
Influence of Ripening on Polyphenolic Content, Degradative, and Browning Enzymes in Cantaloupe Varieties (C. Melo, L.)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Reagents and Standards
2.3. Physico-Chemical Properties
2.4. Bioactive Compounds and Antioxidant Activity
2.5. Enzymatic Activity
2.6. Statistical Analysis
3. Results and Discussion
3.1. Physico-Chemical Characteristics, Soluble Solids, pH, and Organic Acids
3.2. Bioactive Compounds and Antioxidant Activity
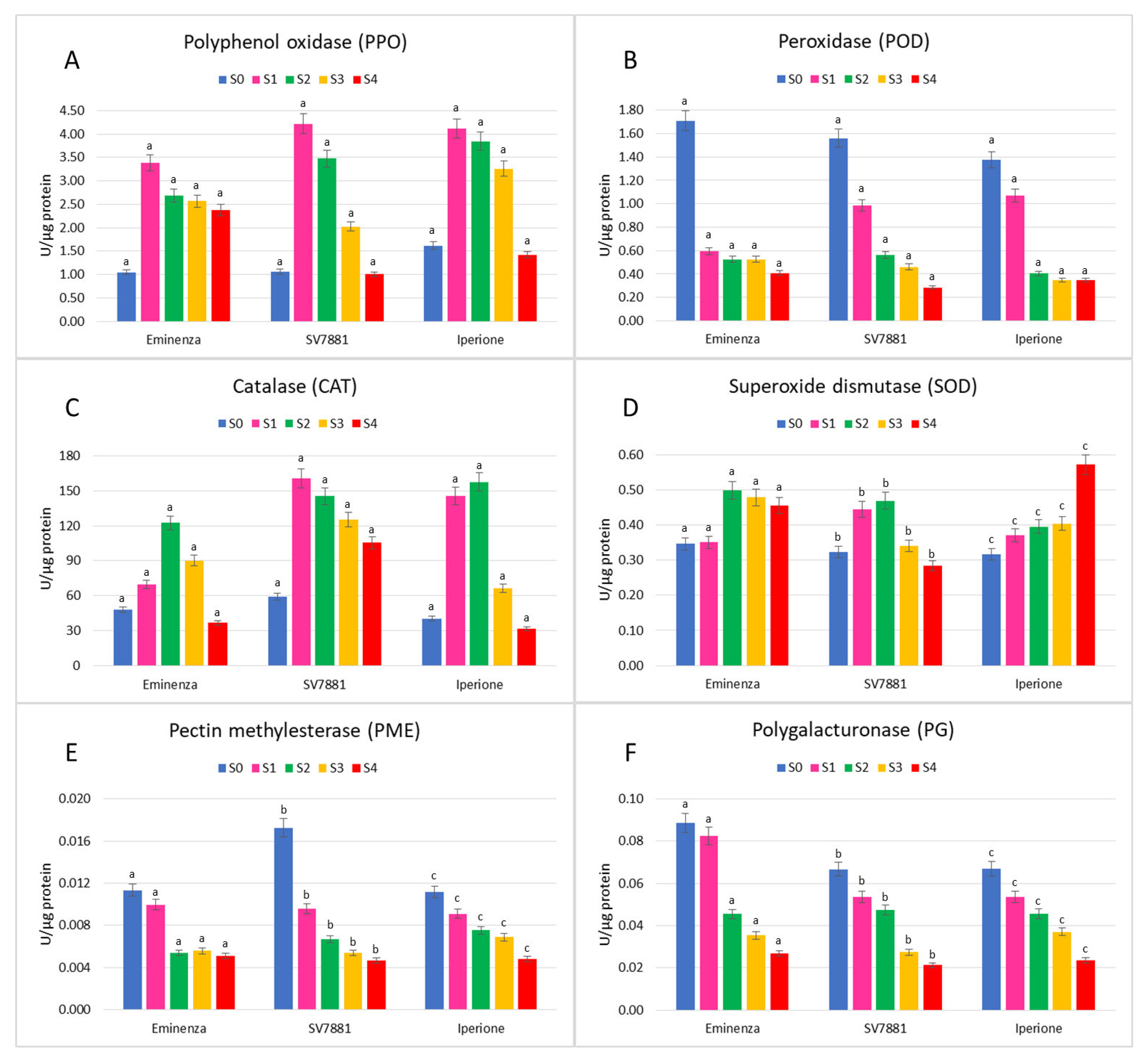
3.3. Enzymatic Activity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ismail, H.I.; Chan, K.W.; Mariod, A.A.; Ismail, M. Phenolic content and antioxidant activity of cantaloupe (Cucumis melo) methanolic extracts. Food Chem. 2010, 119, 643–647. [Google Scholar] [CrossRef]
- Maietti, A.; Tedeschi, P.; Stagno, C.; Bordiga, M.; Travaglia, F.; Locatelli, M.; Arlorio, M.; Brandolini, V. Analytical traceability of melon (Cucumis melo var. reticulatus): Proximate composition, bioactive compounds, and antioxidant capacity in relation to cultivar, plant physiology state, and seasonal variability. J. Food Sci. 2012, 77, C646–C652. [Google Scholar] [PubMed]
- Fundo, J.F.; Miller, F.A.; Garcia, E.; Santos, J.R.; Silva, C.L.; Brandão, T.R. Physicochemical characteristics, bioactive compounds and antioxidant activity in juice, pulp, peel and seeds of cantaloupe melon. J. Food Meas. Charact. 2018, 12, 292–300. [Google Scholar]
- Vouldoukis, I.; Lacan, D.; Kamate, C.; Coste, P.; Calenda, A.; Mazier, D.; Conti, M.; Dugas, B. Antioxidant and anti-inflammatory properties of a Cucumis melo LC. extract rich in superoxide dismutase activity. J. Ethnopharmacol. 2004, 94, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Faostat, Food and Agriculture Organization of the United States. Available online: http://www.fao.org/home/en/ (accessed on 30 April 2020).
- Lester, G.E. Antioxidant, sugar, mineral, and phytonutrient concentrations across edible fruit tissues of orange-fleshed honeydew melon (Cucumis melo L.). J. Agric. Food Chem. 2008, 56, 3694–3698. [Google Scholar] [CrossRef]
- Sroka, Z.; Cisowski, W. Hydrogen peroxide scavenging, antioxidant and anti-radical activity of some phenolic acids. Food Chem. Toxicol. 2003, 41, 753–758. [Google Scholar] [CrossRef]
- Shahidi, F.; Naczk, M. Phenolics in Food and Nutraceuticals; CRC Press: Boca Raton, FL, USA, 2004. [Google Scholar]
- Tachakittirungrod, S.; Okonogi, S.; Chowwanapoonpohn, S. Study on antioxidant activity of certain plants in Thailand: Mechanism of antioxidant action of guava leaf extract. Food Chem. 2007, 103, 381–388. [Google Scholar] [CrossRef]
- Schieber, A.; Stintzin, F.C.; Carle, R. By-products of plant food processing as a source of functional compounds—Recent developments. Trends Food Sci. Technol. 2001, 12, 401–413. [Google Scholar] [CrossRef]
- Galanakis, C.M. Recovery of high added-value components from food wastes: Conventional, emerging technologies and commercialized applications. Trends Food Sci. Technol. 2012, 26, 68–87. [Google Scholar] [CrossRef]
- Amaro, A.L.; Oliveira, A.; Almeida, D.P. Biologically active compounds in melon: Modulation by preharvest, post-harvest, and processing factors. In Processing and Impact on Active Components in Food; Academic Press: Cambridge, MA, USA; Elsevier: Amsterdam, The Netherlands, 2015; pp. 165–171. [Google Scholar]
- Masia, A. Superoxide dismutase and catalase activities in apple fruit during ripening and post-harvest and with special reference to ethylene. Physiol. Plant. 1998, 104, 668–672. [Google Scholar] [CrossRef]
- Jimenez, A.; Creissen, G.; Kular, B.; Firmin, J.; Robinson, S.; Verhoeyen, M.; Mullineaux, P. Changes in oxidative processes and components of the antioxidant system during tomato fruit ripening. Planta 2002, 214, 751–758. [Google Scholar] [CrossRef]
- Huang, R.; Xia, R.; Hu, L.; Lu, Y.; Wang, M. Antioxidant activity and oxygen-scavenging system in orange pulp during fruit ripening and maturation. Sci. Hortic. 2007, 113, 166–172. [Google Scholar] [CrossRef]
- Yemenicioglu, A.; Özkan, M.; Cemeroglu, B. Some characteristics of polyphenol oxidase and peroxidase from taro (Colocasia antiquorum). Turk. J. Agric. For. 1999, 23, 425–430. [Google Scholar]
- Ioannou, I. Prevention of enzymatic browning in fruit and vegetables. Eur. Sci. J. 2013, 9, 30. [Google Scholar]
- Zhang, X.; Shao, X. Characterisation of polyphenol oxidase and peroxidase and the role in browning of loquat fruit. Czech. J. Food Sci. 2015, 33, 109–117. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Chen, L.; Pang, L.; Chen, X.; Jia, X.; Li, X. Ultrasound treatment inhibits browning and improves antioxidant capacity of fresh-cut sweet potato during cold storage. RSC Adv. 2020, 10, 9193–9202. [Google Scholar] [CrossRef] [Green Version]
- Toivonen, P.M.; Brummell, D.A. Biochemical bases of appearance and texture changes in fresh-cut fruit and vegetables. Postharvest Biol. Tec. 2008, 48, 1–14. [Google Scholar]
- S2ingh, B.; Suri, K.; Shevkani, K.; Kaur, A.; Kaur, A.; Singh, N. Enzymatic browning of fruit and vegetables: A review. In Enzymes in Food Technology; Springer: Singapore, 2018; pp. 63–78. [Google Scholar]
- Chisari, M.; Barbagallo, R.N.; Spagna, G. Characterization and role of polyphenol oxidase and peroxidase in browning of fresh-cut melon. J. Agric. Food Chem. 2007, 56, 132–138. [Google Scholar] [CrossRef]
- Chisari, M.; Barbagallo, R.N.; Spagna, G. Characterization of polyphenol oxidase and peroxidase and influence on browning of cold stored strawberry fruit. J. Agric. Food Chem. 2007, 55, 3469–3476. [Google Scholar] [CrossRef]
- Huber, D.J. Polyuronide degradation and hemicellulose modifications in ripening tomato fruit. J. Am. Soc. Hortic. Sci. 1983, 108, 405–409. [Google Scholar]
- Chisari, M.; Silveira, A.C.; Barbagallo, R.N.; Spagna, G.; Artés, F. Ripening stage influenced the expression of polyphenol oxidase, peroxidase, pectin methylesterase and polygalacturonase in two melon cultivars. Int. J. Food Sci. Technol. 2009, 44, 940–946. [Google Scholar] [CrossRef]
- Menon, S.V.; Ramana Rao, T.V. Health-promoting components and related enzyme activities of muskmelon fruit during its development and ripening. J. Food Biochem. 2014, 38, 415–423. [Google Scholar] [CrossRef]
- Lamikanra, O. Enzymatic effects on flavor and texture of fresh-cut fruits and vegetables. In Fresh-Cut Fruits and Vegetables: Science, Technology and Market; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2002; pp. 125–186. [Google Scholar]
- Moon, K.M.; Kwon, E.B.; Lee, B.; Kim, C.Y. Recent trends in controlling the enzymatic browning of fruit and vegetable products. Molecules 2020, 25, 2754. [Google Scholar] [CrossRef]
- Prusky, D.; Lichter, A. Activation of quiescent infections by postharvest pathogens during transition from the biotrophic to the necrotrophic stage. FEMS Microbiol. Lett. 2007, 268, 1–8. [Google Scholar] [CrossRef]
- Alkan, N.; Fortes, A.M. Insights into molecular and metabolic events associated with fruit response to post-harvest fungal pathogens. Front. Plant. Sci. 2015, 6, 889. [Google Scholar] [CrossRef] [Green Version]
- Conesa, M.À.; Fullana-Pericàs, M.; Granell, A.; Galmés, J. Mediterranean long shelf-life landraces: An untapped genetic resource for tomato improvement. Front. Plant. Sci. 2020, 10, 1651. [Google Scholar] [CrossRef]
- Khabbazi, S.D.; Khabbazi, A.D.; Cevik, V.; Ergül, A. Genetic engineering of horticultural crops contributes to the improvement of crop nutritional quality and shelf life. In Transgenic Technology Based Value Addition in Plant Biotechnology; Elsevier: Amsterdam, The Netherlands, 2020; p. 247. [Google Scholar]
- Farcuh, M.; Copes, B.; Le-Navenec, G.; Marroquin, J.; Jaunet, T.; Chi-Ham, C.; Van Deynze, A. Texture diversity in melon (Cucumis melo L.): Sensory and physical assessments. Postharvest Biol. Technol. 2020, 159, 111024. [Google Scholar] [CrossRef]
- AOAC. Association of Official Analytical Chemist—Official Methods of Analysis, 14th ed.; Association of Official Analytical Chemists: Virginia, VA, USA, 1984; pp. 414–422. [Google Scholar]
- Vella, F.M.; Cautela, D.; Laratta, B. Characterization of polyphenolic compounds in cantaloupe melon by-products. Foods 2019, 8, 196. [Google Scholar] [CrossRef] [Green Version]
- Singleton, V.L.; Rossi, J.A., Jr. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Arnow, L.E. Colorimetric determination of the components of 3,4-dihydroxyphenylalaninetyrosine mixtures. J. Biol. Chem. 1937, 118, 531–537. [Google Scholar] [CrossRef]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoids contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Vella, F.M.; Laratta, B.; La Cara, F.; Morana, A. Recovery of bioactive molecules from chestnut (Castanea sativa Mill.) by-products through extraction by different solvents. Nat. Prod. Res. 2018, 32, 1022–1032. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Blois, M.S. Antioxidant determination by the use of a stable free radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Furumo, N.C.; Furutani, S. A simple method for assaying total protein, polyphenol oxidase and peroxidase activity from ‘Kaimana’ Litchi chinensis Sonn. J. Hawaii. Pac. Agric. 2008, 15, 1–7. [Google Scholar]
- Pinheiro, D.T.; Silva, A.L.D.; Silva, L.J.D.; Sekita, M.C.; Dias, D.C.F.D.S. Germination and antioxidant action in melon seeds exposed to salt stress. Pesqui. Agropecuária Trop. 2016, 46, 336–342. [Google Scholar] [CrossRef] [Green Version]
- Hagerman, A.E.; Austin, P.J. Continuous spectrophotometric assay for plant pectin methyl esterase. J. Agric. Food Chem. 1986, 34, 440–444. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Albuquerque, B.; Lidon, F.C.; Barreiro, M.G. A case study on the flavour properties of melon (Cucumis melo L.) cultivars. Fruits 2006, 61, 333–339. [Google Scholar] [CrossRef] [Green Version]
- Magwaza, L.S.; Opara, U.L. Analytical methods for determination of sugars and sweetness of horticultural products–A review. Sci. Hortic. 2015, 184, 179–192. [Google Scholar] [CrossRef]
- Tadmor, Y.; Burger, J.; Yaakov, I.; Feder, A.; Libhaber, S.E.; Portnoy, V.; Meir, A.; Tzuri, G.; Sa’ar, U.; Rogachev, I.; et al. Genetics of flavonoid, carotenoid, and chlorophyll pigments in melon fruit rinds. J. Agric. Food Chem. 2010, 58, 10722–10728. [Google Scholar] [CrossRef]
- Santos-Buelga, C.; Scalbert, A. Proanthocyanidins and tannin-like compounds-nature, occurrence, dietary intake, and effects on nutrition and health. J. Sci. Food Agric. 2000, 80, 1094–1117. [Google Scholar] [CrossRef]
- Buzzini, P.; Arapitsas, P.; Goretti, M.; Branda, E.; Turchetti, B.; Pinelli, P.; Ieri, F.; Romani, A. Antimicrobial and antiviral activity of hydrolysable tannins. Mini-Rev. Med. Chem. 2008, 8, 1179–1187. [Google Scholar] [CrossRef]
- Gülçin, I.; Huyut, Z.; Elmastaş, M.; Aboul-Enein, H.Y. Radical scavenging and antioxidant activity of tannic acid. Arab. J. Chem. 2010, 3, 43–53. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Koo, N.; Min, D.B. Reactive oxygen species, aging, and antioxidative nutraceuticals. Compr. Rev. Food Sci. Food Saf. 2004, 3, 21–33. [Google Scholar] [CrossRef]
- Holderbaum, D.F.; Kon, T.; Kudo, T.; Guerra, M.P. Enzymatic browning, polyphenol oxidase activity, and polyphenols in four apple cultivars: Dynamics during fruit development. HortScience 2010, 45, 1150–1154. [Google Scholar] [CrossRef] [Green Version]
- Murata, M.; Tsurutani, M.; Tomita, M.; Homma, S.; Kaneko, K. Relationship between apple ripening and browning: Changes in polyphenol content and polyphenol oxidase. J. Agric. Food Chem. 1995, 43, 1115–1121. [Google Scholar] [CrossRef]
- Brandelli, A.; Lopes, C.H. Polyphenoloxidase activity, browning potential and phenolic content of peaches during postharvest ripening. J. Food Biochem. 2005, 29, 624–637. [Google Scholar] [CrossRef]
- Chang, W.H.; Hwang, Y.J. Effect of ethylene treatment on the ripening, polyphenol oxidase activity and water-soluble tannin content of Taiwan northern banana at different maturity stages and the stability of banana polyphenol oxidase. Acta Hortic. 1990, 275, 603–610. [Google Scholar] [CrossRef]
- Weemaes, C.; Ludikhuyze, L.; Van den Broeck, I.; Hendrickx, M. Kinetic study of antibrowning agents and pressure inactivation of avocado polyphenoloxidase. J. Food Sci. 1999, 64, 823–827. [Google Scholar] [CrossRef]
- Cheema, S.; Sommerhalter, M. Characterization of polyphenol oxidase activity in Ataulfo mango. Food Chem. 2015, 171, 382–387. [Google Scholar] [CrossRef]
- Menon, S.V.; Ramana Rao, T.V. Nutritional quality of muskmelon fruit as revealed by its biochemical properties during different rates of ripening. Int. Food Res. J. 2012, 19, 1621–1628. [Google Scholar]
- Prasanna, V.; Prabha, T.N.; Tharanathan, R.N. Fruit ripening phenomena—An overview. Crit. Rev. Food Sci. Nutr. 2007, 47, 1–19. [Google Scholar] [CrossRef]
- Payasi, A.; Mishra, N.N.; Chaves, A.L.S.; Singh, R. Biochemistry of fruit softening: An overview. Physiol. Mol. Biol. Plants 2009, 15, 103–113. [Google Scholar] [CrossRef] [Green Version]
- Quiroga, M.; Guerrero, C.; Botella, M.A.; Barceló, A.; Amaya, I.; Medina, M.I.; Alonso, F.J.; de Forchetti, S.M.; Tigier, H.; Valpuesta, V. A tomato peroxidase involved in the synthesis of lignin and suberin. Plant. Physiol. 2000, 122, 1119–1128. [Google Scholar] [CrossRef] [Green Version]
- Bernards, M.A.; Summerhurst, D.K.; Razem, F.A. Oxidases, peroxidases and hydrogen peroxide: The suberin connection. Phytochem. Rev. 2004, 3, 113–126. [Google Scholar] [CrossRef]
- Zimmermann, P.; Heinlein, C.; Orendi, G.; Zentgraf, U. Senescence specific regulation of catalase in Arabidopsis thaliana (L.) heynh. Plant. Cell Environ. 2006, 29, 1049–1060. [Google Scholar]
- Ben-Amor, B.; Latché, A.; Bouzayen, M.; Pech, J.C.; Romojaro, F. Inhibition of ethylene biosynthesis by antisense ACC oxidase RNA prevents chilling injury in Charentais cantaloupe melons. Plant. Cell Environ. 1999, 22, 1579–1586. [Google Scholar] [CrossRef]
- Mondal, K.; Sharma, N.S.; Malhotra, S.P.; Dhawan, K.; Singh, R. Antioxidant systems in ripening tomato fruits. Biol. Plant. 2004, 48, 49–53. [Google Scholar] [CrossRef]
- Rogiers, S.Y.; Kumar, G.N.M.; Knowles, N.R. Maturation and ripening of fruit of Amelanchier alnifolia Nutt. are accompanied by increasing oxidative stress. Ann. Bot. 1998, 81, 203–211. [Google Scholar] [CrossRef] [Green Version]
- Baker, J.E. Superoxide dismutase in ripening fruits. Plant. Physiol. 1976, 58, 644–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selvaraj, Y.; Kumar, R. Studies on fruit softening enzymes and polyphenol oxidase activity in ripening mango (Mangifera indica L.) fruit. J. Food Sci. Technol. 1989, 26, 218–222. [Google Scholar]
- Prabha, T.N.; Yashoda, H.M.; Prasanna, V.; Jagadeesh, B.H.; Bimba Jain, M.V. Carbohydrate metabolism in relation to textural softening during fruit ripening. Trends Carbohyd. Chem. 2000, 6, 89–95. [Google Scholar]

{kind=link}
| Cultivar | Stage | Total Soluble Solids (°Brix) | pH | Acidity (g/100 mL of Acid Citric) |
|---|---|---|---|---|
| Eminenza | S0 | 7.75 ± 0.92 a | 5.70 ± 0.02 a | 0.134 ± 0.009 a |
| S1 | 11.00 ± 0.57 a | 6.23 ± 0.02 a | 0.108 ± 0.005 a | |
| S2 | 13.30 ± 0.85 a | 6.44 ± 0.02 a | 0.099 ± 0.003 a | |
| S3 | 16.50 ± 0.28 a | 6.55 ± 0.02 a | 0.073 ± 0.003 a | |
| S4 | 17.65 ± 0.35 a | 6.57 ± 0.02 a | 0.068 ± 0.004 a | |
| SV7881 | S0 | 6.60 ± 0.42 a | 5.85 ± 0.01 b | 0.106 ± 0.003 b |
| S1 | 10.10 ± 0.42 a | 6.64 ± 0.02 b | 0.082 ± 0.005 b | |
| S2 | 12.85 ± 0.78 a | 6.81 ± 0.02 b | 0.073 ± 0.003 b | |
| S3 | 13.70 ± 0.57 a | 6.86 ± 0.01 b | 0.066 ± 0.005 b | |
| S4 | 15.90 ± 0.14 a | 6.68 ± 0.02 b | 0.059 ± 0.002 b | |
| Iperione | S0 | 6.90 ± 0.42 a | 5.69 ± 0.01 a | 0.097 ± 0.004 c |
| S1 | 10.40 ± 0.37 a | 6.03 ± 0.03 a | 0.080 ± 0.003 c | |
| S2 | 13.10 ± 0.67 a | 6.59 ± 0.02 a | 0.056 ± 0.002 c | |
| S3 | 15.05 ± 0.21 a | 6.64 ± 0.02 a | 0.053 ± 0.005 c | |
| S4 | 16.35 ± 0.25 a | 7.01 ± 0.01 a | 0.040 ± 0.003 c |
| Cultivar | Stage | Total Polyphenols (µg GAE/mg) | Ortho-diphenols (µg CAE/mg) | Flavonoids (µg CE/mg) | Tannins (µg GAE/mg) | Antioxidant Power (µg AAE/mg) * | EC50 (mg/mL) ** |
|---|---|---|---|---|---|---|---|
| Eminenza | S0 | 2.22 ± 0.03 a | 0.36 ± 0.01 a | 0.24 ± 0.01 a | 0.32 ± 0.04 a | 0.45 ± 0.02 a | 38.85 ± 1.41 a |
| S1 | 2.46 ± 0.04 a | 0.58 ± 0.01 a | 0.27 ± 0.01 a | 0.49 ± 0.05 a | 0.73 ± 0.03 a | 37.41 ± 1.01 a | |
| S2 | 2.80 ± 0.07 a | 0.84 ± 0.01 a | 0.51 ± 0.01 a | 0.68 ± 0.08 a | 1.40 ± 0.04 a | 13.70 ± 0.94 a | |
| S3 | 2.62 ± 0.03 a | 0.61 ± 0.02 a | 0.44 ± 0.01 a | 0.39 ± 0.04 a | 0.93 ± 0.02 a | 15.44 ± 0.87 a | |
| S4 | 2.12 ± 0.08 a | 0.34 ± 0.01 a | 0.41 ± 0.01 a | 0.24 ± 0.08 a | 0.43 ± 0.01 a | 22.84 ± 0.88 a | |
| SV7881 | S0 | 2.02 ± 0.04 a | 0.36 ± 0.02 b | 0.29 ± 0.01 b | 0.25 ± 0.04 b | 0.63 ± 0.01 b | 40.75 ± 1.50 a |
| S1 | 2.15 ± 0.05 a | 0.42 ± 0.02 b | 0.34 ± 0.02 b | 0.27 ± 0.05 b | 0.73 ± 0.01 b | 37.19 ± 1.62 a | |
| S2 | 2.33 ± 0.04 a | 0.52 ± 0.01 b | 0.45 ± 0.01 b | 0.33 ± 0.04 b | 0.89 ± 0.02 b | 19.83 ± 0.84 a | |
| S3 | 3.68 ± 0.08 a | 0.73 ± 0.01 b | 0.59 ± 0.01 b | 0.54 ± 0.08 b | 1.48 ± 0.01 b | 17.94 ± 0.88 a | |
| S4 | 2.42 ± 0.01 a | 0.51 ± 0.01 b | 0.46 ± 0.01 b | 0.34 ± 0.04 b | 1.06 ± 0.04 b | 21.82 ± 0.74 a | |
| Iperione | S0 | 1.92 ± 0.06 b | 0.30 ± 0.01 a | 0.18 ± 0.01 c | 0.23 ± 0.06 c | 0.65 ± 0.02 b | 47.05 ± 0.99 a |
| S1 | 2.00 ± 0.01 b | 0.38 ± 0.01 a | 0.20 ± 0.01 c | 0.28 ± 0.03 c | 0.82 ± 0.01 b | 36.73 ± 1.22 a | |
| S2 | 2.40 ± 0.01 b | 0.54 ± 0.01 a | 0.34 ± 0.03 c | 0.53 ± 0.02 c | 1.00 ± 0.02 b | 21.05 ± 0.86 a | |
| S3 | 2.61 ± 0.02 b | 0.67 ± 0.01 a | 0.42 ± 0.01 c | 0.61 ± 0.03 c | 1.14 ± 0.01 b | 20.35 ± 0.41 a | |
| S4 | 2.63 ± 0.02 b | 0.81 ± 0.02 a | 0.48 ± 0.02 c | 0.87 ± 0.03 c | 1.50 ± 0.03 b | 13.40 ± 0.67 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vella, F.M.; Calandrelli, R.; Laratta, B. Influence of Ripening on Polyphenolic Content, Degradative, and Browning Enzymes in Cantaloupe Varieties (C. Melo, L.). Horticulturae 2021, 7, 421. https://doi.org/10.3390/horticulturae7110421
Vella FM, Calandrelli R, Laratta B. Influence of Ripening on Polyphenolic Content, Degradative, and Browning Enzymes in Cantaloupe Varieties (C. Melo, L.). Horticulturae. 2021; 7(11):421. https://doi.org/10.3390/horticulturae7110421
Chicago/Turabian StyleVella, Filomena Monica, Roberto Calandrelli, and Bruna Laratta. 2021. "Influence of Ripening on Polyphenolic Content, Degradative, and Browning Enzymes in Cantaloupe Varieties (C. Melo, L.)" Horticulturae 7, no. 11: 421. https://doi.org/10.3390/horticulturae7110421
APA StyleVella, F. M., Calandrelli, R., & Laratta, B. (2021). Influence of Ripening on Polyphenolic Content, Degradative, and Browning Enzymes in Cantaloupe Varieties (C. Melo, L.). Horticulturae, 7(11), 421. https://doi.org/10.3390/horticulturae7110421