Abstract
In this study, whether the addition of antifreeze protein (AFP) to a cryopreservative solution (plant vitrification solution 2 (PVS2)) is more effective in reducing freezing injuries in Hosta capitata than PVS2 alone at different cold exposure times (6, 24, and 48 h) is investigated. The upregulation of C-repeat binding factor 1 (CBF1) and dehydrin 1 (DHN1) in response to low temperature was observed in shoots. Shoots treated with distilled water (dH2O) strongly triggered gene expression 6 h after cold exposure, which was higher than those expressed in PVS2 and PVS2+AFP. However, 24 h after cold exposure, gene expressions detected in dH2O and PVS2 treatments were similar and higher than PVS2 + AFP. The expression was highest in PVS2+AFP when the exposure time was extended to 48 h. Similarly, nitric reductase activities 1 and 2 (Nia1 and Nia2) genes, which are responsible for nitric oxide production, were also upregulated in low-temperature-treated shoots, as observed for CBF1 and DHN1 expression patterns during cold exposure periods. Based on the gene expression patterns, shoots treated with PVS2+AFP were more likely to resist cold stress, which was also associated with the higher cryopreservation efficiency of PVS2+AFP compared to PVS2 alone. This finding suggests that the improvement of cryopreservation efficiency by AFP could be due to the transcriptional regulation of CBF1, DHN1, Nia1, and Nia2, which might reduce freezing injuries during cryopreservation. Thus, AFP could be potentially used as a cryoprotectant in the cryopreservation of rare and commercially important plant germplasm.
1. Introduction
Hosta species are herbaceous perennial plants. Their decorative foliage, diverse flower colors, attractive inflorescences, and ability to tolerate shade make them popular as groundcover ornamental plants in the landscape industry. Korea is one of the countries where some of these species, such as Hosta capitata, Hosta clausa, Hosta longipes, Hosta minor, and Hosta yingeri, are distributed [1]. However, the viral disease Hosta virus X has been found to infect the leaves of ornamental plants cultivated in Korea [2]. In addition, these species face a steady decline in their natural habitats due to overcollection and global climate change. This increases the risk of their extinction and potentially harms their use as groundcover plants in the landscape industry.
Cryopreservation has been widely used as a novel tool for both germplasm conservation and the production of virus-free plants [3,4,5]. However, severe intracellular ice formation has been the main challenge for the cryopreservation technique. Antifreeze protein (AFP) is added into the loading solution or plant vitrification solution 2 (PVS2) before dipping the biological materials into liquid nitrogen (LN) to increase the survival rate and plant regeneration. This intervention can inhibit the formation of ice crystals and protect cells from freezing injury [6]. Despite its application in the cryopreservation of animal cells [7,8,9], there have been only a few reports of AFP as a cryoprotectant in the cryopreservation of plant tissues and organs [10,11,12]. Jeon et al. [11] reported AFP’s inhibitory role in ice crystal formation in the cryopreservation of chrysanthemum shoot tips. Seo et al. [12] reported the transcriptional regulation of cold regulatory genes, such as C-repeat binding factor 1 (CBF1) and dehydrin 1 (DHN1), in cryopreserved potato shoots by AFP. In addition, AFP’s influence in the transcriptional regulation of CBF1 and DHN1 genes in H. capitata seedlings at a low temperature was also reported [13].
Previous reports have indicated the involvement of nitric oxide (NO) in plant acclimation and freezing tolerance, as NO acts as a signaling molecule in response to abiotic and biotic stresses [14,15]. At a low temperature (4 °C), NO production mainly depends on nitric reductase (NR) activity, which is controlled by Nia1 and Nia2 genes; gene knockout in plants reduced NO production and freezing tolerance [16,17]. NO synthesis in response to a low temperature has also been reported in Lotus japonicus and Pisum sativum [18,19]. However, the transcriptional regulation of Nia1 and Nia2 genes by AFP has not been investigated so far. Thus, it is also interesting to examine the genes involved in the cold tolerance mechanism in this study.
There have been no studies reporting the successful cryopreservation of Hosta species (H. capitata). Therefore, it is necessary to develop an optimal protocol for the successful cryopreservation of this species for germplasm conservation. It is of interest to determine whether AFP acts as a cryoprotectant in cryopreserved tissues. Hence, before cryopreservation, AFP’s role in the transcriptional regulation of genes involved in the cold tolerance mechanism is first tested. Its beneficial role in improving the cryopreservation efficiency of H. capitata is then investigated.
2. Materials and Methods
2.1. Plant Materials
In-vitro-grown H. capitata plants, obtained from a previous study, were kept in a hormone-free Murashige and Skoog (MS) medium [20] and subcultured in the same medium every 4 weeks. Only uniform and healthy plants were selected to obtain meristems (0.5 mm–1.0 cm) used as explants for this experiment.
2.2. AFP Treatment
To determine whether AFP can be used as a cryoprotectant in the cryopreservation process, before cryopreservation was started, its involvement in the low-temperature tolerance mechanism was initially accessed. Briefly, meristems were precultured for 3 h in a liquid MS medium (0.3 M sucrose) on a rotary shaker at 60 rpm. Meristems were dipped in a loading solution (LS; liquid MS medium, 0.4 M sucrose, 2.0 M glycerol (pH 5.8)) [21] for 30 min and then in plant vitrification solution 2 (PVS2; liquid MS medium, 3.28 M glycerol, 2.42 M ethylene glycol, 1.9 M dimethyl sulfoxide, 0.4 M sucrose (pH 5.8)) for 30 min at 25 °C [22] with or without AFP (100 or 0 ng/mL AFP III; A/F Protein, Inc., Waltham, MA, USA). Meristems were air-dried and cultured in a hormone-free MS medium and then kept at a low temperature (4 °C) for 6, 24, and 48 h. Meristems that were not dipped in any solution but kept at room temperature were used as controls. Those dipped in dH2O and placed at 4 °C were used as negative controls (Figure 1). A treatment contained 15 explants with three replications. After low-temperature treatment, meristems from each treatment were collected, placed in 1.5 mL Falcon tubes, immersed in liquid nitrogen (LN), and stored at −80 °C for RNA extraction and gene expression analysis.
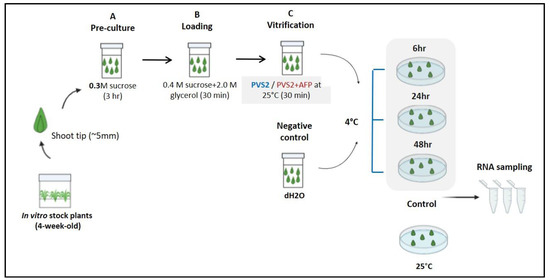
Figure 1.
Schematic diagram showing the application of antifreeze protein (AFP) as a cryoprotectant in the low-temperature tolerance mechanism.
2.3. RNA Extraction and Gene Expression Analysis
RNA was extracted from the meristems of the treatments mentioned above using the RNeasy Plant Mini Kit (Qiagen, Hilden, Germany), as described by Naing et al. [23]. cDNA was synthesized from 1 µg total RNA using an oligo(dT)20 primer. The transcript levels of genes (CBF1, DHN1, Nia1, and Nia2) involved in the low-temperature tolerance mechanism were detected using the Step One Plus Real-Time Polymerase Chain Reaction (PCR) System (Applied Biosystems, Foster City, CA, USA). The actin gene (ACT) was used as the reference gene. The primers and PCR conditions used for gene detection are listed in Table 1. Three independent biological samples per treatment were used for the analysis of each gene.

Table 1.
Primer sequences and PCR conditions used in this study for quantitative real-time PCR (qRT-PCR) analysis of Hosta capitata.
2.4. Cryopreservation
Cryopreservation of meristems was performed following Jeon et al.’s [11] protocol, with some modifications. Briefly, meristems (approximately 5 mm) were excised from 4-week-old in vitro H. capitata. The excised meristems were precultured in MS basal medium containing 0.3 M sucrose for 3 h and then loaded with LS for 30 min. The loaded meristems were treated with the PVS2 solution, with or without AFP (100 ng/mL), for 30 min at 0 °C. Then, meristems treated with the PVS2 solution and AFP were transferred to cryovials (Thermo Scientific, Waltham, MA, USA) containing fresh PVS2 and AFP; those treated with PVS2 solution only were also transferred to a cryovial containing PVS2 solution only. They were immersed in LN for 30 min. Meristems that were not treated with any loading solution but immersed in LN were used as controls. Meristems immersed in LN were rapidly warmed in a water bath at 38 °C for 3 min and then washed for 20 min at 25 °C with unloading solution (MS + 1.2 M sucrose) and washed thrice with dH2O. (Figure 2). There were 15 explants in a cryovial, with three replications.
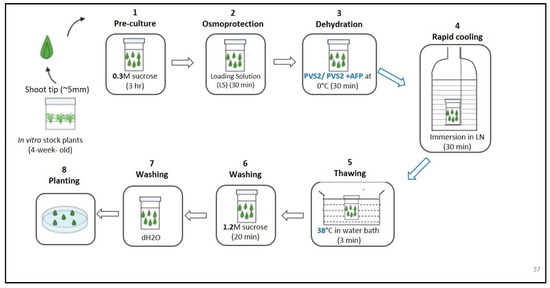
Figure 2.
Schematic diagram showing the steps involved in the cryopreservation of Hosta capitata. LN stands for liquid nitrogen.
2.5. Recovery of Cryopreserved Meristems
Cryopreserved meristems were then transferred to a postculture medium containing NH4+-free MS for 3 days in the dark at 25 ± 2 °C. The recovered shoots were then transferred to an MS basal medium containing α-naphthalene acetic acid and benzyladenine (KIN) for 4 weeks. The survival and shoot regeneration rates, with or without AFP, were compared to controls at 4 weeks postculture. The total number of explants per treatment was 45. Survival rate was defined as the percentage of meristems showing green tissues, and shoot regeneration was defined as the percentage of meristems inducing shoots.
2.6. Statistical Analysis
Data were statistically analyzed using analysis of variance (ANOVA) with SPSS version 11.09 (IBM Corporation, Armonk, NY, USA). Data are presented as means ± standard errors (SEs). Mean separations were carried out using the least significant difference test (LSDT), and significance was determined at the 5% level.
3. Results
3.1. Antifreeze Protein Regulates Genes Involved in the Low Temperature Tolerance Mechanism
The transcript levels of CBF1 and DHN1 genes expressed in shoot tips exposed to room temperature did not vary among treatment times (6, 24, and 48 h) and were significantly lower than those exposed to a low temperature (4 °C). Generally, the expression patterns of the detected genes were similar in trend for this investigation. At the low temperature, shoots treated with dH2O triggered CBF1 and DHN1 gene expression after 6 hr. Similarly, shoots treated with PVS2 also triggered gene expression, but their levels were significantly lower than those in dH2O. Gene induction was also observed in shoots treated with the AFP-containing PVS2 solution, which was even lower than PVS2 (Figure 3 and Figure 4).
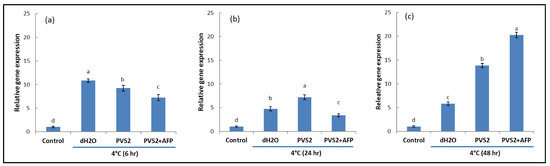
Figure 3.
Transcript levels of CBF1 in meristems of Hosta capitata treated with dH2O and plant vitrification solution 2 (PVS2), with or without AFP (AFP III), at 4 °C for different time intervals ((a): 6 h, (b): 24 h, and (c): 48 h). Plants at 25 °C were used as controls. The actin gene (ACT) was used as the reference gene. Data are presented as the means of three replicates. Error bars indicate standard errors of the means. Means with the same letter are not significantly different by the least significant difference test (LSDT, p < 0.05).
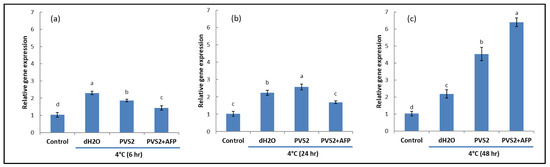
Figure 4.
Transcript levels of DHN1 in meristems of Hosta capitata treated with dH2O and plant vitrification solution 2 (PVS2), with or without AFP (AFP III), at 4 °C for different time intervals ((a): 6 h, (b): 24 h, and (c): 48 h). Plants at 25 °C were used as controls. The actin gene (ACT) was used as the reference gene. Data are presented as the means of three replicates. Error bars indicate standard errors of the means. Means with the same letter are not significantly different by the least significant difference test (LSDT, p < 0.05).
Higher expression was changed from dH2O to PVS2 when the treatment time was 24 h, whereas their expression in PVS2+AFP was still the lowest. When the treatment time was further extended to 48 h, their expression significantly decreased with the dH2O treatment, but not with PVS2, whereas CBF1 and DHN1 gene expression sharply increased with PVS2+AFP and were the highest.
To investigate NO and its involvement in cold acclimation, NR activities that produce NO were detected in shoots at low temperature (4 °C) and room temperature (25 °C) by analyzing the transcript levels of Nia1 and Nia2 genes. As expected, shoots at 4 °C triggered higher gene expression than shoots at 25 °C regardless of exposure times. However, shoots pretreated with PVS2 and PVS2+AFP induced lower expression levels than those treated with dH2O at 6 h after cold exposure, whereas those expressed in PVS2+AFP were the lowest. At 24 h after cold exposure, gene expression in dH2O and PVS2 was not significantly different but higher than PVS2+AFP. Interestingly, their expression in dH2O started to decline when the exposure time was increased to 48 h, but they were still moderately expressed in PVS2 and highly expressed in PVS2+AFP. However, the expression levels between the genes were slightly different for all treatments and exposure times (Figure 5 and Figure 6). These results suggest that low-temperature-induced upregulation of Nia1 and Nia2 results in NO production in plants at a low temperature; at 6 and 24 h exposure, the low-temperature-induced gene upregulation was lower in shoots pretreated with PVS2+AFP. Taken together, AFP is likely to be used for the reduction of cold stress in this species.
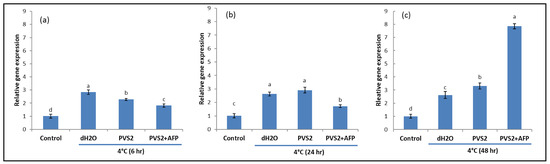
Figure 5.
Transcript levels of Nia1 in meristems of Hosta capitata treated with dH2O and plant vitrification solution 2 (PVS2), with or without AFP (AFP III), at 4 °C for different time intervals ((a): 6 h, (b): 24 h, and (c): 48 h). Plants at 25 °C were used as controls. The actin gene (ACT) was used as the reference gene. Data are presented as the means of three replicates. Error bars indicate standard errors of the means. Means with the same letter are not significantly different by the least significant difference test (LSDT, p < 0.05).
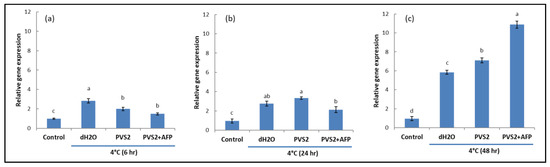
Figure 6.
Transcript levels of Nia2 in meristems of Hosta capitata treated with dH2O and plant vitrification solution (PVS2), with or without AFP (AFP III), at 4 °C for different time intervals ((a): 6 h, (b): 24 h, and (c): 48 h). Plants at 25 °C were used as controls. The actin gene (ACT) was used as the reference gene. Data are presented as the means of three replicates. Error bars indicate standard errors of the means. Means with the same letter are not significantly different by the least significant difference test (LSDT, p < 0.05).
3.2. Influence of Antifree Protein on Cryopreservation Efficiency
To further clarify whether the cryoprotective role of AFP improves the cryopreservation efficiency of this species, shoots were loaded with LS, cryoprotected with PVS2 (alone or in combination with AFP), and then immersed in LN. The addition of AFP to the PVS2 solution improved the survival rate (46.7%) and shoot regeneration (28.9%) compared to PVS2 alone (which showed a survival rate of 33.3% and shoot regeneration of 17.8%). However, the controls that were not treated with any cryoprotectant solution but immersed in LN did not survive at all (Table 2). This result suggests that the improvement of cryopreservation of this species by AFP could be through the transcriptional regulation of the genes mentioned above.
Table 2.
Effect of antifreeze protein III (AFP III) on the improvement of the cryopreservation efficiency of Hosta capitata.
4. Discussion
H. capitata, a popular groundcover plant in the landscape industry, has been facing a steady disappearance in natural habitats due to the overcollection of its germplasm and global climate change, which does not favor its natural growth in habitats. These factors increase the risk of extinction and potentially harm its use as a groundcover plant in the landscape industry. Unfortunately, to maintain its germplasm for long periods, an effective storage method for this species is not yet available. The cryopreservation technique has been widely used as a promising tool for the long-term preservation of genetic resources of various plants, including fruit trees, landscape ornamental plants, and cut flowers [11,24,25,26,27], due to reduced labor and space required for long-term storage. However, intracellular ice formation that occurs during cryopreservation remains a challenge. According to the results of preliminary works (unpublished data), cryopreservation of this species was unsuccessful, although several attempts have been made. Many studies have recently reported the cryoprotective role of AFP in cryopreservation and low-temperature treatment studies of horticultural plants [1,11,12,28]. Moreover, Cutler et al. [29] earlier reported that AFP’s vacuum infiltration into potato leaves also caused them to show increased cold tolerance compared to those infiltrated with water (control). Therefore, it was hypothesized that the cryopreservation of this species would be successful using AFP III in the cryopreservation process. Hence, it is of interest to apply AFP to the cryopreservation of H. capitata. The mechanism underlying AFP’s role as a cryoprotectant, by transcriptional analysis of genes involved in the cold tolerance mechanism, was investigated.
Before cryopreservation was started, to investigate whether the addition of AFP to the PVS2 solution would reduce freezing damage compared to PVS2 alone in the cryopreservation process, shoots were treated with PVS2 and PVS2+AFP, whereas shoots treated with dH2O were used as negative controls. Shoots were placed at 4 °C for 6, 24, and 48 h, respectively; untreated shoots placed at 25 °C were used as controls. Transcriptional analysis of CBF1, DHN1, Nia1, and Nia2 in shoots involved in the cold tolerance mechanism was then performed. Compared to 25 °C, upregulation of CBF1 and DHN1 at a low temperature was observed in this study, regardless of exposure times, as reported in previous studies [12,13,30,31,32,33,34,35,36,37]. Thus, it is suggested that CBF1 and DHN1 are low-temperature-inducible genes involved in the cold acclimation process of this Hosta species. Despite exposure to a low temperature for 6 h, higher gene expression was observed in dH2O compared to PVS2 and PVS2+AFP. It could be that shoots treated with dH2O were more responsive to cold than those treated with PVS2 and PVS2+AFP, which have cryoprotective activities. Therefore, the former triggered higher gene expression levels to adapt to the cold condition. Transcriptional control of genes by AFP in potato and the same species at a low temperature was recently reported [12,13]. When the exposure time was increased to 24 h, gene expression in dH2O and PVS2 was similar and higher than PVS2+AFP. It seemed that the presence of AFP in PVS2 is not required to trigger the genes as much as dH2O and PVS2 to adapt to the cold. However, dH2O treatment could not highly trigger the genes at 48 h after cold treatment, whereas PVS2 still triggered higher expression. This can be explained by the fact that the dH2O treatment was no longer able to resist the cold with increasing exposure times. However, the presence of AFP in the PVS2 solution was more likely to tolerate the cold than PVS2 alone because PVS2+AFP started to trigger the genes at 48 h, with higher expression compared to PVS2. Taken together, shoots treated with the AFP-containing PVS2 solution could reduce freezing injuries by enhancing the genes’ transcript levels.
Previous studies have indicated increased NO production in plants in response to a low temperature [14,15]. In this study, the upregulation of Nia1 and Nia2 genes that indirectly favor NO production in shoots at a low temperature was noticed. Shoots treated with dH2O triggered higher gene expression than those treated with PVS2 and PVS2+AFP at 6 h after cold exposure. It was likely that the former was more sensitive to the cold than the latter, thus immediately producing a higher amount of NO via gene upregulation to be used as signaling molecules in the early cold acclimation process. The latter’s lower sensitivity to cold stress could be due to the cryoprotective effect of PVS2 and AFP. Thus, they did not immediately need to trigger a higher gene expression level for NO production. The authors of [17] also observed NO production in Arabidopsis within 1 to 4 h after cold exposure, whereas NO production was positively associated with expression levels of Nia1 and Nia2 genes, and plants with a knockout of these genes (nia1/nia2 mutants) could not sufficiently produce NO at a low temperature. Increased NO production was likely to continue in shoots treated with dH2O until 24 h after cold exposure because gene expression levels did not significantly decline compared to 6 h. In contrast to 6 h, PVS2 treatment triggered higher expression at 24 h, suggesting a need for high content NO to adapt to cold stress when the exposure time was extended. At 48 h after cold exposure, a higher decline of the expression levels in dH2O than in PVS2 treatments could be due to the no-longer-high productivity of NO by shoots treated with dH2O at long exposure, compared to PVS2, which has cryoprotective activity. The presence of AFP in PVS2 was likely to have more resistance to cold stress than PVS2 alone, as the expression levels induced by AFP were more significant than those induced by PVS2 alone despite long exposure to cold. Despite NO production by cold exposure for 24 or 48 h [16,19], this is the first report of AFP controlling NO production at a low temperature. In this study, the expression patterns of CBF1, DHN1, Nia1, and Nia2 in response to a low temperature were similar. Cantrel et al. [17] also observed that CBF1 expression was positively associated with those of Nia1/Nia2, whereas CBF1 was greatly reduced in nia1/nia2 mutant plants at a low temperature compared to wild-type plants. These results further confirm the evidence for AFP’s potential role in reducing chilling injuries in the cold acclimation process. When AFP was used as a cryoprotectant in the cryopreservation of this species, the presence of AFP in PVS2 gave higher cryopreservation efficiency results than PVS2 alone, suggesting its beneficial role as a cryoprotectant in the cryopreservation of H. capitata.
5. Conclusions
The upregulation of CBF1, DHNI, Nia1, and Nia2 genes in H. capitata shoots in response to a low temperature was observed, demonstrating an important role in the cold stress tolerance mechanism. The gene expression patterns in shoots varied depending on the type of treatments and cold exposure times. Shoots treated with PVS2+AFP were likely insensitive to cold stress because they did not strongly trigger the genes at 6 and 24 h after cold exposure, whereas those treated with dH2O and PVS2 did so. Therefore, based on the gene expression patterns, the addition of AFP to PVS2 could be more effective in cold protection than PVS2 alone, as also confirmed by the higher cryopreservation efficiency of PVS2+AFP compared to PVS2 alone. This study suggests that AFP can be a potent cryoprotectant in the cryopreservation of rare and commercially important plant germplasm.
Author Contributions
Conceptualization, A.H.N.; methodology, P.P.W.P.; data curation, P.P.W.P.; writing—original draft preparation, P.P.W.P.; writing—review and editing, A.H.N.; project administration, K.I.P. and C.K.K. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a 2021 Yeungnam University Research Grant.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pe, P.P.W.; Kyu, S.W.; Naing, A.H.; Park, K.I.; Chung, M.Y.; Kim, C.K. Antifreeze proteins promote the germination of low temperature-treated petunia seeds via regulation of antioxidant- and proline-related genes. J. Plant Biotechnol. 2020, 47, 203–208. [Google Scholar] [CrossRef]
- Ryu, K.H.; Park, M.H.; Lee, J.S. Occurrence of mosaic disease of Hosta plants caused by Hosta virus X. Plant Pathol. 2002, 18, 313–316. [Google Scholar] [CrossRef]
- Helliot, B.; Panis, B.; Poumay, Y.; Swennen, R.; Lepoivre, P.; Frison, E. Cryopreservation for the elimination of cucumber mosaic and banana streak viruses from banana (Musa spp.). Plant Cell Rep. 2002, 20, 1117–1122. [Google Scholar] [CrossRef]
- Wang, Q.C.; Mawassi, M.; Li, P.; Gafny, R.; Sela, I.; Tanne, E. Elimination of grapevine virus A (GVA) by cryopreservation of in vitro-grown shoot tips of Vitis vinifera L. Plant Sci. 2003, 165, 321–327. [Google Scholar] [CrossRef]
- Wang, Q.C.; Valkonen, J.P.T. Elimination of two viruses which interact synergistically from sweetpotato by shoot tip culture and cryotherapy. J. Virol. Methods 2008, 154, 135–145. [Google Scholar] [CrossRef]
- Chapsky, L.; Rubinsky, B. Kinetics of antifreeze protein-induced ice growth inhibition. FEBS Lett. 1997, 412, 241–324. [Google Scholar] [CrossRef]
- Arav, A.; Shehu, D.; Mattioli, M. Osmotic and cytotoxic study of vitrification of immature bovine oocytes. J. Reprod Fertil. 1993, 99, 353–358. [Google Scholar] [CrossRef]
- Yeh, Y.; Feeney, R.E. Antifreeze proteins: Structures and mechanisms of function. Chem. Rev. 1996, 96, 601–618. [Google Scholar] [CrossRef]
- Naing, A.H.; Kim, C.K. A brief review of applications of antifreeze proteins in cryopreservation and metabolic genetic engineering. 3 Biotech 2019, 9, 329. [Google Scholar] [CrossRef]
- Wang, J.H.; Bian, H.W.; Zhang, Y.X.; Cheng, H.P. The dual effect of antifreeze protein on cryopreservation of rice (Oryza sativa l.) embryogenic suspension cells. Cryo Lett. 2001, 22, 175–182. [Google Scholar]
- Jeon, S.M.; Naing, A.H.; Park, K.I.; Kim, C.K. The effect of antifreeze protein on the cryopreservation of chrysanthemums. Plant Cell Tissue Organ Cult. 2015, 123, 665–671. [Google Scholar] [CrossRef]
- Seo, J.H.; Naing, A.H.; Jeon, S.M.; Kim, C.K. Anti-freezing-protein type III strongly infuences the expression of relevant genes in cryopreserved potato shoot tips. Plant Mol. Biol. 2018, 97, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Pe, P.P.W.; Naing, A.H.; Park, K.I.; Chung, M.Y.; Kim, C.K. The role of antifreeze proteins in the regulation of genes involved in the response of Hosta capitata to cold. 3 Biotech 2019, 9, 335. [Google Scholar] [CrossRef]
- Besson-Bard, A.; Pugin, A.; Wendehenne, D. New insights into nitric oxide signaling in plants. Annu. Rev. Plant Biol. 2008, 59, 21. [Google Scholar] [CrossRef]
- Neill, S.; Bright, J.; Desikan, R.; Hancock, J.; Harrison, J.; Wilson, I. Nitric oxide evolution and perception. J. Exp. Bot. 2008, 59, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.G.; Chen, L.; Zhang, L.L.; Zhang, W.H. Nitric reductase dependent nitric oxide production is involved in cold acclimation and freezing tolerance in Arabidopsis. Plant Physiol. 2009, 151, 755–767. [Google Scholar] [CrossRef] [PubMed]
- Cantrel, C.; Vazquez, T.; Puyaubert, J.; Rezé, N.; Lesch, M.; Kaiser, W.M.; Dutilleul, C.; Guillas, I.; Zachowski, A.; Baudouin, E. Nitric oxide participates in cold-responsive phosphosphingolipid formation and gene expression in Arabidopsis thaliana. New Phytol. 2011, 189, 415–427. [Google Scholar] [CrossRef]
- Shimoda, Y.; Nagata, M.; Suzuki, A.; Abe, M.; Sato, S.; Kato, T.; Tabata, S.; Higashi, S.; Uchiumi, T. Symbiotic rhizobium and nitric oxide induce gene expression of non-symbiotic hemoglobin in Lotus japonicas. Plant Cell Physiol. 2005, 46, 99–107. [Google Scholar] [CrossRef]
- Corpas, F.J.; Chaki, M.; Fernández-Ocan, A.; Valderrama, R.; Palma, J.M.; Carreras, A.; Begara-Morales, J.C.; Airaki, M.; A del Río, L.; Barroso, J.B. Metabolism of reactive nitrogen species in Pea plants under abiotic stress conditions. Plant Cell Physiol. 2008, 49, 1711–1722. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Hirai, D.; Sakai, A. Cryopreservation of in vitro-grown axillary shoot-tip meristems of mint (Mentha spicata L.) by encapsulation vitrification. Plant Cell Rep. 1999, 19, 15–155. [Google Scholar] [CrossRef] [PubMed]
- Sakai, A.; Kobayashi, S.; Oiyama, I. Cryopreservation of nucellar cells of navel orange (Citrus sinensis Osb. Var. brasiliensis Tanaka) by vitrification. Plant Cell Rep. 1990, 9, 30–33. [Google Scholar] [CrossRef]
- Naing, A.H.; Park, K.I.; Ai, T.N.; Chung, M.Y.; Han, J.S.; Kang, Y.W.; Kim, C.K. Overexpression of snapdragon Delila (Del) gene in tobacco enhances anthocyanin accumulation and abiotic stress tolerance. BMC Plant Biol. 2017, 17, 65. [Google Scholar] [CrossRef] [PubMed]
- Halmagyi, A.; Deliu, C. Cryopreservation of carnation (Dianthus caryophyllus L.) shoot tips by encapsulation-vitrification. Sci Hort. 2007, 113, 300–306. [Google Scholar] [CrossRef]
- Nikishina, T.V.; Popova, E.V.; Vakhrameeva, M.G.; Varlygina, T.I.; Kolomeitseva, G.L.; Burov, A.V.; Popovich, E.A.; Shirokov, A.I.; Shumilov, V.Y.; Popov, A.S. Cryopreservation of seeds and protocorms of rare temperate orchids. Russ. J. Plant Physiol. 2007, 54, 121–127. [Google Scholar] [CrossRef]
- Zhao, Y.; Wu, Y.; Chang, Y.; Reed, B.M. Cryopreservation of Fruit and Ornamental Trees. In Plant Cryopreservation: A Practical Guide; Reed, B.M., Ed.; Springer: New York, NY, USA, 2008. [Google Scholar] [CrossRef]
- Le Bras, C.; Le Besnerais, P.-H.; Hamama, L.; Grapin, A. Cryopreservation of ex-vitro-grown Rosa chinensis ‘Old Blush’ buds using droplet-vitrification and encapsulation-dehydration. Plant Cell Tiss Organ Cult. 2014, 116, 235–242. [Google Scholar] [CrossRef]
- Kyu, S.W.; Naing, A.H.; Pe, P.P.W.; Park, K.I.; Kim, C.K. Tomato seeds pretreated with Antifreeze protein type I (AFP I) promotes the germination under cold stress by regulating the genes involved in germination process. Plant Signal. Behav. 2019, 14, e1682796. [Google Scholar] [CrossRef]
- Cutler, A.J.; Saleem, M.; Kendall, E.; Gusta, L.V.; Georges, F.; Fletcher, G.L. Winter flounder antifreeze protein improves the cold hardiness of plant tissues. J. Plant Physiol. 1989, 135, 351–354. [Google Scholar] [CrossRef]
- Pennycooke, J.C.; Cheng, H.; Roberts, S.M.; Yang, Q.; Rhee, S.Y.; Stockinger, E.J. The low temperature-responsive, Solanum CBF1 genes maintain high identity in their upstream regions in a genomic environment undergoing gene duplications, deletions, and rearrangements. Plant Mol. Biol. 2008, 67, 483–497. [Google Scholar] [CrossRef] [PubMed]
- Benedict, C.; Skinner, J.S.; Meng, R.; Chang, Y.; Bhalerao, N.P.A.; Finn, C.E.; Chen, T.H.H.; Hurry, V. The CBF1-dependent low temperature signalling pathway, regulon and increase in freeze tolerance are conserved in Populus spp. Plant Cell Environ. 2006, 29, 1259–1272. [Google Scholar] [CrossRef]
- Choi, D.W.; Rodriguez, E.M.; Close, T.J. Barley CBF3 gene identification, expression pattern, and map location. Plant Physiol. 2002, 129, 1781–1787. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Fowler, S.G.; Cheng, H.; Lou, Y.; Rhee, S.Y.; Stockinger, E.J.; Thomashow, M.F. Freezing-sensitive tomato has a functional CBF cold response pathway, but a CBF regulon that differs from that of freezing-tolerant Arabidopsis. Plant J. 2004, 39, 905–919. [Google Scholar] [CrossRef]
- Bhattarai, T.; Fettig, S. Isolation and characterization of a dehydrin gene from Cicer pinnatifidum, a drought-resistant wild relative of chickpea. Physiol. Plant. 2005, 123, 452–458. [Google Scholar] [CrossRef]
- Welling, A.; Rinne, P.; Viherä-Aarnio, A.; Kontunen-Soppela, S.; Heino, P.; Palva, E.T. Photoperiod and temperature differentially regulate the expression of two dehydrin genes during overwintering of birch (Betula pubescens Ehrh.). J. Exp. Bot. 2004, 55, 507–516. [Google Scholar] [CrossRef]
- Baudo, M.M.; Meza-Zepeda, L.A.; Palva, E.T.; Heino, P. Induction of homologous low temperature and ABA-responsive genes in frost-resistant (Solanum commersonii) and frost-sensitive (Solanum tuberosum cv. Bintje) potato species. Plant Mol. Biol. 1996, 30, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Jaglo-Ottosen, K.R.; Gilmour, S.J.; Zarka, D.G.; Schabenberger, O.; Thomashow, M.F. Arabidopsis CBF1 overexpression induces COR genes and enhances freezing tolerance. Science 1998, 280, 104–106. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).