Abstract
Drought will likely become more frequent and intense in Europe due to climate change, which may worsen Mn2+ and P deficiencies found in high pH soils. In this context, research investigating the effectiveness of ammonium-based nitrogen fertilizers treated with nitrification inhibitors (NIs) in alleviating Mn2+ and P deficiencies in such soils has been done. However, studies considering the impact of drought periods and soil texture on this topic are lacking. Therefore, we carried out a study addressing this research gap. Maize plants were grown in a greenhouse experiment, and the experimental setup comprised three factors consisting of soil texture (sand and silt loam), soil moisture (sufficient and drought), and DMPP application (with and without DMPP). The measured variables were bulk and rhizosphere soil pH, Mn2+ availability, maize biomass yield, and shoot concentration of selected macro- and micronutrients. DMPP increased shoot biomass production by 60% in silt loam under drought but not in sand soil texture. In addition, DMPP increased Mn2+ and P shoot concentrations by 38% and 21%, respectively, in the silt loam soil texture under drought. In contrast, DMPP did not alleviate the negative impact of drought on plant biomass production, Mn2+ and P shoot concentration in the sand soil texture. In conclusion, DMPP application is effective in alleviating Mn2+ and P deprivation in high pH soil subjected to drought. However, this effect was soil texture-dependent and observed in the silt loam rather than in the sand soil texture.
1. Introduction
Deficiency of Mn2+ and P is a common threat in high pH and humic soils in Northern Germany, the Netherlands, and Denmark [1,2]. Furthermore, drought might become more common in these regions due to climate change, which may worsen Mn2+ and P deficiencies, resulting in significant crop yield losses. For example, Grillakis [3] suggested that a reduction in soil moisture is expected to increase the frequency, duration, and spatial expansion of drought in Europe. In addition, the number of drought events is predicted to increase from 40 to 88, considering historical data (1966–2006) and the predictions for 2060–2099, respectively [3,4]. During soil drying, the increased oxygen (O2) availability stimulates Mn2+ oxidation, which may exacerbate Mn2+ deficiency in soils with high pH and elevated organic matter content. High soil organic matter immobilizes Mn2+, forming stable complexes and therefore decreasing Mn2+ availability. Additionally, diminished nutrient uptake via mass flow and diffusion can aggravate Mn2+ and P deprivation under drought in such soils.
A supply of high pH soils with ammonium-nitrogen (NH4+-N)-based fertilizers has been shown to enhance Mn2+ and P availability and uptake through nitrification-induced acidification of bulk soil [5,6]. Conversely, in such instances where the microbial oxidation of NH4+ occurs, the H+ cotransport with nitrate uptake causes the rhizosphere alkalinization. Consequently, this process renders Mn2+ and P less available for uptake and may reduce plant development, growth, and yield production.
Therefore, application of nitrification inhibitors (NIs) along with sole NH4+-N nutrition has been suggested to increase Mn2+ and P availability in high pH soils due to the rhizosphere acidification [7,8,9,10,11]. In addition, among commonly utilized NIs, 3,4-dimethylpyrazole phosphate (DMPP) has been suggested to have superior effectiveness in suppressing nitrification relative to other NIs such as dicyandiamide [12].
A meta-analysis including 92 peer-reviewed manuscripts demonstrated that the addition of NIs to NH4+-N sources increases P uptake by 8.5% [13]. Rhizosphere acidification is especially important for P due to its low soil mobility once it moves within the soil by diffusion; therefore, increasing its availability near the root surface is of paramount importance. Furthermore, unlike P, previous studies indicated that Mn2+ uptake is not favored by the application of NIs and rhizosphere acidification relative to supplying unstabilized NH4+-N [5]. This probably occurs because O2 consumed during nitrification plays an important role in Mn reduction due to the development of anaerobic microsites in the bulk soil, while NH4+ antagonistically affects Mn2+ uptake [1,14]. Moreover, in contrast to P, Mn2+ can move from the bulk soil through mass flow for plant uptake [15].
The aforementioned studies were carried out under sufficient soil moisture conditions. Therefore, the effectiveness of NIs in alleviating P and Mn2+ deficiencies in high pH soils under drought remains unclear. Drought decreases nutrient mass flow [16] and diffusivity [17], which aggravates Mn2+ and P deficiencies in high pH soils with high organic matter content [16,18,19]. Thus, identifying whether NIs can mitigate Mn2+ and P deficiency under drought is important for managing these soils in a future scenario considering climate change.
Soil texture may impact the effectiveness of NIs in alleviating Mn2+ and P deficiencies under drought. Fine-textured soils hold more water than coarse-textured ones at the same matric potential. Thus, the latter are more vulnerable to nutrient deficiency under drought because reducing soil water content decreases mass flow and diffusion. For instance, soils with water contents of 0.4 g cm−3 and 0.13 g cm−3 needed 10 μM P and 200 μM P, respectively, to reach the same P uptake by maize plants [20,21]. Moreover, fine texture has been suggested to decrease DMPP effectiveness, probably due to its adsorption to the negatively charged soil colloids and enhanced microbial activity in these soils that lead to faster DMPP biodegradation [12]. On the basis of the aforementioned, it is unknown if fine soil texture benefits the DMPP effect on increasing Mn2+ and P availability, as it holds more water, or if it has a negative impact due to its ability to lower the effectiveness of the inhibitor itself. Moreover, to the best of our knowledge, studies on this topic are lacking.
It is unknown whether the DMPP supply can alleviate Mn2+ and P deficiency and consequently promote their shoot concentration through rhizosphere acidification under drought conditions in fine and coarse-textured soils. The present study was conducted with sand and silt loam soil textures to investigate the effect of NH4+-N, with or without DMPP, on maize shoot biomass yield, pH, and Mn2+ availability in bulk and rhizosphere soils, as well as P and Mn2+ maize shoot concentrations. We hypothesized that (1) stabilizing NH4+-N by DMPP alleviates Mn2+ and P deficiencies in maize plants cultivated on high pH soils under drought, and (2) this effect is soil texture dependent.
2. Methods and Materials
2.1. Location and Soil Description
The study involved a greenhouse experiment carried out from March to April 2024 at the Experimental Station of the Institute of Plant Nutrition and Soil Science, Kiel University, Germany. Two differently textured soils collected in different locations in Northern Germany were investigated. The silt loam soil was collected from Friedrichskoog (54°01′34″ N, 8°86′10″ E), while the sandy soil was collected from Grevenkrug (54°12′45″ N, 10°0′37″ E) using a mechanized caterpillar excavator at a depth of 0 to 30 cm. A summary of elementary soil characteristics is shown in Table 1.

Table 1.
Elementary soil characteristics.
2.2. Experimental Setup
An experiment was set under a completely randomized design (CRD) with three factors consisting of soil texture (sand and silt loam), soil moisture (sufficient and drought), and DMPP application (with and without DMPP). Lime was added at a rate of 4 g CaCO3 kg soil−1 and evenly mixed with sand using a concrete mixer to achieve an alkaline pH similar to that of silt loamy soil. This liming rate was based on our previous study using the same sandy soil, in which the pH response of this soil to increasing lime rates was described in detail [6]. Liming was performed two weeks before fertilizer application, pots filling, and sowing. Additionally, N was applied to the soils at a rate of 1 g N pot−1, representing 167 mg N kg−1 soil. Moreover, DMPP was added to the N fertilizer at 1% of the added NH4+-N, while macro- and micronutrients were applied to the soils at the rates shown in Table S1. A cement mixer was used to incorporate lime, nutrients, and DMPP, which were applied on the same day the pots were filled with soil. Nutrient and DMPP were dissolved in deionized water for application. The use of DMPP for stabilization of NH4+-N was due to its high effectiveness, as reported in previous studies [12].
Before soil fertilization, soil from each texture was air-dried and sieved through a 2 mm sieve to remove stones and plant debris. Thereafter, fertilizers and DMPP were dissolved in deionized water and mixed with the soil using a concrete mixer. Subsequently, 5 L Mitscherlich pots were filled with 6 kg soil and initially irrigated to maintain WHC at 60% to allow optimum germination, sprouting, and establishment of the seedlings. Half of the pots were kept at 60% WHC throughout the experiment, and the other half were allowed to reach 30% WHC due to evapotranspiration to simulate drought conditions. For the 30% WHC treatment, soil moisture was kept at 60% WHC for seven days after sowing to ensure optimal germination and plant establishment. Only after that was the soil allowed to dry naturally until the 30% WHC was reached, 10 days after sowing (three days of natural drying). Soil moisture in each pot was monitored through daily weighing, and water loss could be replaced accordingly. The 30% WHCs for sand and silt loam soil textures were 0.08 g g−1 and 0.10 g g−1, respectively, while the 60% WHCs were 0.15 g g−1 and 0.21 g g−1 for sand and silt loam correspondingly.
Further, maize (Zea mays L.) cultivar KWS KEOPS was sown, and after germination, three uniform plants were kept in each pot. During the cultivation period, the plants were subjected to a day/night temperature of 22/16 °C, a photoperiod of 16/8 h (day/night), and a relative humidity of 65% and 85% at day and night, respectively. Lastly, the plants were harvested 28 days after sowing. On the harvesting day, the plants were at the vegetative stage and had five fully expanded leaves (Figure S1).
2.3. pH and Mn2+ Sampling and Measurement in Bulk and Rhizosphere Soils
Using a soil auger, samples referred to as bulk soil were taken from both soil textures 14 (V2 plant stage) and 28 (V5 plant stage) days after sowing. Additionally, rhizosphere soil was collected on day 28, and it was considered to be the soil tightly attached to the roots after manual shaking. Subsequently, the rhizosphere soil was meticulously removed from the roots using a brush, air-dried, and kept for laboratory analyses.
Soil samples were mixed with 0.01 M CaCl2 at a ratio of 1:2.5 and shaken for 120 min at 60 rpm. Then, the samples were filtered, and pH was measured in a pH meter (inoLab® Multi 9310 IDS, Xylem Analytics, Weilheim, Germany). With the same extracts, Mn2+ was measured by Atomic Absorption Spectroscopy (AAS, Thermo S Series, UniGreenScheme, Caerphilly, UK) as adapted from Dreyer et al. [5].
2.4. Bulk Soil Sampling and Measurement for Concentrations of NH4+-N and NO3−-N
Bulk soil samples were mixed with 1 M KCl in a ratio of 1:10 and filtered after a 1 h shaking. Thereafter, the concentration of NH4+-N and NO3−-N in the extracts was measured using a continuous flow analyzer (San++ Continuous Flow Analyzer, Skalar, Breda, The Netherlands). In addition, sub-samples of the bulk soils were oven-dried at 105 °C for 24 h for soil moisture content determination.
2.5. Plant Tissue Sample Preparation and Analysis
Fresh plant shoot samples were oven-dried for a week at a temperature of 65 °C. Subsequently, the samples were weighed to determine shoot biomass production and ground. Additionally, based on plant material digestion procedures provided by Jezek et al. [22], 200 mg of dry plant material powder was transferred into digestion tubes, and 10 mL 69% HNO3 acid (ROTIPU-RAN Supra for ICP, 69, Karlsruhe, Germany) was added. The sample mixtures were digested for 45 min in an 1800 W microwave system (MARS 6 Xpress; CEM, Matthews, MC, USA) before dilution of the digestate to 100 mL miscible solution using Milli-Q water. Finally, shoot concentrations of P, K+, Ca2+, Mg2+, S, Mn2+, Fe2+, Zn2+, and Cu2+ were analyzed using ICP-OES (Agilent Technologies 5800 Series, Santa Clara, CA, USA).
For shoot N concentration analysis, the ground oven-dried shoot tissue samples were measured in a CNS elemental analyzer (Flash EA 1112 NCS, Thermos Fisher Scientific, Waltham, MA, USA).
2.6. Statistical Analyses
The results of the response variables were subjected to the Shapiro–Wilk normality test (p ≤ 0.05). Then, a 3-way ANOVA (F test, p ≤ 0.05) was performed for the factors of soil texture, moisture, and DMPP addition. Finally, the response variables with significant effects were submitted to the Tukey test (p ≤ 0.05) using the Statistix 10 software [23].
3. Results
3.1. Maize Shoot Biomass Production
Drought stress reduced maize shoot biomass yield by 38% and 65% in sand and silt loam soil textures, respectively, compared with sufficient moisture treatment (Figure 1). In addition, applying DMPP did not change maize biomass production in the investigated soil textures under sufficient moisture and in the sand soil under drought. In contrast, DMPP application compared with the without DMPP treatment increased shoot biomass production by 60% in the silt loam soil under drought.
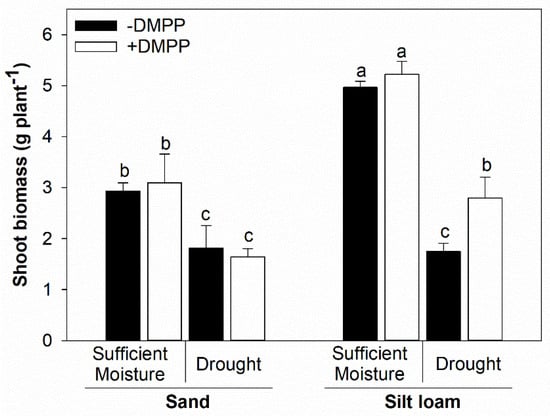
Figure 1.
Shoot biomass of maize plants grown in sand and silt loam soil textures under sufficient moisture (60% WHC) and drought (30% WHC) and fertilized with ammonium sulfate without and with DMPP addition (−DMPP, +DMPP). Vertical bars represent the mean ± standard deviation (n = 4). Different letters represent significant differences between means according to the Tukey test (p ≤ 0.05).
3.2. Soil pH and Mn2+ Availability
The moisture contents in both soils did not affect the rhizosphere soil pH of both soil textures (Figure 2A). Furthermore, a reduction in the rhizosphere soil pH was observed in both soil textures with the addition of DMPP, irrespective of soil moisture. The treatments generally did not affect the bulk soil pH (Figure 2B).
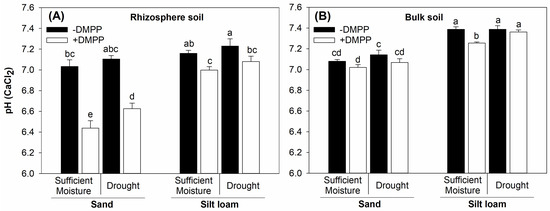
Figure 2.
Rhizosphere (A) and bulk soil (B) pH (CaCl2) of sand and silt loam soil textures under sufficient moisture (60% WHC) and drought (30% WHC), and fertilized with ammonium sulfate without and with DMPP addition (−DMPP, +DMPP). Vertical bars represent the mean ± standard deviation (n = 4). Different letters represent significant differences between means according to the Tukey test (p ≤ 0.05).
Further, the rhizosphere soil of both textures indicated an 8- to 38-fold Mn2+ greater availability compared to bulk soil (Figure 3A,B). Under silt loam, Mn2+ in the rhizosphere was higher relative to sand, though it was not affected by soil moisture and DMPP addition. Contrarily, DMPP increased Mn2+ availability in the rhizosphere soil of sand irrespective of the soil moisture contents. There were no effects of DMPP and moisture content on Mn2+ in the bulk soil of both textures.
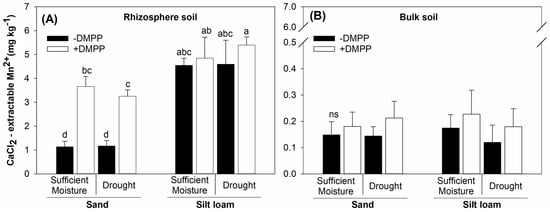
Figure 3.
Rhizosphere (A) and bulk (B) soil CaCl2–extractable Mn2+ in sand and silt loam soil textures under sufficient moisture (60% WHC) and drought (30% WHC), and fertilized with ammonium sulfate (AS) without and with DMPP addition (−DMPP, +DMPP). Vertical bars represent the mean ± standard deviation (n = 4). Different letters represent significant differences between means according to the Tukey test (p ≤ 0.05). ns represents a non-significant effect according to the analysis of variance (p ≤ 0.05).
3.3. Maize Shoot Concentrations of Macro- and Micronutrients
Maize shoot concentration of Mn2+ decreased from sand to silt loam soil (Figure 4B). Notably, under sand, DMPP led to increased Mn2+ shoot concentration in sufficient soil moisture content, but not under drought. In addition, DMPP increased Mn2+ shoot concentration by up to 38% under drought in the silt loam soil texture.
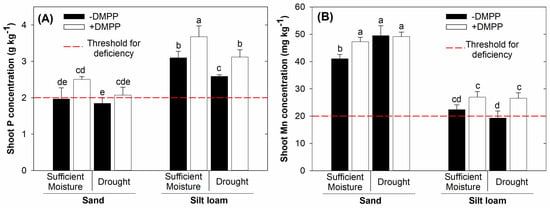
Figure 4.
Shoot P (A) and Mn2+ (B) concentrations in maize plants grown on sand and silt loam soil textures under sufficient moisture (60% WHC) and drought (30% WHC), and fertilized with ammonium sulfate without and with DMPP addition (−DMPP, +DMPP). Vertical bars represent the mean ± standard deviation (n = 4). Different letters represent significant differences between means according to the Tukey test (p ≤ 0.05).
In contrast to Mn2+, P shoot concentration increased from sand to silt loam soils (Figure 4A). Under silt loam, sufficient soil moisture increased P shoot concentration relative to drought. Further, in the silt loam texture, DMPP promoted P shoot concentration by up to 21% and 50% in drought stress and sufficient moisture, respectively. Contrastingly, under sand soil, P shoot concentration was not affected by soil moisture and DMPP.
According to the results presented in Table 2, in general, DMPP supply did not impact K+, Mg2+, Fe2+, Zn2+, and Cu2+ shoot concentration. Moreover, DMPP decreased Ca2+ in plant tissue under both soil textures, while it increased S shoot concentration in sand texture.
Table 2.
Macro- and micronutrient concentrations in shoot tissue of maize plants grown on sand and silt loam soil textures under sufficient moisture (60% WHC) and drought (30% WHC), fertilized with ammonium sulfate (AS) without and with DMPP addition (−DMPP, +DMPP).
Shoot N concentration was not affected by moisture content, whereas DMPP addition increased shoot N concentration under drought soil conditions in the sand soil (Figure 5). Additionally, there were no effects from moisture contents and DMPP treatments on shoot N concentration in silt loam soil. Furthermore, soil texture did not impact the observed results.
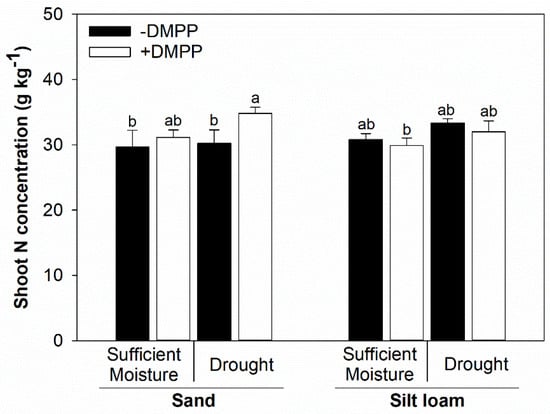
Figure 5.
Shoot total N concentration in maize plants grown on sand and silt loam soil textures under sufficient moisture (60% WHC) and drought (30% WHC), and fertilized with ammonium sulfate without and with DMPP addition (−DMPP, +DMPP). Vertical bars represent the mean ± standard deviation (n = 4). Different letters represent significant differences between means according to the Tukey test (p ≤ 0.05).
3.4. Soil Concentrations of NH4+-N and NO3−-N
Nitrification was higher in silt loam compared with sand soil texture, irrespective of the moisture contents (Figure 6). DMPP inhibited nitrification in sand soil regardless of the moisture content. Under silt loam, DMPP suppressed nitrification, especially during the first two weeks, but NO3−-N accumulated in the soil after this period despite DMPP application.
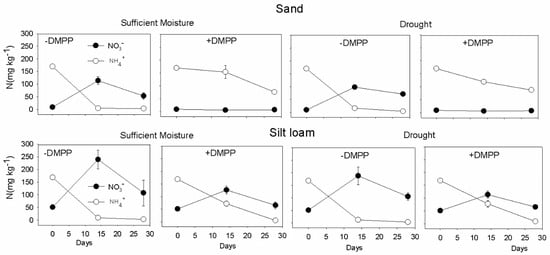
Figure 6.
Soil NH4+-N and NO3−-N concentrations in sand and silt loam soil textures under sufficient moisture (60% WHC) and drought (30% WHC), and fertilized with ammonium sulfate without and with DMPP addition (−DMPP, +DMPP). Vertical bars represent the mean ± standard deviation (n = 4).
4. Discussion
Mn2+ and P deficiencies are found in soils with high pH and organic matter content in Northern Germany, the Netherlands, and Denmark [1,2]. In addition, climate change is likely to increase the frequency and duration of drought events, which can further decrease nutrient uptake via mass flow and diffusion, aggravating Mn2+ and P deficiencies [16,24]), and promoting crop yield losses [25]. Moreover, soil drying favors Mn2+ oxidation, which can worsen Mn2+ deficiency. In this context, the effectiveness of applying NIs to alleviate Mn2+ and P deficiencies in soils with higher pH has been investigated under sufficient moisture conditions [5,6], but studies under drought are lacking. Similarly, the impact of soil texture on the effectiveness of NIs in alleviating Mn2+ and P deficiencies under drought is poorly understood. The present study indicated that DMPP can alleviate Mn2+ and P deficiencies under drought; however, this effect was soil texture-dependent, and the likely underlying mechanisms are discussed below.
4.1. DMPP-Induced Rhizosphere Acidification Alleviates P and Mn2+ Deprivation in Silt Loam Soil with Higher pH
Results of this study show that DMPP-stabilized NH4+-N increased shoot biomass production by 60% in silt loam under drought stress conditions, but not in sand soil texture (Figure 3B). DMPP-stabilized NH4+-N in this context alleviated the effects of moisture deprivation on plant growth in silt loam texture. Such results probably can be explained by increased Mn2+ availability and uptake caused by DMPP (Figure 4B), given the role of Mn2+ in water molecule-splitting in the light phase of photosynthesis [26,27]. In that respect, DMPP in silt loam soil under drought rescued maize from Mn2+ deficiency, increasing its shoot concentration by up to 38%. Noteworthy, the average Mn2+ shoot concentration was very close to or below the threshold for deficiency (Figure 4B) [28], suggesting that DMPP played a pivotal role in preventing Mn2+ deficiency in this soil texture. Moreover, higher P shoot concentrations in the DMPP treatment relative to those without DMPP (Figure 4A) may also have contributed to the aforementioned effect.
In addition to Mn2+ availability, DMPP-stabilized NH4+-N improved P availability and increased its shoot concentration by 21% in the silt loam soil under drought. This probably occurred due to the rhizosphere acidification resulting from NH4+ uptake-proton release into the rhizosphere. These findings confirmed previous studies showing that applying NIs is beneficial for P availability and uptake [10,11,13,29]. This is because P availability is favored in slightly acidic soils with a pH around 6.5 [30]. At pH values higher than 6.5, P availability decreases due to precipitation of phosphate with exchangeable Ca2+. Moreover, pH values below 6.5 stimulated phosphate binding to Al hydroxides or precipitation with exchangeable Al3+ [30].
4.2. Soil Texture Impacts the Effectiveness of DMPP in Alleviating P and Mn2+ Deficiencies
The positive effects of DMPP on alleviating maize growth reduction due to Mn2+ and P deprivation under drought were found in the silt loam and not in the sand soil texture. Thus, the data indicated that the impact of DMPP-induced rhizosphere acidification is strongly dependent on soil texture. This might be explained by the fact that drought in sandy soils impairs nutrient mass flow and diffusion more severely due to their lower WHC relative to fine-textured soils. In the present study, the WHC from sand and silt loam soil textures were 0.08 g g−1 and 0.10 g g−1, respectively. Therefore, the rhizosphere acidification caused by DMPP probably has negligible impacts on maize P uptake under drought conditions in sandy soils. For instance, previous studies indicated that soils with water contents of 0.4 g cm−3 and 0.13 g cm−3 needed 10 μM P and 200 μM P, respectively, to reach the same P uptake by maize plants [20,21].
Interestingly, DMPP showed lower effectiveness in suppressing nitrification and in inducing rhizosphere acidification in the silt loam relative to the sand soil texture. For instance, DMPP caused a significant decrease in the rhizosphere pH of both soils; however, it led to a sharper reduction in the sand (−0.48 units) compared to the silt loam texture (−0.15 units). Such results can be explained by the lower buffering capacity of coarse-textured relative to fine-textured soils [31]. In addition, the silt loam soil had a higher NO3−-N concentration (51 mg kg−1) than the sand soil (9 mg kg−1) at the beginning of the experiment, which may have attenuated DMPP effects on rhizosphere acidification of the fine-textured soil. Finally, the effectiveness of DMPP in inhibiting NO3−-N accumulation was lower in the silt loam relative to the sand soil (Figure 6). This likely occurred because fine-textured soils may promote DMPP adsorption to negatively charged soil colloids [12] and enhance its biodegradation, especially under warm temperatures [32].
5. Conclusions
This study supported the hypothesis that DMPP alleviates Mn2+ and P deficiencies in maize plants cultivated on soils with higher pH under drought. However, this effect was soil texture-dependent and observed in fine-textured rather than coarse-textured soil. Studies investigating the effectiveness of DMPP on alleviating Mn2+ and P deficiencies in high-pH soils were lacking; therefore, our data provide new insights into the potential of using nitrification inhibitors to attenuate the negative impacts of more frequent and severe droughts caused by climate change on plant growth. The present study also indicated that the use of NIs to alleviate Mn2+ and P deficiencies should be site-specific, as their effectiveness depends on both soil and environmental factors. Consequently, multiyear and multisite studies will be necessary to developing site-specific recommendations. Finally, future research should focus on NI-driven alleviation of Mn2+ and P deficiencies in high-pH soils with different drought severity and duration. Moreover, experiments that consider the whole crop life cycle and drought occurrence at different timings, including plant vegetative and reproductive stages, are also needed.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/nitrogen7020034/s1. Figure S1: Phenotype of maize plants grown in sand and silt loam soil textures under sufficient moisture (60% WHC) and drought (30% WHC), fertilized with ammonium sulfate without and with DMPP addition (−DMPP, +DMPP).; Table S1: Macro- and micronutrients applied to the soil (g kg soil−1).
Author Contributions
M.E.E.: Funding acquisition; conceptualization; methodology; investigation; visualization; and writing—original draft. P.L.R.: Supervision; visualization; writing—review and editing. B.P.: Supervision; writing—review and editing. K.H.M.: Supervision; funding acquisition; resources; project administration; writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Katholischer Akademischer Ausländer-Dienst, grant number [MEE Doctoral scholarship 2021–2024].
Data Availability Statement
The datasets generated during and/or analyzed during the current study are available from the corresponding authors on reasonable request.
Acknowledgments
ME Etabo thanks the doctoral scholarship provided by Katholischer Akademischer Ausländer-Dienst.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Husted, S.; Thomsen, M.U.; Mattsson, M.; Schjoerring, J.K. Influence of nitrogen and sulphur form on manganese acquisition by barley (shape Hordeum vulgare). Plant Soil 2005, 268, 309–317. [Google Scholar] [CrossRef]
- Mulder, E.G.; Gerretsen, F.C. Soil manganese in relation to plant growth. In Advances in Agronomy; Elsevier: Amsterdam, The Netherlands, 1952; Volume 4, pp. 221–277. [Google Scholar] [CrossRef]
- Grillakis, M.G. Increase in severe and extreme soil moisture droughts for Europe under climate change. Sci. Total Environ. 2019, 660, 1245–1255. [Google Scholar] [CrossRef]
- Brouder, S.M.; Volenec, J.J. Nutrition of plants in a changing climate. In Marschner’s Mineral Nutrition of Plants; Elsevier: Amsterdam, The Netherlands, 2023; pp. 723–750. [Google Scholar] [CrossRef]
- Dreyer, M.; Wichmann, M.; Rischen, M.; Görlach, B.M.; Ehmke, A.; Pitann, B.; Mühling, K.H. Ammonium-driven nitrification plays a key role in increasing Mn availability in calcareous soils. J. Plant Nutr. Soil Sci. 2020, 183, 389–396. [Google Scholar] [CrossRef]
- Etabo, M.E.; Ribeiro, P.L.; Pitann, B.; Mühling, K.H. Manganese and phosphorus maize shoot concentrations are differently affected by nitrification inhibitor-driven rhizosphere acidification. Soil Use Manag. 2024, 40, e13054. [Google Scholar] [CrossRef]
- Graham, R.D.; Hannam, R.J.; Uren, N.C. (Eds.) Manganese in Soils and Plants; Springer: Dordrecht, The Netherlands, 1988. [Google Scholar] [CrossRef]
- Marschner, H. Marschner’s Mineral Nutrition of Higher Plants; Academic Press: Cambridge, MA, USA, 2012. [Google Scholar] [CrossRef]
- Pedersen, I.F.; Sørensen, P.; Rasmussen, J.; Withers, P.J.A.; Rubæk, G.H. Fertilizer ammonium: Nitrate ratios determine phosphorus uptake by young maize plants. J. Plant Nutr. Soil Sci. 2019, 182, 541–551. [Google Scholar] [CrossRef]
- Thomson, C.J.; Marschner, H.; Römheld, V. Effect of nitrogen fertilizer form on pH of the bulk soil and rhizosphere, and on the growth, phosphorus, and micronutrient uptake of bean. J. Plant Nutr. 1993, 16, 493–506. [Google Scholar] [CrossRef]
- Vogel, C.; Sekine, R.; Huang, J.; Steckenmesser, D.; Steffens, D.; Huthwelker, T.; Borca, C.N.; Pradas Del Real, A.E.; Castillo-Michel, H.; Adam, C. Effects of a nitrification inhibitor on nitrogen species in the soil and the yield and phosphorus uptake of maize. Sci. Total Environ. 2020, 715, 136895. [Google Scholar] [CrossRef]
- Barth, G.; Von Tucher, S.; Schmidhalter, U.; Otto, R.; Motavalli, P.; Ferraz-Almeida, R.; Meinl Schmiedt Sattolo, T.; Cantarella, H.; Vitti, G.C. Performance of nitrification inhibitors with different nitrogen fertilizers and soil textures. J. Plant Nutr. Soil Sci. 2019, 182, 694–700. [Google Scholar] [CrossRef]
- Li, S.; Liu, Y.; Sha, Z.; Li, S.; Yang, Q. Adding nitrification inhibitors to N fertilisers induces rhizosphere acidification and enhances P acquisition: A meta-analysis. Eur. J. Agron. 2023, 151, 126967. [Google Scholar] [CrossRef]
- Coleto, I.; Marín-Peña, A.J.; Urbano-Gámez, J.A.; González-Hernández, A.I.; Shi, W.; Li, G.; Marino, D. Interaction of ammonium nutrition with essential mineral cations. J. Exp. Bot. 2023, 74, 6131–6144. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Razavi, B.S. Rhizosphere size and shape: Temporal dynamics and spatial stationarity. Soil Biol. Biochem. 2019, 135, 343–360. [Google Scholar] [CrossRef]
- Li, S.; Yang, L.; Huang, X.; Zou, Z.; Zhang, M.; Guo, W.; Addo-Danso, S.D.; Zhou, L. Mineral nutrient uptake, accumulation, and distribution in Cunninghamia lanceolata in response to drought stress. Plants 2023, 12, 2140. [Google Scholar] [CrossRef]
- Suriyagoda, L.D.B.; Ryan, M.H.; Renton, M.; Lambers, H. Plant responses to limited moisture and phosphorus availability. In Advances in Agronomy; Elsevier: Amsterdam, The Netherlands, 2014; Volume 124, pp. 143–200. [Google Scholar] [CrossRef]
- Alejandro, S.; Höller, S.; Meier, B.; Peiter, E. Manganese in plants: From acquisition to subcellular allocation. Front. Plant Sci. 2020, 11, 300. [Google Scholar] [CrossRef] [PubMed]
- Wood, T.E.; Silver, W.L. Strong spatial variability in trace gas dynamics following experimental drought in a humid tropical forest. Glob. Biogeochem. Cycles 2012, 26, GB3005. [Google Scholar] [CrossRef]
- Cox, M.S.; Barber, S.A. Soil phosphorus levels needed for equal P uptake from four soils with different water contents at the same water potential. Plant Soil 1992, 143, 93–98. [Google Scholar] [CrossRef]
- Marschner, P.; Rengel, Z. Nutrient availability in soils. In Marschner’s Mineral Nutrition of Plants; Elsevier: Amsterdam, The Netherlands, 2023; pp. 499–522. [Google Scholar] [CrossRef]
- Jezek, M.; Geilfus, C.M.; Bayer, A.; Mühling, K.H. Photosynthetic capacity, nutrient status, and growth of maize (Zea mays L.) upon MgSO4 leaf-application. Front. Plant Sci. 2015, 5, 781. [Google Scholar] [CrossRef]
- Statistix®. Statistix 10 Analytical Software; Statistix: Tallahassee, FL, USA, 2013. [Google Scholar]
- Smith, M.R.; Dinglasan, E.; Veneklaas, E.; Polania, J.; Rao, I.M.; Beebe, S.E.; Merchant, A. Effect of drought and low P on yield and nutritional content in common bean. Front. Plant Sci. 2022, 13, 814325. [Google Scholar] [CrossRef]
- Homyak, P.M.; Allison, S.D.; Huxman, T.E.; Goulden, M.L.; Treseder, K.K. Effects of drought manipulation on soil nitrogen cycling: A meta-analysis. JGR Biogeosci. 2017, 122, 3260–3272. [Google Scholar] [CrossRef]
- Barber, J. Towards a full understanding of water splitting in photosynthesis. Int. J. Photoenergy 2004, 6, 43–51. [Google Scholar] [CrossRef]
- Greife, P.; Schönborn, M.; Capone, M.; Assunção, R.; Narzi, D.; Guidoni, L.; Dau, H. The electron–proton bottleneck of photosynthetic oxygen evolution. Nature 2023, 617, 623–628. [Google Scholar] [CrossRef] [PubMed]
- Broadley, M.; Brown, P.; Cakmak, I.; Rengel, Z.; Zhao, F. Function of nutrients. In Marschner’s Mineral Nutrition of Higher Plants; Elsevier: Amsterdam, The Netherlands, 2012; pp. 191–248. [Google Scholar] [CrossRef]
- Jing, J.; Zhang, F.; Rengel, Z.; Shen, J. Localized fertilization with P plus N elicits an ammonium-dependent enhancement of maize root growth and nutrient uptake. Field Crops Res. 2012, 133, 176–185. [Google Scholar] [CrossRef]
- Penn, C.J.; Camberato, J.J. A critical review on soil chemical processes that control how soil pH affects phosphorus availability to plants. Agriculture 2019, 9, 120. [Google Scholar] [CrossRef]
- Bordoloi, R.; Das, B.; Yam, G.; Pandey, P.K.; Tripathi, O.P. Modeling of water holding capacity using readily available soil characteristics. Agric. Res. 2019, 8, 347–355. [Google Scholar] [CrossRef]
- Ribeiro, P.L.; Singh, A.; Sagervanshi, A.; Naeem, A.; Mühling, K.H. High soil moisture rather than drying-rewetting cycles reduces the effectiveness of nitrification inhibitors in mitigating N2O emissions. Biol Fertil Soils 2024, 60, 627–638. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.