

Decadal Changes in the Annual Growth of Two Commercial Ommastrephid Species in the Pacific Ocean



Abstract
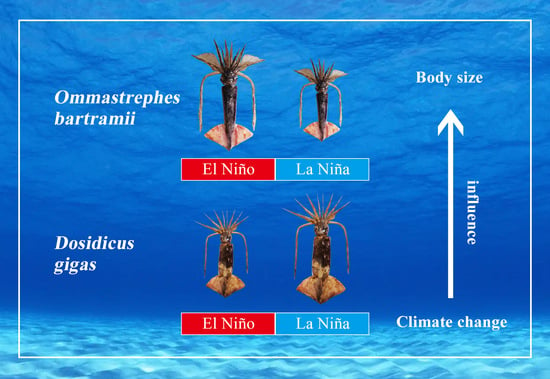
1. Introduction
2. Materials and Methods
2.1. Sampling and Measurement
2.2. The Relationship between Mantle Length and Body Weight (LWR)
2.3. Model Comparison
2.4. Relative Condition Factor
2.5. Climatic Index Data
3. Results
3.1. Mantle Length Distribution
3.2. LWR
3.3. Recommended LWR and Temporal and Sexual Variations in LWR
3.4. The Relative Condition Factor (Kn)
4. Discussion
4.1. Body Size of the Two Squids
4.2. LWR of Two Squids and Their Sexual and Interannual Variations
4.3. Relative Condition Factors of Two Squids
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, X.J.; Lu, H.J.; Liu, B.L.; Tian, S.Q. Current exploitation and some scientific issues in the sustainable utilization of Ommastrephidae. J. Shanghai Ocean. Univ. 2012, 21, 831–840. [Google Scholar]
- Arkhipkin, A.I.; Rodhouse, P.G.K.; Pierce, G.J.; Sauer, W.; Sakai, M.; Allcock, L.; Arguelles, J.; Bower, J.R.; Castillo, G.; Ceriola, L.; et al. World squid fisheries. Rev. Fish. Sci. Aquac. 2015, 23, 92–252. [Google Scholar] [CrossRef]
- Wang, J.T.; Chen, X.J.; Tanaka, K.; Cao, J.; Chen, Y. Environmental influences on commercial oceanic ommastrephid squids: A stock assessment perspective. Sci. Mar. 2017, 81, 37–47. [Google Scholar] [CrossRef]
- Boyle, P.R. Cephalopod biology in the fisheries context. Fish. Res. 1990, 8, 303–321. [Google Scholar] [CrossRef]
- Rodhouse, P.G. Large-scale range expansion and variability in ommastrephid squid populations: A review of environmental links. CalCOFI Rep. 2008, 49, 83–89. Available online: https://oceanrep.geomar.de/id/eprint/53855 (accessed on 5 September 2022).
- Yu, W.; Chen, X.J.; Liu, L.W. Synchronous Variations in Abundance and Distribution of Ommastrephes bartramii and Dosidicus gigas in the Pacific Ocean. J. Ocean. Univ. China 2021, 20, 695–705. [Google Scholar] [CrossRef]
- Parry, M. Feeding behavior of two ommastrephid squids Ommastrephes bartramii and Sthenoteuthis oualaniensis off Hawaii. Mar. Ecol. Prog. Ser. 2006, 318, 229–235. [Google Scholar] [CrossRef]
- Cao, J.; Chen, X.J.; Liu, B.L.; Tian, S.Q.; Qian, W.G. Review on the relationship between stock recruitment of squid and oceanographic environment. J. Shanghai Ocean Univ. 2010, 19, 232–239. [Google Scholar]
- Caccavo, J.A.; Christiansen, H.; Constable, A.J.; Ghigliotti, L.; Trebilco, R.; Brooks, C.M.; Cotte, C.; Desvignes, T.; Dornan, T.; Jones, C.D.; et al. Productivity and Change in Fish and Squid in the Southern Ocean. Front. Ecol. Evol. 2021, 9, 624918. [Google Scholar] [CrossRef]
- Chemshirova, I.; Hoving, H.J.; Arkhipkin, A. Temperature effects on size, maturity, and abundance of the squid Illex argentinus (Cephalopoda, Ommastrephidae) on the Patagonian Shelf. Estuar. Coast. Shelf Sci. 2021, 255, 107343. [Google Scholar] [CrossRef]
- Wang, Y.; Han, P.W.; Fang, Z.; Chen, X.J. Climate-induced life cycle and growth variations of neon flying squid (Ommastrephes bartramii) in the North Pacific Ocean. Aquac. Fish. 2021, 8, 211–220. [Google Scholar] [CrossRef]
- Yu, W.; Wen, J.; Chen, X.J.; Gong, Y.; Liu, B.L. Trans-Pacific multidecadal changes of habitat patterns of two squid species. Fish. Res. 2021, 233, 105762. [Google Scholar] [CrossRef]
- Murata, M. Oceanic resources of squids. Mar. Freshw. Behav. Phy 1990, 18, 19–71. [Google Scholar] [CrossRef]
- Chen, X.J.; Liu, B.L.; Chen, Y. A review of the development of Chinese distant-water squid jigging fisheries. Fish. Res. 2008, 89, 211–221. [Google Scholar] [CrossRef]
- Fang, Z.; Han, P.W.; Wang, Y.; Chen, Y.Y.; Chen, X.J. Interannual variability of body size and beak morphology of the squid Ommastrephes bartramii in the North Pacific Ocean in the context of climate change. Hydrobiologia 2021, 848, 1295–1309. [Google Scholar] [CrossRef]
- Bower, J.R.; Ichii, T. The red flying squid (Ommastrephes bartramii): A review of recent research and the fishery in Japan. Fish. Res. 2005, 76, 39–55. [Google Scholar] [CrossRef]
- Fang, Z.; Thompson, K.; Jin, Y.; Chen, X.J.; Chen, Y. Preliminary analysis of beak stable isotopes (δ13C and δ15N) stock variation of neon flying squid, Ommastrephes bartramii, in the North Pacific Ocean. Fish. Res. 2016, 177, 153–163. [Google Scholar] [CrossRef]
- Fang, Z.; Li, J.H.; Thompson, K.; Hu, F.F.; Chen, X.J.; Liu, B.L.; Chen, Y. Age, growth, and population structure of the red flying squid (Ommastrephes bartramii) in the North Pacific Ocean, determined from beak microstructure. Fish. Bull. 2016, 114, 34–44. [Google Scholar] [CrossRef]
- Han, P.W.; Fang, Z.; Li, N.; Chen, X.J. Migration Route Reconstruction of Different Cohorts of Ommastrephes bartramii in the North Pacific Based on Statolith Microchemistry. Front. Mar. Sci. 2022, 9, 832639. [Google Scholar] [CrossRef]
- Yu, W.; Chen, X.J.; Yi, Q.; Gao, G.; Chen, Y. Impacts of climatic and marine environmental variations on the spatial distribution of Ommastrephes bartramii in the Northwest Pacific Ocean. Acta Oceanol. Sin. 2016, 35, 108–116. [Google Scholar] [CrossRef]
- Wang, Y.; Han, P.W.; Chen, X.J.; Fang, Z. Interannual and ontogenetic stage differences on trophic ecology of neon flying squid Ommastrephes bartramii inferred from stable isotope analyses in beaks. Fish. Res. 2022, 249, 106252. [Google Scholar] [CrossRef]
- Argüelles, J.; Rodhouse, P.G.; Villegas, P.; Castillo, G. Age, growth and population structure of the jumbo flying squid Dosidicus gigas in Peruvian waters. Fish. Res. 2001, 54, 51–61. [Google Scholar] [CrossRef]
- Taipe, A.; Yamashiro, C.; Mariategui, L.; Rojas, P.; Roque, C. Distribution and concentrations of jumbo flying squid (Dosidicus gigas) off the Peruvian coast between 1991 and 1999. Fish. Res. 2001, 54, 21–32. [Google Scholar] [CrossRef]
- Hu, G.Y.; Yu, W.; Li, B.L.; Chen, X.J.; Chen, Y.; Li, J.H. Impacts of El Niño on the somatic condition of Humboldt squid based on the beak morphology. J. Ocean. Limnol. 2019, 37, 1440–1448. [Google Scholar] [CrossRef]
- Liu, B.L.; Chen, X.J.; Yi, Q. A comparison of fishery biology of jumbo flying squid, Dosidicus gigas outside three exclusive economic zones in the Eastern Pacific Ocean. Chin. J. Oceanol. Limnol. 2013, 31, 523–533. [Google Scholar] [CrossRef]
- Nigmatullin, C.M.; Nesis, K.N.; Arkhipkin, A.I. A review of the biology of the jumbo squid Dosidicus gigas (Cephalopoda: Ommastrephidae). Fish. Res. 2001, 54, 9–19. [Google Scholar] [CrossRef]
- Argüelles, J.; Tafur, R.; Taipe, A.; Villegas, P.; Keyl, F.; Dominguez, N.; Salazar, M. Size increment of jumbo flying squid Dosidicus gigas mature females in Peruvian waters, 1989–2004. Prog. Oceanogr. 2008, 79, 308–312. [Google Scholar] [CrossRef]
- Ibáñez, C.M.; Sepúlveda, R.D.; Ulloa, P.; Keyl, F.; Pardo-Gandarillas, M.C. The biology and ecology of the jumbo squid Dosidicus gigas (Cephalopoda) in Chilean waters: A review. Lat. Am. J. Aquat. Res. 2015, 43, 402–414. [Google Scholar] [CrossRef]
- Chen, P.; Chen, X.J.; Yu, W.; Lin, D.M. Interannual Abundance Fluctuations of Two Oceanic Squids in the Pacific Ocean Can Be Evaluated Through Their Habitat Temperature Variabilities. Front. Mar. Sci. 2021, 8, 770224. [Google Scholar] [CrossRef]
- Jackson, G.; Moltschaniwskyj, N. Spatial and temporal variation in growth rates and maturity in the Indo-Pacific squid Sepioteuthis lessoniana (Cephalopoda: Loliginidae). Mar. Biol. 2022, 140, 747–754. [Google Scholar] [CrossRef]
- Forsythe, J.W. Accounting for the effect of temperature on squid growth in nature: From hypothesis to practice. Mar. Freshw. Res. 2004, 55, 331–339. [Google Scholar] [CrossRef]
- Arkhipkin, A.; Argüelles, J.; Shcherbich, Z.; Yamashiro, C. Ambient temperature influences adult size and life span in jumbo squid (Dosidicus gigas). Can. J. Fish. Aquat. Sci. 2015, 72, 400–409. [Google Scholar] [CrossRef]
- Takahara, H.; Kidokoro, H.; Sakurai, Y. High temperatures may halve the lifespan of the Japanese flying squid, Todarodes pacificus. J. Nat. Hist. 2017, 51, 2607–2614. [Google Scholar] [CrossRef]
- Hoving, H.J.T.; Gilly, W.F.; Markaida, U.; Benoit-Bird, K.; Brown, Z.W.; Daniel, P.; Field, W.F.; Parassenti, L.; Liu, B.L.; Campos, B. Extreme plasticity in life-history strategy allows a migratory predator (jumbo squid) to cope with a changing climate. Glob. Chang. Biol. 2013, 19, 2089–2103. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Chen, X.J.; Chen, C.S.; Zhang, Y. Impacts of oceanographic factors on interannual variability of the winter-spring cohort of neon flying squid abundance in the Northwest Pacific Ocean. Acta Oceanol. Sin. 2017, 36, 48–59. [Google Scholar] [CrossRef]
- Feng, Z.P.; Yu, W.; Chen, X.J. Concurrent habitat fluctuations of two economically important marine species in the Southeast Pacific Ocean off Chile in relation to ENSO perturbations. Fish. Oceanogr. 2021, 31, 123–134. [Google Scholar] [CrossRef]
- Zepeda-Benitez, V.Y.; Morales-Bojórquez, E.; Díaz-Uribe, J.G.; Nevárez-Martínez, M.O.; Hernández-Herrera, A.; López-Martínez, J. Implementation of catch-at-age model for the jumbo squid Dosidicus gigas. Ecol. Model. 2017, 344, 6–16. [Google Scholar] [CrossRef]
- Granados-Amores, J.; Salinas-Zavala, C.A.; Flores-ortega, J.R.; Díaz-Santana-Iturrios, M. Length-weight relationship and condition factor for 7 loliginid squid species in Mexican waters. Cienc. Mar. 2019, 45, 175–180. [Google Scholar] [CrossRef]
- Seçer, B.; Sungur, S.; Çiçek, E.; Mouludi-Saleh, A.; Eagderi, S. Length-weight relationship and condition factor of endemic genus Seminemacheilus (Teloestei = Nemacheilidae) for Turkey. Limnol. Freshw. Biol. 2021, 4, 1152–1155. [Google Scholar] [CrossRef]
- Acarli, D.; Kale, S.; Çakır, K. Length–Weight Relationships of Eighteen Fishes and a Cephalopod from Gökçeada Island, Northern Aegean Sea, Turkey. Thalassas 2022, 38, 479–486. [Google Scholar] [CrossRef]
- Le Cren, E.D. The length-weight relationship and seasonal cycle in gonad weight and condition in the Perch (Perca fluviatilis). J. Anim. Ecol. 1951, 20, 201–219. [Google Scholar] [CrossRef]
- Froese, R. Cube law, condition factor and weight-length relationships: History, meta-analysis and recommendations. J. Appl. Ichthyol. 2006, 22, 241–253. [Google Scholar] [CrossRef]
- Jisr, N.; Younes, G.; Sukhn, C.; El-Dakdouki, M.H. Length-weight relationships and relative condition factor of fish inhabiting the marine area of the Eastern Mediterranean city, Tripoli-Lebanon. Egypt. J. Aquatic. Res. 2018, 44, 299–305. [Google Scholar] [CrossRef]
- Lipinski, M.R.; Underhill, L.G. Sexual maturation in squid: Quantum or continuum. S. Afr. J. Mar. Sci. 1995, 15, 207–223. [Google Scholar] [CrossRef]
- Han, P.W.; Li, N.; Fang, Z.; Chen, X.J. Heterogeneity of mantle length-body mass relationship in different Ommastrephes bartramii populations based on linear mixed model. S. China Fish. Sci. 2020, 16, 12–20. [Google Scholar] [CrossRef]
- Ma, Q.Y.; Tian, S.Q.; Han, D.Y.; Richard, K.; Gao, C.H.; Liu, W.C. Growth and maturity heterogeneity of three croaker species in the East China Sea. Reg Stud. Mar. Sci. 2021, 41, 101483. [Google Scholar] [CrossRef]
- Schielzeth, H.; Dingemanse, N.J.; Nakagawa, S.; Westneat, D.F.; Allegue, H.; Teplitsky, C.; Réale, D.; Dochtermann, N.A.; Garamszegi, L.Z.; Araya-Ajoy, Y.G. Robustness of linear mixed-effects models to violations of distributional assumptions. Methods Ecol. Evol. 2020, 11, 1141–1152. [Google Scholar] [CrossRef]
- Qian, S.S.; Cuffney, T.F.; Alameddine, I.; McMahon, G.; Reckhow, K.H. On the application of multilevel modeling in environmental and ecological studies. Ecology 2010, 91, 355–361. [Google Scholar] [CrossRef]
- Xu, H.; Sun, Y.J.; Wang, X.J.; Wang, J.; Fu, Y. Linear mixed-effects models to describe individual tree crown width for China-fir in Fujian province, southeast China. PLoS ONE 2015, 10, e0122257. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Akaike, H. A New Look at the Statistical Model Identification. IEEE Trans. Automat. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Cai, K.; Richard, K.D.; Ma, Q.Y.; Han, X.B.; Qin, S. Growth Heterogeneity of Chub Mackerel (Scomber japonicus) in the Northwest Pacific Ocean. J. Mar. Sci. Eng. 2022, 10, 301. [Google Scholar] [CrossRef]
- Ferreri, G.A.B. Length-Weight Relationships and Condition Factors of the Humboldt Squid (Dosidicus Gigas) from the Gulf of california and the Pacific Ocean. J. Shellfish Res. 2014, 33, 769–780. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, X.J.; Fang, Z. Effects of marine environment variation on the growth of neon flying squid (Ommastrephes bartramii) in the north Pacific Ocean. J. Fish. China 2022, 46, 569–582. [Google Scholar] [CrossRef]
- Chen, X.J.; Zhao, X.H.; Chen, Y. Influence of El Niño/La Niña on the western winter-spring cohort of neon flying squid (Ommastrephes bartramii) in the northwestern Pacific Ocean. ICES J. Mar. Sci. 2007, 64, 1152–1160. [Google Scholar] [CrossRef]
- Nishikawa, H.; Toyoda, T.; Masuda, S.; Ishikawa, Y.; Sasaki, Y.; Igarashi, H.; Sakai, M.; Seito, M.; Awaji, T. Wind-induced stock variation of the neon flying squid (Ommastrephes bartramii) winter-spring cohort in the subtropical North Pacific Ocean. Fish. Oceanogr. 2015, 24, 229–241. [Google Scholar] [CrossRef]
- Jones, J.B.; Pierce, G.J.; Saborido-Rey, F.; Brickle, P.; Kuepper, F.C.; Shcherbich, Z.N.; Arkhipkin, A.I. Size-dependent change in body shape and its possible ecological role in the Patagonian squid (Doryteuthis gahi) in the Southwest Atlantic. Mar. Biol. 2019, 166, 54. [Google Scholar] [CrossRef]
- Ning, X.; Lu, H.J.; Liu, K.; Chen, Z.Y.; Chen, X.J. Fisheries biological characteristics of Japanese common squid (Todarodes pacificus) in spring in the La Niña a year of 2018 in the East China Sea. J. Fish. China 2020, 44, 1676–1684. [Google Scholar] [CrossRef]
- Waluda, C.M.; Rodhouse, P.G.K. Remotely sensed mesoscale oceanography of the Central Eastern Pacific and recruitment variability in Dosidicus gigas. Mar. Ecol. Prog. Ser. 2006, 310, 25–32. [Google Scholar] [CrossRef]
- Robinson, C.J.; Gómez-Gutiérrez, J.; Markaida, U.; Gilly, W.F. Prolonged decline of jumbo squid (Dosidicus gigas) landings in the Gulf of California is associated with chronically low wind stress and decreased chlorophyll a after El Niño 2009–2010. Fish. Res. 2016, 173, 128–138. [Google Scholar] [CrossRef]
- Hu, G.Y.; Chen, X.J.; Fang, Z. Effect of individual growth on beak morphometry of jumbo flying squid, Dosidicus gigas off the Peruvian Exclusive Economic Zone. J. Fish. China 2016, 40, 36–44. [Google Scholar] [CrossRef]
- Markaida, U.; Quiñónez-Velázquez, C.; Sosa-Nishizaki, O. Age, growth and maturation of jumbo squid Dosidicus gigas (Cephalopoda: Ommastrephidae) from the Gulf of California, Mexico. Fish. Res. 2004, 66, 31–47. [Google Scholar] [CrossRef]
- O’Dor, R.K. Chapter 11. Squid life-history strategies. In Squid Recruitment Dynamic; Rodhouse, P.D., Dawe, E.G., O’Dor, R.K., Eds.; FAO: Rome, Italy, 1998; pp. 233–254. [Google Scholar]
- Froese, R.; Tsikliras, A.C.; Stergiou, K.I. Editorial Note on Weight–Length Relations of Fishes. Acta Ichthyol. Piscat. 2011, 41, 261–263. [Google Scholar] [CrossRef]
- Hile, R. Age and growth of the cisco, Leucichthys artedi (le Sueur), in the lakes of the Northeastern highlands, Wisconsin. Bull. Bur. Fish. 1936, 48, 211–317. [Google Scholar]
- Keyl, F.; Argüelles, J.; Mariategui, L.; Tafur, R.; Wolff, M.; Yamashiro, C. A hypothesis on range expansion and spatio-temporal shifts in size-at-maturity of jumbo squid (Dosidicus gigas) in the Eastern Pacific Ocean. CCOFI. Rep. 2008, 49, 119–128. [Google Scholar]
- Qian, M.T.; Gong, J.W.; Fan, J.T.; Yu, W.; Chen, X.J.; Qian, W.G. Variations in the abundance and spatial distribution of Ommastrephes bartramii in the Northwest Pacific Ocean based on photosynthetic active radiation. Haiyang Xuebao 2020, 42, 44–53. [Google Scholar] [CrossRef]
- Vasconcelos, P.; Pereira, F.; Carvalho, A.N.; Gaspar, M.B. Weight-length relationships and relative growth of the cuttlefish (Sepia officinalis): Causes and effects of hypoallometry. Thalassas 2018, 34, 323–331. [Google Scholar] [CrossRef]
- Chen, J.Y.; Ching, T.Y.; Chen, C.S. Inter-annual variability in growth and maturation of the swordtip squid Uroteuthis edulis in Yilan Bay off northeastern Taiwan. J. Mar. Sci. Technol. 2022, 30, 158–171. [Google Scholar] [CrossRef]
- Dawe, E.G.; Beck, P.C. Population structure, growth, and sexual maturation of short-finned squid (Illex illecebrosus) at Newfoundland. Can. J. Fish. Aquat. Sci. 1997, 54, 137–146. [Google Scholar] [CrossRef]
- Chen, X.J.; Han, F.; Zhu, K.; Punt, A.E.; Lin, D.M. The breeding strategy of female jumbo squid Dosidicus gigas: Energy acquisition and allocation. Sci. Rep. 2020, 10, 9639. [Google Scholar] [CrossRef] [PubMed]
- Fauziyah.; Purwiyanto, A.I.S.; Agustriani, F.; Putri, W.A.E. Growth aspect of squid (Loligo chinensis) from the Banyuasin Coastal Waters, South Sumatra, Indonesia. Ecol. Montenegrina 2020, 27, 1–10. [Google Scholar] [CrossRef]
- Pecl, G.T.; Moltschaniwskyj, N.A.; Tracey, S.R.; Jordan, A.R. Inter-annual plasticity of squid life history and population structure: Ecological and management implications. Oecologia 2004, 139, 515–524. [Google Scholar] [CrossRef] [PubMed]
- Morato, T.; Afonso, P.; Loirinho, P.; Barreiros, J.P.; Sanstos, R.S.; Nash, R.D.M. Length-weight relationships for 21 costal fish species of the Azores, Northeastern Atlantic. Fish. Res. 2001, 50, 297–302. [Google Scholar] [CrossRef]
- Yatsu, A.; Midorikawa, S.; Shimada, T.; Uozumi, Y. Age and growth of the neon flying squid, Ommastrephes bartrami, in the North Pacific Ocean. Fish. Res. 1997, 29, 257–270. [Google Scholar] [CrossRef]
- Nigmatullin, C.M.; Markaida, U. Oocyte development, fecundity and spawning strategy of large sized jumbo squid Dosidicus gigas (Oegopsida: Ommastrephinae). J. Mar. Biol. Assoc. UK 2009, 89, 789–801. [Google Scholar] [CrossRef]
- Nishikawa, H.; Igarashi, H.; Ishikawa, Y.; Sakai, M.; Kato, Y.; Ebina, M.; Usui, N.; Kamachi, M.; Awaji, T. Impact of paralarvae and juveniles feeding environment on the neon flying squid (Ommastrephes bartramii) winter-spring cohort stock. Fish. Oceanogr. 2014, 23, 289–303. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Ommastrephes bartramii | Dosidicus gigas | ||
|---|---|---|---|---|
| Female | Male | Female | Male | |
| 2008 | 0 | 0 | 304 | 72 |
| 2009 | 729 | 703 | 565 | 146 |
| 2010 | 1194 | 1317 | 470 | 114 |
| 2011 | 324 | 278 | 925 | 343 |
| 2012 | 301 | 464 | 469 | 217 |
| 2013 | 247 | 223 | 285 | 167 |
| 2014 | 150 | 103 | 221 | 142 |
| 2015 | 529 | 494 | 370 | 257 |
| 2016 | 362 | 368 | 214 | 135 |
| 2017 | 287 | 326 | 66 | 38 |
| 2018 | 116 | 45 | 0 | 0 |
| Total | 4239 | 4321 | 3889 | 1631 |
| Model | Equation | Log-Transformed | Ommastrephes bartramii | Dosidicus gigas | ||
|---|---|---|---|---|---|---|
| AIC | RMSE | AIC | RMSE | |||
| GLM | W = a × Lb | ln(W) = ln(a) + b × ln(L) | −16,614 | 0.09165 | −9064 | 0.10641 |
| Y.I | W = [a × exp(ReY.I)] × Lb | ln(W) = [ln(a) + ReY.I] + b × ln(L) | −17,044 | 0.08914 | −9429 | 0.10244 |
| S.I | W = [a × exp(ReS.I)] × Lb | ln(W) = [ln(a) + ReS.I] + b × ln(L) | −16,650 | 0.09132 | −9144 | 0.10538 |
| Y and S.I | W = [a × exp(ReY.I) × exp(ReS.I)] × Lb | ln(W) = [ln(a) + ReY.I + ReS.I] + b × ln(L) | −17,073 | 0.08887 | −9542 | 0.10119 |
| Y.S | W = a × L(b + ReY.S) | ln(W) = ln(a) + (b + ReY.S) × ln(L) | −17,018 | 0.08919 | −9408 | 0.1025 |
| S.S | W = a × L(b + ReS.S) | ln(W) = ln(a) + (b + ReS.S) × ln(L) | −16,649 | 0.09132 | −9147 | 0.10535 |
| Y and S.S | W = a × L(b + ReY.S + ReS.S) | ln(W) = ln(a) + (b + ReY.S + ReS.S) × ln(L) | −17,062 | 0.08893 | −9539 | 0.10121 |
| Y.I and S | W = [a × exp(ReY.I)] × L(b + ReY.S) | ln(W) = [ln(a) + ReY.I] + (b + ReY.S) × ln(L) | −17,171 | 0.08823 | −9605 | 0.10033 |
| S.I and S | W = [a × exp(ReS.I)] × L(b + ReS.S) | ln(W) = [ln(a) + ReS.I] + (b + ReS.S) × ln(L) | −16,646 | 0.09131 | −9163 | 0.10515 |
| Y and S.I and S | W = [a × exp(ReY.I) × exp(ReS.I)] × L(b + ReY.S + ReS.S) | ln(W) = [ln(a) + ReY.I + ReS.I] + (b + ReY.S + ReS.S) × ln(L) | −17,216 | 0.08794 | −9748 | 0.09893 |
| Fishing Ground | Anomalous Climate Conditions | Year |
|---|---|---|
| Ommastrephes bartramii | El Niño event | 2009, 2015 |
| La Niña event | 2010, 2011 | |
| Dosidicus gigas | El Niño event | 2014, 2015, 2017 |
| La Niña event | 2010, 2013, 2017 |
| Species | Variables | Fixed Effects | Random Effects | Estimate of Parameter | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| ln(a) | b | ln(a) | b | a (×10−5) | b | |||||
| Estimate | Confidence Interval | Estimate | Confidence Interval | |||||||
| Ommastrephes bartramii | Year | 2009 | −11.2281 | 3.1395 | 0.5018 | [0.2622, 0.7414] | −0.0851 | [−0.1268, −0.0433] | 2.1961 | 3.0544 |
| 2010 | −0.3874 | [−0.6083, −0.1677] | 0.0656 | [0.0270, 0.1043] | 0.9025 | 3.2050 | ||||
| 2011 | −0.7620 | [−1.0769, −0.4474] | 0.1331 | [0.0776, 0.1886] | 0.6205 | 3.2725 | ||||
| 2012 | 0.7982 | [0.4996, 1.0959] | −0.1391 | [−0.1917, −0.0864] | 2.9536 | 3.0003 | ||||
| 2013 | −0.1458 | [−0.4983, 0.2065] | 0.0275 | [−0.0355, 0.0905] | 1.1491 | 3.1670 | ||||
| 2014 | −0.4229 | [−0.9091, 0.0623] | 0.0746 | [−0.0128, 0.1621] | 0.8710 | 3.2140 | ||||
| 2015 | −0.6278 | [−0.8639, −0.3927] | 0.1121 | [0.0708, 0.1535] | 0.7096 | 3.2515 | ||||
| 2016 | −0.0406 | [−0.3187, 0.2377] | 0.0025 | [−0.0463, 0.0513] | 1.2766 | 3.1420 | ||||
| 2017 | 0.1710 | [−0.1675, 0.5106] | −0.0310 | [−0.0906, 0.0283] | 1.5775 | 3.1084 | ||||
| 2018 | 0.9155 | [0.4047, 1.4297] | −0.1600 | [−0.2495, −0.0711] | 3.3211 | 2.9794 | ||||
| Sex | F | −0.0940 | [−0.2563, 0.0646] | 0.0158 | [−0.0109, 0.0431] | 1.2102 | 3.1553 | |||
| M | 0.0940 | [−0.0646, 0.2563] | −0.0158 | [−0.0431, 0.0109] | 1.4606 | 3.1237 | ||||
| Dosidicus gigas | Year | 2008 | −10.8720 | 3.0629 | −0.1811 | [−0.5479, 0.1859] | 0.0331 | [−0.0311, 0.0972] | 1.5838 | 3.0960 |
| 2009 | −0.8622 | [−1.1057, −0.6182] | 0.1469 | [0.1033, 0.1903] | 0.8016 | 3.2098 | ||||
| 2010 | −1.0330 | [−1.2639, −0.8018] | 0.1788 | [0.1373, 0.2202] | 0.6756 | 3.2418 | ||||
| 2011 | −0.6823 | [−0.9087, −0.4556] | 0.1218 | [0.0804, 0.1632] | 0.9595 | 3.1847 | ||||
| 2012 | −0.8741 | [−1.1847, −0.5629] | 0.1574 | [0.1022, 0.2126] | 0.7921 | 3.2204 | ||||
| 2013 | 0.6522 | [0.2367, 1.0677] | −0.1153 | [−0.1902, −0.0404] | 3.6444 | 2.9476 | ||||
| 2014 | 1.1261 | [0.6235, 1.6281] | −0.1902 | [−0.2806, −0.0997] | 5.8535 | 2.8727 | ||||
| 2015 | 0.6666 | [0.4012, 0.9321] | −0.1158 | [−0.1647, −0.0668] | 3.6972 | 2.9472 | ||||
| 2016 | 0.2332 | [−0.1436, 0.6101] | −0.0554 | [−0.1245, 0.0137] | 2.3968 | 3.0076 | ||||
| 2017 | 0.9545 | [0.2488, 1.6590] | −0.1613 | [−0.2891, −0.0334] | 4.9308 | 2.9016 | ||||
| Sex | F | −0.1175 | [−0.2307, −0.0045] | 0.0240 | [0.0009, 0.0470] | 1.6878 | 3.0869 | |||
| M | 0.1175 | [0.0021, 0.2330] | −0.0240 | [−0.0475, −0.0004] | 2.1351 | 3.0390 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, P.; Li, J.; Dan, Y.; Liu, B.; Chen, X.; Fang, Z. Decadal Changes in the Annual Growth of Two Commercial Ommastrephid Species in the Pacific Ocean. Fishes 2022, 7, 280. https://doi.org/10.3390/fishes7050280
Han P, Li J, Dan Y, Liu B, Chen X, Fang Z. Decadal Changes in the Annual Growth of Two Commercial Ommastrephid Species in the Pacific Ocean. Fishes. 2022; 7(5):280. https://doi.org/10.3390/fishes7050280
Chicago/Turabian StyleHan, Peiwu, Jianhua Li, Yating Dan, Bilin Liu, Xinjun Chen, and Zhou Fang. 2022. "Decadal Changes in the Annual Growth of Two Commercial Ommastrephid Species in the Pacific Ocean" Fishes 7, no. 5: 280. https://doi.org/10.3390/fishes7050280
APA StyleHan, P., Li, J., Dan, Y., Liu, B., Chen, X., & Fang, Z. (2022). Decadal Changes in the Annual Growth of Two Commercial Ommastrephid Species in the Pacific Ocean. Fishes, 7(5), 280. https://doi.org/10.3390/fishes7050280