
Genome-Wide Characterization of Laccase Gene Family from Turnip and Chinese Cabbage and the Role in Xylem Lignification in Hypocotyls


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Growth, Sample Collection, and Preparation
2.2. Free-Hand Section
2.3. Identification of the LAC Proteins in Turnip and Chinese Cabbage
2.4. Phylogenetic, Motif Analysis, and Gene Exon/Intron Structure Determination
2.5. Phylogenetic-Tree Construction of LACs among Different Species
2.6. Chromosome Location of LAC Genes in Turnip and Chinese Cabbage
2.7. Total RNA Isolation and Gene Expression of BrrLACs and BraLACs
2.8. Statistical Analysis
3. Results
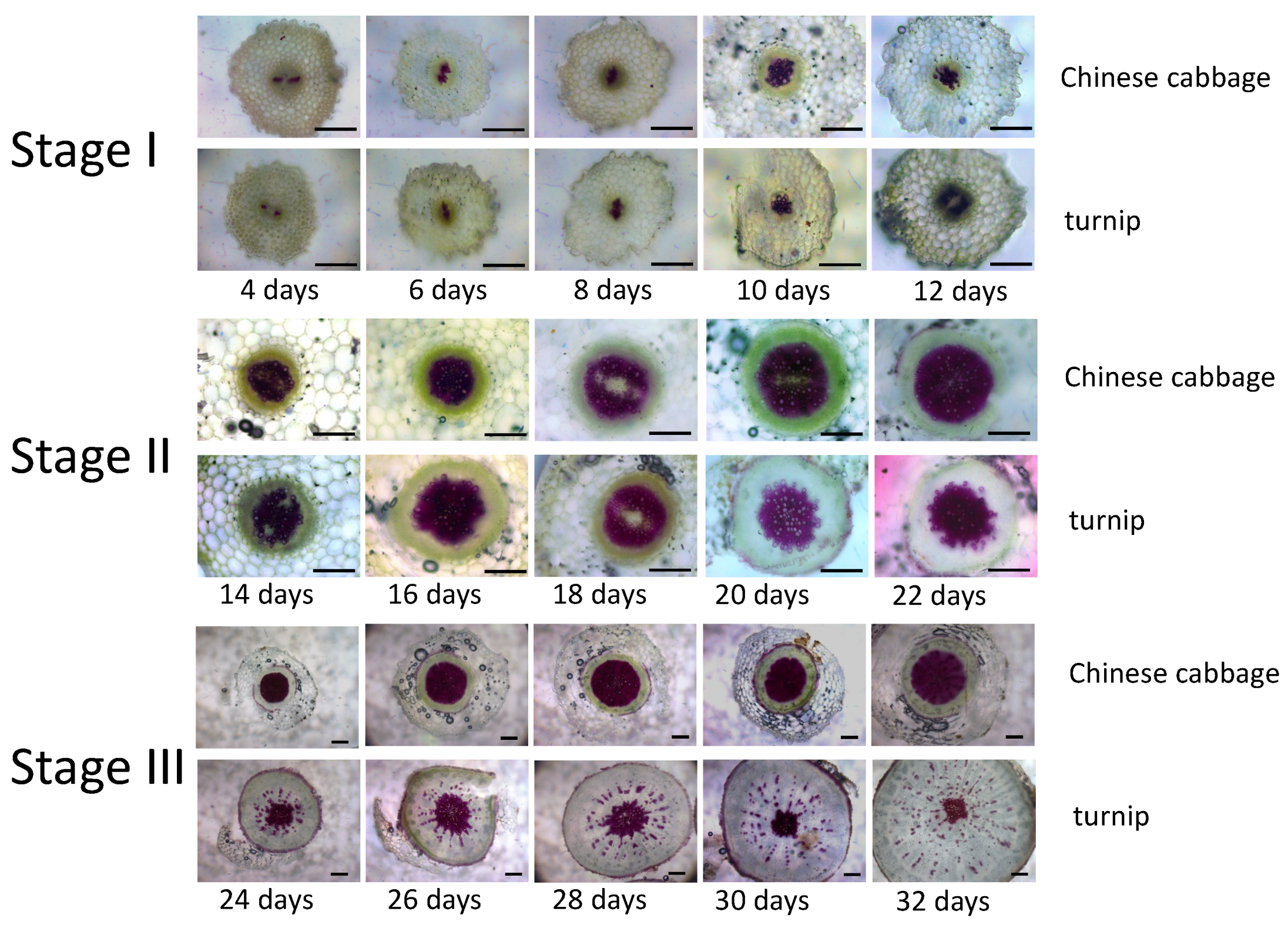
3.1. Hypocotyl Anatomical Characteristics during Early Developmental Stage in Turnip and Chinese Cabbage
3.2. Identification of LACCASE (LAC) in Turnip and Chinese Cabbage
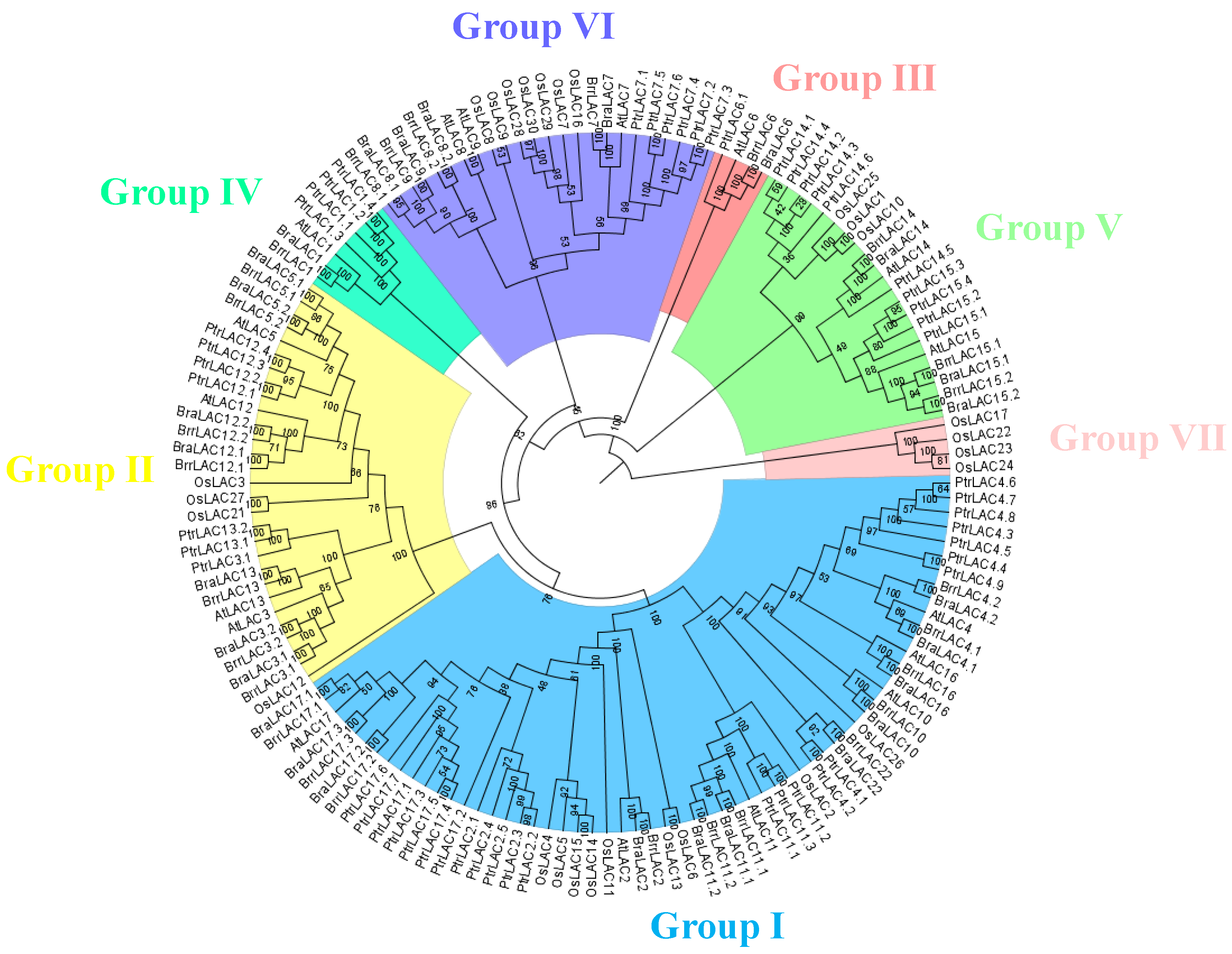
3.3. Phylogenetic Analysis
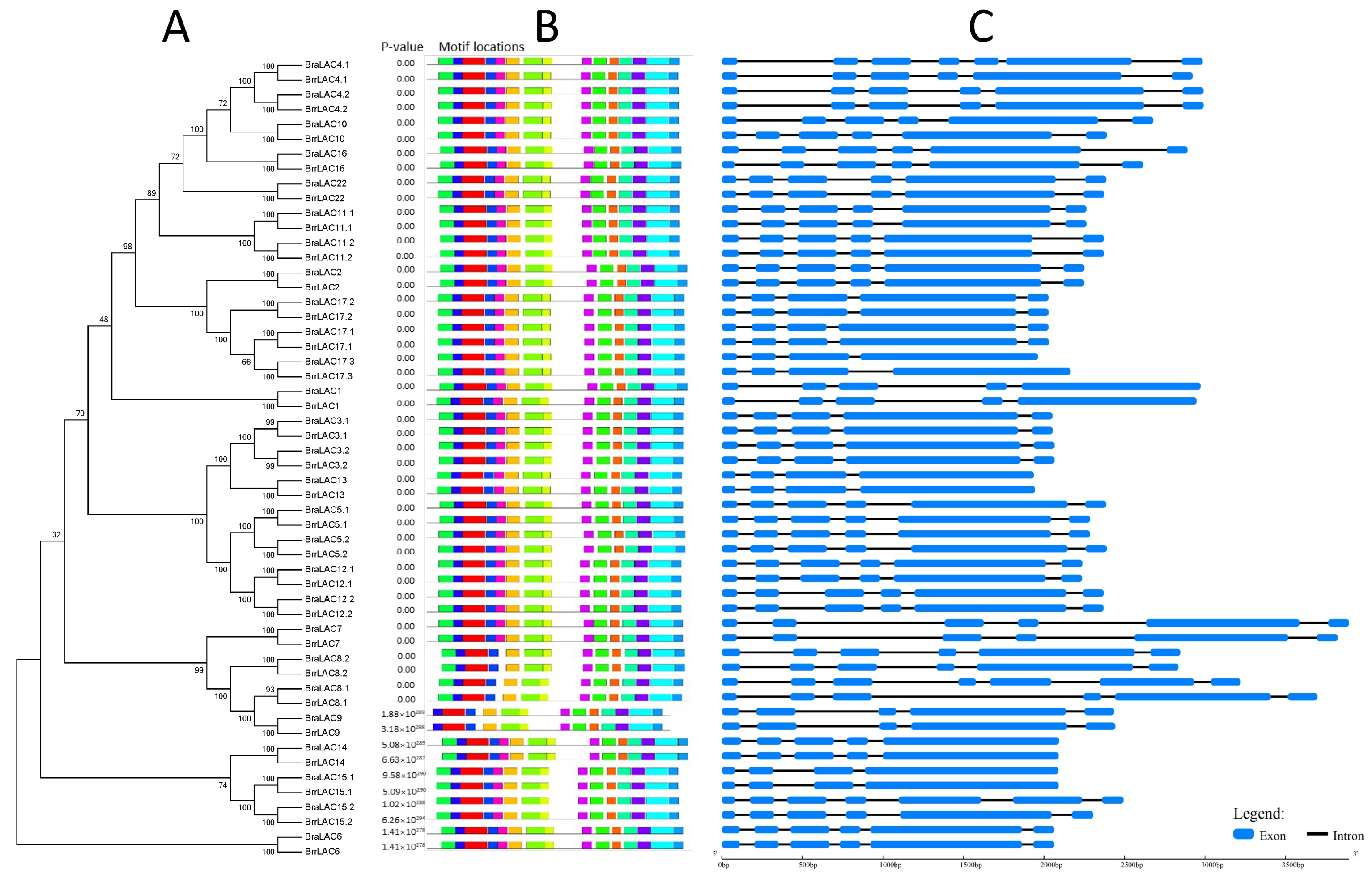
3.4. Gene Structure and Conserved Motif Analysis of LACs in Turnip and Chinese Cabbage
3.5. Chromosomal Localization
3.6. Tissue-Specific Expression Patterns of Laccase Genes in Turnip and Chinese Cabbage
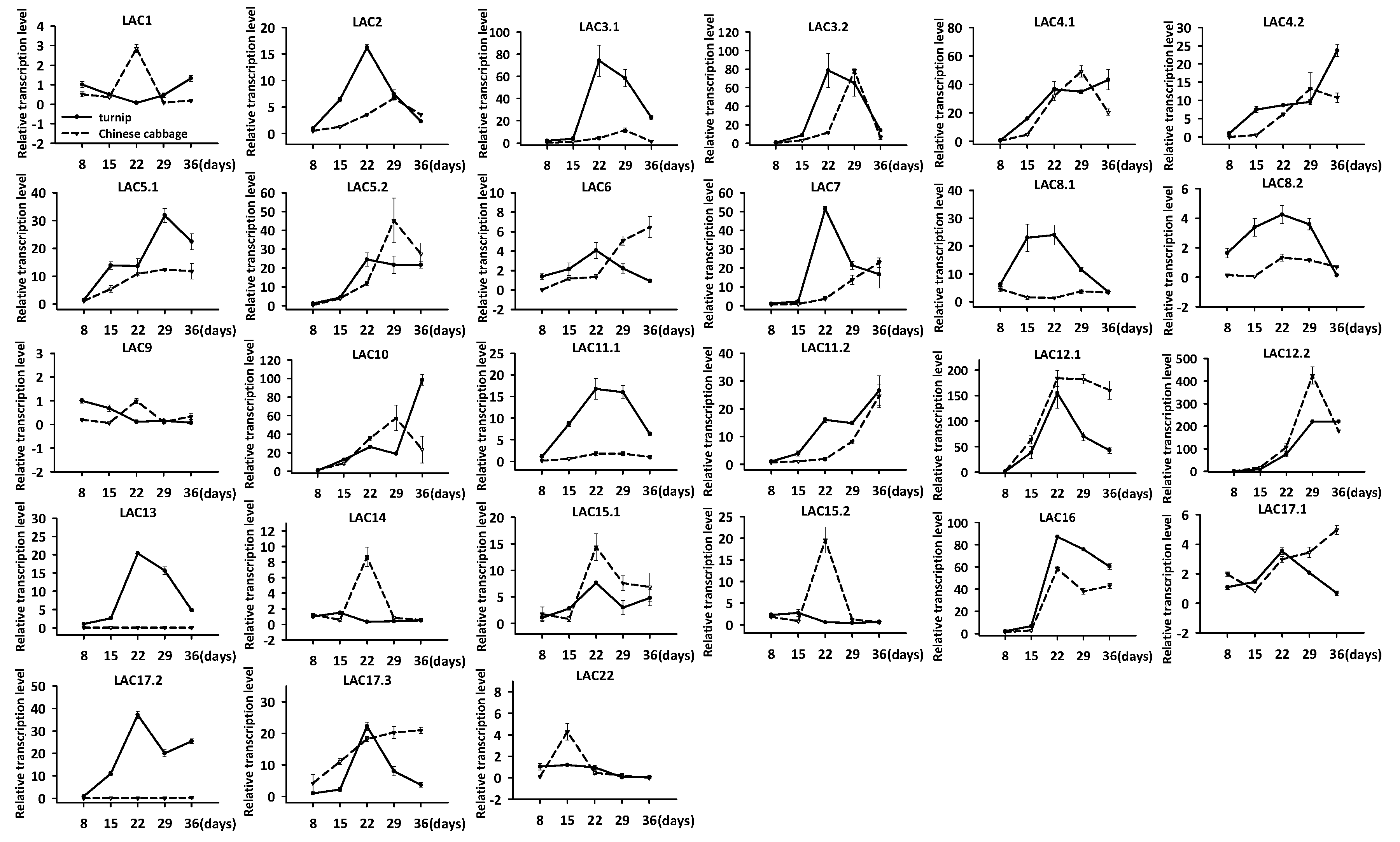
3.7. Expression Analysis of LAC Genes during Early Hypocotyl Development Stage in Turnip and Chinese Cabbage
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Nagaharu, U.; Nagaharu, N. Genome analysis in Brassica with special reference to the experimental formation of B. napus and peculiar mode of fertilization. Jpn. J. Bot. 1935, 7, 389–452. [Google Scholar]
- Labana, K.; Gupta, M. Importance and origin. In Breeding Oilseed Brassicas; Springer: Berlin/Heidelberg, Germany, 1993; pp. 1–7. [Google Scholar]
- Zierer, W.; Ruscher, D.; Sonnewald, U.; Sonnewald, S. Tuber and Tuberous Root Development. In Annual Review of Plant Biology; Merchant, S.S., Ed.; Springer: Berlin/Heidelberg, Germany, 2021; Volume 72, pp. 551–580. [Google Scholar]
- Liu, Y.Y.; Wen, J.; Ke, X.C.; Zhang, J.; Sun, X.D.; Wang, C.T.; Yang, Y.P. Gibberellin inhibition of taproot formation by modulation of DELLA-NAC complex activity in turnip (Brassica rapa var. rapa). Protoplasma 2021, 258, 925–934. [Google Scholar] [CrossRef] [PubMed]
- Diamantidis, G.; Effosse, A.; Potier, P.; Bally, R. Purification and characterization of the first bacterial laccase in the rhizospheric bacterium Azospirillum lipoferum. Soil Biol. Biochem. 2000, 32, 919–927. [Google Scholar] [CrossRef]
- Polashock, J.J.; Anagnostakis, S.L.; Milgroom, M.G.; Hillman, B.I. Isolation and Characterization of a Virus-Resistant Mutant of Cryphonectria-Parasitica. Curr. Genet. 1994, 26, 528–534. [Google Scholar] [CrossRef] [PubMed]
- Fernandez Larrea, J.; Stahl, U. Isolation and characterization of a laccase gene from Podospora anserina. Mol. Gen. Genet. 1996, 252, 539–551. [Google Scholar]
- Wahleithner, J.A.; Xu, F.; Brown, K.M.; Brown, S.H.; Golightly, E.J.; Halkier, T.; Kauppinen, S.; Pederson, A.; Schneider, P. The identification and characterization of four laccases from the plant pathogenic fungus Rhizoctonia solani. Curr. Genet. 1996, 29, 395–403. [Google Scholar] [CrossRef]
- Savinova, O.S.; Moiseenko, K.V.; Vavilova, E.A.; Chulkin, A.M.; Fedorova, T.V.; Tyazhelova, T.V.; Vasina, D.V. Evolutionary Relationships Between the Laccase Genes of Polyporales: Orthology-Based Classification of Laccase Isozymes and Functional Insight from Trametes hirsute. Front. Microbiol. 2019, 10, 152. [Google Scholar] [CrossRef] [Green Version]
- Bao, W.; O’Malley, D.M.; Whetten, R.; Sederoff, R.R. A laccase associated with lignification in loblolly pine xylem. Science 1993, 260, 672–674. [Google Scholar] [CrossRef]
- Liang, M.; Haroldsen, V.; Cai, X.; Wu, Y. Expression of a putative laccase gene, ZmLAC1, in maize primary roots under stress. Plant Cell Environ. 2006, 29, 746–753. [Google Scholar] [CrossRef]
- O’Malley, D.M.; Whetten, R.; Bao, W.; Chen, C.L.; Sederoff, R.R. The role of of laccase in lignification. Plant J. 1993, 4, 751–757. [Google Scholar] [CrossRef]
- Mayer, A.M.; Staples, R.C. Laccase: New functions for an old enzyme. Phytochemistry 2002, 60, 551–565. [Google Scholar] [CrossRef]
- Berthet, S.; Demont-Caulet, N.; Pollet, B.; Bidzinski, P.; Cezard, L.; Le Bris, P.; Borrega, N.; Herve, J.; Blondet, E.; Balzergue, S.; et al. Disruption of LACCASE4 and 17 results in tissue-specific alterations to lignification of Arabidopsis thaliana stems. Plant Cell 2011, 23, 1124–1137. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.; Nakashima, J.; Chen, F.; Yin, Y.B.; Fu, C.X.; Yun, J.F.; Shao, H.; Wang, X.Q.; Wang, Z.Y.; Dixon, R.A. LACCASE Is Necessary and Nonredundant with PEROXIDASE for Lignin Polymerization during Vascular Development in Arabidopsis. Plant Cell 2013, 25, 3976–3987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balasubramanian, V.K.; Rai, K.M.; Thu, S.W.; Hii, M.M.; Mendu, V. Genome-wide identification of multifunctional laccase gene family in cotton (Gossypium spp.); expression and biochemical analysis during fiber development. Sci. Rep. 2016, 6, 34309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Q.; Min, L.; Yang, X.; Jin, S.; Zhang, L.; Li, Y.; Ma, Y.; Qi, X.; Li, D.; Liu, H. Laccase GhLac1 modulates broad-spectrum biotic stress tolerance via manipulating phenylpropanoid pathway and jasmonic acid synthesis. Plant Physiol. 2018, 176, 1808–1823. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.Y.; Lee, C.; Hwang, S.-G.; Park, Y.C.; Lim, H.L.; Jang, C.S. Overexpression of the OsChI1 gene, encoding a putative laccase precursor, increases tolerance to drought and salinity stress in transgenic Arabidopsis. Gene 2014, 552, 98–105. [Google Scholar] [CrossRef]
- Ping, X.K.; Wang, T.Y.; Lin, N.; Di, F.F.; Li, Y.Y.; Jian, H.J.; Wang, H.; Lu, K.; Li, J.N.; Xu, X.F.; et al. Genome-Wide Identification of the LAC Gene Family and Its Expression Analysis Under Stress in Brassica napus. Molecules 2019, 24, 1985. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Li, G.; Zheng, K.J.; Zhu, X.B.; Ma, J.J.; Wang, D.M.; Tang, K.Q.; Feng, X.X.; Leng, J.T.; Yu, H.; et al. The Soybean Laccase Gene Family: Evolution and Possible Roles in Plant Defense and Stem Strength Selection. Genes 2019, 10, 701. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.Q.; Luo, L.; Wang, X.X.; Shen, Z.G.; Zheng, L.Q. Comprehensive Analysis of Rice Laccase Gene (OsLAC) Family and Ectopic Expression of OsLAC10 Enhances Tolerance to Copper Stress in Arabidopsis. Int. J. Mol. Sci. 2017, 18, 209. [Google Scholar] [CrossRef]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C-T method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.Y.; Basset, N.; Petrasch, S.; Zhang, N.W.; Bucher, J.; Shen, S.X.; Zhao, J.J.; Bonnema, G. What makes turnips: Anatomy, physiology and transcriptome during early stages of its hypocotyl-tuber development. Hortic. Res.-Engl. 2019, 6, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, N.W.; Zhao, J.J.; Lens, F.; de Visser, J.; Menamo, T.; Fang, W.; Xiao, D.; Bucher, J.; Basnet, R.K.; Lin, K.; et al. Morphology, Carbohydrate Composition and Vernalization Response in a Genetically Diverse Collection of Asian and European Turnips (Brassica rapa subsp rapa). PLoS ONE 2014, 9, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, V.; Sergeeva, L.; Ligterink, W.; Aloni, R.; Zemach, H.; Doron-Faigenboim, A.; Yang, J.; Zhang, P.; Shabtai, S.; Firon, N. Gibberellin Promotes Sweetpotato Root Vascular Lignification and Reduces Storage-Root Formation. Front. Plant Sci. 2019, 10, 1320. [Google Scholar] [CrossRef]
- Wang, G.L.; Que, F.; Xu, Z.S.; Wang, F.; Xiong, A.S. Exogenous gibberellin enhances secondary xylem development and lignification in carrot taproot. Protoplasma 2017, 254, 839–848. [Google Scholar] [CrossRef] [PubMed]
- Que, F.; Wang, G.L.; Feng, K.; Xu, Z.S.; Wang, F.; Xiong, A.S. Hypoxia enhances lignification and affects the anatomical structure in hydroponic cultivation of carrot taproot. Plant Cell Rep. 2018, 37, 1021–1032. [Google Scholar] [CrossRef]
- Wang, X.W.; Wang, H.Z.; Wang, J.; Sun, R.F.; Wu, J.; Liu, S.Y.; Bai, Y.Q.; Mun, J.H.; Bancroft, I.; Cheng, F.; et al. The genome of the mesopolyploid crop species Brassica rapa. Nat. Genet. 2011, 43, U1035–U1157. [Google Scholar] [CrossRef] [Green Version]
- Cheng, F.; Wu, J.; Wang, X.W. Genome triplication drove the diversification of Brassica plants. Hortic. Res.-Engl. 2014, 1, 14024. [Google Scholar] [CrossRef] [Green Version]
- Beilstein, M.A.; Nagalingum, N.S.; Clements, M.D.; Manchester, S.R.; Mathews, S. Dated molecular phylogenies indicate a Miocene origin for Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2010, 107, 18724–18728. [Google Scholar] [CrossRef] [Green Version]
- Sena, J.S.; Giguere, I.; Boyle, B.; Rigault, P.; Birol, I.; Zuccolo, A.; Ritland, K.; Ritland, C.; Bohlmann, J.; Jones, S.; et al. Evolution of gene structure in the conifer Picea glauca: A comparative analysis of the impact of intron size. BMC Plant Biol. 2014, 14, 95. [Google Scholar]
- Heyn, P.; Kalinka, A.T.; Tomancak, P.; Neugebauer, K.M. Introns and gene expression: Cellular constraints, transcriptional regulation, and evolutionary consequences. Bioessays 2015, 37, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Swinburne, I.A.; Miguez, D.G.; Landgraf, D.; Silver, P.A. Intron length increases oscillatory periods of gene expression in animal cells. Gene Dev. 2008, 22, 2342–2346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranocha, P.; Chabannes, M.; Chamayou, S.; Danoun, S.; Jauneau, A.; Boudet, A.M.; Goffner, D. Laccase down-regulation causes alterations in phenolic metabolism and cell wall structure in poplar. Plant Physiol. 2002, 129, 145–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, S.F.; Fan, C.F.; Li, X.H.; Li, Y.; Hu, J.; Li, C.F.; Luo, K.M. LACCASE14 is required for the deposition of guaiacyl lignin and affects cell wall digestibility in poplar. Biotechnol. Biofuels 2020, 13, 197. [Google Scholar] [CrossRef]
- Bryan, A.C.; Jawdy, S.; Gunter, L.; Gjersing, E.; Sykes, R.; Hinchee, M.A.; Winkeler, K.A.; Collins, C.M.; Engle, N.; Tschaplinski, T.J.; et al. Knockdown of a laccase in Populus deltoides confers altered cell wall chemistry and increased sugar release. Plant Biotechnol. J. 2016, 14, 2010–2020. [Google Scholar] [CrossRef]
- Liu, Y.D.; Cao, S.; Liu, X.T.; Li, Y.; Wang, B.; Sun, Y.; Zhang, C.; Guo, X.R.; Li, H.; Lu, H. PtrLAC16 plays a key role in catalyzing lignin polymerization in the xylem cell wall of Populus. Int. J. Biol. Macromol. 2021, 188, 983–992. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | CDS (bp) | Molecular Weight (kDa) | pI | GRAVY | Instability Index (%) |
|---|---|---|---|---|---|
| BrrLAC1 | 1719 | 64.4 | 9.03 | −0.268 | 31.66 |
| BrrLAC2 | 1737 | 64.2 | 9.47 | −0.091 | 35.57 |
| BrrLAC3.1 | 1713 | 64.0 | 9.46 | −0.269 | 41.82 |
| BrrLAC3.2 | 1713 | 64.0 | 9.42 | −0.249 | 40.58 |
| BrrLAC4.1 | 1677 | 61.5 | 9.36 | −0.085 | 33.09 |
| BrrLAC4.2 | 1680 | 61. 8 | 9.41 | −0.065 | 30.91 |
| BrrLAC5.1 | 1710 | 63.1 | 8.69 | −0.141 | 36.25 |
| BrrLAC5.2 | 1725 | 63.7 | 8.92 | −0.142 | 35.12 |
| BrrLAC6 | 1710 | 63.7 | 7.96 | −0.178 | 34.69 |
| BrrLAC7 | 1707 | 62.6 | 8.95 | 0.030 | 37.95 |
| BrrLAC8.1 | 1755 | 65.0 | 7.29 | −0.066 | 36.43 |
| BrrLAC8.2 | 1776 | 65.9 | 7.67 | −0.044 | 35.40 |
| BrrLAC9 | 1623 | 60.2 | 5.99 | −0.116 | 42.55 |
| BrrLAC10 | 1680 | 61.2 | 9.49 | −0.034 | 33.40 |
| BrrLAC11.1 | 1683 | 62.1 | 8.75 | −0.072 | 33.67 |
| BrrLAC11.2 | 1686 | 62.6 | 8.91 | −0.114 | 35.96 |
| BrrLAC12.1 | 1698 | 62.5 | 9.49 | −0.172 | 29.22 |
| BrrLAC12.2 | 1698 | 62.5 | 9.10 | −0.134 | 30.60 |
| BrrLAC13 | 1701 | 62.9 | 6.65 | −0.149 | 42.51 |
| BrrLAC14 | 1743 | 65.8 | 9.71 | −0.296 | 29.67 |
| BrrLAC15.1 | 1680 | 63.4 | 9.04 | −0.194 | 35.88 |
| BrrLAC15.2 | 1683 | 63.3 | 9.03 | −0.157 | 34.88 |
| BrrLAC16 | 1698 | 62.5 | 9.14 | −0.052 | 35.26 |
| BrrLAC17.1 | 1722 | 63.7 | 9.28 | −0.168 | 33.69 |
| BrrLAC17.2 | 1719 | 63.5 | 9.27 | −0.147 | 34.10 |
| BrrLAC17.3 | 1722 | 63.7 | 9.33 | −0.120 | 33.72 |
| BrrLAC22 | 1683 | 61.8 | 8.47 | −0.081 | 27.47 |
| BraLAC1 | 1740 | 65.2 | 9.09 | −0.254 | 31.61 |
| BraLAC2 | 1737 | 64.2 | 9.47 | −0.091 | 35.57 |
| BraLAC3.1 | 1713 | 64.0 | 9.46 | −0.277 | 41.48 |
| BraLAC3.2 | 1713 | 64.1 | 9.48 | −0.265 | 40.58 |
| BraLAC4.1 | 1686 | 61.8 | 9.37 | −0.081 | 33.10 |
| BraLAC4.2 | 1680 | 61.8 | 9.41 | −0.061 | 31.49 |
| BraLAC5.1 | 1710 | 63.1 | 8.69 | −0.143 | 36.85 |
| BraLAC5.2 | 1725 | 63.8 | 8.99 | −0.158 | 35.27 |
| BraLAC6 | 1710 | 63.7 | 7.96 | −0.178 | 34.69 |
| BraLAC7 | 1707 | 62.7 | 8.94 | 0.015 | 37.38 |
| BraLAC8.1 | 1761 | 65.2 | 7.30 | −0.042 | 36.55 |
| BraLAC8.2 | 1776 | 66.0 | 7.30 | −0.049 | 35.31 |
| BraLAC9 | 1623 | 60.1 | 5.96 | −0.114 | 43.10 |
| BraLAC10 | 1680 | 61.2 | 9.49 | −0.034 | 33.40 |
| BraLAC11.1 | 1683 | 62.0 | 8.63 | −0.069 | 33.92 |
| BraLAC11.2 | 1686 | 62.6 | 8.91 | −0.114 | 35.96 |
| BraLAC12.1 | 1698 | 62.7 | 9.52 | −0.165 | 28.94 |
| BraLAC12.2 | 1698 | 62.5 | 9.10 | −0.134 | 30.60 |
| BraLAC13 | 1701 | 62.9 | 6.52 | −0.149 | 42.97 |
| BraLAC14 | 1743 | 65.8 | 9.73 | −0.301 | 27.60 |
| BraLAC15.1 | 1680 | 63.5 | 9.15 | −0.208 | 35.94 |
| BraLAC15.2 | 1683 | 63.2 | 9.03 | −0.159 | 34.55 |
| BraLAC16 | 1698 | 62.5 | 9.15 | −0.052 | 35.39 |
| BraLAC17.1 | 1722 | 63.7 | 9.18 | −0.150 | 34.17 |
| BraLAC17.2 | 1719 | 63.4 | 9.28 | −0.119 | 35.45 |
| BraLAC17.3 | 1722 | 63.6 | 9.33 | −0.119 | 34.14 |
| BraLAC22 | 1683 | 61.8 | 8.47 | −0.079 | 26.04 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, J.; Liu, Y.; Yang, S.; Yang, Y.; Wang, C. Genome-Wide Characterization of Laccase Gene Family from Turnip and Chinese Cabbage and the Role in Xylem Lignification in Hypocotyls. Horticulturae 2022, 8, 522. https://doi.org/10.3390/horticulturae8060522
Wen J, Liu Y, Yang S, Yang Y, Wang C. Genome-Wide Characterization of Laccase Gene Family from Turnip and Chinese Cabbage and the Role in Xylem Lignification in Hypocotyls. Horticulturae. 2022; 8(6):522. https://doi.org/10.3390/horticulturae8060522
Chicago/Turabian StyleWen, Jing, Yuanyuan Liu, Shuda Yang, Yongping Yang, and Chuntao Wang. 2022. "Genome-Wide Characterization of Laccase Gene Family from Turnip and Chinese Cabbage and the Role in Xylem Lignification in Hypocotyls" Horticulturae 8, no. 6: 522. https://doi.org/10.3390/horticulturae8060522
APA StyleWen, J., Liu, Y., Yang, S., Yang, Y., & Wang, C. (2022). Genome-Wide Characterization of Laccase Gene Family from Turnip and Chinese Cabbage and the Role in Xylem Lignification in Hypocotyls. Horticulturae, 8(6), 522. https://doi.org/10.3390/horticulturae8060522