG-400) and right (BUG-294) pedicles with basal parts of antlers belonging to the same individual. The pedicles are rather long: their medial length exceeds the lateromedial diameter. The transverse sections of the pedicles are regularly circular.
The facial fragment of cranium BUG-417 is heavily damaged and deformed. The specimen belongs to a senile individual with the deeply worn right tooth series (P2-M2) that does not provide meaningful dental morphological characters. The ethmoidal vacuity is large and attains ca. 45 mm in length and just a little shorter than the length of the upper premolar row (56.5 mm). The preorbital gland pit is moderately deep and as large as the ethmoidal vacuity.
Another facial fragment of cranium “GR-963, c. 8, N II, 140” preserves the posterior parts of nasal bones and the maxillary bones with deeply worn P2-M3 (sin) and P2, P4-M2 (dx). This specimen is characterized by the rather short nasal bones, which do not reach the imaginary line connecting the anterior edges of the orbits. The distance between the posterior edges of nasal bones and the anterior orbit line is quite significant and it amounts to 2 cm. The ethmoidal vacuities are rather large (not preserved completely) and have a long (more than 2 cm) contact with the nasal bones. Preorbital gland pits are moderately deep with blurred edges. The frontal bone is somewhat swallowed in its anterior part and depressed above the orbit. The length of the left upper cheek tooth row (P2-M3) amounts to 116.7 mm, while the length of the left molar series (M1-M3) to 67.5 mm.
The cranial fragment “GR-963, c. 6, N II, 277” belongs to a senile individual as well. It preserves the tooth series P2-M1 (sin) and P3-M3 (dx), the larger part of the left maxillary bone, and the part of the left premaxillary bone. The state of preservation of the specimen is poor, however, it retains the small empty upper canine alveolus filled with sediment. The distance between the canine alveolus and P2 amounts to 74.0 mm.
The badly damaged female skull from Valea Grăunceanului (ISER; no collection number) was briefly described as Arvernoceros sp. [20]. The skull is crushed and its nasal and premaxillary bones are missing. Nonetheless, the preserved frontal bones show that nasal bones were rather short and they did not reach the imaginary line connecting the anterior edges of orbits. The orbitofrontal part of the skull is moderately long: the anterior edge of the orbit is situated above the posterior lobe of M3. The basioccipitale is broad in pharyngeal tubercles and bell-shaped (Figure 4). The length of the right upper tooth row is 130.0 mm (L P2-P4, 58.8 mm, L M1-M3, 77.2 mm); the breadth of the occipital condyles is 72.3 mm; and, the palatal width between M3 is 70.7 mm.
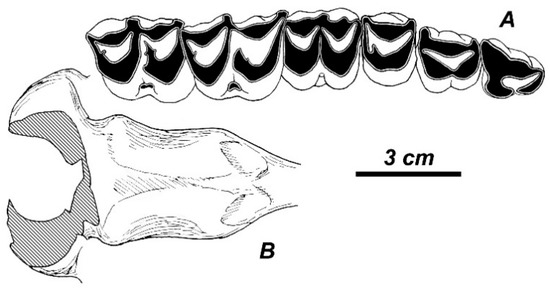
Figure 4.
Rucervus radulescui sp. nov. from Valea Grăunceanului: the occlusal view of the left upper tooth row (A) and the basioccipitale (B) of the female skull from Valea Grăunceanului (no number, ISER).
Mandible. Mandibles of R. radulescui are characterized by the moderately high horizontal ramus; the height of the mandibular horizontal ramus attains 42.6–59.6% (n = 3) of the lower molar series length and overlaps with the proportion values of Eucladoceros from Sénèze (46.2–56.4%, n = 6). The only complete mandible, BUG-30108, is characterized by the moderate length of the diastema that equals the length of the lower molar series (Table 2). Its diastema is relatively shorter than in Eucladoceros from Sénèze that in most cases has a diastema slightly longer than the M1-M3 series, and is much shorter than in modern Rucervus duvaucelii: the diastema in the male individual 1854-16 (MNHN) of this species is as long as the entire lower tooth row P2-M3.
Table 2.
Measurements of the mandibles of Rucervus radulescui sp. nov. from Valea Grăunceanului (GR, BUG) and Liventsovka (RGU), Rucervus gigans sp. nov. from Apollonia-1 (APL), and large-sized Rucervus sp. from Liventsovka (RGU).
Dentition. The description of upper tooth morphology is based on a small series of uncatalogued upper tooth series (five specimens) and isolated teeth (P2, P3, P4, M2) from the collection of Museum of Oltenia and the cranial fragments with dentitions (including the above described damaged female skull) being stored in the Institute of Speleology of Bucharest. The size of dental remains generally corresponds to the size of E. ctenoides from Sénèze and Rucervus colberti from the Siwaliks, and are significantly larger than Rucervus simplicidens from the Siwaliks (Table 3).
Table 3.
Upper cheek teeth measurements of Early Pleistocene Rucervus radulescui sp. nov. and Rucervus gigans sp. nov. compared with large deer from the Siwaliks. The data on Rucervus simplicidens and Rucervus colberti (Siwaliks) are from Colbert [36].
Upper canines. The splanchnocranium fragment GR-963-c.b6-n.II-277 of an old individual is poorly preserved, but is of a special interest, since it shows the presence of an alveolus of a small upper canine. The specimen preserves the heavily worn tooth series P2-M1 (sin) and P3-M3 (dx), most part of the left maxillary bone and a part of the left premaxillary bone. The filled with sediment left canine alveolus is visible on the palatal side of the splanchnocranium. The distance between P2 and the upper canine amounts to 74.0 mm.
Upper premolars. A cleft is present at the lingual side of P2 and its protocone and hypocone are completely separated each from another (Figure 4).
P3 is pronouncedly asymmetric; its hypocone and protocone are separated with a well-marked isthmus. The vertical groove on the lingual wall of the tooth crown is deep and sharp. The hypocone is supplemented with an additional enamel fold from the inner side (Figure 4).
P4 is slightly asymmetric; its hypocone and protocone are separated by a weakly expressed isthmus. A vertical shallow groove on the lingual wall of P4 divides the hypocone and the protocone. The hypocone is supplemented with an inner enamel fold, which may vanish in deeply worn teeth. In less worn teeth, a small enamel islet is visible in the hypocone (Figure 4 and Figure 5).

Figure 5.
Rucervus radulescui sp. nov. from Valea Grăunceanului: the fragment of right maxilla of a young individual with P4-M3 (no number, MO); (A) lingual view; and, (B) occlusal view.
Upper molars. Upper molars are supplemented with a hypoconal spur (which is weak in M3), as well as with small enamel folds (up to three) on the posterior wing of the protocone (Figure 4 and Figure 5). The entostyle is variable from very little to large leaf-shaped with serrated edges. The strong continuous lingual cingulum is never present, however, in some specimens, upper molars have weak enamel folds on the base of lingual side reminding a weak discontinuous cingulum (Figure 5). The molar crowns are comparatively narrow: the crown length exceeds crown breadth (Table 3).
Lower premolars. The lower premolar series is relatively long: the premolar/molar index varies between 64.7 and 72.5% (M = 67.8%, n = 9). The lower tooth series proportion is very close to those of the deer samples from Untermassfeld (M = 65.1%, n = 26) and Perrier-Etouaires (M = 68.1%, n = 9), and relatively longer than in the sample of Eucladoceros from Sénèze (M = 63.3%, n = 17) (Figure 6A). P4 normally is primitive, non-molarized; its parastilid and paraconid are separated; metaconid is rounded and somewhat extended posteriorly due to the extension of its posterior wing. In one case (the specimen BUG-30108), the metaconid of P4 gets in touch with the entoconid, and it almost closes the valley between metaconid and entoconid (Figure 7).
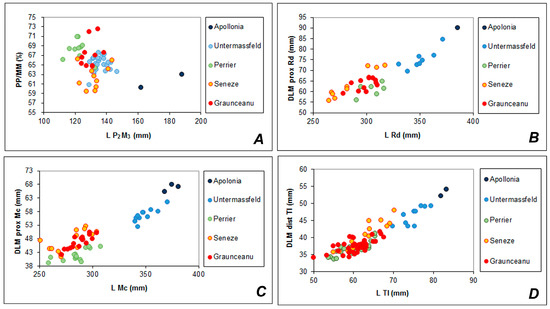
Figure 6.
(A) Lower tooth row size plotted against relative premolar length and limb bone lengths (B, radius (Rd); C, metacarpus (Mc); D, talus (Tl)) plotted against epiphyseal breadth. The data on Rucervus giulii from Unrermassfeld are adapted from Kahlke [37]; the data on Rucervus gigans from Apollonia-1 are adapted from Croitor and Kostopoulos [38]; the data on Eucladoceros ctenoides from Sénèze and on the mixed sample from Perrier-Etouaires are adapted from Croitor and Popescu [39].
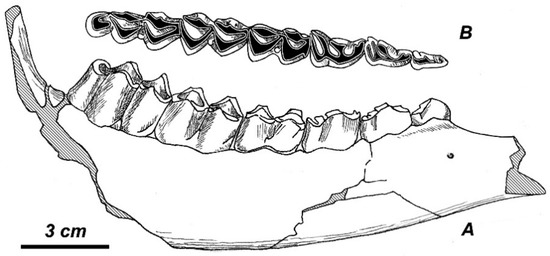
Figure 7.
Rucervus radulescui sp. nov.: right mandibular ramus BUG-30108 (MO): (A) side view; (B) grinding surface of tooth row. The drawing does not show the anterior part of diastemal portion, which was found later.
P2 and P3 represent the typical for the cervines shear functional type. P3 shows a certain degree of specialization: its metaconid is well-expressed, obliquely directed toward the posterior, and terminated with rounded extension, which gets in touch with entoconide, as one can see on the specimen BUG-30108 (Figure 7).
Lower molars. Lower molars are supplemented with well-developed pillar-like ectostylids. M3 bears normally only one anterior ectostylid.
Limb bones. The long limb bones do not show any specific morphological adaptations (Figure 8, Table 4). The metapodial diaphysis are somewhat laterally compressed and are extended in the dorsoplantar direction. This is particularly true for the metatarsal mid-shaft cross-section, which is asymmetric and significantly extended in the dorsoplantar direction (Figure 8III-E). The measurements of radius broadly overlap with those of the samples from Sénèze and Perrier-Etouaires (Figure 6B–D). One can notice that the epiphyses of R. radulescui are relatively broader than those of the perrieri/ardei sample.
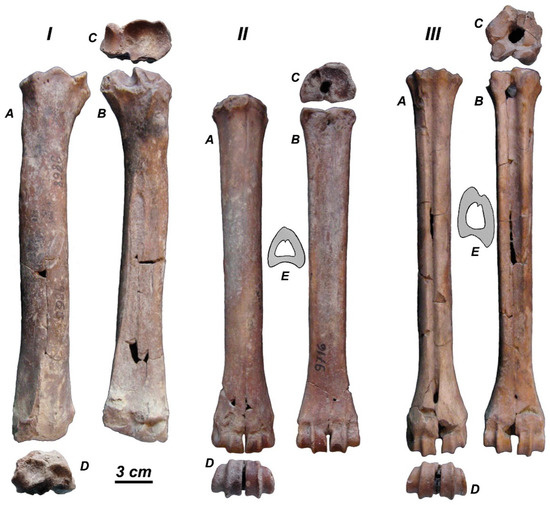
Figure 8.
Rucervus radulescui sp. nov., limb bones from Valea Grăunceanului (MO): (I) the left radius BUG-9868; (II) the left metacarpus BUG-9716; (III) the left metatarsus BUG-9639; A, dorsal view; B, plantar view; C, proximal view; D, distal view; E, midshaft section shape (from the distal fragment of metacarpal without collection number, MO; and, the proximal fragment of metatarsal BUG-9656).
Table 4.
Rucervus radulescui sp. nov. from Valea Grăunceanului: measurements of long bones.
The metacarpal length of R. radulescui is similar to that of the sample from Perrier-Etouaires, however, the metacarpal epiphyses are broader and perfectly correspond to the measurements of Eucladoceros from Sénèze. The metacarpal bones of R. radulescui are significantly smaller and shorter than those from Untermassfeld and the samples under discussion do not overlap (Figure 6C).
The measurements and proportions of the tali of R. radulescui (Table 5) are very close to those of the perrieri/ardei samples from Perrier-Etouaires and broadly overlap with the sample of Eucladoceros from Sénèze, except for the largest specimens (Figure 6D), which, apparently, represent the large form, originally described as Cervus senezensis Depéret, 1910 [40]. The tali of R. radulescui are significantly smaller than those of the large deer from Untermassfeld and the samples under consideration do not overlap. The calcanei of the deer from Valea Grăunceanului show the same intermediary biometric position between the samples from Perrier-Etouaires and Sénèze, which, in their turn, also broadly overlap.
Table 5.
Rucervus radulescui sp. nov. from Valea Grăunceanului: measurements of talus.
3.2.3. Other Findings of Rucervus radulescui
Fossil remains of a similar deer form have been found in the Salcia Quarry (Moldova) and they are stored in the Institute of Zoology of the Academy of Sciences of Moldova. This is a proximal weathered fragment of shed antler (52-403), which perfectly fits the shape, size, and proportions of the sample from Valea Grăunceanului. Most likely, some of the postcranial remains from Salcia also belong to R. radulescui: the proximal fragment of radius 52-1515 (DLM prox, 61.8 mm; DAP prox, 33.6 mm), which falls among the large specimens from Valea Grăunceanului, and the proximal fragment of metatarsus 52-335 (DLM prox, 35.4 mm; DAP prox, 41.0 mm), which corresponds to the smallest specimens of R. radulescui from the type locality.
Quite abundant remains of R. radulescui have been yielded in the Khaprovian deposits from the Azov Sea Area. A quite complete antler GIN 301-12 (now destroyed) was discovered near the Morskaya Station (Russia). This specimen was reported by Alekseyeva [32] as Cervus philisi. The figure provided by Alekseyeva [32] apparently represents the outlines of the medial and lateral sides of the antler redrawn from a photograph. The antler is characterized by a high position of the first tine and a small bilobed distal palmation, which, obviously, is partially destroyed. Titov [34] reported the remains of this deer as a juvenile or small-sized form of Arvernoceros sp. This arbitrary definition is caused by the fact that the Khaprovian deposits have yielded remains of another similar, but much larger, cervid form. Although the fragmentary character of the fossil remains did not allow for Titov [34] to make an adequate systematic interpretation, he cautiously provided the antler measurements of the “large” and the “small” or “juvenile” Arvernoceros separately.
One can assume that the “juvenile” Arvernoceros from the Khaprovian fauna is a cervid form very similar to, or even identical with, the deer from Valea Grăunceanului. The basal fragment of left antler RGU-836 (Figure 9) is very close in size and shape to the antlers from Valea Grăunceanului. The antler beam is set obliquely with respect to the burr and somewhat directed sideward. The antler becomes quite flat in the area of the first ramification and continues into a beam with the circular cross-section and the compressed from the sides first tine. The first tine and the main beam are connected by the specific sharp rib (the morphological character also noticed by Titov [34]). The angle of the first ramification is ca. 95°. The fragment of left frontal bone RGU-759 with a small basal portion of antler is characterized by a relatively long pedicle—the length measured from the posteromedial side exceeds the pedicle diameter—with the circular cross-section (Table 1). Here, also should be mentioned the weathered frontal fragment of skull RGU-774 that belongs to an older individual with more robust pedicles (Table 6). The frontal bones are somewhat concaved before the pedicles, however, the interfrontal suture is not elevated. The pedicles are not bordered by a bony ridge at their bases from the medial side, as in Praemegaceros, and they are quite divergent: the angle between the pedicle axes amounts to 110°. The pronounced divergence of the pedicles distinguishes this cervid form from R. (Arvernoceros) ardei, which is conversely characterized by a minor divergence of pedicles [25].
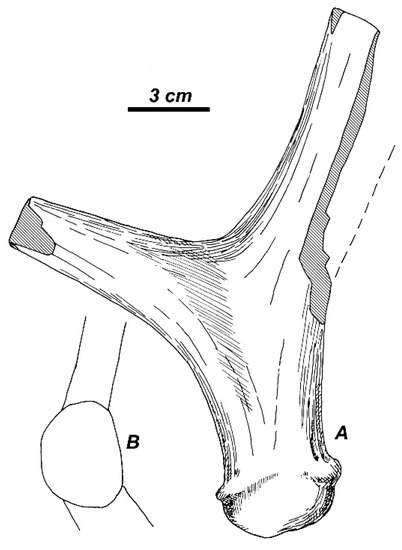
Figure 9.
Rucervus radulescui sp. nov. from Liventsovka: basal fragment of the left shed antler RGU-836 (University of Rostov-upon-Don, UR); (A) lateral view; and, (B) basal view.
Table 6.
Cranial measurements and proportions of Rucervus gigans sp. nov. from Apollonia-1 (APL) and Kalamoto (KAL, adapted from Tsoukala & Chatzopoulou [41]) as compared to large-sized Rucervus sp. from Liventsovka (RGU).
The fragment of left ramus of mandible RGU-578 from Liventsovka preserved a part of tooth series P4-M2 (Figure 10). The fourth premolar is primitive, unmolarized, however, it shows some specific features that are shared with the deer from Valea Grăunceanului: its metaconid is long and is obliquely directed to the posterior due to the strong extension of its posterior wing. Molars are supplemented with small ectostylids. The fragment of right mandibular ramus shows the same dental morphology, which is even more accentuated due to the lesser degree of dental wear. Both metaconid and entoconid are elongated and obliquely extended towards the posterior. The fragment of left hemimandible RGU-394 with P3-M1 presenting a more advanced degree of wear is characterized by the fusion of entoconid and entostylid in P3 and P4 that closes the posterior valley of the premolars, and the fusion of paraconid and metaconid that closes the anterior premolar valley, so that P4 becomes molarized.
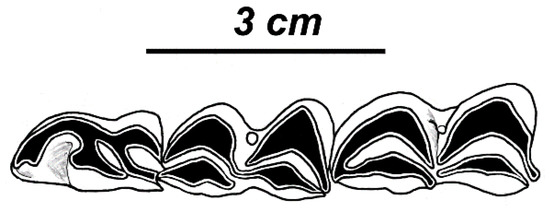
Figure 10.
Rucervus radulescui sp. nov. from Liventsovka: dental morphology of P4-M2 of the left ramus of mandible RGU-578 (UR).
The cervid remains from the Early Pleistocene of Volakas, Sesklo, Dafnero, and Gerakarou (Greece), as reported by Kostopoulos and Athanassiou [35] as Eucladoceros ctenoides should also be mentioned in this discussion. Antler remains are not described from the above-mentioned faunas, while postcranial bones and teeth belong to a cervid form, which is slightly smaller than the deer from Valea Grăunceanului and generally corresponds to the size of perrieri/ardei sample from Perrier-Etouaires. The incomplete lower tooth row from Gerakarou (Figure 3, [33]) is characterized by the presence of a simple unmolarized P4. The metaconid of P3 is very large and sloped towards the posterior and gets in touch with the entoconid, as in the dental material from Valea Grăunceanului and Liventsovka. Possibly, this peculiar specialization of premolar morphology permitted to indicate the more advanced evolutionary character of the Greek material if compared to Eucladoceros from Sénèze [33]. The missing data on antler morphology and comparatively small size of the Greek material warn against the definite systematical conclusions. However, the lower tooth row from Gerakarou shows specific morphological characters of R. radulescui and therefore should be ascribed to this species. Most probably, the postcranial and dental remains from Volakas, Sesklo, and Dafnero reported as Eucladoceros [33] also belong to R. radulescui. The postcranial cervid remains from Varshets and Slivnitsa (Bulgaria), as reported by Spassov [42] as a rather small Eucladoceros sp. require a further comparative study in order to reveal their taxonomic position.
To conclude this chapter, it is necessary to add that antler remains (the most diagnostic cervid skeletal remains) of typical E. ctenoides with comb-like bauplan have not been recorded in Eastern Europe so far.
3.3. The New Large-Sized Deer Species from Apollonia-1
3.3.1. Taxonomic Description
Subgenus: Rucervus Hodgson, 1838.
Diagnosis. the first tine is situated comparatively low above the burr; the height of the first ramification is less than twice the basal diameter of antler; the crown part of antler is relatively longer (no less than 40% of the antler beam length) and often is not palmated.
Type species. the subgenus is nominotypical.
Composition. Rucervus duvaucelii (Cuvier, 1823), Rucervus simplicidens (Lydekker, 1876), Rucervus colberti (Azzaroli, 1954), Rucervus schomburgki (Blyth, 1863), Rucervus gigans sp. nov.
Distribution. Pliocene–Holocene, Southern Eurasia.
Remark. Possibly Arvernoceros insolitus Vekua et al., 2010 also should be included in this subgenus or, at list, it is closely related to the southern evolutionary radiation of Rucervus, however, we need more data to confirm the taxonomic and systematic position of this species.
Rucervus gigans sp. nov.
Synonymy. 1997 Megaloceros sp.—Kostopoulos [21], p. 846, Figure 1A–F. 2004 Arvernoceros cf. verestchagini David, 1992—Croitor & Kostopoulos [38], p. 140, Figures 1, 2, 4A and 5.
Derivato nominis. In Latin: giant, or the Giants, mythic race defeated by the Olympians.
Diagnosis. The body size is very large, being similar to that of the giant deer Megaloceros giganteus. The first ramification is situated quite close to the burr (the height of ramification is less than 2 × DAP of antler’s base), flattened, bifurcated, with the vertical orientation of the flattened portion and the fork. The angle of the first ramification is obtuse. The cross-section of the antler beam is circular. Pedicles are moderately long and somewhat deflected backwards, with a circular cross-section. Lower fourth premolar (P4) is simple. The relative length of lower premolars is relatively long: premolar/molar ratio normally exceeds 60%. Upper molars are supplemented with well-developed triangular entostyles, which are supplemented with a weak lingual cingulum at least in the M3. Protoconal fold in the upper molars is not present. Mandibular diastema is somewhat shorter than lower molar series. Metapodials are relatively long; the length of the metacarpus is comparable with that of the metatarsus
Differential diagnosis. The new species is significantly larger than R. simplicidens and R. colberti from the Siwalik fauna. R. gigans is distinguished from R. radulescui by the significantly larger body size and the low position of the bifurcated first tine. The new species is distinguished from Praemegaceros pliotarandoides that is found in the same fauna, by the primitive unmolarized P4, the flattened and bifurcated first tine situated on the anterior side of the beam close to the burr (not cylinder-shaped, inserted on the anteromedial side of the beam, as in P. pliotarandoides), the absence of vestigial subbasal tine, and the horizontal diastemal part of mandible (the symphisal part is not elevated as in Praemegaceros). R. gigans is distinguished from R. (Arvernoceros) verestchagini by the lower position of its first tine, which is bifurcated, not extended into a small palmation and trifurcated as in the latter species.
Locus typicus. Apollonia-1, 60 km NE to Thessaloniki, Greece (Figure 1, [22]).
Stratum typicum: the fluviolacustrine sediments of Platanochori Formation (Premygdonian group), the final stage of the Lower Pleistocene, MNQ 20, 1.3–1.0 Ma [21,22,23,24].
Syntypes: The left hemimandible APL-384 with P3-M3 and destroyed ascending part (Figure 1D, [21]; Figure 5, [38]); the left maxilla APL-274 with full cheek tooth series P2-M3 (Figure 4A, [38]); the basal fragment of antler APL-357 with almost complete basal tine and partially destroyed burr (Figure 1B, [21]; Figure 1, [38]); the right metacarpus APL-385 (Figure 1E, [21]); the left metacarpus APL-334 (Figure 1F, [21]); and the frontal fragment with preserved basal portions of antlers APL-212 (Figure 1A, [21]; Figure 2, [38]). The syntypes are stored in the School of Geology of the University of Thessaloniki (Greece).
3.3.2. Morphological Description
The detailed description is published by Croitor and Kostopoulos [38]. Here, I focus the attention upon the diagnostic characters that support the proposed new taxonomic status of the large-sized deer from Apollonia and reveal its systematic position and phylogenetic relationships.
The size of the left mandibular ramus APL-384 (Figure 11) is extremely large: the length of its cheek tooth row exceeds the analogous measurements taken from the sample of Megaloceros giganteus stored in the NHML (LP2-M3, 161.1–173.1 mm, n = 12). Its mandibular body is relatively low for its size and is not pachyostotic (Table 2). The diastemal part of the mandible is relatively short: the length of the diastema is slightly shorter than the length of the lower molar series. P4 is simple: its metaconid is moderately extended anteroposteriorly. Paraconid and parastylid of P3 and P4 are fused and form the specific isolated enamel islet.
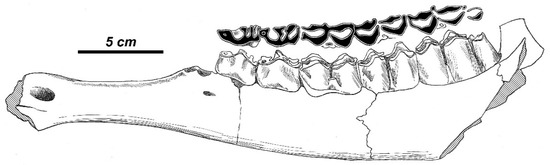
Figure 11.
Rucervus gigans sp. nov. from Apollonia-1: the lateral view of the left mandibular ramus APL-384 (syntype; School of Geology of the University of Thessaloniki, SGUT) and the shape of the occlusal surface of toothrow.
The mandibular fragment APL-33 belongs to an older individual with a deeply worn dentition (M1 is almost completely worn out). The length of its tooth row falls within the variation range of the Irish sample of M. giganteus stored in the NHML. The entoconid and entostylid of P4 are fused and they close the little posterior valley, as in the deeply worn fourth premolars of R. radulescui from Liventsovka, however, the anterior valley remains open.
The maxilla APL-274 (Figure 12, Table 3) is with the full row of cheek teeth on the advanced stage of wear, however, the meaningful morphological characters are clearly visible. The total tooth row length exceeds the size range of variation of M. giganteus from Ireland (LP2-M3, 141.6–153.3 mm, n = 9; stored in the NHML, MNHN, PMM, NHMB). The lingual wall of P2 is cleft, so that its protocone and hypocone are separated. The inner wall of the protocone of P2 is folded. The lingual wall of P3 has a shallow vertical groove; the protocone and the hypocone of P3 are connected through an isthmus. The inner was of the hypocone of P3 is undulated. The hypocone includes a small enamel islet. The inner wall of hypocone of P4 is supplemented with a large enamel fold. Upper molars are characterized by the well-developed hypoconal spur and the flattened entostyles. In M3, the entostyle is larger than in other molars and it is connected with a very small, but clear lingual cingulum. The molar crowns are comparatively broad: the measurements of their breadth are close to the values of the crown length (Table 2).

Figure 12.
Rucervus gigans sp. nov. from Apollonia-1: the left maxilla APL-274 (syntype; SGUT) with tooth series P2-M3: grinding surface view.
The basal fragment of shed antler APL-357 (Figure 13) is characterized by the very low position of the basal tine and the obtuse angle of the first ramification. The basal tine becomes flattened in its distal portion and then bifurcated. Unlike M. giganteus, which is also characterized by flattened and bifurcated basal tine; the flattened portion of the basal tine in R. gigans is set in the vertical plane.
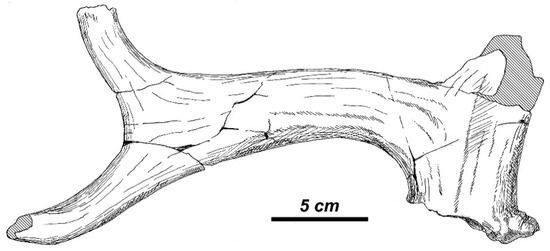
Figure 13.
Rucervus gigans sp. nov. from Apollonia-1: the basal fragment of antler APL-357 (syntype, SGUT).
The frontlet APL-212 (Figure 14A) belongs to a rather young individual; its pedicles are relatively long with the circular transversal section. The pedicles are pointed sidewards and backwards and they are situated in the plane of the frontal bones. The angle of the pedicle divergence is right. The sagittal profile of frontal bones is prominent between the pedicles and slightly concave before the pedicles. The frontal bones become concaved in the sides of the interfrontal suture in the supraorbital area. The interfrontal suture is slightly elevated in front of the pedicles. The frontal part of the skull is relatively broad with respect to the braincase breadth measured behind the pedicles. This characteristic distinguishes the specimen from Apollonia-1 from the “large Arvernoceros” from Liventsovka (Figure 14B) with relatively narrower frontals (Table 6). The antlers are characterized by a position of the basal tine higher than in the previous specimen, suggesting a certain degree of the individual or ontogenetic variation of this character. The antler becomes strongly compressed lateromedially in the area of the first ramification, while the main beam cross-section is circular.
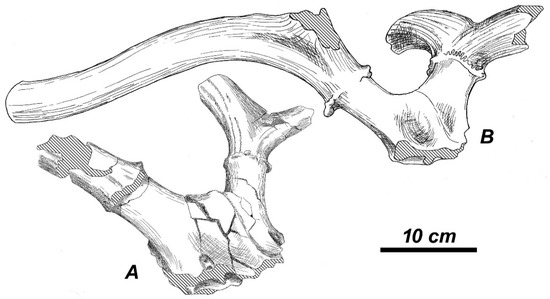
Figure 14.
Antlered frontlets of (A) Rucervus gigans sp. nov. from Apollonia-1 (APL-212, syntype, SGUT) and (B) Rucervus sp. from Liventsovka (RGU-239 liv, UR); oblique views.
The postcranial bones are exceptionally large (Figure 6) and they cannot be confused with any cervid sample considered in the present study. It is necessary to stress the most curious morphological peculiarity of R. gigans that apparently reflects the specific ecological niche of a high level browser occupied by this species [38]: its metacarpals, metatarsals, and radius are too long for a deer of such large size and, respectively, are 10.7%, 11.8%, and 4.2% longer than the corresponding mean lengths of the giant deer from Ireland (the data on Irish giant deer are taken from Croitor et al. [43]). The proportions of limbs in R. gigans are also peculiar: its metacarpals are comparable in length with the radius and metatarsus [38].
4. Discussion
4.1. Plio-Pleistocene Rucervus from Europe
Cervus ardei Croized & Jobert, 1828 from the Late Pliocene of Perrier-Etouaires (France) is the earliest cervid from the European paleontological record that displays a bauplan close to the Rucervus antler. Heintz [25] included Cervus ardei in the new monotypic genus Arvernoceros, which is regarded here as the subgenus of Rucervus. The total length of the type antler of R. (Arvernoceros) ardei (no number, MNHN) attains 610 mm [25]. The first tine in the type specimen is situated rather high above the burr: the first ramification height measures more than twice as much as the DAP of the antler base. The first tine is supplemented with an accessory prong, as in modern R. (Rucervus) duvaucelii (not visible on the Figure 15). This accessory prong may be variable in shape and position and in some cases is situated in the area of ramification. The middle tine is never present. The antler beam is cylinder-shaped and curved backward from the first ramification and then somewhat bowed anteriorly in its distal part. The antler is terminated by a small palmation with three tines (the maximum number of tines recorded for this species), which are pointed upward and forward (Figure 15D) and do not show the typical for Rucervus crown construction with distal tines being inserted on the posterior side of the beam. Nonetheless, the direction of two posterior tine may not be exact, since the antler is reconstructed at the base of each tine. Even if the shape of palmation is reconstructed correctly, it may represent an individual variation, which deviated from the “initial” described for Rucervus antler construction pattern. The distal fragment of antler PET1024 (MNHN) is of interest for this discussion, since it represents a part of palmation resembling the posterior palmation lobe with two terminal prongs in R. radulescui (Figure 16). Therefore, the distal tines of the type specimen of Cervus ardei could be interpreted as the apical tine and the two posterior tines that form a posterior lobe of palmation. Heintz [25] ascribed to R. (Arvernoceros) ardei upper molars with a strongly developed cingulum. Mandibles and postcranial bones of large cervids from Perrier-Etouaires represent an unresolved problem, since the type locality has yielded the remains of another deer species of similar size Praeelaphus perrieri [25,44]. The pedicles of R. (Arvernoceros) ardei are quite long, cylinder-shaped, and little divergent, and therefore represent the most primitive morphological condition within Rucervus. Other cranial characters of R. ardei remain unknown. The specific morphological traits of R. ardei that could be regarded as diagnostic for the subgenus Arvernoceros are the very high position of the first ramification and the relatively short crown part of the antler that attains less than one-third of the total antler length.
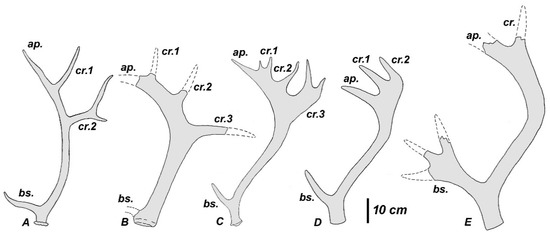
Figure 15.
Antler shape of (A) Rucervus (Rucervus) duvaucelii; (B) Rucervus (Rucervus) simplicidens/colberti; (C) Rucervus (Arvernoceros) radulescui; (D) Rucervus (Arvernoceros) ardei; and, (E) Rucervus (Arvernoceros) verestchagini; bs., basal tine; ap., apical part of beam; cr., crown tine.
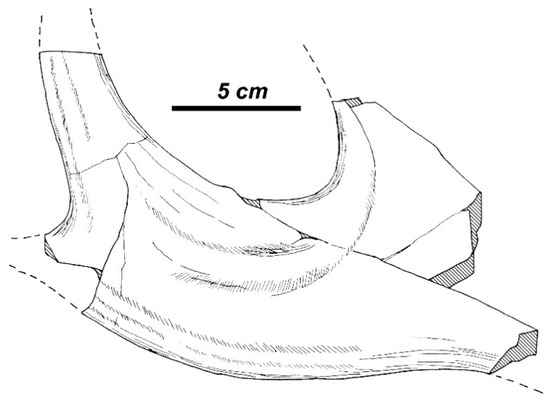
Figure 16.
Rucervus (Arvernoceros) ardei from Perrier-Etouaires: the bifurcated posterior lobe of distal palmation PET-1024 (National Museum of Natural History, Paris, MNHN).
Heintz [25] assumed the phylogenetic relationship between Rucervus ardei and Megaloceros giganteus, however, he acknowledged that this hypothesized phylogenetic link is not demonstrated. Vislobokova [45] supported the direct phylogenetic relationship between Rucervus (Arvernoceros) ardei and M. giganteus. This link, according to the cited authors, is based on such shared characters as the development of small distal palmation with distal tines directed towards the anterior, and the presence of cingulum in the upper molars. Actually, cingulum is a quite variable morphological feature in giant deer and it is well-developed mostly in the better known Irish sample, while the continental forms of M. giganteus are characterized by a very weak lingual cingulum in the upper molars [44], therefore, this character, as well as the apomorphic development of distal palmation serve as weak arguments for the evolutionary link between Arvernoceros and Megaloceros.
R. verestchagini (David, 1992) is a much larger species coming from the Early Pleistocene of Salcia (Moldova). The general antler bauplan is very similar to that of R. ardei, however, besides the extremely large size—the diameter of its antler base is ca. 10 cm—R. verestchagini is characterized by the poor development of crown (Figure 15E), which is represented in the type specimen by a simple fork formed by the apical part of the beam and the thinner posterior tine, and the particularly large and flattened first tine terminated by three prongs [46].
Another comparatively large cervid has been reported from Liventsovka (Russia) as Arvernoceros sp. [34,47]. As R. ardei and R. verestchagini, the deer from Liventsovka is characterized by the high position of the first tine (Figure 14B). The pedicles are cylindrical, robust, and divergent. The shape of basal tine and the distal part of the antler are unknown. Titov [34] reported from Liventsovka some fragments of palmated antlers as Cervidae gen. indet., which, possibly, might belong to this cervid form. The size of the deer from Liventsovka is quite large but smaller than that of R. verestchagini. Since the main diagnostic characters of this deer (the shape of the first tine and the shape of distal portion) remain unknown, this cervid is regarded here as Rucervus (Arvernoceros) sp. The high position of the first tine may suggest its phylogenetic relationship with European forms. The geological age of the large Rucervus from Liventsovka remains unclear, since the Livenzovka collection seems to include material from two different stratigraphic levels at least: one of early Villafranchian age (MN16b) and another one quite younger (MN17) [48,49].
The large-sized deer from the final Villafranchian from Saint-Prest (France), Rosières (France), and Untermassfeld (Germany) is another cervid form that should be included in the group of European Rucervus. This is a conspicuously long-limbed and large-sized deer (larger than European Eucladoceros) with a primitive lower fourth premolar and a practically absent cingulum in the upper molars. This deer was described under different names: Cervus (Megaceros) dupuisi Shehlin, 1912 based on the sample from Rosières [50]; Eucladoceros giulii Kahlke, 1997 based on the sample from Untermassfeld [37]; and, reported by Guerin et al. [51] as Praemegaceros verticornis (Dawkins) from Saint-Prest [19]. According to Azzaroli [52], some mandibles from the Forest Bed Formation (England) of the intermediary between P. verticornis and Praemegaceros dawkinsi size are very similar to Megaceros dupuisi from Rosieres. The mandibles in question from the Forest Bed Formation were ascribed by Azzaroli [52] to Praedama savini (=Megaceros savini according to Azzaroli, 1953). The complete mature antlers of this cervid form are unknown. Kahlke [37] proposed a hypothetical reconstruction of antlers that shows the comb-like bauplan of European Eucladoceros ctenoides. Actually, a “typical” E. ctenoides is present in the roughly coeval with Untermassfeld French site of Ceyssaguet where this species is represented by fine complete antlers and it is characterized by “normal” for European Eucladoceros body size [53]. Unlike E. ctenoides from Ceyssaguet, the cervid from Untermassfeld is characterized by the relatively longer lower premolar series and the unsplit lingual walls in the upper molars [38]. The hypothesized antler reconstruction proposed by Kahlke [37] has been questioned by Croitor and Kostopoulos [38]. The only complete antler from Untermassfeld is juvenile (p. 229, Figure 26, [37]), but it shows some features characteristic of Rucervus (Arvernoceros): the first tine is situated high above the burr, flattened from the sides, and terminated with a bifurcation. The angle of the first ramification is 55°. The main beam is somewhat flattened in the area of distal bifurcation. The general morphological specialization of the antler under discussion recalls R. verestchagini [38].
4.2. Plio-Pleistocene Rucervus from the Siwaliks and Caucasus
The cervid fossil record from the Siwaliks and Caucasus is significant for the understanding of the origin and systematic position of Early Pleistocene deer from southeastern Europe, however, this important source on the geological history of cervids is undeservedly ignored.
Up to seven deer species and forms are reported from the Siwalik Hills. The Siwalik cervid species were established mostly on the morphology of upper molars, the fact that limits the understanding of their taxonomic status and systematic position. As a matter of fact, only three types of antlers have so far been reported from the Siwaliks, which attest the presence of the genera Metacervocerus (the sister genus of modern Axis), Panolia (reported by Azzaroli [54] as Rucervus sp. I and Euctenoceros sp.), and Rucervus [36,54,55]. Another smaller Axis-like deer Cervus rewati is described by Arif et al. [56] from the Upper Siwaliks of Pakistan.
Cervus latidens Lydekker, 1876 was described on the basis of few very large isolated molars [57]. The dental remains of this poorly known ruminant form are as large as those of the deer from Apollonia measurements of M3 (L = 31.2 mm [1.23 in.], D = 25. 4 mm [1.0 in.], the measurements are adapted from Lydekker [57]), but the tooth crown of the Siwalik form is relatively narrower in the linguolabial dimension. This is a dental characteristic correlated with the comparatively more advanced degree of hypsodonty in the Siwalik teeth. Besides that, the figures of upper molars (Pl. VIII, Figures 4, 6, 7, 10, [58]) show the typical bovid characteristics: the anterior and posterior walls of molars are parallel, the entostyle is high and column-shaped, and the hypoconal enamel spur is missing (the hypoconal spur is present in R. gigans). According to Lydekker [58], the initial inclusion of this species to Cervidae was arbitrary and later he included “Cervus latidens” in the bovid genus Oreas [59]. This taxonomic revision was never contested afterwards.
Cervus triplidens Lydekker, 1876 is another questionable ruminant species from the Siwaliks included in the family Cervidae. The species is based on the maxilla with high-crowned M2-M3 ([58]: Pl. VIII, Figures 1 and 2), is characterized by high and strong column-shaped entostyles, and therefore looks rather like a bovid. Koken [60] expressed his doubts about the attribution of the specimen in question to a cervid. It is important to mention that Lydekker [59] himself indicated in the footnote on page 116 the “superficial” similarity of dental morphology of C. triplidens with Boselaphus sp. According to Colbert [36], this species is based on a very meagre material and therefore is of “little value”. Colbert [36] provisionally ascribed to C. triplidens the left maxilla No. 19792 (AMNH) with P3-M3. This specimen shows a typical cervid morphology. The molars are low crowned and are supplemented with a quite strong and flat entostyle (not column-shaped as in the type specimen), but are devoid of a hypoconal spur.
Cervus simplicidens is the only Lydekker’s species from the Siwaliks (Pl. VIII, Figure 3, [57]) that is certainly based on true cervid remains: the upper molars of this species are supplemented by a small lingual cingulum and a small entostyle [54]. The stratigraphic provenance of this cervid form is uncertain (Middle or Upper Siwaliks [36,54]). According to Lydekker [58], the general shape of the molars is very similar to those of the living Rucervus duvaucelii. However, another maxilla with P3-M3 (Pl. VIII, Figure 6, [59]) described as C. simplicidens, again, shows the typical bovid morphological characters: massive column-like entostyles, high crowns with parallel anterior and posterior walls, and no cingulum. Colbert [36] described as C. simplicidens a juvenile cervid right maxilla with deciduous teeth and two molars supplemented with a small entostyle and a hypoconal enamel spur. Azzaroli [54] ascribed to C. simplicidens a fine male skull B.M. 39570 (NHML), noticing the great similarity of its molar morphology with the type specimen. The skull also displays a great morphological affinity—except for the relatively shorter muzzle—with modern R. duvaucelii, the fact that permitted to Azzaroli [54] to include the species under discussion in the genus Rucervus. The available for observation morphological characters of the cranium BM39570 (the small upper canines; the large ethmoidal openings with a long border formed by nasal bones at more than ½ of ethmoidal opening length; the position of the anterior edges of the orbits above the posterior edge of M3) confirm the opinion of Azzaroli [54].
“Cervus” sivalensis Lydekker, 1876 is based on the lower molars that are figured by Lydekker [57] in the Tab. XIII, Figure 5. Originally, Lydekker (ibidem) supposed that this specimen could belong to Cervus triplidens, but in the introduction chapter, which was prepared somewhat later, Lydekker proposes for this specimen a new species name Cervus sivalensis. Later on, Lydekker [59] ascribed to this species the quite worn M2 and M3, which, however, provide the typical cervid morphology and they are supplemented with a small lingual cingulum. According to Lydekker (ibidem), the size and morphology of this specimen agree with those of modern R. duvaucelii. The morphology of the upper third molar figured by Lydekker (p. 122, Figure 8, [59]) strongly recalls the type specimen of Cervus simplicidens and most probably should be ascribed to this species. Azzaroli [54] regarded this specimen (BM48440, NHML) as the holotype of “Cervus” sivalensis, but this is not the case, since the species originally was based on the lower molars figured in Lydekker [57]. Actually, the type specimen of “Cervus” sivalensis provides poor diagnostic characters and its taxonomic status for the moment is unclear since the Siwalik fauna contains another cervid of the similar size, Metacervocerus punjabiensis (Brown, 1926) [61,62]. Colbert [36] ascribed to “Cervus” sivalensis a well-preserved skull No. 19829 (AMNH) and the almost complete antler No. 19807 (AMNH), which strongly resemble R. duvaucelii (Figure 15A,B). The antler is heavily built, with round cross-section; the basal tine is situated very close to the burr; the antler crown is formed by at list three crown tines that were inserted on the posterior side of the distal part of the beam [36], as in modern R. duvaucelii. The upper molars of the skull are deeply worn, however, as one can see on the photographic image (Figure 145, [36]), there is a weak but clear lingual cingulum [54].
Azzaroli [54] proposed a new species name Cervus colberti for the skull No. 19829 (AMNH), since, according to the cited author, it is distinguished from R. simplicidens by a somewhat larger size. Azzaroli (ibidem) distinguishes his new species from “Cervus” sivalensis by smaller cingulum and more rounded protocone and hypocone. Actually, the degree of cingulum expression may be an individually variable character, while the lingual outlines of protocone and hypocone depend on the stage of molar wear, which seems to be very advanced in the type specimen of Rucervus colberti (Figure 145, [36]). The proposed in my previous work [55] body mass estimation based on the figure of skull 19829 (AMNH), as published by Colbert [36], is exaggerated. In fact, the body size of R. colberti is not very different from that of R. simplicidens and it generally corresponds to the size of Eucladoceros. It is difficult to say whether R. colberti (Azzaroli, 1954) is indeed a larger advanced form closely related to R. simplicidens (Lydekker, 1876) since the measurements of the discussed cervid remains from the Siwaliks stored in NHML [54] are still missing. There is a probability that we are dealing with only one species R. simplicidens and R. colberti is its junior synonym.
The paleontological record of the Caucasian region brings several poorly known and poorly understood ruminant forms reported as cervids. Some of them seem to be related to the Siwalik species, however, a direct comparative study was never done.
Pseudalces mirandus Flerov, 1962 is an imperfectly known very large ruminant from southeastern Europe based on very poor cranial material from the mixed Pliocene fauna of Kosiakino Quarry (South Russia, Stavropol). The new genus and the new species were proposed by Flerov [63] on the basis of the orbitofrontal part of the skull with upper left tooth row P4-M3. Flerov and Shevyreva [64] suggested the morphological affinity of Pseudalces with Eucladoceros but excluded the possibility of the direct phylogenetic relationship between those two genera. Vislobokova [65] regards P. mirandus as a side phylogenetic branch of true elks (Alceini). The similar large ruminant form was described by Vekua [66] from the late Early Pliocene of Kvabebi (Georgia) as Pseudalces sp. The dental remains from Kosiakino Quarry and Kvabebi belong to a rather large ruminant (the estimated body mass exceeds 400 kg) and they show some morphological peculiarities that cast doubts on their attribution to a cervid: the protocone in the upper molars has no additional enamel fold as it could be expected in primitive elks, while the premolars remain relatively large, distinguishing the specimens under question from Cervinae. The enamel “pockets” from the labial side of the upper molars from Kvabebi remind the morphological condition seen in Giraffa punjabiensis (Figure 195, [36]). The systematical position of P. mirandus from Kosiakino Quarry and Pseudalces sp. from Kvabebi requires a revision and possibly those remains actually belong to Giraffidae.
The somewhat smaller ruminant from Kvabebi reported by Vekua [66] as Eucladoceros sp. is a cervid similar in body size to Eucladoceros from Sénèze. This deer form is characterized by the certain distance between the antler burr and the first tine, and the laterally compressed antler portion in the area of the first ramification (p. 202, Figure 50, [66]). The basal part of the first tine is flattened and forms a right angle with the main beam, which, in its turn, is sharply bent backwards from the first ramification, as in Rucervus from the Siwaliks figured by Azzaroli [54]. Pedicles are quite short (the pedicle diameter exceeds the pedicle length), cylindrical, and divergent, with an angle of divergence of 95°. The metatarsal bone from Kvabebi (p. 202, Figure 51, [66]) fits the size and proportions of R. radulescui. The mandible K-1022 described as Pseudalces sp. (p. 207, Figure 53, [66]) certainly belongs to this cervid form. This specimen is characterized by the low mandibular body, the primitive shape of P4, and the relatively long premolar series. The mandible from Kvabebi is distinguished from the sample of Eucladoceros from Sénèze by its relatively longer premolar series, which is regarded here as a primitive morphological condition. The size and dental row proportion of the specimen K-1022 is very similar to those of R. radulescui from Valea Grăunceanului.
Arvernoceros insolitus Vekua et al., 2010 from Early Pleistocene of Dmanisi (Georgia) is a large-sized deer that is characterized by the very low position of the first tine and the distal palmation, which is developed on the posterior side of the beam and bears small posterior digitations [67]. The distalmost part of the tine is terminated with a very long anterior tine and another broader and more flattened tine separated from the anterior tine and the posterior palmation by broad and deep notches. The first tine is situated very close to the burr; the middle tine is not present. The very low position of the first tine approaches this deer to Rucervus simplicidens/colberti and R. gigans. Therefore, the South Asian origin of this species is quite probable. Nonetheless, the lack of sufficient data does not allow for the moment to demonstrate the relationships of A. insolitus with Rucervus from the Siwaliks and Apollonia-1.
Titov and Shvyreva [68] described another large-sized deer species Megaloceros stavropolensis from the Late Villafranchian of Ciscaucasia. This species is based on a large antler with the large fan-shaped distal palmation terminated by strong and long tines on its posterior portion. The first tine is situated at a certain distance from the burr. There are no middle and posterior tines. The cross-section of the beam is irregularly triangular. The antler beam bends backwards from the first ramification and is gently bowed sidewards [68]. The antler shape generally is very different from that of A. insolitus and it cannot be regarded as an individual morphological variant of the same species. Titov and Shvyreva [68] regard this cervid as a transitional form between R. (Arvernoceros) ardei and M. giganteus, thus following Heintz’s [25] hypothesized relationship between Arvernoceros and Megaloceros. Apparently, this cervid form is of East Asian origin. The general shape of palmation recalls the small-sized Dama sericus Teilhard de Chardin & Trassaert, 1937 from the Plio-Pleistocene formations of South-Eastern Shansi, China [44]. Grubb [69] included D. sericus in the genus Arvernoceros, and, in accordance with this viewpoint, M. stavropolensis was also included in Arvernoceros [44]. Nonetheless, it is possible that the Ciscaucasian deer is closely related to the Asian Sinomegaceros lineage, which, in its turn, could be phylogenetically related to Rucervus [44].
4.3. Morphological Similarities and Geographic Distribution of Rucervus radulescui and Rucervus gigans
The available cranial characteristics of R. radulescui generally correspond to the cranial morphology of modern R. duvaucelii: the nasal bones of the Romanian species are rather short and they do not reach the anterior orbital line, the preorbital fossae are large, the ethmoidal openings are equally large with long (ca. 1/2 of the orifice length) contact with the nasal bones, the basioccipital bone is broadened in the pharyngeal tuberosities. The difference with modern species concerns mostly the relatively shorter diastema in R. radulescui, which apparently is correlated with the comparatively shortened muzzle. The palatal width between the third molars of R. radulescui (70.7 mm; the female skull from ISER) seems to be somewhat broader than that in R. simplicidens (the skull No. 19829, AMNH), which amounts to 55 mm (measured from the Figure 145 in Colbert [36]).
R. radulescui shows the specific antler bauplan characteristic of the genus Rucervus (the missing middle tine, the posterior insertion of the crown tines that may occasionally be bifurcated in their distal parts). The distal portion of the antler is transformed into the relatively large bilobed palmation, however, the initial Rucervus-type construction of the antler crown is easily recognisable. The antler specialization is seen in the loss of the accessory prong on the first tine and the cylindrical shape of the first tine. This morphological variant is found in the antler 34590 (NHML) from Perrier-Etouaires, described as Cervus cylindroceros Dawkins, 1878. The distal portion of C. cylindroceros is terminated with a simple fork formed by the longer anterior tine, which may be interpreted as the continuation of the main beam, and the shorter posterior tine [70]. According to Heintz [25], C. cylindroceros represents an early ontogenetic stage of antler development of R. (Arvernoceros) ardei.
The antler proportions of R. radulescui permit to group this cervid form with European species R. ardei and R. verestchagini. These species share the relatively high position of the first (basal) tine and the very compact or even poor (as in R. verestchagini) development of the crown part. The crown part (palmation) attains less than 30% of the antler length in the type specimen of R. ardei and apparently the antler from Valea Grăunceanului is characterized by similar proportions. These shared antler characteristics permit to keep Arvernoceros Heintz, 1970 as a subgenus of Rucervus.
The dentition of R. radulescui remains quite primitive in its relatively long lower premolar series and the low degree of molarization of P4. The lost cingulum in the upper molars and the more complicated morphology of P3 and P4 represent the advanced dental features of R. radulescui. The dental diagnostic differences between R. radulescui and E. ctenoides are quite weak and are clearly visible when a large serial material is analyzed. Generally, R. radulescui is distinguished from E. ctenoides by the relatively longer lower premolar series and the stronger development of entoconid and posterior wing of metaconid in P3 and P4 with a trend towards the fusion of the posterior premolar elements. It should be noted that Eucladoceros generally shows a trend towards the stronger development of the anterior wing of the metaconid in P4 and its occasional fusion with the paraconid [25,53].
Unlike the species of the subgenus Arvernoceros, the South Asian Rucervus species, which correspondingly are included here in the nominotypical subgenus Rucervus, are characterized by the very low position of the first tine and the proportionally larger crown part of the antler. The crown part of the antler attains almost ½ of the antler length in R. duvaucelii [29,30,69], and apparently this is the case for R. simplicidens/colberti. The crown part of antler in R. schomburgki is even relatively longer (Figure 3, [69]).
The complete antlers of R. gigans are unknown, however, the low position of its first ramification resembles the antler from the Siwaliks figured by Colbert [36]. The dentition of R. gigans shares with R. simplicidens the weak development of cingulum in the upper third molar and the presence of a well-developed flat entostyle. These details of antler and dental morphology suggest that R. gigans should be included in the subgenus Rucervus being a peculiar South Asian element of the fauna from Apollonia-1.
4.4. Early Pleistocene Dispersals of Ruminants in Southeastern Europe
The earliest occurrence of Rucervus in Western Eurasia has been recorded in the Early Pliocene of Weze-1 (Poland) [62] and Kvabebi (Georgia). This roughly simultaneous occurrence of Rucervus in Eastern Europe and Transcaucasia suggests the general dispersal event of this genus in Western Eurasia and probably is the starting point of divergence between the subgenera Rucervus and Arvernoceros. Apparently, the origin of R. radulescui is related to the evolutionary branch of Rucervus evolved in southeastern Europe, since this species shares some details of its antler morphology with other species from the European continent (R. ardei, R. verestchagini), and its area of distribution includes the Balkan Peninsula and the North Pontic area. The western border of the distribution area, probably, is shaped by the ecological incompatibility with E. ctenoides. The West European cervid is characterized by a similar body size and a similar degree of evolutionary specialization. One can assume that those two deer generally depended on similar ecological resources, as it was demonstrated by the study of forage habits of large Early Pleistocene cervids from Western Europe and the Balkans [71]. The analysis of the dental wear of E. ctenoides from Chilhac, Sénèze, and Saint-Vallier and that of the cervid from Gerakarou—regarded in the present study as R. radulescui—shows the similar range of forage resource exploitation extending from browsing to mixed feeding [71]. However, according to the morphofunctional study of the postcranial skeleton that was carried out by Curran [5,6], R. radulescui and E. ctenoides were adapted to different types of landscape. The geometric morphometric analysis of limb bones demonstrated that E. ctenoides from Saint-Vallier and Sénèze inhabited the intermediate open environment, while R. radulescui was adapted to open and probably mountainous habitats [5,6]. Therefore, R. radulescui may be regarded as an ecological vicar species of E. ctenoides.
The associated with R. radulescui fauna is quite archaic with a certain oriental paleobiogeographic influence. The associated with R. radulescui fauna from Valea Grăunceanului is similar to those from Varshets and Slivnitsa, and it rather represents an archaic Villafranchian association of quite specialized and stenobiont species that existed in Europe before the “wolf event” (sensu Azzaroli [15]) or Pachycrocuta faunal turnover [16]. This is the case of sabertooth cats [4], Acinonyx pardinensis [72], the Villafranchian bovids Pliotragus, Gazellospira, and Procamptoceras, and the small Axis-like cervid Metacervocerus rhenanus [3]. According to Spassov [7], the wave of dispersals of new bovid forms from the East characterize the early Late Villafranchian Balkan faunas and indicates the progressive aridification of the region. The presence of Mitilanotherium, Gazellospira, Ovis, and Manis gives to the “pre-Pachycrocuta” Balkan faunas a rather Central and East Asian affinity [3,36,73].
The “post-Pachycrocuta” Balkan faunas show a greater similarity with Transcaucasian coeval faunas. The fauna from Fântâna lui Mitilan (MN18) and the middle faunal horizon (T-2) of the Irimești Area (Romania) are interesting by the earliest well-dated record of the large-sized deer Praemegaceros obscurus in Europe. The faunal assemblages from Fântâna lui Mitilan and T-2 of Irimești are correlated with Tasso Faunal Unit and Psekups [10,74] and share with the fauna from Dmanisi several cervid species with a quite restricted biogeographic distribution. P. obscurus is known by its abundant remains from Salcia (Moldova), Pietrafitta (Italy), and Forest Bed Formation (England). Some rare and very fragmentary remains of P. obscurus are also known from the Tamanian fauna of Semibalki-1 (Russia) [75]. This is the only species of the genus reported from the Early Pleistocene of the Near East [75]. The age of P. obscurus from Ubeidiya (Israel) generally corresponds to that of the findings of this deer from Italy. The remains of P. obscurus from Dmanisi (Georgia) [67] are older and they suggest the South Asian origin of this species. It is quite probable that P. obscurus is phylogenetically close to Panolia sp. from the Siwaliks [44].
Cervus nestii and Alces gallicus are two other cervid species shared by the faunas from Fântâna lui Mitilan and Dmanisi [73,76,77]. C. nestii is closely related to modern Cervus elaphus but shows much smaller body size and simple antlers lacking the bez tine and terminated with the simple fork [44]. The fine complete antler of this species from Dmanisi was described by Kahlke [78] as Cervus abesalomi [44,76]. The distribution of C. nestii in Europe was rather restricted and included only the Balkans and the Italian Peninsula. This species is regarded here as a warm-loving South Asian faunal element.
Alces gallicus is a side branch of Alces lineage with quite specialized large antlers, but primitive dentition with unmolarized P3 and Palaeomeryx fold in lower molars [44]. The remains of this species are known from Liventsovka (Russia) and Kuruksai (Tadjikistan) [34,79], therefore, the Central Asian origin for this species is suggested. The occurrence of A. gallicus in Dmanisi is interesting, since it indicates the two-way faunal exchange between Eastern Europe and the Transcaucasian Land. Some poor remains of Alces ex gr. minor-gallicus have been also reported from the Late Villafranchian fauna of the Denizli basin (Turkey) [80]. The faunal exchange between the Balkans and the Anatolian-Caucasian Land probably occurred due to the temporary closing of Bosporus that supposedly took place during this geological period [7] and it could be triggered by the 41 kyr glacial cycles. Spassov [7] indicated a presence of a Late Villafranchian paleobiogeographic unity of Southeastern Europe and Southwestern Asia analogous to the Greco-Iranian Late Miocene paleobiogeographic province. Apparently, the Balkan and Caucasian-Anatolian glacial refugia acted as communicating vessels during the cold phases. This is also the case of the Italian glacial refugium, which was biogeographically more open for faunal exchanges with the eastern refugia during the cold phases due to the sea level drop and the deforestation of the Dinaric Alps [39,73].
Possibly, the temporary closure of Bosporus during one of the cold phases of 41 kyr glacial cycles also enabled early hominins to disperse into the Balkan Peninsula.
The northern Peri-Pontic path of southern warm loving species dispersals (including Homo ex gr. erectus), as hypothesized by Ivanova et al. [81] and Strait et al. [9], seems to be less probable. First of all, the Late Villafranchian climate cooling that was triggered by the higher amplitude 41 kyr cycles caused the increase of seasonality, and, for the first time in Europe, the drop of the mean temperature of cold months below freezing point, as it was shown for Central Europe [82]. It is necessary to note that water obstacles should be practically insurmountable for warm-loving species in the conditions of cold climate. Therefore, the unfavourable climate conditions could impede warm-loving terrestrial mammals to disperse northward from the Caucasian mountain range. The mountain range of the Greater Caucasus protected the Caucasian land from the North and apparently created specific mild climate conditions, as in the case of large European southern peninsulas protected from the north by the Alpine-Himalayan mountain belt [73]. On the other hand, the Caucasian land at that time was a rather large peninsula that was separated from the West and from the East by Kujalnik (Black) and Akchagyl (Caspian) seas, and by the Manych Straight from the North [83]. This also explains the refugial character of the area of distribution of the endemic Pliocene holdover deer Praeelaphus sp. that survived in the fauna from Dmanisi (=Cervus ex gr. Arvernoceros ardei fide Vekua [84]) [44].
The dispersals of south Asian ruminants toward the Balkan Peninsula continued during the final stage of Late Villafranchian. This is the case of R. gigans from Greece, which coexists with P. pliotarandoides, a large-sized deer of supposedly Central Asian origin [44].
Bison (Eobison) tamanensis/Bison (Eobison) sp. with the larger area of distribution (Greece, Italy, Moldova, Ukraine, and South Russia) is another large Late Villafranchian ruminant that entered southeastern Europe from South Asia. Apparently, this primitive bison is related to B. (Eobison) georgicus from Dmanisi and B. (Eobison) sivalensis from Siwaliks.
The paleobiogeographic origin of the small-sized deer from the Early Pleistocene of the Near East and Anatolia is not clear due to the insufficient data. The antler remains from Ubeidya [85] and the Denizli basin [80] remind the European Metacervocerus rhenanus in their size and the high position of the first ramification. This cervid genus is known from the Siwalik fauna as the rather large species Metacervocerus punjabiensis, which is characterized by its pronounced mesodonty [61]. However, it is difficult to estimate the difference in the relative cheek tooth crowns height from the figures that are provided by Geraads [85] and Brown [61] first of all because the figured specimens are characterized by a different stage of wear, but if the difference in crown height of those two species will be demonstrated, the direct phyletic relationship between M. rhenanus and M. punjabiensis should be ruled out. Both M. rhenanus and M. punjabiensis are very close to Axis shansius from Shansi (China) [86]. The Chinese species is as large as M. punjabiensis but is distinguished by brachyodont dentition. The molarized P4 from Ubeidya figured by Geraads (Pl. II, [85]) may attest to the presence of the genus Dama in this fauna.
The problem of small-sized deer from the Near East is complicated by the presence of the Axis-like Cervus rewati from the Upper Siwaliks described on the basis of a mandible [56]. The antler morphology of this species remains unknown.
5. Conclusions
The antler bauplan remains quite conservative in the genus Rucervus during at list last three million years and it changes very little even in the specialized species with palmated antlers. The dispersal of the typical representatives of Rucervus in western Eurasia took place in the Early Pliocene and gave a regional radiation of medium to large species grouped in the subgenus Arvernoceros and represented by R. ardei from the Late Pliocene of Europe, R. verestchagini from the Early Pleistocene of Eastern Europe, R. giulii/dupuisi from the late Early Pleistocene of Europe, and the new species R. radulescui from Early Pleistocene of southeastern Europe. These species share the very high position of the first tine above the burr and the comparatively short crown part of their antlers. The crown may be poorly developed, as in R. verestchagini, or form a more or less developed distal palmation, as in R. ardei and R. radulescui.
The South Asian radiation of Rucervus is characterized by the low position of the first tine and relatively long crown part of the antler. Those species (R. duvaucelii, R. simplicidens, R. colberti, R. schomburgki, and the new species R. gigans) are included here in the nominotypical subgenus.
The described in the present study species R. radulescui and R. gigans come essentially from the same geographic area but they represent different paleobiogeographic unities: the “pre-Pachycrocuta” Villafranchian fauna of southeastern Europe with the central Asian paleobiogeographic influence, and the “post-Pachycrocuta” Villafranchian fauna with the clear south Asian paleobiogeographic influence. Apparently, the faunal exchanges between southeastern Europe and the Near East and southern Asia were controlled by the complex interaction of geographic obstacles, such as Bosporus and Manych Straight, the climate barrier from the north of the Greater Caucasus range, and the 41 kyr glacial cycles that repeatedly closed the Bosporus and thus triggered the two-way faunal exchange between southeastern Europe and the Near East, and, apparently, the further westwards dispersal of the archaic hominins in Eurasia.
Funding
The research was supported by the Royal Society (the Bonhote Grant, 2001), the NATO Science Programme/The Ministry of Economy of Greece (2001), and the Institute of Cultural Heritage of the Academy of Sciences of Moldova.
Acknowledgments
Many thanks to Vera Severianovna Bajgusheva, Aurelian Popescu, Georges Koufos, Dimitris S. Kostopoulos and Vadim Titov for their kind help during my work with collections under their care. I thank also Jean-Philippe Brugal and Nikos Solounias for sharing with me their opinions on some questionable specimens of fossil ruminants.
Conflicts of Interest
The author declares no conflicts of interest.
References
- Geist, V. Deer of the World: Their Evolution, Behaviour, and Ecology; Stackpole Books: Mechanicsburg, PA, USA, 1998; pp. 1–421. ISBN 0-8117-0496-3. [Google Scholar]
- Ungar, P.S.; Grine, F.E.; Teaford, M.F. Diet in Early Homo: A Review of the Evidence and a New Model of Adaptive Versatility. Annu. Rev. Anthrop. 2006, 35, 209–228. [Google Scholar] [CrossRef]
- Brugal, J.-P.; Croitor, R. Evolution, Ecology, and Biochronology of Herbivore Associations in Europe during the Last 3 Million Years. Quaternaire 2007, 18, 129–152. [Google Scholar] [CrossRef]
- Croitor, R.; Brugal, J.-P. Ecological and evolutionary dynamics of the carnivore community in Europe during the last 3 million years. Quat. Int. 2010, 212, 98–108. [Google Scholar] [CrossRef]
- Curran, S.C. Hominin Paleoecology and Cervid Ecomorphology. Ph.D. Dissertation, University of Minnesota, Minneapolis, MN, USA, 2009. [Google Scholar]
- Curran, S.C. Exploring Eucladoceros ecomorphology using geometric morphometrics. Anatom. Rec. 2015, 298, 291–313. [Google Scholar] [CrossRef] [PubMed]
- Spassov, N. The Plio-Pleistocene vertebrate fauna in South-Eastern Europe and the megafaunal migratory waves from the east to Europe. Rev. Paléobiol. 2003, 22, 197–229. [Google Scholar]
- Spassov, N. Southeastern Europe as a route for the earliest dispersal of Homo toward Europe: Ecological Conditions and the Timing of the First Human Occupation of Europe. In Paleoanthropology of the Balkans and Anatolia; Harvati, K., Roksandic, M., Eds.; Springer: Dordrecht, The Netherlands, 2016; pp. 281–290. ISBN 978-94-024-0873-7. [Google Scholar]
- Strait, D.S.; Orr, C.M.; Hodgkins, J.; Spassov, N.; Gurova, M.; Miller, C.; Tzankov, T. The Human Fossil Record of Bulgaria and the Formulation of Biogeographic Hypotheses. In Paleoanthropology of the Balkans and Anatolia; Harvati, K., Roksandic, M., Eds.; Springer: Dordrecht, The Netherlands, 2016; pp. 69–78. ISBN 978-94-024-0873-7. [Google Scholar]
- Radulescu, C.; Samson, P.M. The Plio-Pleistocene Mammalian Succession of the Olteț Valley, Dacic Basin, Romania. Quartärpaläontologie 1990, 8, 225–232. [Google Scholar]
- Radulescu, C.; Samson, P.M. Biochronology and evolution of the Early Pliocene to the Early Pleistocene mammalian faunas of Romania. Boll. Soc. Paleont. Ital. 2001, 40, 285–291. [Google Scholar]
- Ghenea, C. The Pliocene-Pleistocene boundary in Romania. In The Pleistocene Boundary and the Beginning of the Quaternary; Van Couvering, J.A., Ed.; Cambridge University Press: Cambridge, UK; New York/Port Chester, NY, USA; Melbourne/Sydney, Australia, 1997; Volume 41, pp. 216–220. ISBN 0521341159. [Google Scholar]
- Radulescu, C.; Samson, P.M.; Petculescu, A.; Stiuca, E. Pliocene large mammals of Romania. Coloq. Paleontol. 2003, 1, 549–558. [Google Scholar]
- Radulescu, C.; Samson, P.M.; Stiuca, E. Cadre biostratigraphique au Paléolithique inférieur en Roumanie. Quaternaire 1998, 9, 283–290. [Google Scholar] [CrossRef]
- Azzaroli, A. Quaternary mammals and the “End-Villafranchian” dispersal event—A turning point in the history of Eurasia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1983, 44, 113–139. [Google Scholar] [CrossRef]
- Martínez-Navarro, B. Early Pleistocene faunas of Eurasia and hominid dispersals. In Out of Africa I: Who? When? and Where? Vertebrate Paleobiology and Paleoanthropology Series; Fleagle, J.G., Shea, J.J., Grine, F.E., Baden, A.L., Leakey, R.E., Eds.; Springer: New York, NY, USA, 2010; pp. 207–224. ISBN 978-90-481-9036-2. [Google Scholar]
- Samson, P.M. Les Equides fossiles de Roumanie. Geol. Rom. 1975, 14, 165–352. [Google Scholar]
- Frost, S.; Ting, N.; Harcourt-Smith, W.; Delson, E. The positional and locomotor behavior of Paradolichopithecus arvernensis as inferred from the functional morphology of the postcrania. Am. J. Primatol. 2005, 66, S134–S135. [Google Scholar]
- Croitor, R. Systematical Position and Evolution of the Genus Arvernoceros (Cervidae, Mammalia) from Plio-Pleistocene of Eurasia. Muzeul Olteniei Craiova Oltenia Studii si communicari S tiintele Naturii 2009, 25, 375–382. [Google Scholar]
- Croitor, R. Deer from Late Miocene to Pleistocene of Western Palearctic: Matching fossil record and molecular phylogeny data. Zitteliana 2014, 32, 115–153. [Google Scholar]
- Kostopoulos, D.S. The Plio-Pleistocene artiodactyls (Vertebrata, Mammalia) of Macedonia 1. The fossiliferous site “Apollonia 1”, Mygdonia basin of Greece. Geodiversitas 1997, 19, 845–875. [Google Scholar]
- Koufos, G.D. New Material and Revision of the Carnivora, Mammalia from the Lower Pleistocene Locality Apollonia 1, Greece. Quaternary 2018, 1, 6. [Google Scholar] [CrossRef]
- Koufos, G.D.; Kostopoulos, D.S. Biochronology and succession of the Plio-Pleistocene macromammalian localities of Greece. Mémoires et travaux de l’Institut de Montpellier 1997, 21, 619–634. [Google Scholar]
- Koufos, G.D.; Syrides, G.E.; Kostopoulos, D.S.; Koliadimou, K.K. Preliminary results about the stratigraphy and the paleoenvironment of Mygdonia basin, Macedonia, Greece. Geobios 1995, 18, 243–249. [Google Scholar] [CrossRef]
- Heintz, E. Les Cervidés Villafranchiens de France et d’Espagne; Mémoires du Muséum national d’Histoire naturelle, Série C; Editions du Museum National d’Histoire Naturelle: Paris, France, 1970; t. I, 330p, t. II, 206p. [Google Scholar]
- Von den Driesch, A. A Guide to the Measurement of Animal Bones from Archaeological Sites; Peabody Museum Press: Salem, MA, USA, 1976; Volume 1, pp. 1–137. ISBN 9780873659505. [Google Scholar]
- Janis, C.M. Correlation of cranial and dental variables with body size in ungulates and macropodoids. In Body Size in Mammalian Paleobiology: Estimation and Biological Implications; Damuth, J., MacFadden, B.J., Eds.; Cambridge University Press: Cambridge, UK, 1990; pp. 255–299. ISBN 9780521360999. [Google Scholar]
- Hodgson, B.H. Proceedings of Learned Societies. Linnaean Society, Feb. 20, 1838. Ann. Nat. Hist. 1838, 1, 152–154. [Google Scholar]
- Hodgson, B.H. Specific name and character of a new species of Cervus, discovered by B. H. Hodgson, Esq. in 1825, and indicated in his Catalogue by the local name of Bahraiya. J. Asiatic Soc. Bengal 1835, 4, 648–649. [Google Scholar]
- Hodgson, B.H. Note on Cervus duvaucelii Cuvier, or C. elaphoides and Bahraiya of Hodgson. J. Asiatic Soc. Bengal 1836, 5, 240–242. [Google Scholar]
- Pocock, R.I. The Larger Deer of British India. Part II. J. Bombay Nat. Hist. Soc. 1943, 43, 553–572. [Google Scholar]
- Alekseyeva, L.I. Early Anthropogene Theriofauna of East Europe. Trans. Geol. Inst. 1977, 300, 1–214. (In Russian) [Google Scholar]
- Kostopoulos, D.S.; Athanassiou, A. In the shadow of bovids: Suids, cervids and giraffids from the Plio-Pleistocene of Greece. Quaternaire Hors-Serie 2005, 2, 179–190. [Google Scholar]
- Titov, V.V. Late Pliocene Large Mammals from Northeastern Sea of Azov Region; SSC RAS Publishing: Rostov-upon-Don, Russia, 2008; p. 264. ISBN 978-902982-47-0. (In Russian) [Google Scholar]
- Radulescu, C. Artiodactyles du Pliocène et du Pléistocène inférieur de Roumanie. Quaternaire Hors-série 2005, 2, 191–200. [Google Scholar]
- Colbert, E.H. Siwalik Mammals in the American Museum of Natural History. Trans. Am. Philos. Soc. 1935, 26, 1–401. [Google Scholar] [CrossRef]
- Kahlke, H.-D. Die Cerviden-Reste aus dem Unterpleistozän von Untermaßfeld. In Das Pleistozän von Untermaßfeld bei Meininghen (Thüringen); Verlag des Römisch-Germanischen Zentralmuseums Mainz: Mainz, Germany, 1997; Volume 40, pp. 181–275. [Google Scholar]
- Croitor, R.; Kostopoulos, D.S. On the Systematic Position of the Large-Sized Deer from Apollonia, Early Pleistocene, Greece. Palaeontol. Zeitsch. 2004, 78, 137–159. [Google Scholar] [CrossRef]
- Croitor, R.; Popescu, A. Large-sized ruminants from the Early Pleistocene of Leu (Oltenia, Romania) with remarks on biogeographical aspects of the “Pachycrocuta event”. Neues Jahrb. Geol. Paläontol.-Abhandlungen 2011, 261, 353–371. [Google Scholar] [CrossRef]
- Depéret, C.; Mayet, L. Le gisement de Senèze et sa faune paléomammalogique. Assoc. Fr. Avanc. Sci. C. R. 1911, 2, 261–263. [Google Scholar]
- Tsoukala, E.; Chatzopoulou, K. A New Early Pleistocene (Latest Villafranchian) Site with mammals in Kalamoto (Mygdonia Basin, Macedonia, Greece)—Preliminary Report. Mitt. Komm. Quartärforsch. Österr. Acad. Wiss. 2005, 14, 213–233. [Google Scholar]
- Spassov, N. Brief review of the Pliocene ungulate fauna of Bulgaria. Quaternaire Hors-Série 2005, 2, 201–212. [Google Scholar]
- Croitor, R.; Stefaniak, K.; Pawłowska, K.; Ridush, B.; Wojtal, P.; Stach, M. Giant deer Megaloceros giganteus Blumenbach, 1799 (Cervidae, Mammalia) from Palaeolithic of Eastern Europe. Quat. Int. 2014, 326–327, 91–104. [Google Scholar] [CrossRef]
- Croitor, R. Plio-Pleistocene Deer of Western Palearctic: Taxonomy, Systematics, Phylogeny; Inst. Zool. of the Acad. of Sc. of Moldova: Chișinău, Moldova, 2018; 140p. [Google Scholar]
- Vislobokova, I.A. Fossil deer of Eurasia. Trans. Paleont. Inst. 1990, 240, 1–206. (In Russian) [Google Scholar]
- David, A. A new deer species (Cervidae, Mammalia) from the late Pliocene deposits of Moldova. Bull. Acad. Şti. Rep. Moldova Ser. Şti. Chim. Biol. 1992, 1, 67–68. (In Russian) [Google Scholar]
- Bajgusheva, V.S. On skull construction of the large-sized deer of the Khaprovian mammal assemblage. In Paleoteriologia; Tatarinov, L.P., Ed.; Nauka: Moscow, Russia, 1994; pp. 236–252. ISBN 5-02-005723-1. (In Russian) [Google Scholar]
- Bajgusheva, V.S.; Titov, V.V.; Tesakov, A.S. The sequence of Plio-Pleistocene mammal faunas from the south Russian Plain (the Azov Region). Boll. Soc. Palaeontol. Ital. 2001, 40, 133–138. [Google Scholar]
- Vislobokova, I.; Sotnikova, M.; Dodonov, A. Late Miocene-Pliocene mammalian faunas of Russia and neighbouring countries. Boll. Soc. Palaeontol. Ital. 2001, 40, 307–313. [Google Scholar]
- Stehlin, H.G. Les sables de Rosières, près Saint-Florent (Cher). II. Paléontologie. Bull. Soc. Géol. Fr. 1912, 12, 198–212. [Google Scholar]
- Guerin, C.; Dewolf, Y.; Lautridou, J.-P. Revision of a famous site: Saint-Prest (Chartres, France). Geobios 2003, 36, 55–82. [Google Scholar] [CrossRef]
- Azzaroli, A. The Deer of the Weybourne Crag and Forest Bed of Norfolk. Bull. Br. Mus. 1953, 2, 1–96. [Google Scholar]
- Croitor, R.; Bonifay, M.-F. Etude préliminaire des cerfs du gisement pléistocène inférieur de Ceyssaguet (Haute-Loire). Paléo 2001, 13, 129–144. [Google Scholar]
- Azzaroli, A. Critical observations upon Siwalik Deer. Proc. Linn. Soc. Lond. 1954, 165, 75–83. [Google Scholar] [CrossRef]
- Croitor, R. Description of a new deer species (Cervidae, Mammalia) from the Early Pliocene of Eastern Europe, with a review of early dispersals and palaeobiogeography of the subfamily Cervinae. Neues Jahrb. Geol. Paläontol.-Abhandlungen 2017, 283, 85–108. [Google Scholar] [CrossRef]
- Arif, M.; Shah, S.M.I.; de Vos, J. Cervus rewati sp. nov. (Mammalia, Cervidae) from the Upper Siwaliks of Pakistan. Mem. Geol. Surv. Pak. 1992, 17, 13–23. [Google Scholar]
- Lydekker, R. Indian Tertiary and post-Tertiary Vertebrata. Part II. Molar Teeth and other remains of Mammalia. Palaeontol. Ind. Mem. Geol. Surv. Ind. 1876, 1, 19–87. [Google Scholar]
- Lydekker, R. Indian Tertiary and post-Tertiary Vertebrata. Part IV. Supplement to crania of ruminants. Palaeontol. Ind. Mem. Geol. Surv. Ind. 1880, 1, 172–181. [Google Scholar]
- Lydekker, R. Indian Tertiary and post-Tertiary Vertebrata. Palaeontol. Ind. Mem. Geol. Surv. Ind. 1884, 3, 1–258. [Google Scholar]
- Koken, E. Ueber fossile Saugethiere aus China. Dames und Kayser Palaeontologische Abhandlungen 1885, 3, 31–114. [Google Scholar]
- Brown, B. A new deer form from the Siwaliks. Am. Mus. Novit. 1926, 242, 1–6. [Google Scholar]
- Croitor, R.; Stefaniak, K. Early Pliocene deer of Central and Eastern European regions and inferred phylogenetic relationships. Palaeontographica 2009, 287, 1–39. [Google Scholar] [CrossRef]
- Flerov, K.K. Family Cervidae Gray 1821. In Basics of Paleontology, Mammals; Orlov, Y.A., Gromova, V.I., Eds.; Nedra: Moscow, Russia, 1962; Volume 13, pp. 368–378. ISBN 978-5-458-31474-9. (In Russian) [Google Scholar]
- Flerov, K.K.; Shevyreva, N.S. Pseudalces, Pliocene deer from Ciscaucasus. Paleont. J. 1963, 3, 120–123. (In Russian) [Google Scholar] [CrossRef]
- Vislobokova, I.A. Elks in the Pliocene of the USSR. Quartärpaläontologie 1986, 6, 239–242. [Google Scholar]
- Vekua, A. Kvabebi Fauna of Akchagylian Vertebrates; Nauka: Moscow, Russia, 1972; UDK 566(118.2); 351p. (In Russian) [Google Scholar]
- Vekua, A.; Bendukidze, O.; Kiladze, S. Gigantic deer in Plio-Pleistocene. Proc. Georgian Nation Mus. 2010, 2, 38–53. (In Georgian) [Google Scholar]
- Titov, V.V.; Shvyreva, A.K. Deer of the Genus Megaloceros (Mammalia, Cervidae) from the Lower Pleistocene of Ciscaucasia. Paleont. J. 2016, 1, 82–90. (In Russian) [Google Scholar]
- Grubb, P. Cervidae of Southeast Asia. In Horns, Pronghorns, and Antlers: Evolution, Morphology, Physiology, and Social Significance; Bubenik, G.A., Bubenik, A.B., Eds.; Springer: New York, NY, USA, 1990; pp. 169–179. ISBN 978-1-4613-8968-2. [Google Scholar]
- Dawkins, W.B. Contributions to the history of the Deer of the European Miocene and Pliocene strata. Q. J. Geol. Soc. 1878, 34, 402–420. [Google Scholar] [CrossRef]
- Berlioz, É.; Kostopoulos, D.S.; Blondel, C.; Merceron, G. Feeding ecology of Eucladoceros ctenoides as a proxy to track regional environmental variations in Europe during the early Pleistocene. C. R. Palevol. 2018, 17, 320–332. [Google Scholar] [CrossRef]
- Spassov, N. Acinonyx pardinensis (Croizet et Jobert) remains from the Middle Villafranchian locality of Varshets (Bulgaria) and the Plio-Pleistocene history of the cheetahs in Eurasia. Estud. Geol. 2011, 67, 245–253. [Google Scholar] [CrossRef][Green Version]
- Croitor, R. Paleobiogeography of early human dispersal in western Eurasia: Preliminary results. C. R. Palevol. 2018, 17, 276–286. [Google Scholar] [CrossRef]
- Radulescu, C.; Samson, P.M. Les mammifères du Pléistocène inferieur d-Izvoru (Departement d’Olt, Roumanie). Quartärpaläontologie 1986, 6, 157–171. [Google Scholar]
- Croitor, R. Taxonomy and systematics of large-sized deer of the genus Praemegaceros Portis, 1920 (Cervidae, Mammalia). Courier-Forschungsinstitut Senckenberg 2006, 256, 91–116. [Google Scholar]
- Bukhsianidze, M. Dmanisi artiodactyl assemblage. In Programme and Abstract Book, Proceedings of the XIV Annual Meeting of the European Association of Vertebrate Palaeontologists, Haarlem, The Netherlands, 6–10 July 2016; Holwerda, F., Madern, A., Voeten, D., van Heteren, A., Meijer, H., den Ouden, N., Eds.; KNAW: Haarlem, The Netherlands, 2016; p. 37. [Google Scholar]
- Bukhsianidze, M.; Hertler, C. Large Mammals from the Akhalkalaki Site. In The Role of the Southern Caucasus on Early Human Evolution and Expansion—Refuge, Hub, or Source Area? In Proceedings of the Workshop, Tbilisi, Georgia, 15–20 October 2013; Bruch, A.A., Lordkipanidze, D., Eds.; Georgian National Museum: Tbilisi, Georgia, 2013; p. 25. [Google Scholar]
- Kahlke, H.-D. Neufunde fon Cerviden-resten aus dem Unterpleistozän von Untermaßfeld. Das Pleistozän von Untermaßfeld bei Meningen (Thüringen), Teil 2. Romisch-Germanisches Zentralmuseum 2001, 40, 461–482. [Google Scholar]
- Vislobokova, I.A.; Sotnikova, M.V.; Erbaeva, M.A. The Villafranchian mammalian faunas of the Asiatic part of former USSR. Il Quaternario 1995, 8, 367–376. [Google Scholar]
- Boulbes, N.; Mayda, S.; Titov, V.V.; Alçiçek, M.C. Les grands mammifères du Villafranchien supérieur des travertins du Bassin de Denizli (Sud-Ouest Anatolie, Turquie). L’Anthropologie 2014, 118, 44–73. [Google Scholar] [CrossRef]
- Ivanova, S.; Gurova, M.; Spassov, N.; Popov, V.; Makedonska, J.; Tzankov, T.; Strait, D.S. Preliminary Findings of the Balkan Paleo Project: Evidence of Human Activity at the “Gateway” of Europe during the Late Pleistocene. Be-JA 2012, 2, 1–24. [Google Scholar]
- Mosbrugger, V.; Utescher, T.; Dilcher, D.L. Cenozoic continental climatic evolution of Central Europe. Proc. Natl. Acad. Sci. USA 2005, 102, 14964–14969. [Google Scholar] [CrossRef] [PubMed]
- Gabunia, L.; Vekua, A.; Lordkipanidze, D. The environmental contexts of early human occupation of Georgia (Transcaucasia). J. Hum. Evol. 2000, 38, 785–802. [Google Scholar] [CrossRef] [PubMed]
- Vekua, A. Die Wirbeltierfauna des Villafranchium von Dmanisi und ihre Biostratigraphische Bedeutung. Jahrb. Römisch-Ger. Zent.mus. Mainz. 1995, 42, 77–180. [Google Scholar]
- Geraads, D. Les Ruminants du Pleistocène d’Oubeidiyeh (Israël). In Les Mammifères du Pléistocène Inférieur de la Vallée du Jourdain à Oubeidiyeh; Tchernov, E., Ed.; Mémoires et Travaux du Centre de Recherche Français de Jerusalem: Paris, France, 1986; Volume 5, pp. 145–173. [Google Scholar]
- Teilhard de Chardin, P.; Trassaert, M. The Piocene Camelidae, Giraffidae, and Cervidae of South Eastern Shansi. Palaeont. Sin. 1937, 1, 1–69. [Google Scholar]
















© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).