
Multi-Parameter Characterization of Disease-Suppressive Bio-composts from Aromatic Plant Residues Evaluated for Garden Cress (Lepidium sativum L.) Cultivation

 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bio-composts
2.2. In Planta Compost Suppressiveness Assay
- The eleven bio-composts, both sterile (twice autoclaved) and not sterile, were supplied at a rate of 30% (vol.) to a standard peat-based growing medium. Non-amended peat was used as a control.
- Isolates of Rhizoctonia solani (AG-4) and Sclerotinia sclerotiorum from the CREA microbial collection (at the Research Center for Vegetables and Ornamental Crops in Pontecagnano Faiano, Italy), maintained on a potato dextrose agar medium (PDA, Oxoid Ltd., Basingstoke, UK), were grown for 21 days on common millet seeds saturated with potato dextrose broth (PDB, Oxoid Ltd., Basingstoke, UK) (1/10 w/w) to prepare the pathogen inoculum to be incorporated into the substrate at a final concentration of 1% (w/w, dry weight).
- The experimental unit consisted of a mini plastic pot (7 cm diam., ~0.1 L vol.) filled with the different substrates, sown with 20 seeds of garden cress cv. Comune (Blumen, Milan, Italy) each and replicated five times for each treatment. Ultimately, there were 11 composts ×2 conditions (autoclaved and non-autoclaved) +2 controls (healthy and infected peat) for a total of 24 treatments, resulting in 120 pots and 2400 seeds. The experiment was repeated.
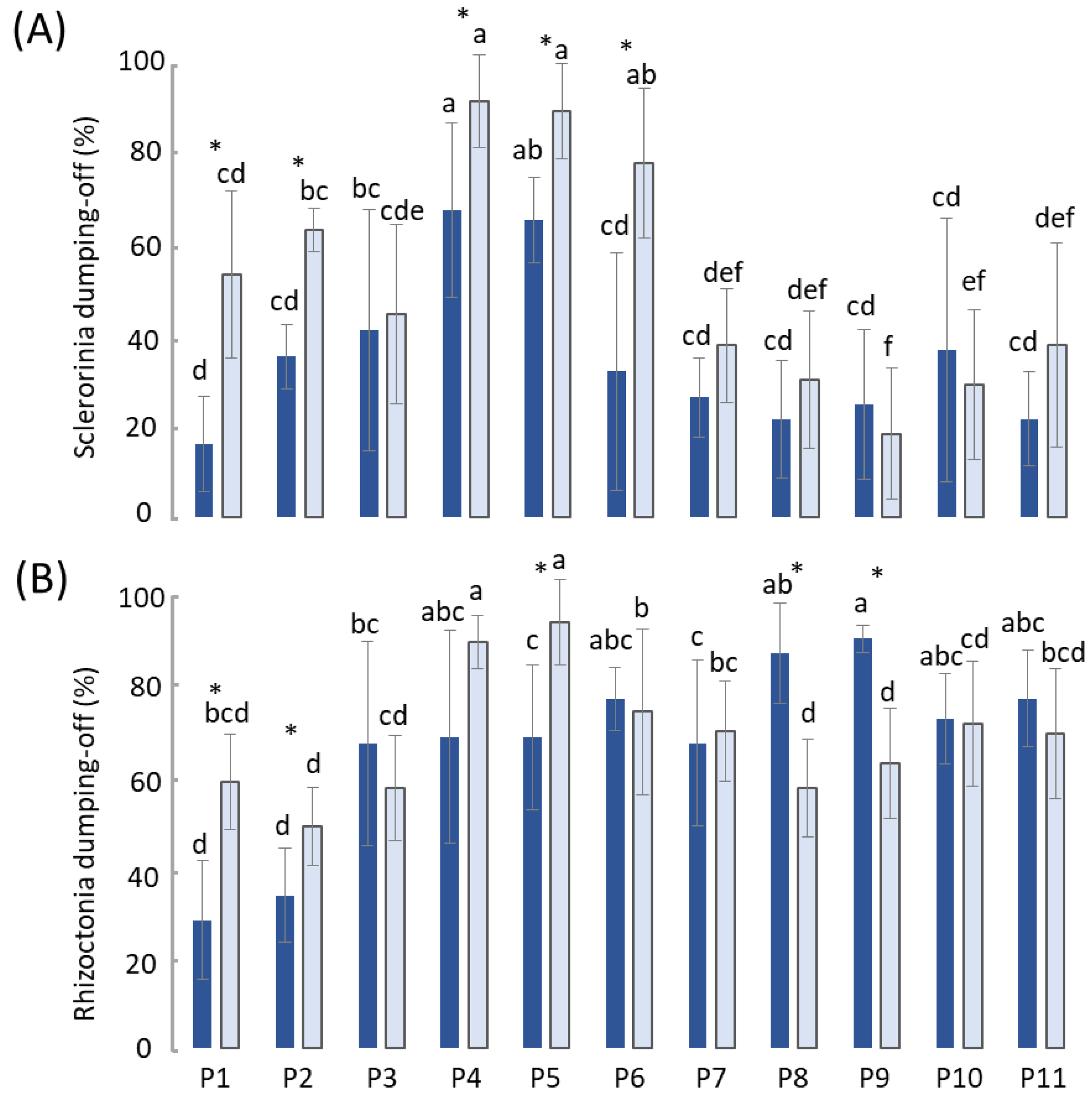
- The sown pots were placed in a climate chamber (25 °C) for 7 days to allow for the emergence of cress seedlings and the development of Sclerotinia and Rhizoctonia damping off (Figure 1).
2.3. Analysis of the Main Physico-Chemical and Biological Components of Compost
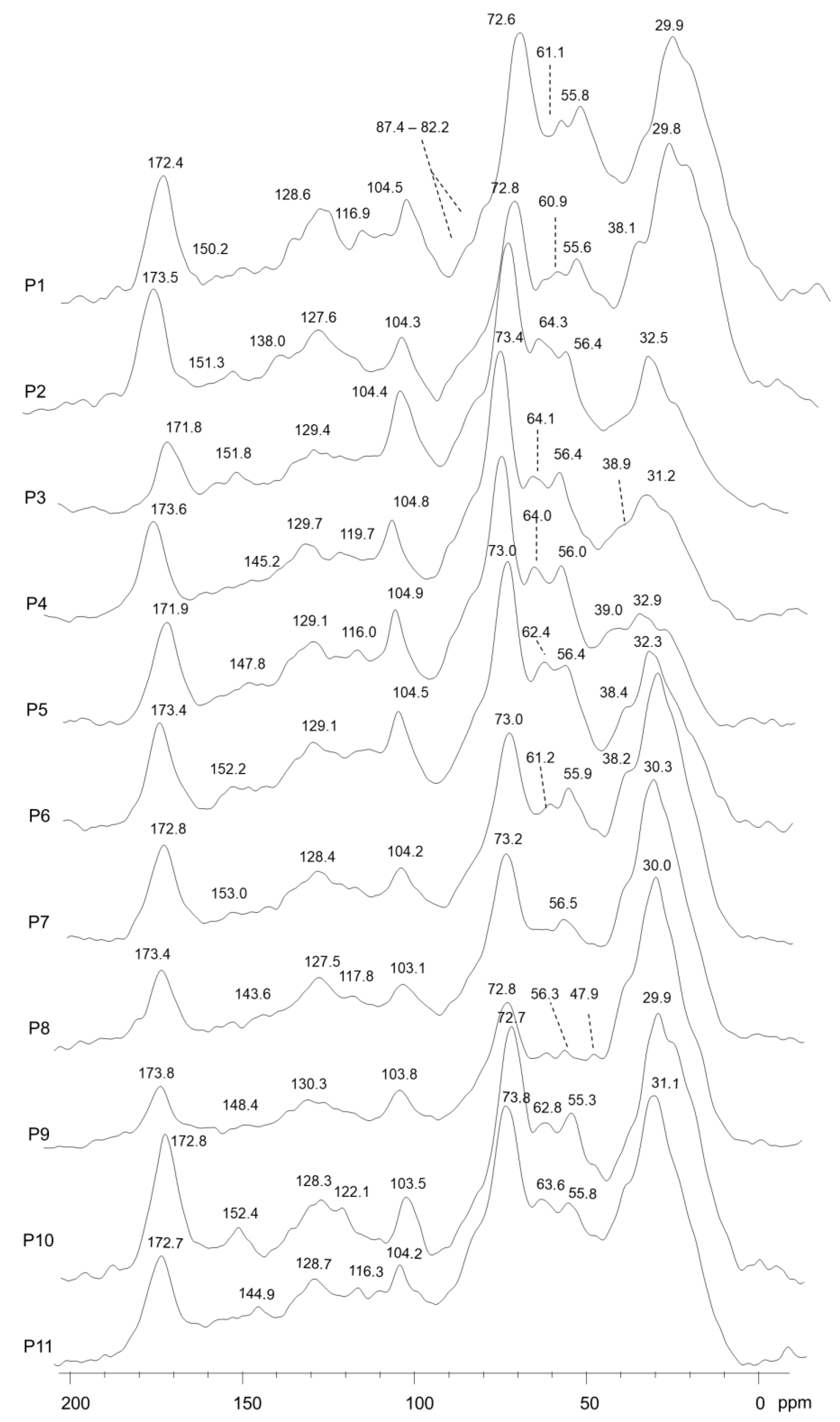
2.4. Analysis of the Molecular Carbon Components of Compost
2.5. Statistical Analysis
3. Results
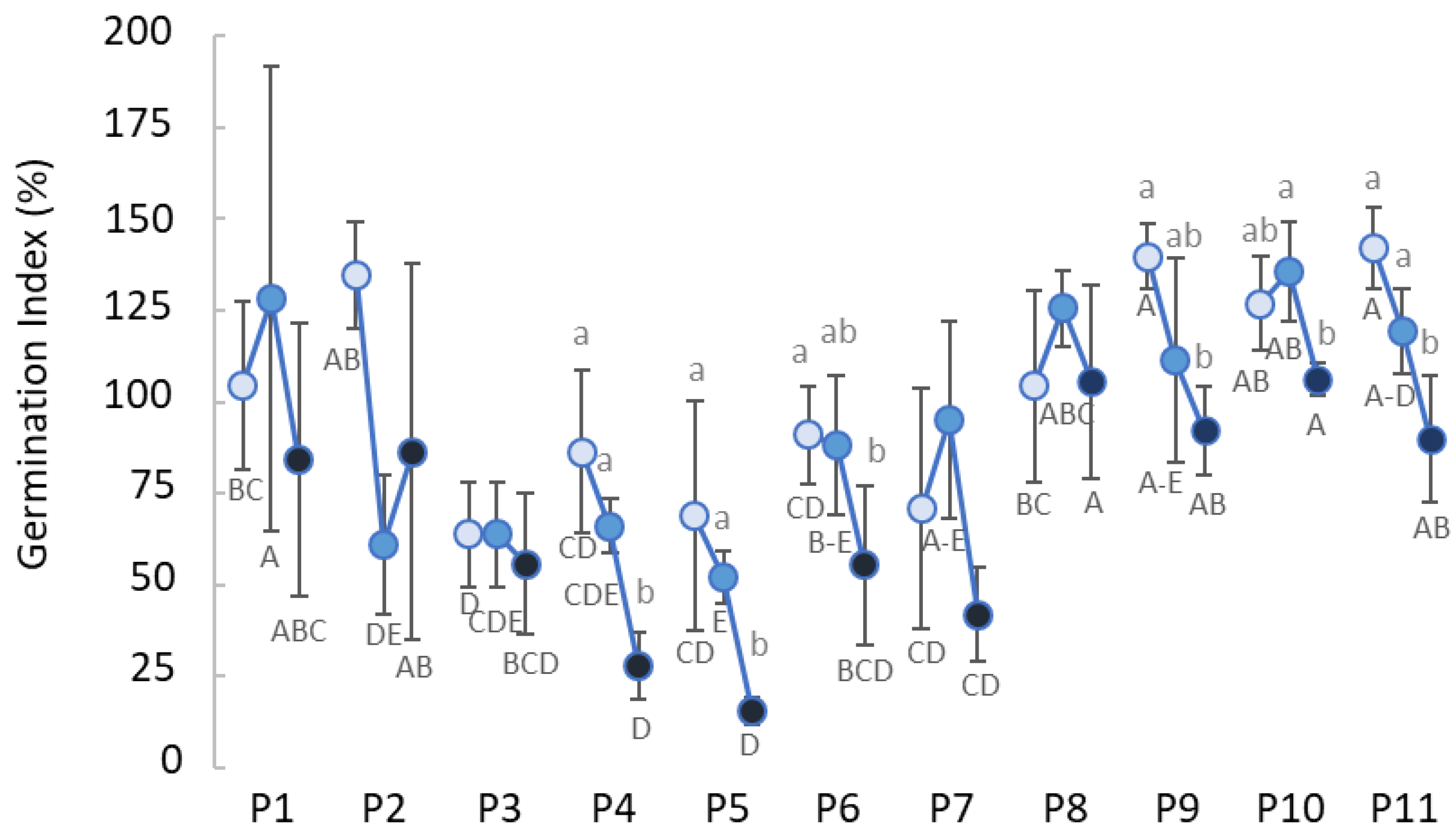
3.1. Compost Suppressiveness
3.2. Phyitotoxicity and Biological Properties of Composts
3.3. Chemical and Molecular Properties of the Composts
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nicola, S.; Hoeberechts, J.; Fontana, E. Ebb-and-flow and floating systems to grow leafy vegetables: A review for rocket, corn salad, garden cress and purslane. Acta Hortic. 2007, 747, 585–593. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G.; De Pascale, S. Sprouts, microgreens and edible flowers as novel functional foods. Agronomy 2021, 11, 2568. [Google Scholar] [CrossRef]
- Santos, J.; Oliveira, M.B.P.P.; Ibáñez, E.; Herrero, M. Phenolic profile evolution of different ready-to-eat baby-leaf vegetables during storage. J. Chromatog. A 2014, 1327, 118–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, T.; Grover, K.; Grewal, I.S. Development and sensory evaluation of ready to eat supplementary food using garden cress (Lepidium sativum) seeds. J. Appl. Nat. Sci. 2016, 8, 1501–1506. [Google Scholar] [CrossRef]
- Ciesielska, K.; Ciesielski, W.; Kulawik, D.; Oszczęda, Z.; Tomasik, P. Cultivation of cress involving water treated under different atmospheres with low-temperature, low-pressure glow plasma of low frequency. Water 2020, 12, 2152. [Google Scholar] [CrossRef]
- Keutgen, N.; Hausknecht, M.; Tomaszewska-Sowa, M.; Keutgen, A.J. Nutritional and sensory quality of two types of cress microgreens depending on the mineral nutrition. Agronomy 2021, 11, 1110. [Google Scholar] [CrossRef]
- Ogórek, R. Enzymatic activity of potential fungal plant pathogens and the effect of their culture filtrates on seed germination and seedling growth of garden cress (Lepidium sativum L.). Eur. J. Plant Pathol. 2016, 145, 469–481. [Google Scholar] [CrossRef]
- Mehta, C.M.; Palni, U.; Franke-Whittle, I.H.; Sharma, A.K. Compost: Its role, mechanism and impact on reducing soil-borne plant diseases. Waste Manag. 2014, 34, 607–622. [Google Scholar] [CrossRef]
- Pane, C.; Zaccardelli, M. Principles of compost-based plant diseases control and innovative new developments. In Composting for Sustainable Agriculture; Maheshwari, D., Ed.; Sustainable Development and Biodiversity; Springer: Cham, Switzerland, 2014; Volume 3. [Google Scholar] [CrossRef]
- Giménez, A.; Fernández, J.A.; Egea-Gilabert, C.; Santísima-Trinidad, A.B.; Ros, M.; Pascual, J.A. Agro-industry composts as growing medium for growing baby-leaf lettuces in a floating system—Added-value to suppress Pythium irregulare. Acta Hortic. 2019, 1242, 791–798. [Google Scholar] [CrossRef]
- Morau, A.; Piepho, H.-P.; Fritz, J. Growth responses of garden cress (Lepidium sativum L.) to biodynamic cow manure preparation in a bioassay. Biol. Agric. Hortic. 2020, 36, 16–34. [Google Scholar] [CrossRef] [Green Version]
- Pane, C.; Piccolo, A.; Spaccini, R.; Celano, G.; Villecco, D.; Zaccardelli, M. Agricultural waste-based composts exhibiting suppressivity to diseases caused by the phytopathogenic soil-borne fungi Rhizoctonia solani and Sclerotinia minor. Appl. Soil Ecol. 2013, 65, 43–51. [Google Scholar] [CrossRef]
- Pane, C.; Spaccini, R.; Piccolo, A.; Scala, F.; Bonanomi, G. Compost amendments enhance peat suppressiveness to Pythium ultimum, Rhizoctonia solani and Sclerotinia minor. Biol. Control 2011, 56, 115–124. [Google Scholar] [CrossRef]
- Aslam, D.N.; Horwath, W.; Vander Gheynst, J.S. Comparison of several maturity indicators for estimating phytotoxicity in compost-amended soil. Waste Manag. 2008, 28, 2070–2076. [Google Scholar] [CrossRef] [PubMed]
- Ventorino, V.; Parillo, R.; Testa, A.; Viscardi, S.; Espresso, F.; Pepe, O. Chestnut green waste composting for sustainable forest management: Microbiota dynamics and impact on plant disease control. J. Environ. Manag. 2016, 166, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Ronga, D.; Francia, E.; Allesina, G.; Pedrazzi, S.; Zaccardelli, M.; Pane, C.; Tava, A.; Bignami, C. Valorization of vineyard by-products to obtain composted digestate and biochar suitable for nursery grapevine (Vitis vinifera L.) production. Agronomy 2019, 9, 420. [Google Scholar] [CrossRef] [Green Version]
- Pane, C.; Spaccini, R.; Piccolo, A.; Celano, G.; Zaccardelli, M. Disease suppressiveness of agricultural greenwaste composts as related to chemical and bio-based properties shaped by different on-farm composting methods. Biol. Control 2019, 137, 104026. [Google Scholar] [CrossRef]
- Pane, C.; Sorrentino, R.; Scotti, R.; Molisso, M.; Di Matteo, A.; Celano, G.; Zaccardelli, M. Alpha and beta-diversity of microbial communities associated to plant disease suppressive functions of on-farm green composts. Agriculture 2020, 10, 113. [Google Scholar] [CrossRef] [Green Version]
- Bignami, C.; Melegari, F.; Zaccardelli, M.; Pane, C.; Ronga, D. Composted solid digestate and vineyard winter prunings partially replace peat in growing substrates for micropropagated highbush blueberry in the nursery. Agronomy 2022, 12, 337. [Google Scholar] [CrossRef]
- De Corato, U. Agricultural waste recycling in horticultural intensive farming systems by on-farm composting and compost-based tea application improves soil quality and plant health: A review under the perspective of a circular economy. Sci. Tot. Environ. 2020, 738, 139840. [Google Scholar] [CrossRef]
- Zaccardelli, M.; Roscigno, G.; Pane, C.; Celano, G.; Di Matteo, M.; Mainente, M.; Vuotto, A.; Mencherini, T.; Esposito, T.; Vitti, A.; et al. Essential oils and quality composts sourced by recycling vegetable residues from the aromatic plant supply chain. Ind. Crop. Prod. 2021, 162, 113255. [Google Scholar] [CrossRef]
- Zaccardelli, M.; Pane, C.; Caputo, M.; Durazzo, A.; Lucarini, M.; Silva, A.M.; Severino, P.; Souto, E.B.; Santini, A.; De Feo, V. Sage species case study on a spontaneous Mediterranean plant to control phytopathogenic fungi and bacteria. Forests 2020, 11, 704. [Google Scholar] [CrossRef]
- Violante, P. Metodi di Analisi Chimica del Suolo; Angeli, F., Ed.; Italian Ministry of Agriculture: Milan, Italy, 2000; p. 536. [Google Scholar]
- Scher, F.M.; Baker, R. Effect of Pseudomonas putida and a synthetic iron chelator on induction of soil suppressiveness to Fusarium Wilt pathogens. Phytopathology 1982, 72, 1567–1573. [Google Scholar] [CrossRef]
- Sadfi, N.; Cherif, M.; Fliss, I.; Boudabbous, A.; Antoun, H. Evaluation of bacterial isolates from salty soils and Bacillus thuringiensis strains for the biocontrol of Fusarium dry rot of potato tubers. J. Plant Pathol. 2001, 83, 101–118. [Google Scholar]
- Mestrelab Research, MestReNova Manual 2010. Manual of MestreNova 6.2. Available online: https://mestrelab.com/ (accessed on 19 June 2022).
- Verrillo, M.; Cozzolino, V.; Spaccini, R.; Piccolo, A. Humic substances from green compost increase bioactivity and antibacterial properties of essential oils in Basil leaves. Chem. Biol. Technol. Agric. 2021, 8, 28. [Google Scholar] [CrossRef]
- Bento, L.R.; Spaccini, R.; Cangemi, S.; Mazzei, P.; de Freitas, B.B.; de Souza, A.E.O.; Moreira, A.B.; Ferreira, O.P.; Piccolo, A.; Bisinoti, M.C. Hydrochar obtained with by-products from the sugarcane industry: Molecular features and effects of extracts on maize seed germination. J. Environ. Manag. 2021, 281, 111878. [Google Scholar] [CrossRef] [PubMed]
- Fregolente, L.G.; Dos Santos, J.V.; Vinci, G.; Piccolo, A.; Moreira, A.B.; Ferreira, O.P.; Bisinoti, M.C.; Spaccini, R. Insights on molecular characteristics of hydrochars by 13C-NMR and off-line TMAH-GC/MS and assessment of their potential use as plant growth promoters. Molecules 2021, 26, 1026. [Google Scholar] [CrossRef]
- de Aquino, A.M.; Canellas, L.P.; da Silva, A.P.S.; Canellas, N.O.; da S Lima, L.; Olivares, F.L.; Piccolo, A.; Spaccini, R. Evaluation of molecular properties of humic acids from vermicompost by 13C-CPMAS-NMR spectroscopy and thermochemolysis–GC–MS. J. Anal. Appl. Pyrol. 2019, 141, 104634. [Google Scholar] [CrossRef]
- Yang, F.; Qi, Y.; Liu, W.; Li, J.; Wang, D.; Fang, L.; Zhang, Y. Separation of five flavonoids from aerial parts of Salvia Miltiorrhiza bunge using HSCCC and their antioxidant activities. Molecules 2019, 24, 3448. [Google Scholar] [CrossRef] [Green Version]
- Kadir, A.; Zheng, G.; Zheng, X.; Jin, P.; Maiwulanjiang, M.; Gao, B.; Aisa, H.A.; Yao, G. Structurally diverse diterpenoids from the roots of Salvia deserta based on nine different skeletal types. J. Nat. Prod. 2021, 84, 1442–1452. [Google Scholar] [CrossRef]
- Aydin, S.K.; Ertaş, A.; Boğa, M.; Erol, E.; Toraman, G.O.A.; Saygı, T.K.; Halfon, B.; Topçu, G. Di-, and triterpenoids isolation and lc-ms analysis of salvia marashica extracts with bioactivity studies. Rec. Nat. Prod. 2021, 15, 463–475. [Google Scholar] [CrossRef]
- Martinez-Balmori, D.; Spaccini, R.; Aguiar, N.O.; Novotny, E.H.; Olivares, F.L.; Canellas, L.P. Molecular characteristics of humic acids isolated from vermicomposts and their relationship to bioactivity. J. Agr. Food Chem. 2014, 62, 11412–11419. [Google Scholar] [CrossRef] [PubMed]
- Bustos-Brito, C.; Joseph-Nathan, P.; Burgueño-Tapia, E.; Martínez-Otero, D.; Nieto-Camacho, A.; Calzada, F.; Yépez-Mulia, L.; Esquivel, B.; Quijano, L. Structure and absolute configuration of abietane diterpenoids from Salvia clinopodioides: Antioxidant, antiprotozoal, and antipropulsive activities. J. Nat. Prod. 2019, 82, 1207–1216. [Google Scholar] [CrossRef] [PubMed]
- Yaris, E.; Balur Adsız, L.; Yener, I.; Tuncay, E.; Yilmaz, M.A.; Akdeniz, M.; Kaplaner, E.; First, M.; Ertas, A.; Kolak, U. Isolation of secondary metabolites of two endemic species: Salvia rosifolia Sm. and Salvia cerino-pruinosa Rech. f. var. elazigensis (Lamiaceae). J. Food Meas. Charact. 2021, 15, 4929–4938. [Google Scholar] [CrossRef]
- Slimestad, R.; Fossen, T.; Brede, C. Flavonoids and other phenolics in herbs commonly used in Norwegian commercial kitchens. Food Chem. 2020, 309, 125678. [Google Scholar] [CrossRef]
- Gajdoš, R. Effects of two composts and seven commercial cultivation media on germination and yield. Compost Sci. Util. 1997, 5, 16–37. [Google Scholar] [CrossRef]
- Shao-qi, Z.; Wei-dong, L.; Xiao, Z. Effects of heavy metals on planting watercress in kailyard soil amended by adding compost of sewage sludge. Process Saf. Environ. 2010, 88, 263–268. [Google Scholar]
- Tuncay, Ö.; Esiyok, D.; Yamur, B.; Okur, B. Yield and quality of garden cress affected by different nitrogen sources and growing period. Afr. J. Agric. Res. 2011, 6, 608–617. [Google Scholar]
- Pauli, A. Antimicrobial properties of essential oil constituents. Int. J. Arom. 2001, 11, 126–133. [Google Scholar] [CrossRef]
- De Almeida, L.F.R.; Frei, F.; Mancini, E.; De Martino, L.; De Feo, V. Phytotoxic activities of mediterranean essential oils. Molecules 2010, 15, 4309–4323. [Google Scholar] [CrossRef] [Green Version]
- Greff, B.; Lakatos, E.; Szigeti, J.; Varga, L. Co-composting with herbal wastes: Potential effects of essential oil residues on microbial pathogens during composting. Crit. Rev. Environ. Sci. Technol. 2021, 51, 457–511. [Google Scholar] [CrossRef]
- Pane, C.; Villecco, D.; Campanile, F.; Zaccardelli, M. Novel strains of Bacillus, isolated from compost and compost-amended soils, as biological control agents against soil-borne phytopathogenic fungi. Biocontrol Sci. Technol. 2012, 22, 1373–1388. [Google Scholar] [CrossRef]
- Escriche, I.; Kadar, M.; Juan-Borrás, M.; Domenech, E. Suitability of antioxidant capacity, flavonoids and phenolic acids for floral authentication of honey. Impact of industrial thermal treatment. Food Chem. 2014, 142, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Doudin, K.; Al-Malaika, S.; Sheena, H.H.; Tverezovskiy, V.; Fowler, P. New genre of antioxidants from renewable natural resources: Synthesis and characterisation of rosemary plant-derived antioxidants and their performance in polyolefins. Polym. Degrad. Stab. 2016, 130, 126–134. [Google Scholar] [CrossRef] [Green Version]
- Palese, A.M.; Pane, C.; Villecco, D.; Zaccardelli, M.; Altieri, G.; Celano, G. Effects of organic additives on chemical, microbiological and plant pathogen suppressive properties of aerated municipal waste compost teas. Appl. Sci. 2021, 11, 7402. [Google Scholar] [CrossRef]
- Scaglia, B.; Nunes, R.R.; Rezende, M.O.O.; Tambone, F.; Adani, F. Investigating organic molecules responsible of auxin-like activity of humic acid fraction extracted from vermicompost. Sci. Tot. Environ. 2016, 562, 289–295. [Google Scholar] [CrossRef]
- Castaño, R.; Borrero, C.; Avilés, M. Organic matter fractions by SP-MAS 13C NMR and microbial communities involved in the suppression of Fusarium wilt in organic growth media. Biol. Cont. 2011, 58, 286–293. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compost Source | Turning | Pathogen | Control (%) | Reference |
|---|---|---|---|---|
| Viticulture and enological factory residues | Pythium ultimum | 23 * | [13] | |
| Rhizoctonia solani | 42 * | |||
| Sclerotinia minor | 0 | |||
| Organic fraction of differentiated municipal bio-waste | P. ultimum | 53 * | [13] | |
| R. solani | 0 | |||
| S. minor | 54 * | |||
| Organic fraction of undifferentiated municipal bio-waste | P. ultimum | 26 * | [13] | |
| R. solani | 0 | |||
| S. minor | 38 * | |||
| Cow manure | P. ultimum | 47 * | [13] | |
| R. solani | 60 * | |||
| S. minor | 36 * | |||
| Organic fraction of differentiated biowaste + peat 50% v/v | P. ultimum | 53 * | [13] | |
| R. solani | 0 | |||
| S. minor | 19 ns | |||
| Tomato, escarole, woodchip, compost 17.5:15.5:65:2 w/w | FV | R. solani | 77 * | [12] |
| S. minor | 61 * | |||
| Tomato, woodchip, compost 50:48:2 w/w | FV | R. solani | 55 * | [12] |
| S. minor | 29 ns | |||
| Artichoke, woodchip, compost 78:20:2 w/w | FV | R. solani | 46 * | [12] |
| S. minor | 48 * | |||
| Artichoke, fennel, escarole, woodchip, compost 43.5:23.5:11:20:2 w/w | FV | R. solani | 39 * | [12] |
| S. minor | 36 ns | |||
| Urban waste | R. solani | 0 | [12] | |
| S. minor | 0 | |||
| Chestnut leaves, branches, bark, and hulls | R. solani | 45 * | [15] | |
| S. minor | 62 * | |||
| Solid digestate | R. solani | 1 ns | [16] | |
| S. minor | 53 * | |||
| Escarole, crop cardoon, compost 80:18:2 w/w | PA | R. solani | 36 * | [17] |
| S. minor | 66 * | |||
| Escarole, crop cardoon, compost 80:18:2 w/w | FV | R. solani | 42 * | [17] |
| S. minor | 69 * | |||
| Escarole, crop cardoon, compost 80:18:2 w/w | TH | R. solani | 15 ns | [17] |
| S. minor | 32 * | |||
| Leafy vegetables, fennel, woodchip | MT→FV | R. solani | 56 * | [18] |
| S. minor | 84 * | |||
| Maize, livestock waste, woodchip | MT→FV | R. solani | 0 | [18] |
| S. minor | 2 ns | |||
| Leafy vegetables, basil, tomato, watermelon, woodchip | MT→FV | R. solani | 39 * | [18] |
| S. minor | 65 * | |||
| Leafy vegetables, basil, watermelon, woodchip | MT→FV | R. solani | 36 * | [18] |
| S. minor | 95 * | |||
| Leafy vegetables, basil, pumpkin, woodchip | MT→FV | R. solani | 0 | [18] |
| S. minor | 11 ns | |||
| Leafy vegetables, basil, woodchip | MT→FV | R. solani | 0 | [18] |
| S. minor | 36 ns | |||
| Leafy vegetables, basil, watermelon, woodchip | MT→FV | R. solani | 43 * | [18] |
| S. minor | 18 ns | |||
| Leafy vegetables, basil, woodchip | MT→FV | R. solani | 48 * | [18] |
| S. minor | 95 * | |||
| Leafy vegetables, basil, woodchip | MT→FV | R. solani | 57 * | [18] |
| S. minor | 0 | |||
| Leafy vegetables, basil, pumpkin, woodchip | MT→FV | R. solani | 0 | [18] |
| S. minor | 72 * | |||
| Leafy vegetables, artichoke, woodchip | MT→FV | R. solani | 52 * | [18] |
| S. minor | 60 * | |||
| Leafy vegetables, cabbage, walnut husk, woodchip | MT→FV | R. solani | 40 * | [18] |
| S. minor | 72 * | |||
| Leafy vegetables, basil, sorghum, tomato, pumpkin, woodchip | MT→FV | R. solani | 45 * | [18] |
| S. minor | 86 * | |||
| Solid digestate + chips 15:83.3 w/w | R. solani | 12 ns | [19] | |
| S. minor | 59 * |
| Compost | Feedstock | |||
|---|---|---|---|---|
| Wood Chips | Vegetable Residues | Aromatic Plant Material | ||
| P1 | 40% | 30% Escarole (Cichorium endivia L.) | 30% Mixture of Sage (Salvia officinalis L.), Basil (Ocimum basilicum L.), Mint (Mentha x piperita L.) and Parsley (Petroselinum crispum (Mill.) Fuss) | |
| P2 | 40% | 30% Escarole | 30% Mixture of Sage, Basil and Rosemary (Rosmarinus officinalis L.) | Distilled |
| P3 | 20% | 50.5% Parsley 6.2% Rocket (Diplotaxis tenuifolia L.) 3.6% Red radish (Raphanus sativus L.) | 29.9% Basil, 6.2% Thyme (Thymus vulgaris L.), 2% Laurel (Laurus nobilis L.), 1.6% Mint | Distilled |
| P4 | - | 100% Basil | ||
| P5 | - | 100% Basil | Distilled | |
| P6 | 20% | 50% Parsley, 28% Thyme, 8.1% Rosemary, 5.5% Mint, 3.8% Oregano (Origanum vulgare L.), 2.5% Sage, 0.9% Laurel, 0.7% Tarragon (Artemisia dracunculus L.), 0.5% Basil | ||
| P7 | 20% | 17.6% Parsley | 39.3% Basil, 19.7% Rosemary, 16.5% Sage, 6.9% Mint | Distilled |
| P8 | - | 100% Rosemary | ||
| P9 | - | 100% Rosemary | Distilled | |
| P10 | - | 100% Sage | ||
| P11 | - | 100% Sage | Distilled | |
| Compost | Fluoresceine-Diacetate Hydrolysis (µg FDA g−12h−1) | Basal Respiration (μL CO2 s−1 g−1) | Total Fungi (Log CFU g−1) | Total Bacteria (LogCFU g−1) | Thermal-Resistant Bacteria (LogCFU g−1) | Pseudomonas-Like Bacteria (LogCFU g−1) | EC | pH | NO3− (ppm) |
|---|---|---|---|---|---|---|---|---|---|
| P1 | 8.22 ± 0.90 b | 2.73 ± 1.85 | 5.27 ± 0.12 d | 7.68 ± 0.05 bcd | 7.51 ± 0.49 a | 7.95 ± 0.00 cd | 8434.00 b | 8.52 d | 5.87 ± 0.50 def |
| P2 | 8.59 ± 0.33 b | 1.01 ± 0.18 | 5.61 ± 0.06 c | 7.34 ± 0.12 d | 7.26 ± 0.00 ab | 8.30 ± 0.49 bc | 4612.00 e | 9.00 bc | 8.60 ± 0.53 c |
| P3 | 7.03 ± 0.11 c | 1.06 ± 0.00 | 3.56 ± 0.00 g | 7.62 ± 0.31 cd | 6.54 ± 0.27 cd | 7.26 ± 0.30 e | 5123.00 d | 10.23 a | 7.13 ± 1.21 cde |
| P4 | 9.40 ± 0.25 a | 0.77 ± 0.00 | 5.31 ± 0.05 d | 7.61 ± 0.16 cd | 7.10 ± 0.21 ab | 6.75 ± 0.18 f | 11837.00 a | 8.57 d | 5.60 ± 1.93 ef |
| P5 | 8.73 ± 0.13 b | 1.21 ± 1.24 | 5.25 ± 0.09 d | 7.88 ± 0.24 bc | 6.80 ± 0.06 bc | 7.76 ± 0.13 d | 7258.67 c | 10.30 a | 4.80 ± 0.60 f |
| P6 | 8.81 ± 0.36 ab | 0.94 ± 0.00 | 4.86 ± 0.00 e | 7.36 ± 0.17 d | 6.10 ± 0.21 de | 7.27 ± 0.28 e | 4331.00 f | 9.40 b | 13.07 ± 1.10 a |
| P7 | 8.41 ± 0.19 b | 2.26 ± 0.00 | 4.56 ± 0.00 f | 5.93 ± 0.16 e | 5.45 ± 0.20 g | 6.75 ± 0.10 f | 4630.00 e | 8.48 d | 7.60 ± 0.87 cd |
| P8 | 0.94 ± 0.18 de | 0.65 ± 0.27 | 6.16 ± 0.15 ab | 8.94 ± 0.32 a | 4.87 ± 0.12 h | 8.79 ± 0.21 a | 2282.00 h | 8.85 cd | 11.07 ± 2.34 b |
| P9 | 0.54 ± 0.26 e | 0.58 ± 0.34a | 6.01 ± 0.00 b | 7.57 ± 0.28 cd | 5.57 ± 0.15 fg | 9.01 ± 0.00 a | 8.99 i | 8.94 c | 2.13 ± 0.61 g |
| P10 | 1.28 ± 0.29 d | 0.37 ± 0.43 | 5.26 ± 0.21 d | 7.86 ± 0.13 bc | 5.42 ± 0.30 g | 8.73 ± 0.00 ab | 3646.00 g | 9.16 bc | 7.47 ± 0.42 cde |
| P11 | 1.32 ± 0.27 d | 1.00 ± 0.22 | 6.35 ± 0.09 a | 7.98 ± 0.13 b | 5.88 ± 0.04 ef | 7.72 ± 0.32 d | 3567.67 g | 9.26 b | 12.13 ± 0.99 ab |
| Sign. | *** | ns | *** | *** | *** | *** | *** | *** | *** |
| Compost | Carboxyl-C | O-Aryl-C | Aryl-C | O-Alkyl-C | CH3O/C_N | Alkyl-C | HBa | A/OAa | LigRa |
|---|---|---|---|---|---|---|---|---|---|
| (190–160 ppm) | (160–140 ppm) | (140–110 ppm) | (110–60 ppm) | (60–45 ppm) | (45–0 ppm) | ||||
| P1 | 7.2 | 2.1 | 12.6 | 31.3 | 13.0 | 33.9 | 1.2 | 1.1 | 6.2 |
| P2 | 8.2 | 2.9 | 11.5 | 26.8 | 12.3 | 38.2 | 1.4 | 1.4 | 4.3 |
| P3 | 5.1 | 3.7 | 12.1 | 44.0 | 11.7 | 23.4 | 0.8 | 0.5 | 3.1 |
| P4 | 8.1 | 3.8 | 14.3 | 41.1 | 11.8 | 21.0 | 0.8 | 0.5 | 3.1 |
| P5 | 7.9 | 3.8 | 15.5 | 42.4 | 11.4 | 19.0 | 0.8 | 0.4 | 3.0 |
| P6 | 7.4 | 3.3 | 15.1 | 38.7 | 10.9 | 24.6 | 0.9 | 0.6 | 3.3 |
| P7 | 7.2 | 2.5 | 11.8 | 31.0 | 11.8 | 35.7 | 1.3 | 1.2 | 4.7 |
| P8 | 6.5 | 2.2 | 11.4 | 30.1 | 12.0 | 37.8 | 1.3 | 1.3 | 5.4 |
| P9 | 6.3 | 2.2 | 9.5 | 28.0 | 10.5 | 43.6 | 1.5 | 1.6 | 4.8 |
| P10 | 9.5 | 3.1 | 10.4 | 30.7 | 11.5 | 34.7 | 1.2 | 1.1 | 3.8 |
| P11 | 7.7 | 5.3 | 12.0 | 33.6 | 11.3 | 30.2 | 1.1 | 0.9 | 2.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pane, C.; Spaccini, R.; Caputo, M.; De Falco, E.; Zaccardelli, M. Multi-Parameter Characterization of Disease-Suppressive Bio-composts from Aromatic Plant Residues Evaluated for Garden Cress (Lepidium sativum L.) Cultivation. Horticulturae 2022, 8, 632. https://doi.org/10.3390/horticulturae8070632
Pane C, Spaccini R, Caputo M, De Falco E, Zaccardelli M. Multi-Parameter Characterization of Disease-Suppressive Bio-composts from Aromatic Plant Residues Evaluated for Garden Cress (Lepidium sativum L.) Cultivation. Horticulturae. 2022; 8(7):632. https://doi.org/10.3390/horticulturae8070632
Chicago/Turabian StylePane, Catello, Riccardo Spaccini, Michele Caputo, Enrica De Falco, and Massimo Zaccardelli. 2022. "Multi-Parameter Characterization of Disease-Suppressive Bio-composts from Aromatic Plant Residues Evaluated for Garden Cress (Lepidium sativum L.) Cultivation" Horticulturae 8, no. 7: 632. https://doi.org/10.3390/horticulturae8070632
APA StylePane, C., Spaccini, R., Caputo, M., De Falco, E., & Zaccardelli, M. (2022). Multi-Parameter Characterization of Disease-Suppressive Bio-composts from Aromatic Plant Residues Evaluated for Garden Cress (Lepidium sativum L.) Cultivation. Horticulturae, 8(7), 632. https://doi.org/10.3390/horticulturae8070632