

Host Habitat as a Dominant Role in Shaping the Gut Microbiota of Wild Crucian Carp (Carassius auratus)

Abstract
1. Introduction
2. Materials and Methods
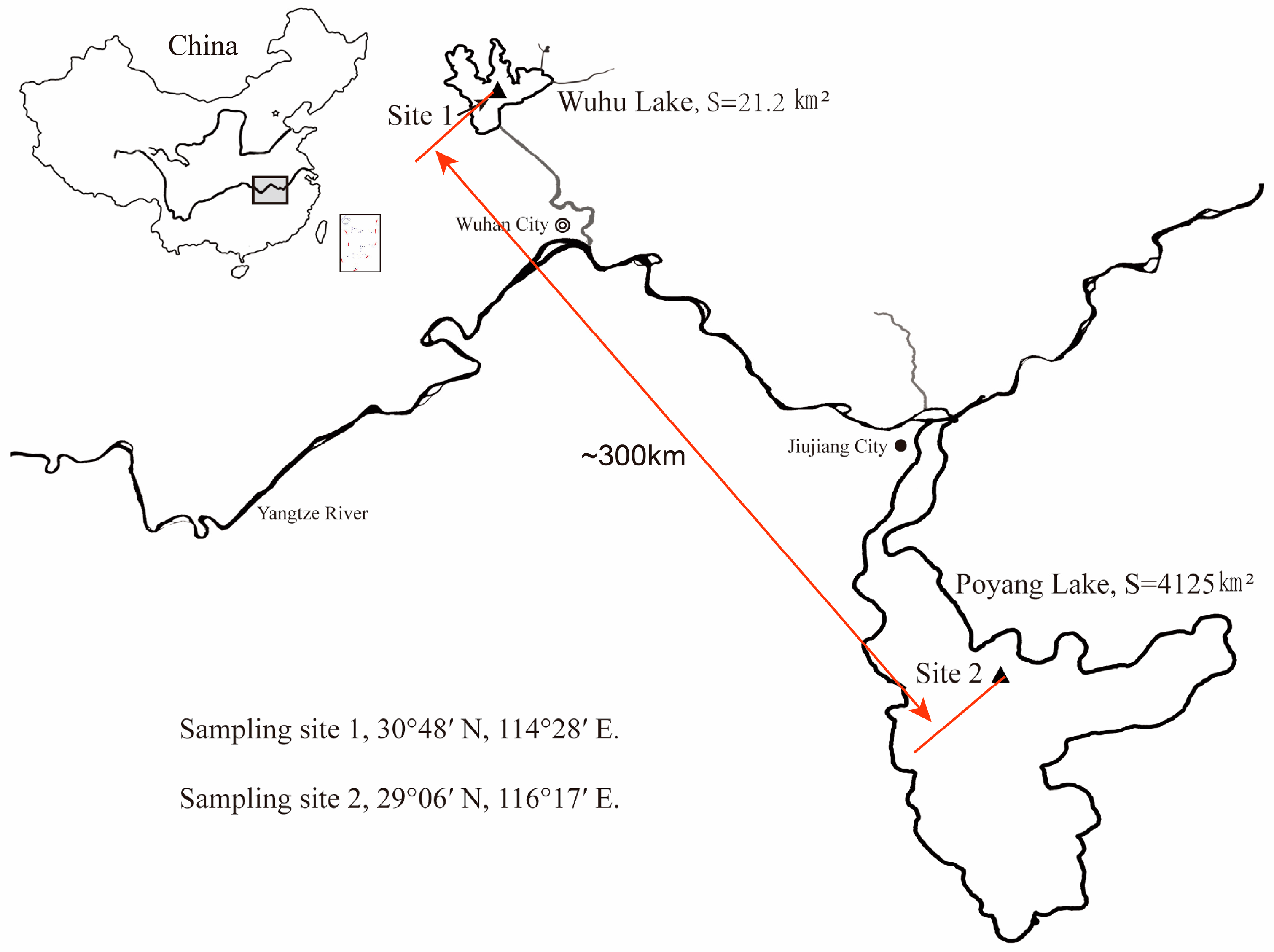
2.1. Sample Collection
2.2. DNA Isolation
2.3. PCR and Sequencing Methods
2.4. Statistical Analysis
3. Results
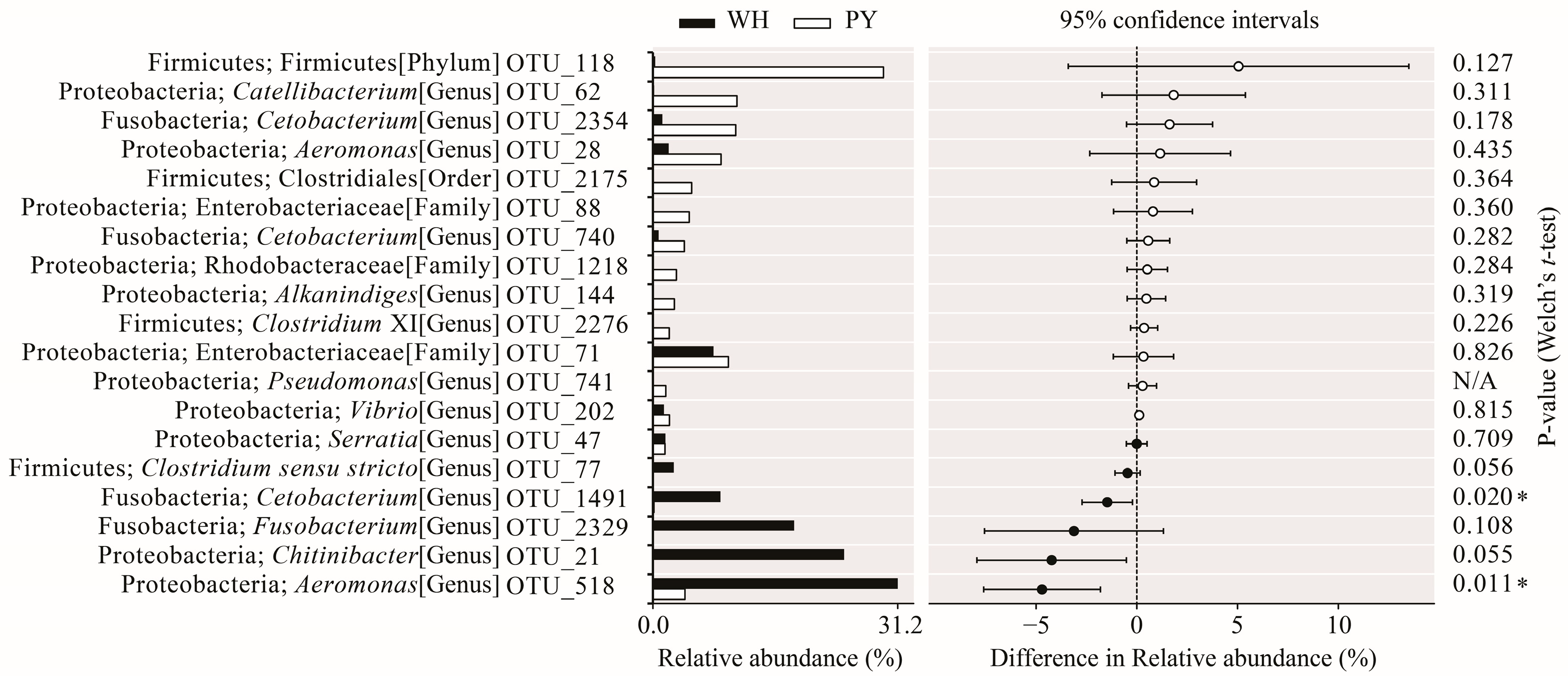
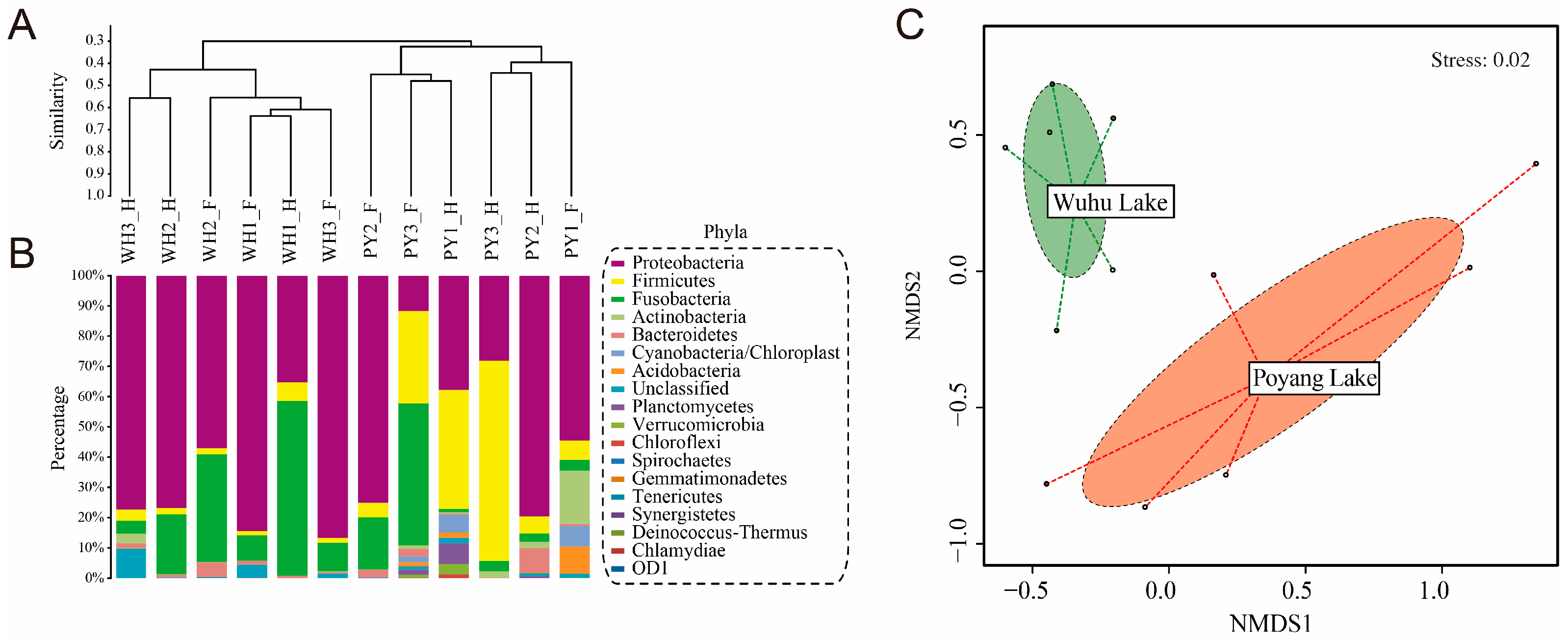
3.1. Characterization of the Gut Microbiota
3.2. Microbial Diversity Varied with Host Habitats
3.3. Microbial Community Dissimilarity between Gut Content Samples
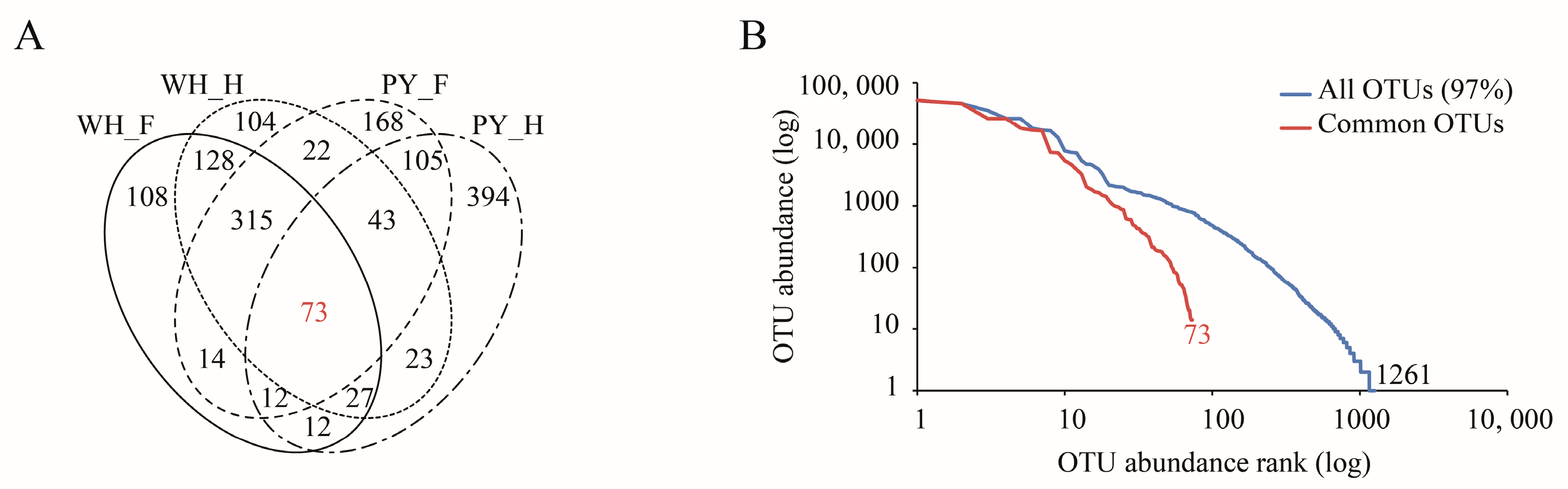
3.4. Identification of a Common Gut Microbiota
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ding, S.L.; Chi, M.M.; Scull, B.P.; Rigby, R.; Schwerbrock, N.M.J.; Magness, S.; Jobin, C.; Lund, P.K. High-Fat Diet: Bacteria Interactions Promote Intestinal Inflammation Which Precedes and Correlates with Obesity and Insulin Resistance in Mouse. PLoS ONE 2010, 5, e12191. [Google Scholar] [CrossRef] [PubMed]
- Chang, X.L.; Shen, Y.H.; Yun, L.L.; Wang, X.F.; Feng, J.C.; Yang, G.K.; Meng, X.L.; Zhang, J.X.; Su, X. The antipsychotic drug olanzapine altered lipid metabolism in the common carp (Cyprinus carpio L.): Insight from the gut microbiota-SCFAs-liver axis. Sci. Total Environ. 2023, 856, 159054. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.H.; Lin, G.; Fu, G.H.; Wan, Z.Y.; Lee, M.; Wang, L.; Liu, X.J.; Yue, G.H. The intestinal microbiome of fish under starvation. BMC Genom. 2014, 15, 266. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Zhang, T.; Ren, L.; Fang, D.A.; Xu, D.P. Differential Study of Microbiota in the Gill and Intestine of Silver Carp (Hypophthalmichthys molitrix) from the Algae-Dominated and Hydrophyte-Dominated Areas of Taihu Lake, China. Fishes 2022, 7, 304. [Google Scholar] [CrossRef]
- Xu, Q.; Yang, Z.; Chen, S.Y.; Zhu, W.J.; Xiao, S.Y.; Liu, J.; Wang, H.Q.; Lan, S.L. Effects of Replacing Dietary Fish Meal by Soybean Meal Co-Fermented Using Bacillus subtilis and Enterococcus faecium on Serum Antioxidant Indices and Gut Microbiota of Crucian Carp Carassius auratus. Fishes 2022, 7, 54. [Google Scholar] [CrossRef]
- Romero, J.; Catalan, N.; Ramirez, C.; Miranda, C.D.; Oliva, M.; Flores, H.; Romero, M.S.; Rojas, R. High Abundance of Candidatus Arthromitus in Intestinal Microbiota of Seriolella violacea (Palm Ruff) under Reared Conditions. Fishes 2023, 8, 109. [Google Scholar] [CrossRef]
- Raicu, P.; Taisescu, E.; Bãnãrescu, P. Carassius carassius and C. auratus, a pair of diploid and tetraploid representative species (Pisces, Cyprinidae). Cytologia 1981, 46, 233–240. [Google Scholar] [CrossRef]
- Clements, K.D.; Angert, E.R.; Montgomery, W.L.; Choat, J.H. Intestinal microbiota in fishes: What’s known and what’s not. Mol. Ecol. 2014, 23, 8. [Google Scholar] [CrossRef]
- Li, T.T.; Li, H.; Gatesoupe, F.J.; She, R.; Lin, Q.; Yan, X.F.; Li, J.B.; Li, X.Z. Bacterial Signatures of “Red-Operculum” Disease in the Gut of Crucian Carp (Carassius auratus). Microb. Ecol. 2017, 74, 510–521. [Google Scholar] [CrossRef]
- Yan, B.; Han, J.; Sun, Y.M.; Lei, L.; Yuan, J.; Qiao, Z.X.; Men, J.; Wang, X.; Guo, Y.Y.; Wang, Q.D.; et al. Probiotics ameliorate growth retardation of glyphosate by regulating intestinal microbiota and metabolites in crucian carp (Carassius auratus). Sci. Total Environ. 2022, 851, 158260. [Google Scholar] [CrossRef]
- Liu, H.D.; Liu, M.; Wang, B.J.; Jiang, K.Y.; Jiang, S.; Sun, S.J.; Wang, L. PCR-DGGE analysis of intestinal bacteria and effect of Bacillus spp. on intestinal microbial diversity in kuruma shrimp (Marsupenaeus japonicus). Chin. J. Oceanol. Limn. 2010, 28, 808–814. [Google Scholar] [CrossRef]
- Ni, J.J.; Yu, Y.H.; Zhang, T.L.; Gao, L. Comparison of intestinal bacterial communities in grass carp, Ctenopharyngodon idellus, from two different habitats. Chin. J. Oceanol. Limn. 2012, 30, 757–765. [Google Scholar] [CrossRef]
- Wu, S.G.; Tian, J.Y.; Gatesoupe, F.J.; Li, W.X.; Zou, H.; Yang, B.J.; Wang, G.T. Intestinal microbiota of gibel carp (Carassius auratus gibelio) and its origin as revealed by 454 pyrosequencing. World J. Microbiol. Biotechnol. 2013, 29, 1585–1595. [Google Scholar] [CrossRef]
- Faro, A.; Boj, S.F.; Clevers, H. Fishing for Intestinal Cancer Models: Unraveling Gastrointestinal Homeostasis and Tumorigenesis in Zebrafish. Zebrafish 2009, 6, 361–376. [Google Scholar] [CrossRef] [PubMed]
- Yan, Q.Y.; Van der Gast, C.J.; Yu, Y.H. Bacterial Community Assembly and Turnover within the Intestines of Developing Zebrafish. PLoS ONE 2012, 7, e30603. [Google Scholar] [CrossRef] [PubMed]
- Bencina, M.; Jakopic, M.; Friedrich, J. High-molecular-weight genomic DNA isolation from Doratomyces microsporus and synthesis of a genomic DNA library. Acta Chim. Slov. 2007, 54, 893–899. [Google Scholar]
- Walters, W.A.; Caporaso, J.G.; Lauber, C.L.; Berg-Lyons, D.; Fierer, N.; Knight, R. PrimerProspector: De novo design and taxonomic analysis of barcoded polymerase chain reaction primers. Bioinformatics 2011, 27, 1159–1161. [Google Scholar] [CrossRef]
- Zhou, J.Z.; Wu, L.Y.; Deng, Y.; Zhi, X.Y.; Jiang, Y.H.; Tu, Q.C.; Xie, J.P.; Van Nostrand, J.D.; He, Z.L.; Yang, Y.F. Reproducibility and quantitation of amplicon sequencing-based detection. ISME J. 2011, 5, 1303–1313. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microb. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Faith, D.P.; Minchin, P.R.; Belbin, L. Compositional Dissimilarity as a Robust Measure of Ecological Distance. Vegetatio 1987, 69, 57–68. [Google Scholar] [CrossRef]
- Warton, D.I.; Wright, S.T.; Wang, Y. Distance-based multivariate analyses confound location and dispersion effects. Methods Ecol. Evol. 2012, 3, 89–101. [Google Scholar] [CrossRef]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Welch, B.L. The Generalization of ‘Student’s’ Problem when Several Different Population Variances are Involved. Biometrika 1947, 34, 28–35. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2019. [Google Scholar]
- Bjork, S.J.; Bartholomew, J.L. Invasion of Ceratomyxa shasta (Myxozoa) and comparison of migration to the intestine between susceptible and resistant fish hosts. Int. J. Parasitol. 2010, 40, 1087–1095. [Google Scholar] [CrossRef] [PubMed]
- Gomez, G.D.; Balcazar, J.L. A review on the interactions between gut microbiota and innate immunity of fish. FEMS Immunol. Med. Microbiol. 2008, 52, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Hill, M.J. Diet and the Human Intestinal Bacterial-Flora. Cancer Res. 1981, 41, 3778–3780. [Google Scholar]
- Ni, J.J.; Yan, Q.Y.; Yu, Y.H.; Zhang, T.L. Fish gut microecosystem: A model for detecting spatial pattern of microorganisms. Chin. J. Oceanol. Limn. 2014, 32, 54–57. [Google Scholar] [CrossRef]
- Li, T.T.; Qi, M.T.; Gatesoupe, F.J.; Tian, D.C.; Jin, W.H.; Li, J.; Lin, Q.; Wu, S.J.; Li, H. Adaptation to Fasting in Crucian Carp (Carassius auratus): Gut Microbiota and Its Correlative Relationship with Immune Function. Microb. Ecol. 2019, 78, 6–19. [Google Scholar] [CrossRef]
- Zou, S.; Gong, L.; Khan, T.A.; Pan, L.; Yan, L.; Li, D.; Cao, L.; Li, Y.; Ding, X.; Yi, G.; et al. Comparative analysis and gut bacterial community assemblages of grass carp and crucian carp in new lineages from the Dongting Lake area. Microbiologyopen 2020, 9, e996. [Google Scholar] [CrossRef]
- Wang, S.-X.; Zhang, J.-Y.; Du, X.-K.; Liu, D.-J.; Liu, L.-X.; Shen, X.-H. Comparative analysis of the intestinal microbiota in goldfish and crucian carps between different aquaponics and traditional farming. Aquac. Rep. 2022, 25, 101240. [Google Scholar] [CrossRef]
- Fu, Y.; Liang, X.; Li, D.; Gao, H.; Wang, Y.; Li, W.; Xu, K.; Hu, F. Effect of Dietary Tryptophan on Growth, Intestinal Microbiota, and Intestinal Gene Expression in an Improved Triploid Crucian Carp. Front. Nutr. 2021, 8, 676035. [Google Scholar] [CrossRef] [PubMed]
- Song, H.K.Y.; Lee, S.; Han, D.W.; Kim, J.H. Characterization of the Gut Microbiota of Mackerel Icefish, Champsocephalus gunnari. Fishes 2023, 8, 13. [Google Scholar] [CrossRef]
- Wu, S.; Gao, T.; Zheng, Y.; Wang, W.; Cheng, Y.; Wang, G. Microbial diversity of intestinal contents and mucus in yellow catfish (Pelteobagrus fulvidraco). Aquaculture 2010, 303, 1–7. [Google Scholar] [CrossRef]
- Li, T.; Long, M.; Gatesoupe, F.J.; Zhang, Q.; Li, A.; Gong, X. Comparative analysis of the intestinal bacterial communities in different species of carp by pyrosequencing. Microb. Ecol. 2015, 69, 25–36. [Google Scholar] [CrossRef]
- Roeselers, G.; Mittge, E.K.; Stephens, W.Z.; Parichy, D.M.; Cavanaugh, C.M.; Guillemin, K.; Rawls, J.F. Evidence for a core gut microbiota in the zebrafish. ISME J. 2011, 5, 1595–1608. [Google Scholar] [CrossRef]
- Shanks, O.C.; Kelty, C.A.; Archibeque, S.; Jenkins, M.; Newton, R.J.; McLellan, S.L.; Huse, S.M.; Sogin, M.L. Community structures of fecal bacteria in cattle from different animal feeding operations. Appl. Environ. Microb. 2011, 77, 2992–3001. [Google Scholar] [CrossRef]
- Claesson, M.J.; O’Sullivan, O.; Wang, Q.; Nikkila, J.; Marchesi, J.R.; Smidt, H.; de Vos, W.M.; Ross, R.P.; O’Toole, P.W. Comparative analysis of pyrosequencing and a phylogenetic microarray for exploring microbial community structures in the human distal intestine. PLoS ONE 2009, 4, e6669. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fish ID | Sampling Lake | Weight/g | Body Length/cm |
|---|---|---|---|
| P1 | Poyang | 167.6 | 14.2 |
| P2 | Poyang | 150.2 | 13.3 |
| P3 | Poyang | 161.5 | 13.8 |
| W1 | Wuhu Lake | 142.0 | 12.2 |
| W2 | Wuhu Lake | 168.2 | 14.4 |
| W3 | Wuhu Lake | 170.3 | 14.8 |
| Fish Habitats | Gut Segment | Total Sequences Passed Quality Check | Number of Taxa (OTU Number) | Shannon | Chao1 |
|---|---|---|---|---|---|
| Wuhu lake | Foregut | 88,401 | 214.3 ± 43.0 | 2.1 ± 0.2 | 285.8 ± 33.0 |
| Wuhu lake | Hindgut | 91,029 | 216.0 ± 58.3 | 2.6 ± 0.5 | 306.1 ± 83.2 |
| Poyang lake | Foregut | 110,828 | 197.3 ± 77.8 | 2.9 ± 1.0 | 267.1 ± 77.1 |
| Poyang lake | Hindgut | 146,294 | 280.3 ± 188.3 | 3.0 ± 0.8 | 318.5 ± 193.3 |
| Jaccard Distance-Based Test | Bray–Curtis Distance-Based Test | ||||
|---|---|---|---|---|---|
| WH vs. PY (Host Habitat) | Foregut vs. Hindgut (Gut Segment) | WH vs. PY (Host Habitat) | Foregut vs. Hindgut (Gut Segment) | ||
| MRPP | Delta | 0.840 | 0.895 | 0.745 | 0.825 |
| P | 0.005 | 0.246 | 0.008 | 0.252 | |
| ANOSIM | R | 0.585 | 0.113 | 0.585 | 0.113 |
| P | 0.007 | 0.152 | 0.002 | 0.151 | |
| PERMANOVA | F | 2.507 | 1.123 | 3.481 | 1.231 |
| P | 0.008 | 0.272 | 0.004 | 0.218 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Huang, X.; Zhao, L.; Cai, W.; Yu, Y.; Zhang, J. Host Habitat as a Dominant Role in Shaping the Gut Microbiota of Wild Crucian Carp (Carassius auratus). Fishes 2023, 8, 369. https://doi.org/10.3390/fishes8070369
Li X, Huang X, Zhao L, Cai W, Yu Y, Zhang J. Host Habitat as a Dominant Role in Shaping the Gut Microbiota of Wild Crucian Carp (Carassius auratus). Fishes. 2023; 8(7):369. https://doi.org/10.3390/fishes8070369
Chicago/Turabian StyleLi, Xinghao, Xueli Huang, Liya Zhao, Wei Cai, Yuhe Yu, and Jin Zhang. 2023. "Host Habitat as a Dominant Role in Shaping the Gut Microbiota of Wild Crucian Carp (Carassius auratus)" Fishes 8, no. 7: 369. https://doi.org/10.3390/fishes8070369
APA StyleLi, X., Huang, X., Zhao, L., Cai, W., Yu, Y., & Zhang, J. (2023). Host Habitat as a Dominant Role in Shaping the Gut Microbiota of Wild Crucian Carp (Carassius auratus). Fishes, 8(7), 369. https://doi.org/10.3390/fishes8070369