Abstract
In dairy processing environments, many bacterial species form biofilms on surfaces and equipment. Among them, Listeria monocytogenes and Pseudomonas spp. could be present in mixed-species biofilms with increased resistance to disinfectants. This study aimed to evaluate the interactions between L. monocytogenes and P. fluorescens in dual-species biofilms simulating dairy processing conditions as well as the capability of P. fluorescens to produce the blue pigment. The biofilm-forming capability of single- and mixed-cultures was evaluated on polystyrene (PS) and stainless steel (SS) surfaces at 12 °C for 168 h. Biofilm biomass was assessed by crystal violet staining, the planktonic and sessile cells were quantified in terms of Colony Forming Unit (CFU), and the carbohydrates were quantified by the anthrone method. The biofilms were also observed through Confocal Laser Scanning Microscopy (CLSM) analysis. Results showed that only P. fluorescens was able to form biofilms on PS. In dual-species biofilms at the end of the incubation time, a lower biomass compared to P. fluorescens mono-species was observed. On the SS, the biofilm cell population of L. monocytogenes was higher in the dual-species than in the mono-species, particularly after 48 h. Carbohydrates in the dual-species system were higher than those of the mono-species and were also revealed at 168 h. The production of blue pigment by P. fluorescens in the Ricotta-based model system was revealed in both single cultures and co-cultures and was confirmed by the CLSM results, showing agglomeration, probably linked to the blue pigment. Our study suggests that the interactions between the two species can influence biofilm formation, but not the capability of P. fluorescens to produce blue pigment.
1. Introduction
Microbial biofilms are three-dimensional structures of various bacteria that adhere to biotic or abiotic surfaces and differentiate into complex communities, embedded within extracellular polymeric substances (EPS) [1]. The relevance of microbial biofilms has been described in different fields including the food industry where biofilms are responsible for potential food contamination, corrosion, and economic losses [2]. Particularly in the dairy industry, many bacterial species adhere to and form biofilms on surfaces and equipment. Among them, Listeria monocytogenes and different species of Pseudomonas [3] are worthy of attention. Listeria monocytogenes is a ubiquitous pathogen, able to colonize, and persist on common surfaces in food processing environments [4]; it can also contaminate a wide variety of foods such as dairy products. The microbiota of refrigerated foods is dominated by selected microorganisms, including Pseudomonas spp. In particular, P. fluorescens can form biofilms on the surfaces of dairy processing plants [5] and produce pigments such as pyoverdine, pyocyanin, and indigoidine, which are responsible for dairy products’ blue discolouration [6].
Biofilms found in nature are generally formed by two or more microbial species; in fact, multi-species biofilms are commonly encountered in food and food-related environments [7]. Multi-species biofilms possess properties unavailable in mono-species biofilms, which may provide advantages to microorganisms such as an increase in tolerance against stressful conditions and the capability to degrade organic compounds [8].
The objective of this study was to evaluate dual-species biofilms formed by L. monocytogenes and P. fluorescens in a system simulating real conditions encountered in dairy processing. This was done by using (i) surfaces of PS and SS; (ii) L. monocytogenes and blue-pigmenting P. fluorescens strains, isolated from dairy products; (iii) a Ricotta-based model system as the growth medium; and (iv) 12 °C as the incubation temperature.
2. Materials and Methods
Eight strains of Listeria monocytogenes were tested together with one strain of blue-pigmenting Pseudomonas fluorescens pf5 isolated from mozzarella cheese [5]. L. monocytogenes strains, previously isolated from dairy products (LM 1-2-3-4) and dairy plants (LM 5-6-7-8), were typed and serotyped according to the Pulsed-Field Gel Electrophoresis (PFGE) and the US/FDA Bacteriological Analytical Manual [9]. To simulate a dairy processing environment, a Ricotta-based model system was prepared following the method described by de Carvalho et al. (2015) [10].
In order to examine the biofilm-forming capability of the strains in mono- and dual-species and to select one combination, the biofilm study was first assessed on PS. The bacterial suspensions (105 CFU/mL), prepared in the Ricotta-based model system , were aliquoted on PS microtitre plates and then incubated at 12 °C for 168 h. The total biomass was quantified at 590 nm by crystal violet assay [5].
To evaluate biofilm formation on stainless steel (AISI 304 coupons), L. monocytogenes LM5 strain was chosen and combined with P. fluorescens pf5. Sterile glass containers with coupons and the inoculated Ricotta-based model system were incubated at 12 °C for 168 h. At 0, 48, 72, 96, and 168 h, planktonic cells were enumerated on selective media for Pseudomonas spp. (Pseudomonas agar base) and for L. monocytogenes (Agar Listeria according to Ottaviani & Agosti). The enumeration of cells in biofilms (sessile cells) was performed by scraping the surfaces with cotton swabs to collect and enumerate the cells [11].
The EPS were extracted as previously reported by Abdallah et al. (2015) [12], with some modifications, while carbohydrates quantification was carried out following the anthrone method [13]. The absorbance of the samples at 625 nm was measured and the results were presented in µg/cm2. Mono- and dual-species biofilm structures were observed by CLSM according to the method described by Rossi et al. (2018) [2]. The biofilms were stained with the LIVE/DEAD BacLight Bacterial Viability kit. The fluorescence of pyoverdine, the siderophore produced by P. fluorescens, was also checked [14]. Data of biofilm assays were subjected to analysis of variance (ANOVA) and pair-comparison within the same group was achieved by applying Dunnett’s test procedure at *p <0.05 using XLSTAT ver. 2017 (Addinsoft, Paris, France).
3. Results and Discussion
3.1. Serotype and Pulsotype of L. monocytogenes Strains
Four serotypes (1/2a, 1/2b, 1/2c, and 4b) were identified among the eight L. monocytogenes strains. The most prevalent serotype was 1/2b (for strains isolated from both food and environmental sources), then 1/2a (for food strains) and 1/2c (for environmental strains), followed by 4b (for the mozzarella cheese isolate). A total of eight ApaI and eight AscI PFGE types were distinguished, thus revealing that the strains isolated from food products and the environment were gentically different and heterogeneous.
3.2. Biofilm Formation on the Polystyrene Surface
None of the eight L. monocytogenes strains were able to form biofilms on PS (data not shown). However, numerous studies have demonstrated that this pathogen is able to form biofilms on various surfaces [15,16], and a previous study reported low Optical Density (OD) values for L. monocytogenes biofilms [17]. On the other hand, P. fluorescens exhibited good biofilm formation capacity, with increased biomass during incubation time. However, a different behaviour was observed for the species in combination, with variability among the strains. At the end of the incubation period, biofilms in dual-species systems were significantly lower than the single ones (* p < 0.05), and a higher biofilm biomass for the combinations P. fluorescens—L. monocytogenes LM5 was noticed after 72 h. With respect to the blue discolouration, P. fluorescens blue pigment production was observed both in single and mixed cultures after 72 h. Based on the obtained results, the L. monocytogenes LM5 and P. fluorescens pf5 combination was selected for subsequent analysis.
3.3. Biofilm Formation on the Stainless Steel Surface and Enumeration of Planktonic and Sessile Cells
The results of L. monocytogenes LM5 and P. fluorescens pf5 planktonic and sessile cells on SS coupons are presented in Figure 1.
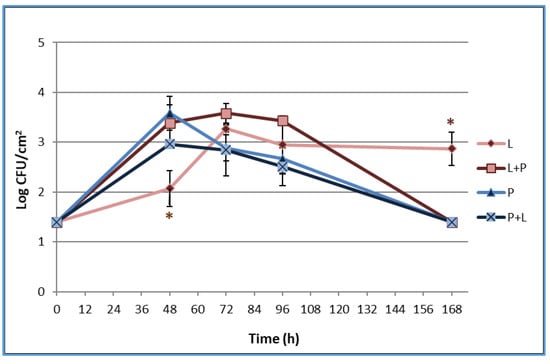
Figure 1.
Dynamics of sessile cells of L. monocytogenes LM5 and P. fluorescens pf5 in mono- and dual-species conditions on stainless steel (SS) coupons at 12 °C for 168 h. The asterisk (*) means statistically significant difference between the mono- and dual-species of each strains for the same incubation time (* p < 0.05). L: L. monocytogenes in single-species; L + P: L. monocytogenes in dual-species; P: P. fluorescens in single-species; P + L: P. fluorescens in dual-species.
Regarding the planktonic phenotype (data not shown), the presence of P. fluorescens determined a slight decrease of L. monocytogenes counts at 48 and 96 h. P. fluorescens showed a greater increase in load over time compared to L. monocytogenes, and planktonic counts did not significantly differ between mono- and dual-species.
The results regarding the sessile populations (Figure 1) showed that L. monocytogenes was able to adhere on the SS surface, probably because of the affinity established between the charge of the cell surface and the anchoring site. In fact, at low temperatures, L. monocytogenes increases the hydrophilicity and, therefore, the affinity to hydrophilic surfaces such as steel [18]. The pathogen load in mono-species increased up to 3.27 log CFU/cm2 at 72 h with no particular changes during the time. In multi-species conditions, the presence of P. fluorescens statistically (*p <0.05) increased the pathogen biofilms after 48 h when it reached a sessile population of 3.39±0.36 log CFU/cm2. This result is in line with those reported by Puga et al. (2018) [17], who observed a stimulation of L. monocytogenes adhesion in mixed-culture biofilms with P. fluorescens. The authors linked the positive effects on L. monocytogenes to Pseudomonas production of proteinases able to mobilize essential amino acids. At the end of the experimental time, L. monocytogenes LM5 sessile cells in the mixed-culture dropped to 1.4 log CFU/cm2. The observed fast cellular dispersal for multi-species biofilms could have been stimulated from the early achievement of a high biofilm level with no extra nutrient supplementation [17]. The P. fluorescens sessile population reached 3.58 ± 0.34 log CFU/cm2 at 48 h and decreased over time, reaching 1.4 log CFU/cm2 after 168 h, with no statistically significant differences among single- and mixed-cultures. The blue pigment production of P. fluorescens was observed starting from 72 h both in single- and in mixed-cultures (Figure 2) when the highest load of the spoilage microrganism (planktonic cells) was detected (data not shown). In agreement with our findings, Andreani et al. (2015) [19] observed an evident blue pigment in broth when Pseudomonas counts reached about 7 × 108 CFU/mL and concluded that the blue pigment production took place in the late logarithmic phase.

Figure 2.
(A) Blue pigment colour appearance during the evaluation of biofilm formation on SS coupons in glass containers. From the left: control, P. fluorescens pf5, L. monocytogenes LM5, and the dual-species system. (B) Blue pigment colour evolution during the CLSM analysis: (B1) 48 h, (B2) 72 h, (B3) 96 h, and (B4) 168 h.
3.4. EPS Analysis by Carbohydrates Quantification
The total amount of carbohydrates in the biofilms was affected by the time and the species involved in biofilm formation (Figure 3). The biofilm carbohydrates content increased over time in single-species biofilms, with the greatest increase occurring at 96 h. No carbohydrates were revealed at 168 h for both single-species biofilms. In dual-species conditions, a higher yield in carbohydrates in comparison with the single species was detected at 72 h. Remarkably, the carbohydrates of the dual-species biofilms were also revealed at the end of the experimental time. Puga et al. (2018) [17] reported that the inclusion of L. monocytogenes to the already established P. fluorescens biofilms increased matrix production.
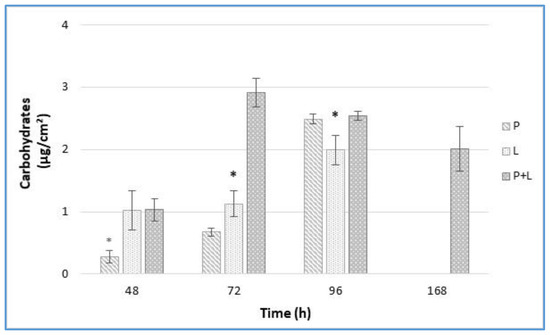
Figure 3.
Carbohydrates amount (µg/cm2) from L. monocytogenes LM5 and P. fluorescens pf5 biofilms on SS coupons in mono- and dual-species at 12 °C for 168 h. The asterisk (*) means statistically significant difference between the mono- and dual-species of each strains for the same incubation time (* p < 0.05). P: P. fluorescens in single-species; L: L. monocytogenes in single-species; P + L: P. fluorescens and L. monocytogenes in dual-species.
3.5. Confocal Laser Scanning Microscopy Analysis
As observed from CLSM analysis (Figure 4), no three-dimensional biofilm architecture was revealed at 168 h. In fact, Figure 4 shows green agglomerates containing damaged or dead cells (red cells according to the propidium iodide staining) and detached P. fluorescens cells (the blue colour of pyoverdine fluorescence). The fact that the agglomerates were not clearly identifiable as cells, and that they were present only in the samples with P. fluorescens, suggests that they could depend on blue pigment formation. In addition, this particular behaviour was observed starting from 72 h in correspondence with the blue pigment formation by P. fluorescens. Figure 2a–d revealed that blue pigment discolouration starting from 72 h turned into a green/grey colour. The colour change highlights the possible reduction of indigoidine to leucoindigoidine, which is considered a chemical marker of blue discolouration [20].
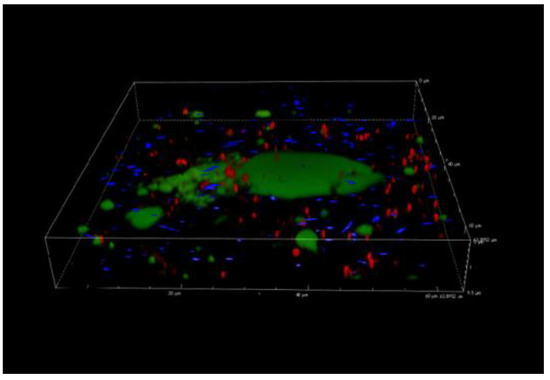
Figure 4.
Confocal laser scanning microscopy analysis of L. monocytogenes LM5 and P. fluorescens pf5 biofilms in dual-species conditions after 168 h at 12 °C.
4. Conclusions
Our results showed that the behaviour of planktonic and sessile populations on SS coupons was strongly dependent on the culture conditions (mono-/dual-species). In fact, the presence of P. fluorescens increased the L. monocytogenes sessile population and the total EPS carbohydrates amount on the SS coupons. Nevertheless, further studies including the comparison of the gene expression between single- and dual- species biofilms and interactions in terms of the volatilome would be useful to provide more information on the inter-species consortium.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data available on request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Kassinger, S.J.; Van Hoek, M.L. Biofilm architecture: An emerging synthetic biology target. Synth. Syst. Biotechnol. 2020, 5, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Rossi, C.; Chaves-López, C.; Serio, A.; Anniballi, F.; Valbonetti, L.; Paparella, A. Effect of Origanum vulgare essential oil on biofilm formation and motility capacity of Pseudomonas fluorescens strains isolated from discolored Mozzarella cheese. J. Appl. Microbiol. 2018, 124, 1220–1231. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, G.S.; Lopes, D.R.G.; Andre, C.; Silva, C.C.; Baglinière, F.; Vanetti, M.C.D. Multispecies biofilm formation by the contaminating microbiota in raw milk. Biofouling 2019, 35, 819–831. [Google Scholar] [CrossRef] [PubMed]
- Di Ciccio, P.; Rubiola, S.; Grassi, M.A.; Civera, T.; Abbate, F.; Chiesa, F. Fate of Listeria monocytogenes in the Presence of Resident Cheese Microbiota on Common Packaging Materials. Front. Microbiol. 2020, 11, 830. [Google Scholar] [CrossRef] [PubMed]
- Rossi, C.; Serio, A.; Chaves-López, C.; Anniballi, F.; Auricchio, B.; Goffredo, E.; Goga, B.T.C.; Lista, F.R.; Fillo, S.; Paparella, A. Biofilm formation, pigment production and motility in Pseudomonas spp. isolated from the dairy industry. Food Control. 2018, 86, 241–248. [Google Scholar] [CrossRef]
- Reichler, S.J.; Martin, N.H.; Evanowski, R.L.; Kovac, J.; Wiedmann, M.; Orsi, R. A century of gray: A genomic locus found in 2 distinct Pseudomonas spp. is associated with historical and contemporary color defects in dairy products worldwide. J. Dairy Sci. 2019, 102, 5979–6000. [Google Scholar] [CrossRef] [PubMed]
- Nychas, G.-J.E.; Nychas, G.-J.E.; Koutsoumanis, K. Strain variability in biofilm formation: A food safety and quality perspective. Food Res. Int. 2020, 137, 109424. [Google Scholar] [CrossRef]
- Røder, H.L.; Sørensen, S.J.; Burmølle, M. Studying Bacterial Multispecies Biofilms: Where to Start? Trends Microbiol. 2016, 24, 503–513. [Google Scholar] [CrossRef] [PubMed]
- Londero, A.; Costa, M.; Galli, L.; Brusa, V.; Linares, L.; Prieto, M.; Leotta, G. Characterization and subtyping of Listeria monocytogenes strains from butcher shops. LWT-Food Sci. Technol. 2019, 113, 108363. [Google Scholar] [CrossRef]
- De Carvalho, R.J.; De Souza, G.T.; Honório, V.G.; De Sousa, J.P.; Da Conceição, M.L.; Maganani, M.; De Souza, E.L. Comparative inhibitory effects of Thymus vulgaris L. essential oil against Staphylococcus aureus, Listeria monocytogenes and mesophilic starter co-culture in cheese-mimicking models. Food Microbiol. 2015, 52, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, J.B.D.S.; De Souza, E.L.; Scarano, J.O.A.; De Sousa, J.M.; Lira, M.C.; De Figueiredo, R.C.B.Q.; De Souza, E.L.; Magnani, M.; Souza, N.; Alcântara, J.O.; et al. Efficacy of using oregano essential oil and carvacrol to remove young and mature Staphylococcus aureus biofilms on food-contact surfaces of stainless steel. LWT 2018, 93, 293–299. [Google Scholar] [CrossRef]
- Abdallah, H.M.I.; Asaad, G.F.M.; Nada, S.A.; Taha, H.S.; Seif El-Nasr, M.M. Influence of extract derived in-vitro cell suspension cultures of Echinacea purpurea against some immunosuppressive effects. Res. J. Pharm. Biol. Chem. Sci. 2015, 6, 1136–1143. [Google Scholar]
- Harimawan, A.; Ting, Y.P. Investigation of extracellular polymeric substances (EPS) properties of P. aeruginosa and B. subtilis and their role in bacterial adhesion. Colloids Surf. B Biointerfaces 2016, 146, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Martin, N.; Murphy, S.; Ralyea, R.; Wiedmann, M.; Boor, K. When cheese gets the blues: Pseudomonas fluorescens as the causative agent of cheese spoilage. J. Dairy Sci. 2011, 94, 3176–3183. [Google Scholar] [CrossRef] [PubMed]
- Mendez, E.; Walker, D.K.; Vipham, J.; Trinetta, V. The use of a CDC biofilm reactor to grow multi-strain Listeria monocytogenes biofilm. Food Microbiol. 2020, 92, 103592. [Google Scholar] [CrossRef] [PubMed]
- Skowron, K.; Brożek, K.; Łukasik, M.; Wiktorczyk, N.; Korkus, J.; Gospodarek-Komkowska, E. Assessment of drug susceptibility and biofilm formation ability by clinical strains of Listeria monocytogenes. Disaster Emerg. Med. J. 2020, 5, 12–18. [Google Scholar] [CrossRef]
- Puga, C.H.; Dahdouh, E.; Sanjose, C.; Orgaz, B. Listeria monocytogenes Colonizes Pseudomonas fluorescens Biofilms and Induces Matrix Over-Production. Front. Microbiol. 2018, 9, 1706. [Google Scholar] [CrossRef] [PubMed]
- Chavant, P.; Gaillard-Martinie, B.; Talon, R.; Hébraud, M.; Bernardi, T. A new device for rapid evaluation of biofilm formation potential by bacteria. J. Microbiol. Methods 2007, 68, 605–612. [Google Scholar] [CrossRef] [PubMed]
- Andreani, N.A.; Carraro, L.; Martino, M.E.; Fondi, M.; Fasolato, L.; Miotto, G.; Magro, M.; Vianello, F.; Cardazzo, B. A genomic and transcriptomic approach to investigate the blue pigment phenotype in Pseudomonas fluorescens. Int. J. Food Microbiol. 2015, 213, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Del Olmo, A.; Calzada, J.; Nuñez, M. The blue discoloration of fresh cheeses: A worldwide defect associated to specific contamination by Pseudomonas fluorescens. Food Control. 2018, 86, 359–366. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).