
Combining Cultural Tactics and Insecticides for the Management of the Sweetpotato Whitefly, Bemisia tabaci MEAM1, and Viruses in Yellow Squash

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Evaluation of Pre-Plant Seedling Protection Tactics (Greenhouse)
2.2. Evaluation of Post-plant Protection Tactics (Field)
2.2.1. General
2.2.2. Evaluation of Mulch Types
2.2.3. Evaluation of Row Covers and Insecticides
2.3. Data Analysis
3. Results
3.1. Evaluation of Pre-Plant Seedling Protection Tactics (Greenhouse)
3.2. Evaluation of Post-Plant Protection Tactics (Field)
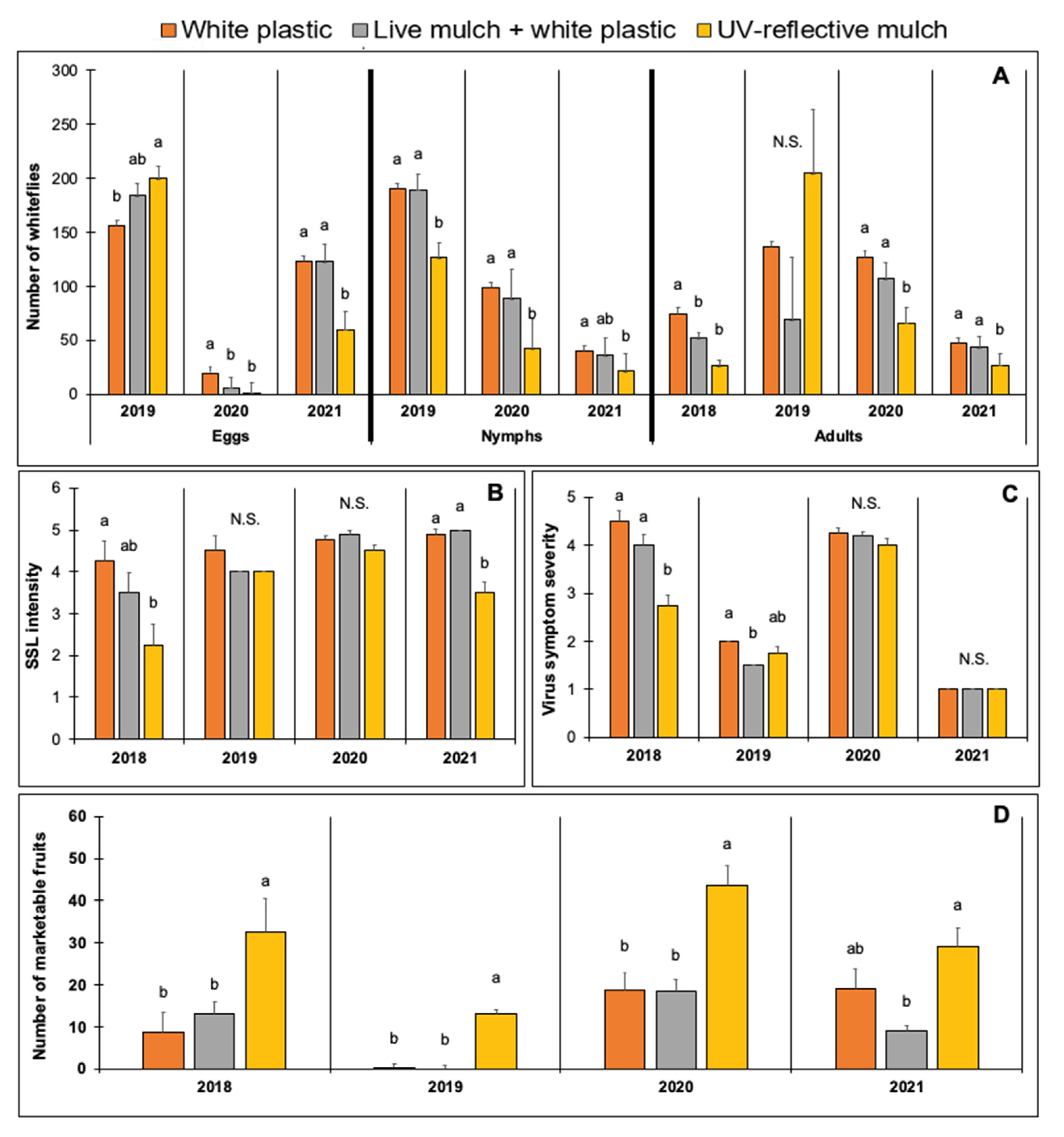
3.2.1. Evaluation of Mulch Types
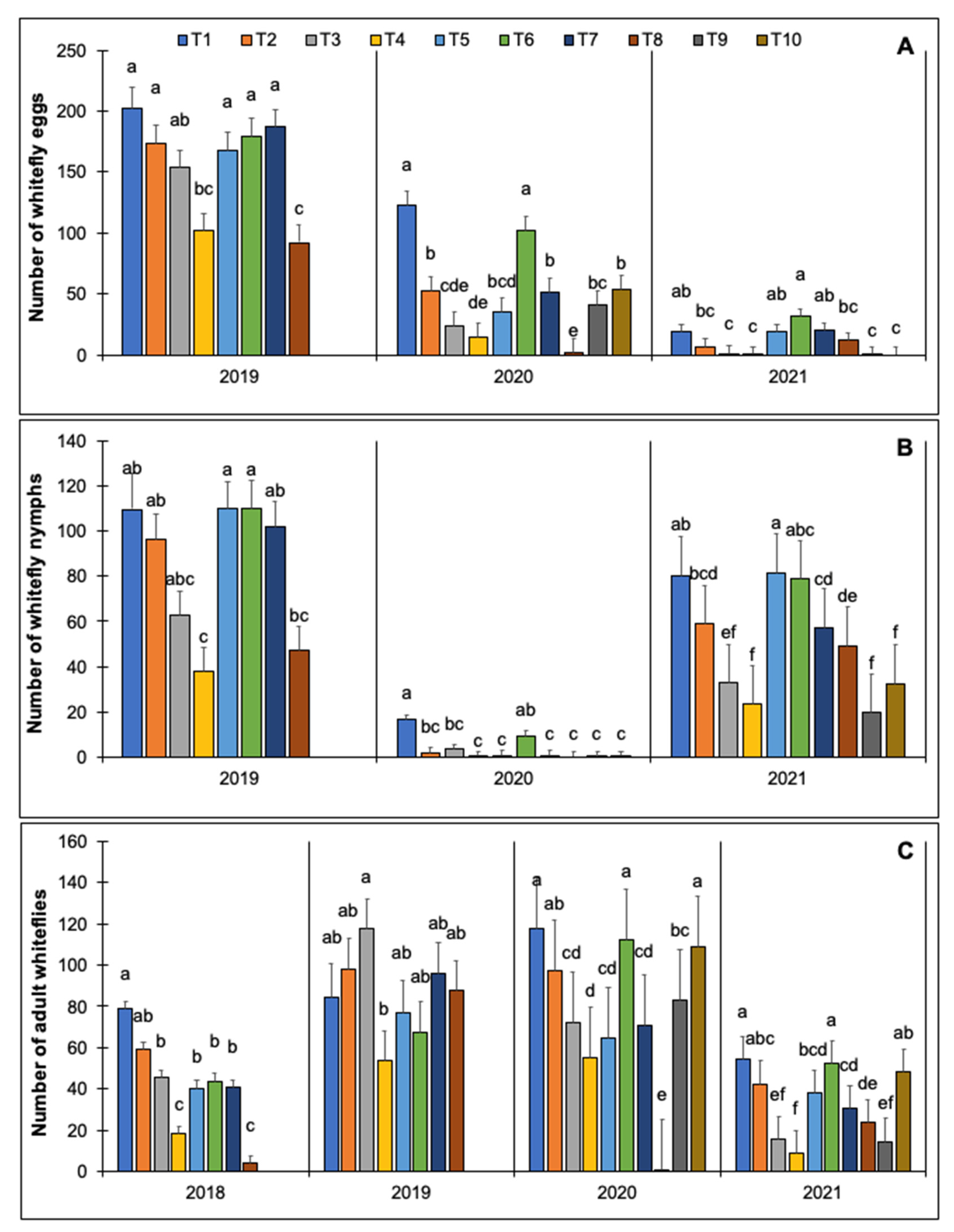
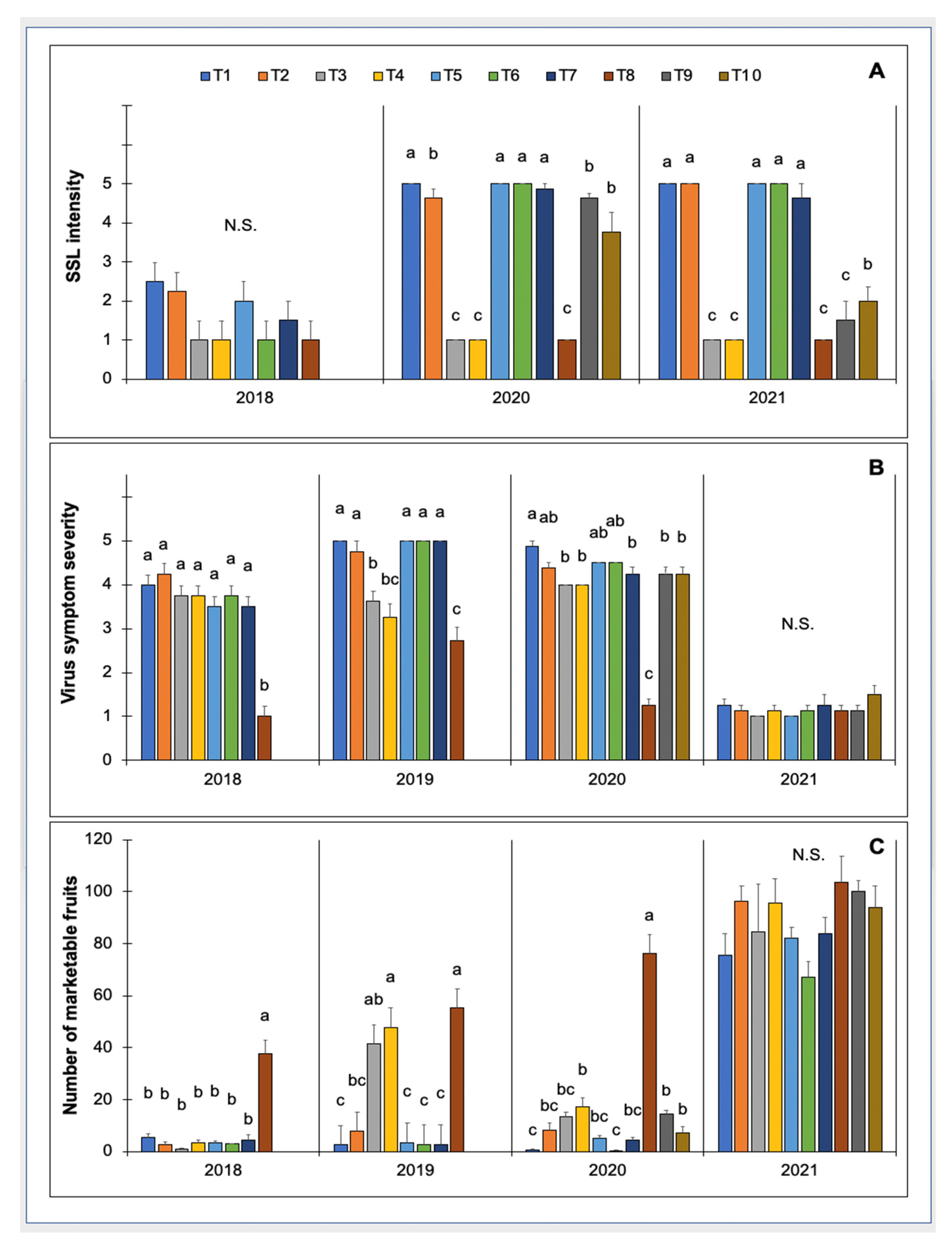
3.2.2. Evaluation of Row Covers and Insecticides
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stubbs, K. Georgia Farm Gate Value Report 2019; University of Georgia Center for Agribusiness and Economic Development: Athens, GA, USA, 2020; pp. 1–170. [Google Scholar]
- Srinivasan, R.; Riley, D.; Diffie, S.; Sparks, A.; Adkins, S. Whitefly population dynamics and evaluation of whitefly-transmitted tomato yellow leaf curl virus (TYLCV)-resistant tomato genotypes as whitefly and TYLCV reservoirs. J. Econ. Entomol. 2012, 105, 1447–1456. [Google Scholar] [CrossRef] [PubMed]
- Coolong, T. Yellow squash and zucchini cultivar evaluation in Georgia. Horttechnology 2017, 27, 296–302. [Google Scholar] [CrossRef]
- Kavalappara, S.R.; Milner, H.; Konakalla, N.C.; Morgan, K.; Sparks, A.N.; McGregor, C.; Culbreath, A.K.; Wintermantel, W.M.; Bag, S. High throughput sequencing-aided survey reveals widespread mixed infections of whitefly-transmitted viruses in cucurbits in Georgia, USA. Viruses 2021, 13, 988. [Google Scholar] [CrossRef]
- Yokomi, R.K.; Hoelmer, K.A.; Osborne, L.S. Relationships between the sweetpotato whitefly and the squash silverleaf disorder. Phytopathology 1990, 80, 895–900. [Google Scholar] [CrossRef]
- Schuster, D.J.; Kring, J.B.; Price, J.F. Association of the sweetpotato whitefly with a silverleaf disorder of squash. HortScience 1991, 26, 155–156. [Google Scholar] [CrossRef]
- McAuslane, H.J.; Chen, J.; Bruce Carle, R.; Schmalstig, J. Influence of Bemisia argentifolii (Homoptera: Aleyrodidae) infestation and squash silverleaf disorder on zucchini seedling growth. J. Econ. Entomol. 2004, 97, 1096–1105. [Google Scholar] [CrossRef]
- Liburd, O.E.; Nyoike, T.W.; Razze, J.M. Biology and management of whiteflies in sustainable field production of cucurbits. EDIS 2008, 2008, 1–3. [Google Scholar] [CrossRef]
- Little, E.L. 2017 Georgia Plant Disease Loss Estimates; University of Georgia: Athens, GA, USA, 2019; pp. 1–21. [Google Scholar]
- Little, E.L. 2018 Georgia Plant Disease Loss Estimates; University of Georgia: Athens, GA, USA, 2020; pp. 1–20. [Google Scholar]
- Little, E.L. 2019 Georgia Plant Disease Loss Estimates; University of Georgia: Athens, GA, USA, 2021; pp. 1–29. [Google Scholar]
- Brown, J.K.; Idris, A.M.; Olsen, M.W.; Miller, M.E.; Isakeit, T.; Anciso, J. Cucurbit leaf curl virus, a new whitefly transmitted geminivirus in Arizona, Texas, and Mexico. Mexico. Plant Dis. 2000, 84, 809. [Google Scholar] [CrossRef]
- Guzman, P.; Sudarshana, M.R.; Seo, Y.-S.; Rojas, M.R.; Natwick, E.; Turini, T.; Mayberry, K.; Gilbertson, R.L. A new bipartite geminivirus (Begomovirus) causing leaf curl and crumpling in cucurbits in the Imperial Valley of California. Plant Dis. 2000, 84, 488. [Google Scholar] [CrossRef]
- Akad, F.; Webb, S.; Nyoike, T.W.; Liburd, O.E.; Turechek, W.; Adkins, S.; Polston, J.E. Detection of Cucurbit leaf crumple virus in Florida cucurbits. Plant Dis. 2008, 92, 648. [Google Scholar] [CrossRef]
- Larsen, R.; Kmiecik, K. First report of cucurbit leaf crumple virus in snap bean in Georgia. In Proceedings of the 2010 American Phytopathological Society Annual Meeting, Charlotte, NC, USA, 7–11 August 2010. [Google Scholar]
- Kuo, Y.-W.; Rojas, M.R.; Gilbertson, R.L.; Wintermantel, W.M. First report of cucurbit yellow stunting disorder virus in California and Arizona, in association with cucurbit leaf crumple virus and squash leaf curl virus. Plant Dis. 2007, 91, 330. [Google Scholar] [CrossRef] [PubMed]
- Gadhave, K.R.; Dutta, B.; Coolong, T.; Sparks, A.N.; Adkins, S.; Srinivasan, R. First report of cucurbit yellow stunting disorder virus in cucurbits in Georgia, United States. Plant Health Prog. 2018, 19, 9–10. [Google Scholar] [CrossRef]
- Fiallo-Olivé, E.; Lett, J.M.; Martin, D.P.; Roumagnac, P.; Varsani, A.; Zerbini, F.M.; Navas-Castillo, J. ICTV virus taxonomy profile: Geminiviridae 2021. J. Gen. Virol. 2021, 102, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, M.; Bar-Joseph, M.; Candresse, T.; Maree, H.J.; Martelli, G.P.; Melzer, M.J.; Menzel, W.; Minafra, A.; Sabanadzovic, S. ICTV virus taxonomy profile: Closteroviridae. J. Gen. Virol. 2020, 101, 364–365. [Google Scholar] [CrossRef]
- Rosen, R.; Kanakala, S.; Kliot, A.; Pakkianathan, B.C.; Farich, B.A.; Santana-Magal, N.; Elimelech, M.; Kontsedalov, S.; Lebedev, G.; Cilia, M.; et al. Persistent, circulative transmission of begomoviruses by whitefly vectors. Curr. Opin. Virol. 2015, 15, 1–8. [Google Scholar] [CrossRef]
- Ghanim, M.; Morin, S.; Czosnek, H. Rate of Tomato yellow leaf curl virus translocation in the circulative transmission pathway of its vector, the whitefly Bemisia tabaci. Phytopathology 2001, 91, 188–196. [Google Scholar] [CrossRef]
- Cicero, J.M.; Brown, J.K. Anatomy of accessory salivary glands of the whitefly Bemisia tabaci (Hemiptera: Aleyrodidae) and correlations to begomovirus transmission. Ann. Entomol. Soc. Am. 2011, 104, 280–286. [Google Scholar] [CrossRef]
- Czosnek, H.; Hariton-Shalev, A.; Sobol, I.; Gorovits, R.; Ghanim, M. The incredible journey of Begomoviruses in their whitefly vector. Viruses 2017, 9, 273. [Google Scholar] [CrossRef]
- Gautam, S.; Gadhave, K.R.; Buck, J.W.; Dutta, B.; Coolong, T.; Adkins, S.; Srinivasan, R. Virus-virus interactions in a plant host and in a hemipteran vector: Implications for vector fitness and virus epidemics. Virus Res. 2020, 286, 198069. [Google Scholar] [CrossRef]
- Célix, A.; Lopez-Sesé, A.; Almarza, N.; Gomez-Guillamon, M.L.; Rodriguez-cerezo, E. Characterization of cucurbit yellow stunting disorder virus, a Bemisia tabaci-transmitted closterovirus. Phytopathology 1996, 86, 1370–1376. [Google Scholar]
- Wisler, G.C.; Duffus, J.E.; Liu, H.; Li, R.H. Ecology and epidemiology of whitefly-transmitted closteroviruses. Plant Dis. 1998, 82, 270–280. [Google Scholar] [CrossRef] [PubMed]
- Hogenhout, S.A.; Ammar, E.D.; Whitfield, A.E.; Redinbaugh, M.G. Insect vector interactions with persistently transmitted viruses. Annu. Rev. Phytopathol. 2008, 46, 327–359. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liang, X.; Wang, X.; Shi, Y.; Gu, Q.; Kuo, Y.W.; Falk, B.W.; Yan, F. Direct evidence for the semipersistent transmission of cucurbit chlorotic yellows virus by a whitefly vector. Sci. Rep. 2016, 6, 36604. [Google Scholar] [CrossRef] [PubMed]
- Schuster, D.J.; Stansly, P.A.; Polston, J.E. Expressions of plant damage of Bemisia. In Bemisia 1995: Taxonomy, Biology, Damage, Control and Management; Gerling, D., Mayer, R.T., Eds.; Intercept: Andover, GB, UK, 1996; pp. 153–165. [Google Scholar]
- Roy, A.; Acharyya, S.; Das, S.; Ghosh, R.; Paul, S.; Srivastava, R.K.; Ghosh, S.K. Distribution, epidemiology and molecular variability of the begomovirus complexes associated with yellow vein mosaic disease of mesta in India. Virus Res. 2009, 141, 237–246. [Google Scholar] [CrossRef]
- Levy, D.; Lapidot, M. Effect of plant age at inoculation on expression of genetic resistance to tomato yellow leaf curl virus. Arch. Virol. 2008, 153, 171–179. [Google Scholar] [CrossRef]
- Hooks, C.R.R.; Valenzuela, H.R.; Defrank, J. Incidence of pests and arthropod natural enemies in zucchini grown with living mulches. Agric. Ecosyst. Environ. 1998, 69, 217–231. [Google Scholar] [CrossRef]
- Webb, S.E.; Linda, S.B. Evaluation of spunbonded polyethylene row covers as a method of excluding insects and viruses affecting fall-grown squash in Florida. J. Econ. Entomol. 1992, 85, 2344–2352. [Google Scholar] [CrossRef]
- Candian, J.S.; Coolong, T.; Dutta, B.; Srinivasan, R.; Sparks, A.; Barman, A.; da Silva, A.L.B.R. Yellow squash and zucchini cultivar selection for resistance to cucurbit leaf crumple virus in the southeastern United States. Horttechnology 2021, 31, 504–513. [Google Scholar] [CrossRef]
- Luckew, A.; Meru, G.; Wang, Y.; Mwatuwa, R.; Kalischuk, M.; Luiz, A.; Ribeiro, B.; McGregor, C. Field evaluation of Cucurbita germplasm for resistance to whiteflies and whitefly-transmitted viruses. HortScience 2022, 57, 337–344. [Google Scholar] [CrossRef]
- Dempsey, M.; Riley, D.G.; Srinivasan, R. Insecticidal effects on the spatial progression of tomato yellow leaf curl virus and movement of its whitefly vector in tomato. J. Econ. Entomol. 2017, 110, 875–883. [Google Scholar] [CrossRef]
- Castle, S.; Palumbo, J.; Prabhaker, N. Newer insecticides for plant virus disease management. Virus Res. 2009, 141, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Riley, D.G.; Srinivasan, R. Integrated management of tomato yellow leaf curl virus and its whitefly vector in tomato. J. Econ. Entomol. 2019, 112, 1526–1540. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, N.E.; Kanan, H.O.; Sugimoto, Y.; Ma, Y.Q.; Inanaga, S. Effect of imidacloprid on incidence of tomato yellow leaf curl virus. Plant Dis. 2001, 85, 84–87. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Prabhaker, N.; Castle, S.; Perring, T.M. Baseline susceptibility of Bemisia tabaci B biotype (Hemiptera: Aleyrodidae) populations from California and Arizona to spirotetramat. J. Econ. Entomol. 2014, 107, 773–780. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.A.; Nagle, C.A.; MacVean, C.A.; McKenzie, C.L. Susceptibility of Bemisia tabaci MEAM1 (Hemiptera: Aleyrodidae) to imidacloprid, thiamethoxam, dinotefuran and flupyradifurone in South Florida. Insects 2016, 7, 57. [Google Scholar] [CrossRef]
- Horowitz, A.R.; Ghanim, M.; Roditakis, E.; Nauen, R.; Ishaaya, I. Insecticide resistance and its management in Bemisia tabaci species. J. Pest Sci. 2020, 93, 893–910. [Google Scholar] [CrossRef]
- Summers, C.G.; Stapleton, J.J. Use of UV reflective mulch to delay the colonization and reduce the severity of Bemisia argentifolii (Homoptera: Aleyrodidae) infestations in cucurbits. Crop Prot. 2002, 21, 921–928. [Google Scholar] [CrossRef]
- Polston, J.E.; Lapidot, M. Management of tomato yellow leaf curl virus: US and Israel perspectives. In Tomato Yellow Leaf Curl Virus Disease; Czosnek, H., Ed.; Springer: New York, NY, USA, 2007; pp. 251–262. [Google Scholar]
- Csizinszky, A.A.; Schuster, D.J.; Polston, J.E. Effect of ultraviolet-reflective mulches on tomato yields and on the silverleaf whitefly. HortScience 1999, 34, 911–914. [Google Scholar] [CrossRef]
- Lapidot, M.; Legg, J.P.; Wintermantel, W.M.; Polston, J.E. Management of whitefly-transmitted viruses in open-field production systems. In Advances in Virus Research; Loebenstein, G., Katis, N., Eds.; Academic Press: Burlington, NJ, USA, 2014; Volume 90, pp. 147–206. ISBN 9780128012468. [Google Scholar]
- Bugg, R.L.; Dutcher, J.D. Warm-season cover crops for pecan orchards: Horticultural and entomological implications. Biol. Agric. Hortic. 1989, 6, 123–148. [Google Scholar] [CrossRef]
- Frank, D.L.; Liburd, O.E. Effects of living and synthetic mulch on the population dynamics of whiteflies and aphids, their associated natural enemies, and insect-transmitted plant diseases in zucchini. Environ. Entomol. 2005, 34, 857–865. [Google Scholar] [CrossRef]
- Hilje, L.; Stansly, P.A. Living ground covers for management of Bemisia tabaci (Gennadius) (Homoptera: Aleyrodidae) and tomato yellow mottle virus (ToYMoV) in Costa Rica. Crop Prot. 2008, 27, 10–16. [Google Scholar] [CrossRef]
- Perring, T.M.; Royalty, R.N.; Farrar, C.A. Floating row covers for the exclusion of virus vectors and the effect on disease incidence and yield of cantaloupe. J. Econ. Entomol. 1989, 82, 1709–1715. [Google Scholar] [CrossRef]
- Costa, H.S.; Johnson, M.W.; Ullman, D.E. Row covers effect on sweetpotato whitefly (Homoptera: Aleyrodidae) densities, incidence of silverleaf, and crop yield in zucchini. J. Econ. Entomol. 1994, 87, 1616–1621. [Google Scholar] [CrossRef]
- Hilje, L.; Costa, H.S.; Stansly, P.A. Cultural practices for managing Bemisia tabaci and associated viral diseases. Crop Prot. 2001, 20, 801–812. [Google Scholar] [CrossRef]
- Roberts, P.; Stansly, P.; Adkins, S.; Kousik, C.S.; Bruton, B. Management of whitefly populations for the control of watermelon vine decline in Florida. Phytopathology 2007, 97, S182. [Google Scholar]
- The IRAC Mode of Action Classification Online. Available online: https://irac-online.org/modes-of-action/ (accessed on 2 February 2022).
- Navas-Castillo, J.; Fiallo-Olivé, E.; Sánchez-Campos, S. Emerging virus diseases transmitted by whiteflies. Ann. Rev. Phytopathol. 2011, 49, 219–248. [Google Scholar] [CrossRef]
- Kavalappara, S.R.; Milner, H.; Sparks, A.N.; McGregor, C.; Wintermantel, W.M.; Bag, S. First report of cucurbit chlorotic yellows virus in association with other whitefly-transmitted viruses in squash (Cucurbita pepo) in Georgia. Plant Dis. 2021, 105, 1862. [Google Scholar] [CrossRef]
- Gilbertson, R.L.; Rojas, M.; Natwick, E. Development of integrated pest management (IPM) strategies for whitefly (Bemisia tabaci)-transmissible geminiviruses. In The Whitefly, Bemisia Tabaci (Homoptera: Aleyrodidae) Interaction with Geminivirus-Infected Host Plants; Thompson, W.M.O., Ed.; Springer Science + Business Media: Berlin/Heidelberg, Germany, 2011; pp. 323–356. ISBN 9789400715240. [Google Scholar]
- Görlach, J.; Volrath, S.; Knauf-Beiter, G.; Hengy, G.; Beckhove, U.; Kogel, K.H.; Oostendorp, M.; Staub, T.; Ward, E.; Kessmann, H.; et al. Benzothiadiazole, a novel class of inducers of systemic acquired resistance, activates gene expression and disease resistance in wheat. Plant Cell 1996, 8, 629–643. [Google Scholar] [CrossRef]
- Csinos, A.S.; Pappu, H.R.; McPherson, R.M.; Stephenson, M.G. Management of tomato spotted wilt virus in flue-cured tobacco with acibenzolar-S-methyl and imidacloprid. Plant Dis. 2001, 85, 292–296. [Google Scholar] [CrossRef]
- Kenney, J.R.; Grandmont, M.-E.; Mauck, K.E. Priming melon defenses with acibenzolar-s-methyl attenuates infections by phylogenetically distinct infected hosts. Viruses 2020, 12, 257. [Google Scholar] [CrossRef]
- Nyoike, T.W.; Liburd, O.E.; Webb, S.E. Suppression of whiteflies, Bemisia tabaci (Hemiptera: Aleyrodidae) and incidence of cucurbit leaf crumple virus, a whitefly-transmitted virus of zucchini squash new to Florida, with mulches and imidacloprid. Fla. Entomol. 2008, 91, 460–465. [Google Scholar] [CrossRef]
- Nasruddin, A.; Agus, N.; Saubil, A.; Jumardi, J.; Rasyid, B.; Siriniang, A.; Nasruddin, A.D.; Firdaus, F.; Said, A.E. Effects of mulch type, plant cultivar, and insecticide use on sweet potato whitefly population in chili pepper. Scientifica 2020, 2020, 6428426 . [Google Scholar] [CrossRef] [PubMed]
- Csizinszky, A.A.; Schuster, D.J.; Kring, J.B. Evaluation of color mulches and oil sprays for yield and for the control of silverleaf whitefly, Bemisia argentifolii (Bellows and Perring) on tomatoes. Crop Prot. 1997, 16, 475–481. [Google Scholar] [CrossRef]
- Mahmoudpour, M.A.; Stapleton, J.J. Influence of sprayable mulch colour on yield of eggplant (Solanum melongena L. cv. Millionaire). Sci. Hortic. 1997, 70, 331–338. [Google Scholar] [CrossRef]
- Vos, J.G.M.; Sumarni, N. Integrated crop management of hot pepper (Capsicum spp.) under tropical lowland conditions: Effects of mulch on crop performance and production. J. Hortic. Sci. 1997, 72, 415–424. [Google Scholar] [CrossRef]
- Summers, C.G.; Stapleton, J.J.; Newton, A.S.; Duncan, R.A.; Hart, D. Comparison of sprayable and film mulches in delaying the onset of aphid-transmitted virus diseases in zucchini squash. Plant Dis. 1995, 79, 1126–1131. [Google Scholar] [CrossRef]
- Murphy, J.F.; Eubanks, M.D.; Masiri, J. Reflective plastic mulch but not a resistance-inducing treatment reduced watermelon mosaic virus incidence and yield losses in squash. Int. J. Veg. Sci. 2009, 15, 3–12. [Google Scholar] [CrossRef]
- Mansour, A.; Al-Musa, A. Tomato yellow leaf curl virus: Host range and virus-vector relationships. Plant Pathol. 1992, 41, 122–125. [Google Scholar] [CrossRef]
- Mehta, P.; Wyman, J.A.; Nakhla, M.K.; Maxwell, D.P. Transmission of tomato yellow leaf curl geminivirus by Bemisia tabaci (Homoptera: Aleyrodidae). J. Econ. Entomol. 1994, 87, 1291–1297. [Google Scholar] [CrossRef]
- Li, M.; Hu, J.; Xu, F.C.; Liu, S.S. Transmission of tomato yellow leaf curl virus by two invasive biotypes and a Chinese indigenous biotype of the whitefly Bemisia tabaci. Int. J. Pest Manag. 2010, 56, 275–280. [Google Scholar] [CrossRef]
- Bugg, R.L.; Phatak, S.C.; Dutcher, J.D. Insects associated with cool-season cover crops in southern Georgia: Implications for pest control in truck-farm and pecan agroecosystems. Biol. Agric. Hortic. 1990, 7, 17–45. [Google Scholar] [CrossRef]
- McNeill, C.A.; Liburd, O.E.; Chase, C.A. Effect of cover crops on aphids, whiteflies, and their associated natural enemies in organic squash. Sustain. Agric. 2012, 36, 382–403. [Google Scholar] [CrossRef]
- Razze, J.M.; Liburd, O.E.; McSorley, R. Preference of Bemisia tabaci biotype B on zucchini squash and buckwheat and the effect of Delphastus catalinae on whitefly populations. Pest Manag. Sci. 2016, 72, 1335–1339. [Google Scholar] [CrossRef]
- Hochmuth, G.J.; Hochmuth, R.C.; Kostewicz, S.; Stall, W. Row Covers for Commercial Vegetable Culture in Florida; University of Florida: Gainesville, FL, USA, 1987; pp. 1–11. [Google Scholar] [CrossRef]
- Poling, E.B.; Fuller, H.P.; Perry, K.B. Frost/freeze protection of strawberries grown on black plastic mulch. HortScience 1991, 26, 15–17. [Google Scholar] [CrossRef]
- Nyoike, T.W.; Liburd, O.E. Effect of living (buckwheat) and UV reflective mulches with and without imidacloprid on whiteflies, aphids and marketable yields of zucchini squash. Int. J. Pest Manag. 2010, 56, 31–39. [Google Scholar] [CrossRef]
- Adkins, S.; Webster, C.G.; Kousik, C.S.; Webb, S.E.; Roberts, P.D.; Stansly, P.A.; Turechek, W.W. Ecology and management of whitefly-transmitted viruses of vegetable crops in Florida. Virus Res. 2011, 159, 110–114. [Google Scholar] [CrossRef]
- Caballero, R.; Schuster, D.J.; Peres, N.A.; Mangandi, J.; Hasing, T.; Trexler, F.; Kalb, S.; Portillo, H.E.; Marçon, P.C.; Annan, I.B. Effectiveness of cyantraniliprole for managing Bemisia tabaci (Hemiptera: Aleyrodidae) and interfering with transmission of tomato yellow leaf curl virus on tomato. J. Econ. Entomol. 2015, 108, 894–903. [Google Scholar] [CrossRef]
- Kousik, C.S.; Adkins, S.T.; Turechek, W.; Roberts, P.D. Use of reflective plastic mulch and insecticide sprays to manage viral watermelon vine decline in Florida, 2007. Plant Dis. Manag. Rep. 2008, 2, V169. [Google Scholar]
- Kousik, C.S.; Adkins, S.; Webster, C.G.; Turechek, W.W.; Stansly, P.; Roberts, P.D. Influence of insecticides and reflective mulch on watermelon vine deqcline caused by squash vein yellowing virus (SqVYV). Plant Health Prog. 2015, 16, 43–49. [Google Scholar] [CrossRef]
- Castle, S.; Palumbo, J.; Merten, P.; Cowden, C.; Prabhaker, N. Effects of foliar and systemic insecticides on whitefly transmission and incidence of cucurbit yellow stunting disorder virus. Pest Manag. Sci. 2017, 73, 1462–1472. [Google Scholar] [CrossRef]
- Smith, H.A.; Nagle, C.A.; MacVean, C.M.; Vallad, G.E.; Van Santen, E.; Hutton, S.F. Comparing host plant resistance, repellent mulches, and at-plant insecticides for management of Bemisia tabaci MEAM1 (Hemiptera: Aleyrodidae) and tomato yellow leaf curl virus. J. Econ. Entomol. 2019, 112, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, A.L.; Kennedy, G.G. The effect of three rates of cyantraniliprole on the transmission of tomato spotted wilt virus by Frankliniella occidentalis and Frankliniella fusca (Thysanoptera: Thripidae) to Capsicum annuum. Crop Prot. 2011, 30, 512–515. [Google Scholar] [CrossRef]
- Jacobson, A.L.; Kennedy, G.G. Effect of cyantraniliprole on feeding behavior and virus transmission of Frankliniella fusca and Frankliniella occidentalis (Thysanoptera: Thripidae) on Capsicum annuum. Crop Prot. 2013, 54, 251–258. [Google Scholar] [CrossRef]
- Cameron, R.; Lang, E.B.; Alvarez, J.M. Use of honeydew production to determine reduction in feeding by Bemisia tabaci (Hemiptera: Aleyrodidae) adults when exposed to cyantraniliprole and imidacloprid treatments. J. Econ. Entomol. 2014, 107, 546–550. [Google Scholar] [CrossRef] [PubMed]
- Nauen, R.; Jeschke, P.; Velten, R.; Beck, M.E.; Ebbinghaus-Kintscher, U.; Thielert, W.; Wölfel, K.; Haas, M.; Kunz, K.; Raupach, G. Flupyradifurone: A brief profile of a new butenolide insecticide. Pest Manag. Sci. 2015, 71, 850–862. [Google Scholar] [CrossRef]
- Schuster, D.J.; Mann, R.S.; Toapanta, M.; Cordero, R.; Thompson, S.; Cyman, S.; Shurtleff, A.; Morris, R.F. Monitoring neonicotinoid resistance in biotype B of Bemisia tabaci in Florida. Pest Manag. Sci. 2010, 66, 186–195. [Google Scholar] [CrossRef]
- Caballero, R.; Cyman, S.; Schuster, D.J. Monitoring insecticide resistance in biotype b of Bemisia tabaci (Hemiptera: Aleyrodidae) in Florida. Fla. Entomol. 2013, 96, 1243–1256. [Google Scholar] [CrossRef]
- Webb, S.E.; Adkins, S.; Reitz, S.R. Semipersistent whitefly transmission of squash vein yellowing virus, causal agent of viral watermelon vine decline. Plant Dis. 2012, 96, 839–844. [Google Scholar] [CrossRef]
- Dutta, B.; Myers, B.; Coolong, T.; Srinivasan, R.; Sparks, A. Whitefly-transmitted plant viruses in South Georgia. Univ. Georg. Coop. Ext. Bull. 2018, 1507, 1–7. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trial | Active Ingredient | Trade Name | IRAC Sub-Group a | Year | Application Rate Per Hectare | |||
|---|---|---|---|---|---|---|---|---|
| 2018 | 2019 | 2020 | 2021 | |||||
| Greenhouse b | Cyantraniliprole | Verimark | 28 Diamide | ✓ | ✓ | 0.95 kg | ||
| Acibenzolar-S-methyl | Actigard | - | ✓ | ✓ | 17.51 g | |||
| Terpene constituents of Chenopodium ambrosioides near ambrosioides extract | Requiem Prime | UNE Botanical essence with unknown/uncertain MoA | ✓ | ✓ | 4.68 L | |||
| Field (Row cover/insecticide only) | Imidacloprid | Admire Pro | 4A Neonicotinoid | ✓ | ✓ | ✓ | ✓ | 0.73 L |
| Cyantraniliprole | Exirel | 28 Diamide | ✓ | ✓ | ✓ | ✓ | 1.50 L | |
| Flupyradifurone | Sivanto Prime | 4D Butenolide | ✓ | ✓ | ✓ | ✓ | 1.02 L | |
| Terpene constituents of Chenopodium ambrosioides near ambrosioides extract | Requiem Prime | UNE botanical essence with unknown/uncertain MoA | ✓ | ✓ | ✓ | ✓ | 7.01 L | |
| Chromobacterium subtsugae strain PRAA4–1 | Grandevo WDG | - | ✓ | ✓ | ✓ | ✓ | 3.36 kg | |
| Paraffinic oil | JMS Stylet-Oil | - | ✓ | ✓ | ✓ | ✓ | 14.03 L | |
| Afidopyropen | Sefina | 9D Pyropropene | ✓ | ✓ | 1.02 L | |||
| Spirotetramat + pyriproxyfen | Senstar | 23 Tetronic/tetramic acid derivative; 7C pyriproxyfen | ✓ | ✓ | 0.73 L | |||
| Treatments | df | 2018 | 2019 | ||
|---|---|---|---|---|---|
| F | p > F | F | p > F | ||
| Number of whitefly eggs | |||||
| Insect exclusion netting (IEN) | 1,72 | 57.57 | <0.0001 | 72.39 | <0.0001 |
| Insecticide/acibenzolar-S-methyl | 3,72 | 0.75 | 0.5260 | 0.42 | 0.7390 |
| IEN*Insecticide/acibenzolar-S-methyl | 3,72 | 0.81 | 0.4940 | 1.20 | 0.3140 |
| Number of whitefly nymphs | |||||
| IEN | 1,72 | 11.53 | 0.0011 | 14.43 | 0.0003 |
| Insecticide/acibenzolar-S-methyl | 3,72 | 0.36 | 0.7840 | 1.37 | 0.2577 |
| IEN*Insecticide/acibenzolar-S-methyl | 3,72 | 0.43 | 0.7317 | 1.30 | 0.2812 |
| Number of whitefly adults | |||||
| IEN | 1,72 | 79.93 | <0.0001 | 216.00 | <0.0001 |
| Insecticide/acibenzolar-S-methyl | 3,72 | 6.50 | 0.0006 | 22.90 | <0.0001 |
| IEN*Insecticide/acibenzolar-S-methyl | 3,72 | 6.50 | 0.0006 | 17.88 | <0.0001 |
| Treatments | Eggs a | Nymphs a | Adults a | |||
|---|---|---|---|---|---|---|
| 2018 | 2019 | 2018 | 2019 | 2018 | 2019 | |
| Main plot (IEN) | ||||||
| IEN | 0.25b | 1.80b | 0.18b | 0.28b | 0.00b | 0.13b |
| No IEN | 34.38a | 15.25a | 4.13a | 2.73a | 1.10a | 2.23a |
| Subplot (Insecticide/acibenzolar-S-methyl) | ||||||
| Non-treated control | 21.65a | 9.75a | 1.80a | 0.90a | 0.95a | 2.15a |
| Acibenzolar-S-methyl (17.51 g/ha) | 12.20a | 8.65a | 2.20a | 2.05a | 0.60ab | 0.70b |
| Terpene constituents of Chenopodium ambrosioides near ambrosioides extract (4.68 L/ha) | 17.35a | 8.45a | 1.50a | 2.25a | 0.20b | 0.70b |
| Cyantraniliprole (0.95 kg/ha) | 18.05a | 7.25a | 3.10a | 0.80a | 0.45b | 1.15b |
| IEN*Insecticide/acibenzolar-S-methyl | ||||||
| No IEN: Non-treated control | 43.10a | 17.90a | 3.60a | 1.50ab | 1.90a | 4.10a |
| No IEN: Acibenzolar-S-methyl | 23.90a | 12.90ab | 3.70a | 3.30ab | 1.20ab | 1.40b |
| No IEN: Terpene constituents of Chenopodium ambrosioides near ambrosioides extract | 34.40a | 16.10a | 3.00a | 4.50ab | 0.40cd | 1.40b |
| No IEN: Cyantraniliprole | 43.10a | 14.10ab | 6.20a | 1.60ab | 0.90bc | 2.00b |
| IEN: Non-treated control | 0.20b | 1.60c | 0.00a | 0.30b | 0.00d | 0.20c |
| IEN: Acibenzolar-S-methyl | 0.50b | 4.40bc | 0.70a | 0.80ab | 0.00d | 0.00c |
| IEN: Terpene constituents of Chenopodium ambrosioides near ambrosioides extract | 0.30b | 0.80c | 0.00a | 0.00b | 0.00d | 0.00c |
| IEN: Cyantraniliprole | 0.00b | 0.40c | 0.00a | 0.00b | 0.00d | 0.30c |
| Treatments by Level | Percent CuLCrV Incidence b |
|---|---|
| Main plot (IEN) | |
| IEN | 2.89b |
| No IEN | 5.39a |
| Subplot (Insecticide/acibenzolar-S-methyl) | |
| Non-treated control | 2.50b |
| Acibenzolar-S-methyl (17.51 g/ha) | 5.94a |
| Terpene constituents of Chenopodium ambrosioides near ambrosioides extract (4.68 L/ha) | 5.78a |
| Cyantraniliprole (0.95 kg/ha) | 2.34b |
| IEN *Insecticide/acibenzolar-S-methyl | |
| No IEN: Non-treated control | 3.13ab |
| No IEN: Acibenzolar-S-methyl | 7.81a |
| No IEN: Terpene constituents of Chenopodium ambrosioides near ambrosioides extract | 7.81a |
| No IEN: Cyantraniliprole | 2.81b |
| IEN: Non-treated control | 1.88b |
| IEN: Acibenzolar-S-methyl | 4.06ab |
| IEN: Terpene constituents of Chenopodium ambrosioides near ambrosioides extract | 3.75ab |
| IEN: Cyantraniliprole | 1.88b |
| Response Variable | 2018 | 2019 | 2020 | 2021 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| df | F | p > F | df | F | p > F | df | F | p > F | df | F | p > F | |
| Whitefly eggs a | - | - | - | 2,56 | 4.32 | 0.0179 | 2,235 | 5.38 | 0.0055 | 2,341 | 31.58 | <0.0001 |
| Whitefly nymphs a | - | - | - | 2,57 | 7.02 | 0.0019 | 2,235 | 14.48 | <0.0001 | 2,341 | 2.82 | 0.0118 |
| Whitefly adults | 2,357 | 26.38 | 0.0011 | 2,717 | 1.38 | 0.3208 | 2,286 | 18.61 | <0.0001 | 2,342 | 7.57 | 0.0006 |
| SSL intensity | 2,9 | 9.80 | 0.0129 | 2,9 | 2.33 | 0.1780 | 2,9 | 1.88 | 0.1680 | 2,9 | 22.23 | <0.0001 |
| Virus symptom severity | 2,9 | 16.71 | 0.0035 | 2,9 | 3.67 | 0.0911 | 2,9 | 2.37 | 0.1090 | 2,9 | 0.95 | 0.4010 |
| Yield | 2,9 | 5.08 | 0.0333 | 2,9 | 64.71 | <0.0001 | 2,33 | 13.06 | <0.0001 | 2,33 | 6.83 | 0.0052 |
| Response Variable | 2018 | 2019 | 2020 | 2021 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| df | F | p > F | df | F | p > F | df | F | p > F | df | F | p > F | |
| Whitefly eggs a | - | - | - | 7,755 | 10.82 | <0.0001 | 9,1160 | 46.45 | <0.0001 | 9,1154 | 11.71 | <0.0001 |
| Whitefly nymphs a | - | - | - | 7,755 | 7.40 | 0.0002 | 9,1160 | 8.75 | <0.0001 | 9,1189 | 16.22 | <0.0001 |
| Whitefly adults | 7,952 | 52.28 | <0.0001 | 7,1536 | 2.73 | 0.0349 | 9,1184 | 26.22 | <0.0001 | 9,1189 | 18.71 | <0.0001 |
| SSL intensity b | 7,24 | 1.69 | 0.1657 | - | - | - | 9,30 | 114.40 | <0.0001 | 9,30 | 69.34 | <0.0001 |
| Virus severity | 7,24 | 19.80 | <0.0001 | 7,24 | 22.41 | <0.0001 | 9,30 | 159.40 | <0.0001 | 9,30 | 0.92 | 0.519 |
| Yield | 7,24 | 33.88 | <0.0001 | 7,24 | 9.80 | <0.0001 | 9,30 | 56.81 | <0.0001 | 9,30 | 1.63 | 0.151 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
LaTora, A.G.; Codod, C.B.; Legarrea, S.; Dutta, B.; Kemerait, R.C., Jr.; Adkins, S.; Turechek, W.; Coolong, T.; da Silva, A.L.B.R.; Srinivasan, R. Combining Cultural Tactics and Insecticides for the Management of the Sweetpotato Whitefly, Bemisia tabaci MEAM1, and Viruses in Yellow Squash. Horticulturae 2022, 8, 341. https://doi.org/10.3390/horticulturae8040341
LaTora AG, Codod CB, Legarrea S, Dutta B, Kemerait RC Jr., Adkins S, Turechek W, Coolong T, da Silva ALBR, Srinivasan R. Combining Cultural Tactics and Insecticides for the Management of the Sweetpotato Whitefly, Bemisia tabaci MEAM1, and Viruses in Yellow Squash. Horticulturae. 2022; 8(4):341. https://doi.org/10.3390/horticulturae8040341
Chicago/Turabian StyleLaTora, Angela Gabrielle, Clarence Bagayao Codod, Saioa Legarrea, Bhabesh Dutta, Robert C. Kemerait, Jr., Scott Adkins, William Turechek, Timothy Coolong, Andre Luiz Biscaia Ribeiro da Silva, and Rajagopalbabu Srinivasan. 2022. "Combining Cultural Tactics and Insecticides for the Management of the Sweetpotato Whitefly, Bemisia tabaci MEAM1, and Viruses in Yellow Squash" Horticulturae 8, no. 4: 341. https://doi.org/10.3390/horticulturae8040341
APA StyleLaTora, A. G., Codod, C. B., Legarrea, S., Dutta, B., Kemerait, R. C., Jr., Adkins, S., Turechek, W., Coolong, T., da Silva, A. L. B. R., & Srinivasan, R. (2022). Combining Cultural Tactics and Insecticides for the Management of the Sweetpotato Whitefly, Bemisia tabaci MEAM1, and Viruses in Yellow Squash. Horticulturae, 8(4), 341. https://doi.org/10.3390/horticulturae8040341