

Otolith Morphometric and Shape Distinction of Three Redfin Species under the Genus Decapterus (Teleostei: Carangidae) from Sulu Sea, Philippines

 ,
,  ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
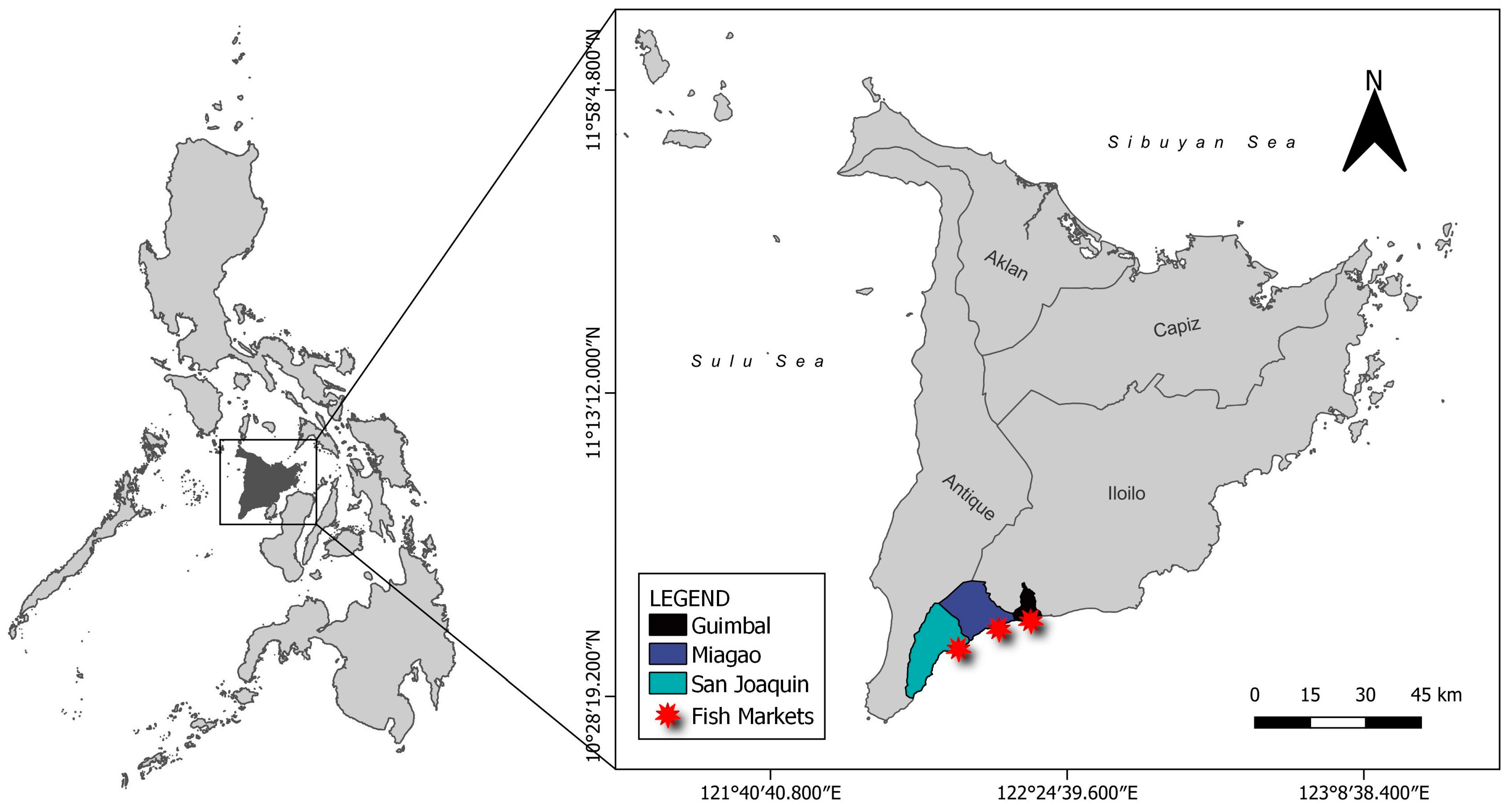
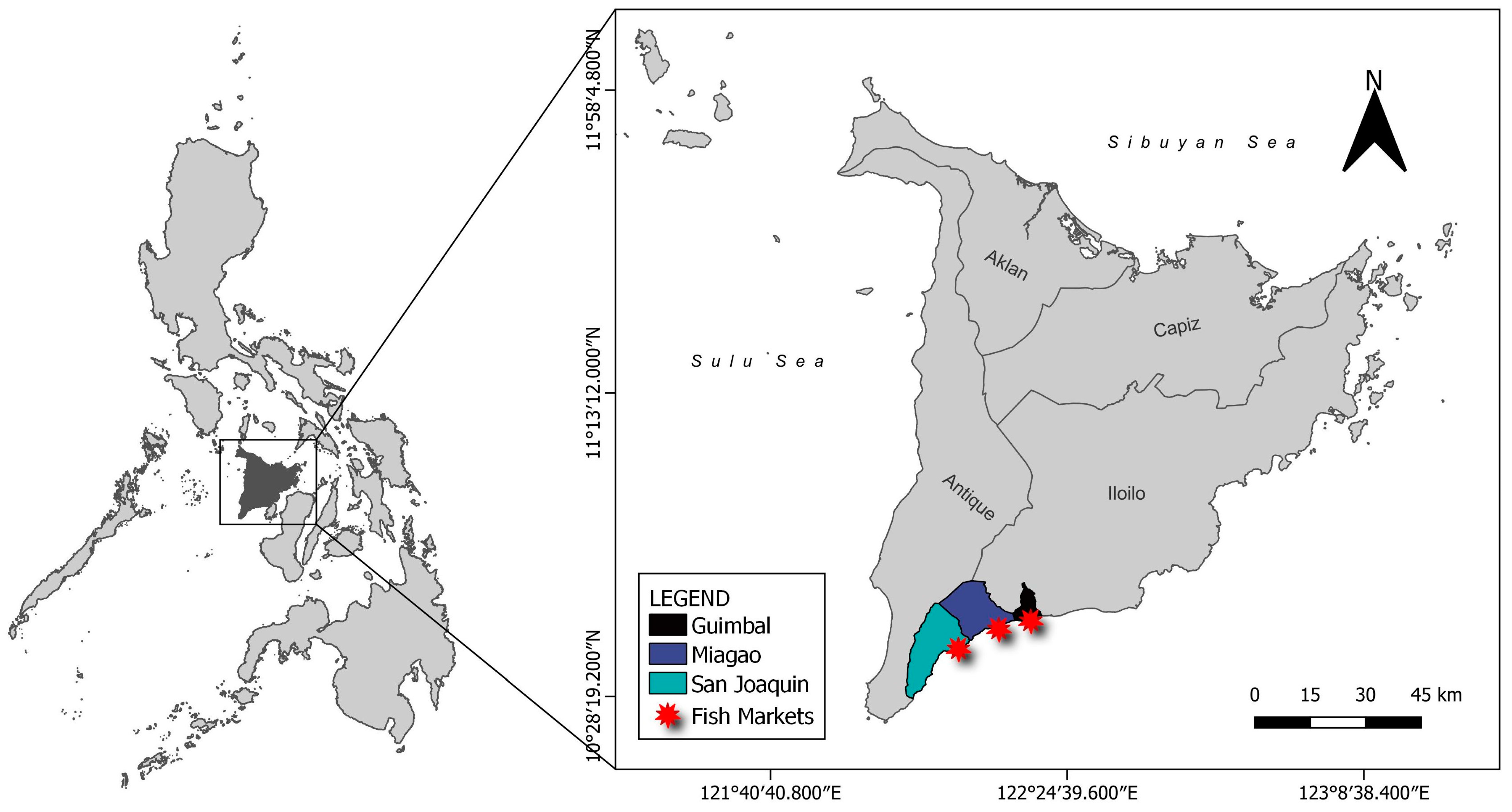
2.1. Sample Collection
2.2. Otolith Extraction and Storage
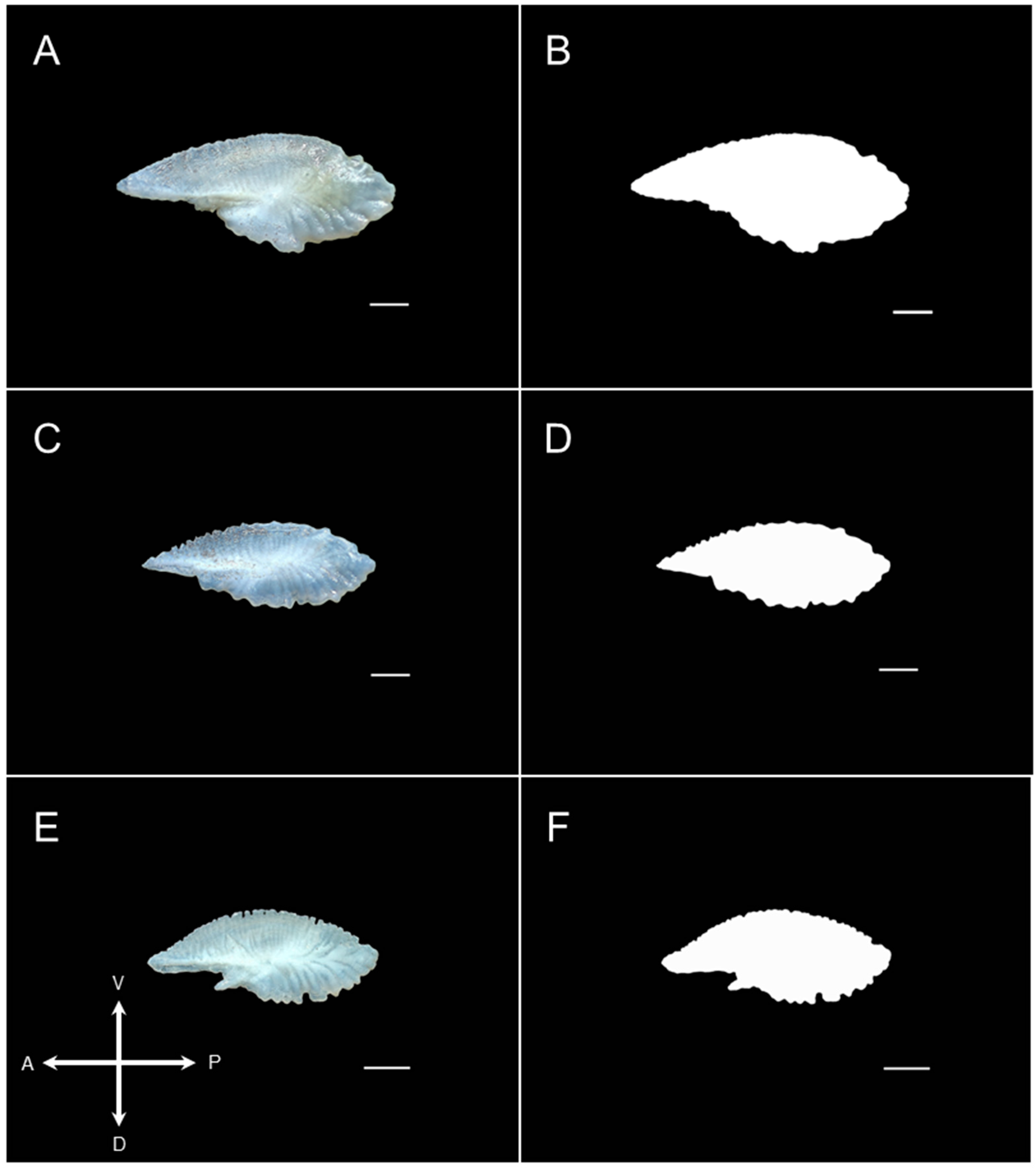
2.3. Sample Imaging
2.4. Otolith Morphometry
2.5. Otolith Shape Analysis
2.6. Data Analyses
3. Results
3.1. Otolith Morphometry
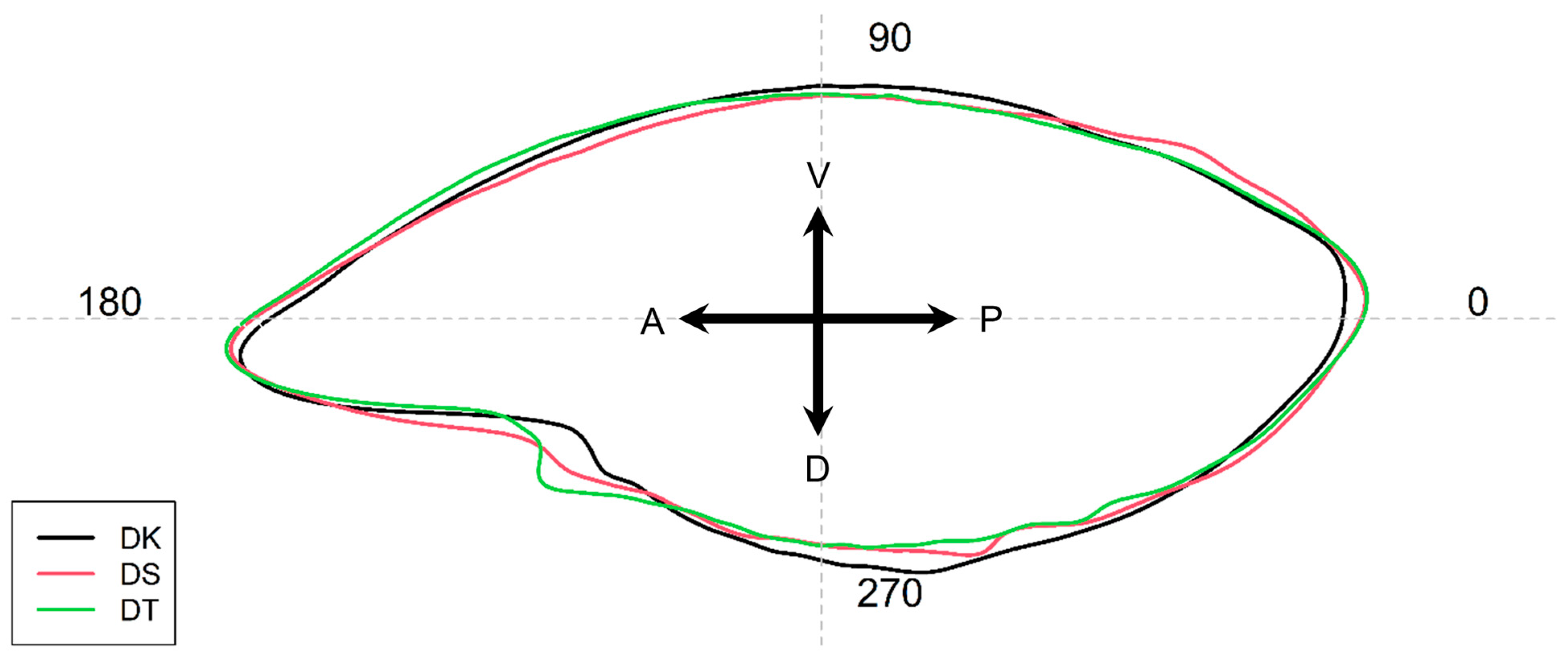
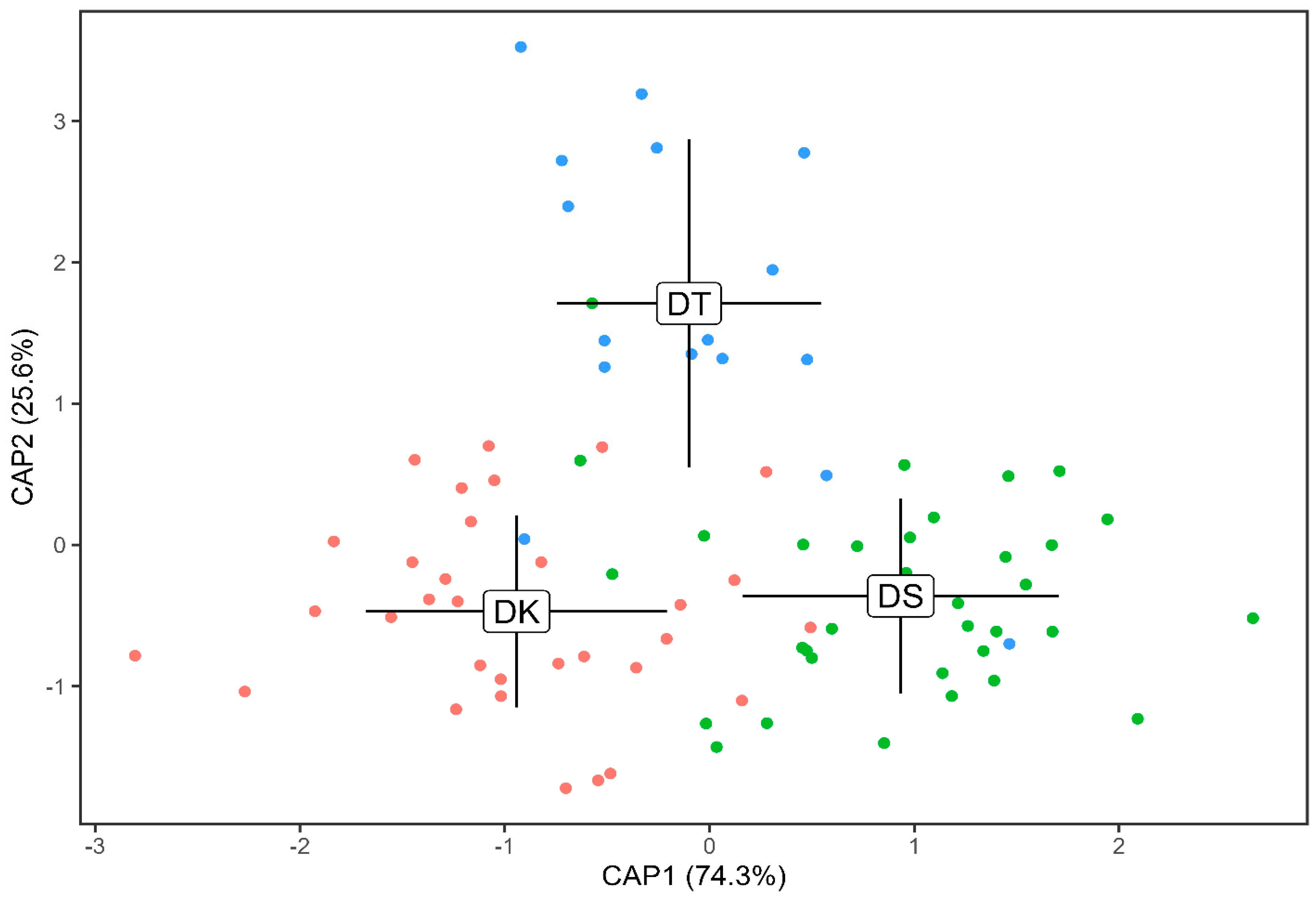
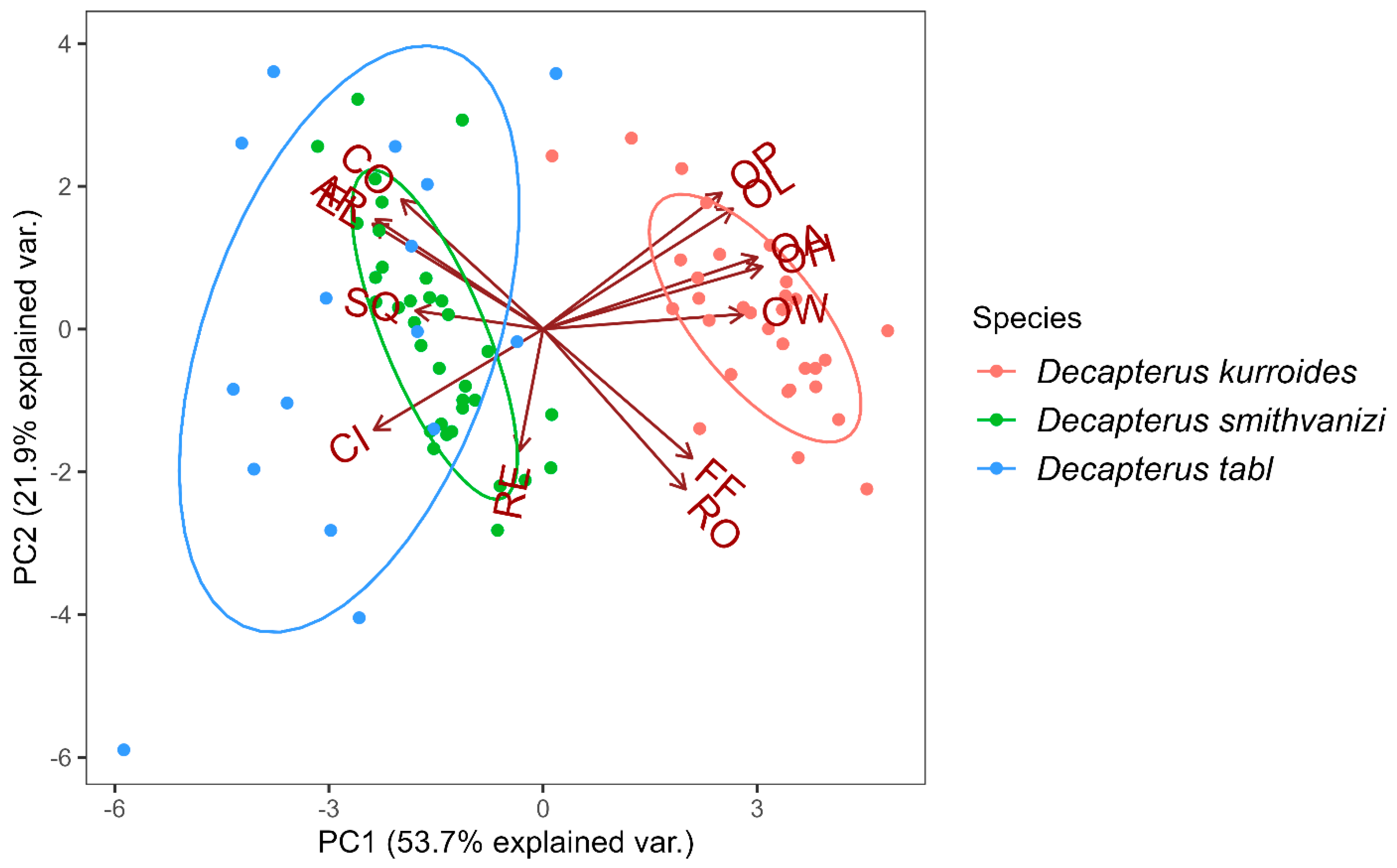
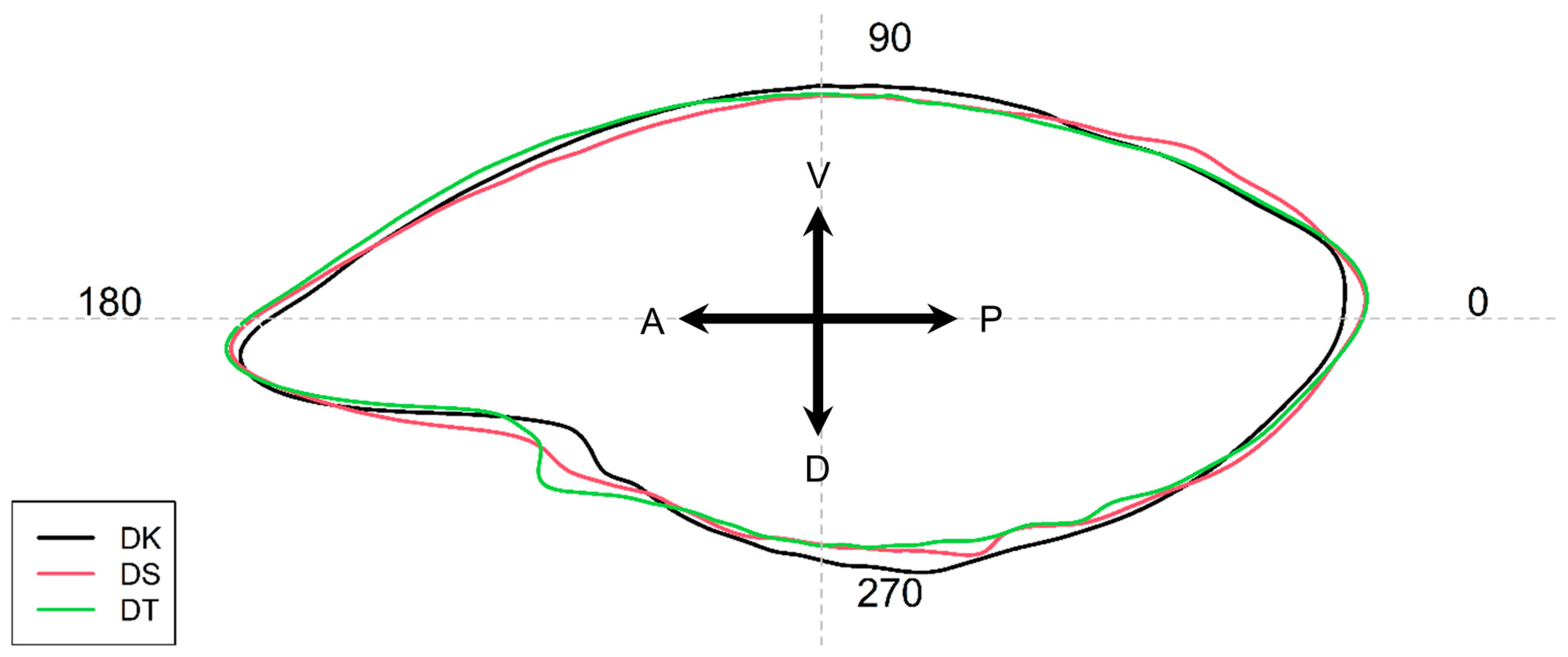
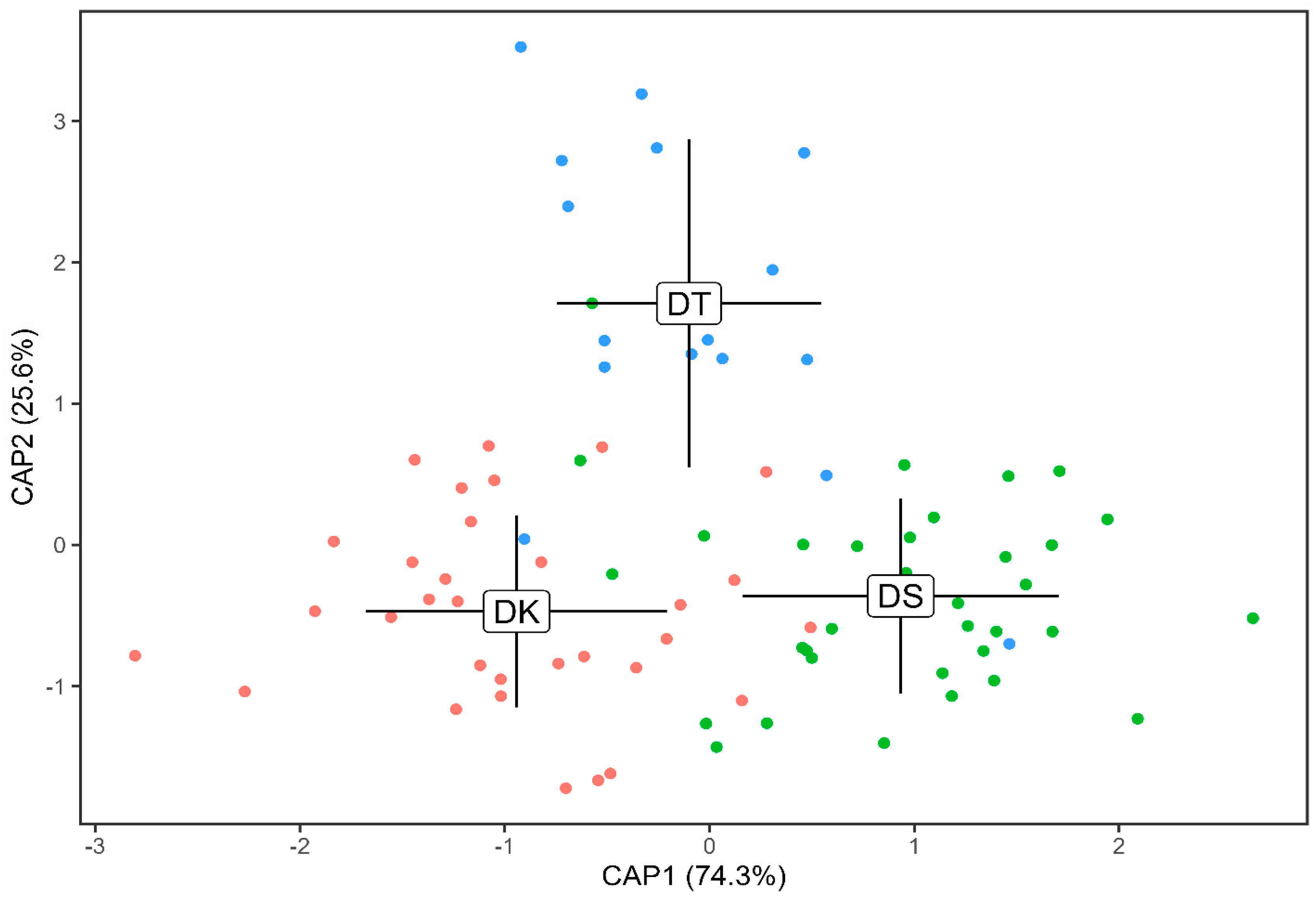
3.2. Otolith Shape Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Popper, A.N.; Ramcharitar, J.; Campana, S.E. Why otoliths? Insights from inner ear physiology and fisheries biology. Mar. Freshw. Res. 2005, 56, 497–504. [Google Scholar] [CrossRef]
- Poznar, M.; Stolarski, J.; Sikora, A.; Mazur, M.; Olesiak-Bańska, J.; Brach, K.; Ozyhar, A.; Dobryszycki, P. Fish otolith matrix macromolecule-64 (OMM-64) and its role in Calcium Carbonate Biomineralization. Cryst. Growth Des. 2020, 9, 5808–5819. [Google Scholar] [CrossRef]
- Edmonds, J.S.; Steckis, R.A.; Moran, M.J.; Caputi, N.; Morita, M. Stock delineation of pink snapper Pagrus auratus and tailor Pomatomus saltatrix from Western Australia by analysis of stable isotope and strontium/calcium ratios in otolith carbonate. J. Fish Biol. 1999, 55, 243–259. [Google Scholar] [CrossRef]
- Ovenden, J.R.; Lloyd, J.; Newman, S.J.; Keenan, C.P.; Slater, L.S. Spatial genetic subdivision between northern Australian and southeast Asian populations of Pristipomoides multidens: A tropical marine fish species. Fish. Res. 2002, 59, 57–69. [Google Scholar] [CrossRef]
- Horne, J.B.; Momigliano, P.; Welch, D.J.; Newman, S.J.; van Herwerden, L. Searching for common threads in threadfins: Phylogeography of Australian polynemids in space and time. Mar. Ecol. Prog. Ser. 2012, 449, 263–276. [Google Scholar] [CrossRef]
- Izzo, C.; Gillanders, B.M.; Ward, T.M. Movement Patterns and Stock Structure of Australian Sardine (Sardinops sagax) off South Australia and the East Coast: Implications for Future Stock Assessment and Management. Final Report to the Fisheries Research and Development Corporation; South Australian Research and Development Institute: West Beach, SA, USA, 2012; pp. 86–87. Available online: https://www.fish.gov.au/Archived-Reports/2014/Documents/Izzo_et_al_2012.pdf (accessed on 11 October 2022).
- Mackenzie, K.; Abaunza, P. Parasites as biological tags. In Stock Identification Methods: Applications in Fishery Science; Cadrin, S.X., Friedland, K.D., Waldman, J.R., Eds.; Elsevier Academic Press: San Diego, CA, USA, 2005; pp. 211–226. [Google Scholar]
- Friedland, K.D.; Cadrin, S.X. Analyses of calcified structures: Texture and spacing patterns. In Stock Identification Methods: Applications in Fishery Science; Cadrin, S.X., Friedland, K.D., Waldman, J.R., Eds.; Elsevier Academic Press: San Diego, CA, USA, 2005; pp. 185–195. [Google Scholar]
- Newman, S.J.; Steckis, R.A.; Edmonds, J.S.; Lloyd, J. Stock structure of the goldband snapper, Pristipomoides multidens (Pisces: Lutjanidae) from the waters of northern and western Australia by stable isotope ratio analysis of sagittal otolith carbonate. Mar. Ecol. Prog. Ser. 2000, 198, 239–247. [Google Scholar] [CrossRef]
- Newman, S.J.; Buckworth, R.C.; Mackie, M.C.; Lewis, P.D.; Wright, I.W.; Williamson, P.C.; Bastow, T.P.; Ovenden, J.R. Spatial subdivision among assemblages of Spanish mackerel, Scomberomorus commerson (Pisces: Scombridae) across northern Australia: Implications for fisheries management. Glob. Ecol. Biogeogr. 2009, 18, 711–723. [Google Scholar] [CrossRef]
- Newman, S.J.; Wright, I.W.; Rome, B.M.; Mackie, M.C.; Lewis, P.D.; Buckworth, R.C.; Ballagh, A.C.; Garrett, R.N.; Stapley, J.; Broderick, D.; et al. Stock structure of grey mackerel, Scomberomorus semifasciatus (Pisces: Scombridae) across northern Australia, based on otolith stable isotope chemistry. Environ. Biol. Fishes 2010, 89, 357–367. [Google Scholar] [CrossRef]
- Begg, G.A. Life history parameters. In Stock Identification Methods: Applications in Fishery Science; Cadrin, S.X., Friedland, K.D., Waldman, J.R., Eds.; Elsevier Academic Press: San Diego, CA, USA, 2005; pp. 119–150. [Google Scholar]
- Campana, S.E. Photographic atlas of fish otolith of the northwest Atlantic Ocean. In Canadian Special Publication of Fisheries and Aquatic Sciences 133; NCR, Research Press: Ottawa, ON, Canada, 2004; p. 284. [Google Scholar]
- Tuset, V.M.; Lombarte, A.; Assis, C.A. Otolith atlas for the western Mediterranean, north, and central eastern Atlantic. Sci. Mar. 2008, 72, 7–198. [Google Scholar] [CrossRef]
- Lombarte, A.; Castellón, A. Interspecific and intraspecific otolith variability in the genus Merluccius as determined by image analysis. Can. J. Zool. 1991, 69, 2442–2449. [Google Scholar] [CrossRef]
- Barnuevo, K.D.E.; Morales, C.J.C.; Calizo, J.K.S.; Delloro, E.S., Jr.; Añasco, C.P.; Babaran, R.P.; Lumayno, S.D.P. Distinct stocks of the redtail scad Decapterus kurroides Bleeker, 1855 (Perciformes: Carangidae) from the northern Sulu and Southern Sibuyan Seas, Philippines revealed from otolith morphometry and shape analysis. Fishes 2023, 8, 12. [Google Scholar] [CrossRef]
- Mille, T.; Mahé, K.; Villanueva, M.C.; De Pontual, H.; Ernande, B. Sagittal otolith morphogenesis asymmetry in marine fishes. J. Fish Biol. 2015, 87, 646–663. [Google Scholar] [CrossRef]
- Kontaş, S.; Bostanci, D.; Yedier, S.; Kurucu, G.; Polat, N. Investigation of fluctuating asymmetry in the four otolith characters of Merlangius merlangus collected from middle Black Sea. Turk. J. Marit. Marine Sci. 2018, 4, 128–138. Available online: https://dergipark.org.tr/en/pub/trjmms/issue/40277/485515 (accessed on 13 January 2023).
- Mejri, M.; Trojette, M.; Jmil, I.; Ben Faleh, A.; Chalh, A.; Quignard, J.-P.; Trabelsi, M. Fluctuating asymmetry in the otolith shape, length, width and area of Pagellus erythrinus collected from the Gulf of Tunis. Cah. Biol. Mar. 2020, 61, 1–7. [Google Scholar] [CrossRef]
- Geladakis, G.; Somarakis, S.; Koumoundouros, G. Differences in otolith shape and fluctuating asymmetry between reared and wild gilthead seabream (Sparus aurata Linnaeus, 1758). J. Fish Biol. 2021, 98, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Jawad, L.A.; Qasim, A.M.; Al-Faiz, N.A. Bilateral asymmetry in size of otolith of Otolithes ruber (Bloch and Schneider, 1801) collected from the marine waters of Iraq. Mar. Poll. Bull. 2021, 165, 112110. [Google Scholar] [CrossRef]
- Mahé, K.; MacKenzie, K.; Ider, D.; Massaro, A.; Hamed, O.; Jurado-Ruzafa, A.; Gonçalves, P.; Anastasopoulu, A.; Jadaud, A.; Mytilineou, C.; et al. Directional bilateral asymmetry in fish otolith: Potential tool to evaluate stock boundaries? Symmetry 2021, 13, 987. [Google Scholar] [CrossRef]
- Vignon, M.; Morat, F. Environmental and genetic determinant of otolith shape revealed by a non-indigenous tropical fish. Mar. Ecol. Prog. Ser. 2010, 411, 231–241. [Google Scholar] [CrossRef]
- Campana, S.E.; Neilson, J.D. Microstructure of fish otoliths. Can. J. Fish. Aquat. Sci. 1985, 42, 1014–1032. [Google Scholar] [CrossRef]
- Mosegaard, H.; Svedäng, H.; Taberman, K. Uncoupling of somatic and otolith growth rates in Arctic Char (Salvelinus alpinus) as an effect of differences in temperature response. Can. J. Fish. Aquat. Sci. 1988, 45, 1514–1524. [Google Scholar] [CrossRef]
- Libungan, L.A.; Oskarsson, G.J.; Slotte, A.; Jacobsen, J.A.; Palsson, S. Otolith shape: A population marker for Atlantic herring Clupea herengus. J. Fish Biol. 2015, 86, 1377–1395. [Google Scholar] [CrossRef] [PubMed]
- Berg, F.; Almeland, O.W.; Skadal, J.; Slotte, A.; Andersson, L.; Folkvord, A. Genetic factors have a major effect on growth, number of vertebrae and otolith shape in Atlantic Herring (Clupea harengus). PLoS ONE 2018, 13, e0190995. [Google Scholar] [CrossRef] [PubMed]
- Capoccioni, F.; Costa, C.; Aguzzi, J.; Menesatti, P.; Lombarte, A.; Ciccotti, E. Ontogenetic and environmental effects on otolith shape variability in three Mediterranean European eel (Anguilla anguilla L.) local stocks. J. Exp. Mar. Biol. Ecol. 2011, 397, 1–7. [Google Scholar] [CrossRef]
- Curcio, N.; Tombari, A.; Capitanio, F. Otolith morphology and feeding ecology of an Antarctic nototheniid, Lepidonotothen larseni. Antarct. Sci. 2013, 26, 124–132. [Google Scholar] [CrossRef]
- Tuset, V.M.; Imondi, R.; Aguado, G.; Otero-Ferrer, J.L.; Santschi, L.; Lombarte, A.; Love, M. Otolith patterns of rockfishes from the northeastern Pacific. J. Morphol. 2015, 276, 458–469. [Google Scholar] [CrossRef]
- Mille, T.; Mahé, K.; Cachera, M.; Villanueva, M.C.; de Pontual, H.; Ernande, B. Diet is correlated with otolith shape in marine fish. Mar. Ecol. Prog. Ser. 2016, 555, 167–184. [Google Scholar] [CrossRef]
- Cardinale, M.; Doering-Arjes, P.; Kastowsky, M.; Mosegaard, H. Effects of sex, stock, and environment on the shape of known-age Atlantic cod (Gadus morhua) otoliths. Can. J. Fish. Aquat. Sci. 2004, 61, 158–167. [Google Scholar] [CrossRef]
- Santos, M.D. Seasonality of commercially important pelagic fishes in the Philippines: Overfishing or climate change? In Proceedings of the International Workshop on Climate and Oceanic Fisheries, Rarotonga, Cook Islands, 3–5 October 2011. [Google Scholar]
- Philippine Statistics Authority. Fisheries Statistics of the Philippines (2018–2020); Philippine Statistics Authority: Quezon City, Philippines, 2020; p. 320.
- Kimura, S.; Katahira, K.; Kuriiwa, K. The red-fin Decapterus group (Perciformes: Carangidae) with the description of a new species, Decapterus smithvanizi. Ichthyol. Res. 2013, 60, 363–379. [Google Scholar] [CrossRef]
- Fricke, R.; Eschmeyer, W.N.; Van der Laan, R. (Eds.) Eschmeyer’s Catalog of Fishes: Genera, Species, References. Electronic Version. 2022. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp (accessed on 14 October 2022).
- Narido, C.I.; Palla, H.P.; Argente, F.A.T.; Geraldino, P.J.L. Population dynamics and fishery of Roughear scad Decapterus tabl Berry 1968 (Perciformes: Carangidae) in Camotes Sea, Central Philippines. Asian Fish. Sci. 2016, 29, 14–27. [Google Scholar] [CrossRef]
- Kimura, S. Family Carangidae: Decapterus tabl. In Commercial and Bycatch Market Fishes of Panay Island, Republic of the Philippines; Motomura, H., Alama, U.B., Muto, N., Babaran, R.P., Ishikawa, S., Eds.; Kagoshima University Museum, Kagoshima, University of the Philippines Visayas, Iloilo, and Research Institute for Humanities and Nature: Kyoto, Japan, 2017; p. 115. [Google Scholar]
- Motomura, H.; Alama, U.B.; Muto, N.; Babaran, R.P.; Ishikawa, S. Commercial and Bycatch Market Fishes of Panay Island, Republic of the Philippines; Kagoshima University Museum, Kagoshima, University of the Philippines Visayas, Iloilo, and Research Institute for Humanities and Nature: Kyoto, Japan, 2017; p. 246. [Google Scholar]
- Smith-Vaniz, W.F.; Carpenter, K.E.; Jiddawi, N.; Borsa, P.; Obota, C.; Yahya, S. Decapterus smithvanizi. IUCN Red List. Threat. Species 2018, e.T123424845A123494632. [Google Scholar] [CrossRef]
- Delloro, E.S., Jr.; Babaran, R.P.; Gaje, A.C.; Cambronero, P.T.; Alama, U.B.; Motomura, H. First record of slender scad, Decapterus smithvanizi (Actinopterygii: Perciformes: Carangidae) from the Philippines. Acta Ichthyol. Piscat. 2021, 51, 233–239. [Google Scholar] [CrossRef]
- Osman, Y.; Mahe, K.; El-Mahdy, S.; Mohammad, A.; Mehanna, S. Relationship between body and otolith morphological characteristics of sabre squirrelfish (Sargocentron spiniferum) from the southern Red Sea: Difference between right and left otoliths. Oceans 2021, 2, 624–633. [Google Scholar] [CrossRef]
- Libungan, L.A.; Pálsson, S. ShapeR: An R package to study otolith shape variation among fish populations. PLoS ONE 2015, 10, e0121102. [Google Scholar] [CrossRef]
- RStudio Team. RStudio: Integrated Development for R; RStudio, PBC: Boston, MA, USA, 2020; Available online: http://www.rstudio.com/ (accessed on 1 December 2022).
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package, Version 2.0-7. R Package. 2013. Available online: https://CRAN.R-project.org/package=vegan/ (accessed on 1 December 2022).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis, 2nd ed.; Springer: Cham, Switzerland, 2016. [Google Scholar]
- The Jamovi Project. Jamovi (Version 2.3.18) [Computer Software]. 2022. Available online: https://www.jamovi.org (accessed on 1 December 2022).
- Zischke, M.T.; Litherland, L.; Tilyard, B.R.; Stratford, N.J.; Jones, E.L.; Wang, Y.G. Otolith morphology of four mackerel species (Scomberomorus spp.) in Australia: Species differentiation and prediction for fisheries monitoring and assessment. Fish. Res. 2016, 176, 39–47. [Google Scholar] [CrossRef]
- Tuset, V.M.; Olivar, M.P.; Otero-Ferrer, J.L.; López-Pérez, C.; Hulley, P.A.; Lombarte, A. Morpho-functional diversity in Diaphus spp. (Pisces: Myctophidae) from the Central Atlantic Ocean: Ecological and evolutionary implications. Deep Sea Res. 2018, 1138, 46–59. [Google Scholar] [CrossRef]
- Deepa, K.P.; Aneesh Kumar, K.V.; Kottnis, O.; Nikki, R.; Bineesh, K.K.; Hashim, M.; Saravanane, N.; Sudhakar, M. Population variations of Opal fish, Bembrops caudimacula Steindachner, 1876 from Arabian Sea and Andaman Sea: Evidence from otolith morphometry. Reg. Stud. Mar. Sci. 2019, 25, 100466. [Google Scholar] [CrossRef]
- Lleonart, J.; Salat, J.; Torres, G.J. 2000. Removing allometric effects of body size in morphological analysis. J. Theor. Biol. 2000, 205, 85–93. [Google Scholar] [CrossRef]
- Elliott, N.G.; Haskard, K.; Koslow, J.A. Morphometric analysis of orange roughy (Hoplostethus atlanticus) off the continental slope of Southern Australia. J. Fish. Biol. 1995, 46, 202–220. [Google Scholar] [CrossRef]
- Sadighzadeh, Z.; Valinassab, T.; Vosugi, G.; Montallebi, A.A.; Fatemi, M.R.; Lombarte, A.; Tuset, V.M. use of otolith shape for stock identification of John’s snapper, Lutjanus johnii (Pisces: Lutjanidae), from Persian Gulf and the Oman Sea. Fish. Res. 2014, 155, 59–63. [Google Scholar] [CrossRef]
- Smith-Vaniz, W.F. Carangidae. In Smith’s Sea Fishes; Smith, M.M., Heemstra, P.C., Eds.; Springer: Berlin, Germany, 1986; pp. 638–661. [Google Scholar]
- de Figueiredo, J.L.; dos Santos, A.P.; Yamaguti, N.; Bernardes, R.A.; Del Bianco Rossi-Wongtschowski, C.L. Peixes da Zona Econômica Exclusiva da Região Sudeste-Sul do Brasil: Levantamento com Rede de Meia-Água; Editora da Universidade de São Paulo, Imprensa Oficial do Estado: São-Paulo, Brazil, 2002; p. 242. [Google Scholar]
- Cervignón, F.; Cipriani, R.; Fischer, W.; Garibaldi, L.; Hendrickx, M.; Lemus, A.J.; Márquez, R.; Putiers, J.M.; Robaina, G.; Rodriguez, B. Fichas Fao de Identificación de Especies Para Los Fines de la Pesca. Guía de Campo de Las Especies Comerciales Marinas Y de Aquas Salobres de la Costa Septentrional de Sur América; FAO: Rome, Italy, 1992; p. 513. [Google Scholar]
- Tuset, V.M.; Lombarte, A.; Gonzalez, J.A.; Pertusa, J.F.; Lorente, M. Comparative morphology of the sagittal otolith in Serranus spp. J. Fish Biol. 2003, 63, 1491–1504. [Google Scholar] [CrossRef]
- Bani, A.; Poursaeid, S.; Tuset, V. Comparative morphology of the sagittal otolith in three species of south Caspian gobies. J. Fish Biol. 2013, 82, 1321–1332. [Google Scholar] [CrossRef]
- Correia, A.T.; Coimbra, A.M.; Damasceino-Oliveira, A. Effect of hydrostatic pressure on otolith growth of early juveniles of Nile Tilapia Oreochromis niloticus. J. Fish. Biol. 2012, 81, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Gagliano, M.; McCormick, M.I. Feeding history influences otolith shape in tropical fish. Mar. Ecol. Prog. Ser. 2004, 278, 291–296. [Google Scholar] [CrossRef]
- Hüssy, K. Otolith shape in juvenile cod (Gadus morhua): Ontogenetic and environmental effects. J. Exp. Mar. Biol. Ecol. 2008, 364, 35–41. [Google Scholar] [CrossRef]
- Qiao, J.; Zhu, R.; Chen, K.; Zhang, D.; Yan, Y.; He, D. Comparative otolith morphology of two morphs of Schizopygopsis thermalis Herzenstein 1891 (Pisces, Cyprinidae) in a Headwater Lake on the Qunghai-Tibet Plateau. Fishes 2022, 7, 99. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | N | Mean Body Weight (g) ± SD | Mean Total Length (cm) ± SD |
|---|---|---|---|
| Decapterus kurroides | 32 | 115.853 ± 85.14 | 20.222 ± 4.90 |
| Decapterus smithvanizi | 34 | 59.682 ± 19.11 | 17.844 ± 1.86 |
| Decapterus tabl | 16 | 100.925 ± 29.42 | 21.863 ± 2.22 |
| OL/TL | X2 | df | W | |
|---|---|---|---|---|
| Decapterus kurroides | Decapterus smithvanizi | 53.0 | 2 | −6.82 *** |
| Decapterus kurroides | Decapterus tabl | 2 | −7.92 *** | |
| Decapterus smithvanizi | Decapterus tabl | 2 | −7.91 *** | |
| OH/OL | ||||
| Decapterus kurroides | Decapterus smithvanizi | 18.8 | 2 | −5.262 *** |
| Decapterus kurroides | Decapterus tabl | 2 | −5.042 ** | |
| Decapterus smithvanizi | Decapterus tabl | 2 | −0.676 |
| X2 | df | |||
|---|---|---|---|---|
| Size-related indices | OW | 65.75 *** | 2 | 0.81 |
| OL | 56.35 *** | 2 | 0.70 | |
| OH | 60.67 *** | 2 | 0.75 | |
| OA | 60.96 *** | 2 | 0.75 | |
| OP | 52.61 *** | 2 | 0.65 | |
| Shape-related indices | RE | 3.36 | 2 | 0.04 |
| SQ | 58.83 *** | 2 | 0.73 | |
| EL | 26.99 *** | 2 | 0.33 | |
| RO | 18.60 *** | 2 | 0.23 | |
| AR | 25.70 *** | 2 | 0.32 | |
| FF | 33.39 *** | 2 | 0.41 | |
| CO | 33.39 *** | 2 | 0.41 | |
| CI | 63.26 *** | 2 | 0.78 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morales, C.J.C.; Barnuevo, K.D.E.; Delloro, E.S., Jr.; Cabebe-Barnuevo, R.A.; Calizo, J.K.S.; Lumayno, S.D.P.; Babaran, R.P. Otolith Morphometric and Shape Distinction of Three Redfin Species under the Genus Decapterus (Teleostei: Carangidae) from Sulu Sea, Philippines. Fishes 2023, 8, 95. https://doi.org/10.3390/fishes8020095
Morales CJC, Barnuevo KDE, Delloro ES Jr., Cabebe-Barnuevo RA, Calizo JKS, Lumayno SDP, Babaran RP. Otolith Morphometric and Shape Distinction of Three Redfin Species under the Genus Decapterus (Teleostei: Carangidae) from Sulu Sea, Philippines. Fishes. 2023; 8(2):95. https://doi.org/10.3390/fishes8020095
Chicago/Turabian StyleMorales, Christian James C., Kyle Dominic E. Barnuevo, Emmanuel S. Delloro, Jr., Roxanne A. Cabebe-Barnuevo, Jenylle Kate S. Calizo, Sanny David P. Lumayno, and Ricardo P. Babaran. 2023. "Otolith Morphometric and Shape Distinction of Three Redfin Species under the Genus Decapterus (Teleostei: Carangidae) from Sulu Sea, Philippines" Fishes 8, no. 2: 95. https://doi.org/10.3390/fishes8020095
APA StyleMorales, C. J. C., Barnuevo, K. D. E., Delloro, E. S., Jr., Cabebe-Barnuevo, R. A., Calizo, J. K. S., Lumayno, S. D. P., & Babaran, R. P. (2023). Otolith Morphometric and Shape Distinction of Three Redfin Species under the Genus Decapterus (Teleostei: Carangidae) from Sulu Sea, Philippines. Fishes, 8(2), 95. https://doi.org/10.3390/fishes8020095