

Comparative Analysis of In Vitro Fermentation Parameters in Total Mixed Rations of Dairy Cows with Varied Levels of Defatted Black Soldier Fly Larvae (Hermetia illucens) as a Substitute for Soybean Meal

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Rumen Fluid Collection and Donor Cows
2.2. Basal Diet and Black Soldier Fly Larvae
2.3. Determination of Chemical Compositions
2.4. In Vitro Experimental Design and Incubation Procedure
2.5. Total Gas Production and Fermentation Parameters
2.6. Determination of In Vitro Digestibility of Dry Matter and Neutral Detergent Fiber
2.7. Determination of Volatile Fatty Acids (VFAs) and Ammonia-Nitrogen (NH3-N)
2.8. Detemination of pH, Methane Emission, and Carbon Dioxide
2.9. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Makkar, H. Feed demand landscape and implications of food-not feed strategy for food security and climate change. Animal 2018, 12, 1744–1754. [Google Scholar] [CrossRef] [PubMed]
- Jayanegara, A.; Novandri, B.; Yantina, N.; Ridla, M. Use of black soldier fly larvae (Hermetia illucens) to substitute soybean meal in ruminant diet: An in vitro rumen fermentation study. Vet. World 2017, 10, 1439–1446. [Google Scholar] [CrossRef] [PubMed]
- Spiller, M.; Muys, M.; Papini, G.; Sakarika, M.; Buyle, M.; Vlaeminck, S.E. Environmental impact of microbial protein from potato wastewater as feed ingredient: Comparative consequential life cycle assessment of three production systems and soybean meal. Water Res. 2020, 171, 115406. [Google Scholar] [CrossRef] [PubMed]
- Hellstrand, S. Animal production in a sustainable agriculture. Environ. Dev. Sustain. 2013, 15, 999–1036. [Google Scholar] [CrossRef]
- Campos, A.; Pereira, O.; Ribeiro, K.; Santos, S.; Valadares Filho, S. Impact of replacing soybean meal in beef cattle diets with inactive dry yeast, a sugarcane by-product of ethanol distilleries and sugar mills. Anim. Feed Sci. Technol. 2014, 190, 38–46. [Google Scholar] [CrossRef]
- Yildiz, E.; Todorov, N. The comparison of the main protein sources for dairy cows: A review. Bulg. J. Agric. Sci. 2014, 20, 428–446. [Google Scholar]
- Hawkey, K.J.; Lopez-Viso, C.; Brameld, J.M.; Parr, T.; Salter, A.M. Insects: A Potential Source of Protein and Other Nutrients for Feed and Food. Annu. Rev. Anim. Biosci. 2021, 9, 333–354. [Google Scholar] [CrossRef]
- Castillo, C.; Hernández, J. Insects in ruminant nutrition as an urgent measure in the light of the scarcity of raw feedstock. Res. Vet. Sci. 2023, 155, 124–125. [Google Scholar] [CrossRef]
- Domingues, C.H.D.F.; Borges, J.A.R.; Ruviaro, C.F.; Gomes Freire Guidolin, D.; Rosa Mauad Carrijo, J. Understanding the factors influencing consumer willingness to accept the use of insects to feed poultry, cattle, pigs and fish in Brazil. PLoS ONE 2020, 15, e0224059. [Google Scholar] [CrossRef]
- Gasco, L.; Biancarosa, I.; Liland, N.S. From waste to feed: A review of recent knowledge on insects as producers of protein and fat for animal feeds. Curr. Opin. Green Sustain. Chem. 2020, 23, 67–79. [Google Scholar] [CrossRef]
- Poma, G.; Cuykx, M.; Amato, E.; Calaprice, C.; Focan, J.F.; Covacia, A. Evaluation of hazardous chemicals inedible insects and insect-based food intended for human consumption. Food Chem. Toxicol. 2017, 100, 70–79. [Google Scholar] [CrossRef] [PubMed]
- van Huis, A. Potential of insects as food and feed in assuring food security. Annu. Rev. Entomol. 2013, 58, 563–583. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.W.; Gulraize; Ali, U.; Ur Rehman, F.; Najeeb, H.; Sohail, M.; Irsa, B.; Muzaffar, Z.; Chaudhry, M.S. Evaluation of standard loose plastic packaging for the management of Rhyzopertha dominica (F.)(Coleoptera: Bostrichidae) and Tribolium castaneum (Herbst)(Coleoptera: Tenebriondiae). J. Insect Sci. 2016, 16, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Przemieniecki, S.W.; Kosewska, A.; Kosewska, O.; Purwin, C.; Lipiński, K.; Ciesielski, S. Polyethylene, polystyrene and lignocellulose wastes as mealworm (Tenebrio molitor L.) diets and their impact on the breeding condition, biometric parameters, metabolism, and digestive microbiome. Sci. Total Environ. 2022, 832, 154758. [Google Scholar] [CrossRef] [PubMed]
- Toral, P.G.; Hervás, G.; González-Rosales, M.G.; Mendoza, A.G.; Robles-Jiménez, L.E.; Frutos, P. Insects as alternative feed for ruminants: Comparison of protein evaluation methods. J. Anim. Sci. Biotechnol. 2022, 13, 21. [Google Scholar] [CrossRef] [PubMed]
- Makkar, H.P.; Tran, G.; Heuzé, V.; Ankers, P. State-of-the-art on use of insects as animal feed. Anim. Feed Sci. Technol. 2014, 197, 1–33. [Google Scholar] [CrossRef]
- Marono, S.; Piccolo, G.; Loponte, R.; Di Meo, C.; Attia, Y.A.; Nizza, A.; Bovera, F. In vitro crude protein digestibility of Tenebrio molitor and Hermetia illucens insect meals and its correlation with chemical composition traits. Ital. J. Anim. Sci. 2015, 14, 3889. [Google Scholar] [CrossRef]
- Ahmed, E.; Fukuma, N.; Hanada, M.; Nishida, T. Insects as Novel Ruminant Feed and a Potential Mitigation Strategy for Methane Emissions. Animals 2021, 11, 2648. [Google Scholar] [CrossRef]
- Yang, Z.; Liu, S.; Xie, T.; Wang, Q.; Wang, Z.; Yang, H.; Li, S.; Wang, W. Effect of Unsaturated Fatty Acid Ratio In Vitro on Rumen Fermentation, Methane Concentration, and Microbial Profile. Fermentation 2022, 8, 540. [Google Scholar] [CrossRef]
- Onetti, S.G.; Shaver, R.D.; McGuire, M.A.; Grummer, R.R. Effect of type and level of dietary fat on rumen fermentation and performance of dairy cows fed corn silage-based diets. J. Dairy Sci. 2001, 84, 2751–2759. [Google Scholar] [CrossRef]
- Association of offical Analytic chemists (AOAC). Official Method of Analytic, 17th ed.; AOAC: Arilington, VA, USA, 2002; Volume 1, pp. 120–155. [Google Scholar]
- Van Soest, P.v.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Krishnamoorthy, U.; Muscato, T.; Sniffen, C.; Van Soest, P. Nitrogen fractions in selected feedstuffs. J. Dairy Sci. 1982, 65, 217–225. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Dairy Cattle, 7th ed.; The National Academies Press: Washington, WA, USA, 2001. [Google Scholar]
- Ahmed, E.; Nishida, T. Optimal Inclusion Levels of Cricket and Silkworm as Alternative Ruminant Feed: A Study on Their Impacts on Rumen Fermentation and Gas Production. Sustainability 2023, 15, 1415. [Google Scholar] [CrossRef]
- Kara, K.; Ozkaya, S.; Guclu, B.K.; Aktug, E.; Demir, S.; Yılmaz, S.; Pirci, G.; Yılmaz, K.; Baytok, E. In vitro ruminal fermentation and nutrient compositions of potato starch by-products. J. Anim. Feed Sci. 2023, 32, 306–315. [Google Scholar] [CrossRef]
- Schofield, P.; Pitt, R.; Pell, A. Kinetics of fiber digestion from in vitro gas production. J. Anim. Sci. 1994, 72, 2980–2991. [Google Scholar] [CrossRef] [PubMed]
- Johnston, J.; Tricarico, J. Practical implications of fiber in dairy rations: Making use of forage fiber. In Proceedings of the 22nd Annual Southwest Nutrition & Management Conference, Tempe, AZ, USA, 22–23 February 2007; pp. 22–23. [Google Scholar]
- Tunkala, B.Z.; DiGiacomo, K.; Alvarez Hess, P.S.; Dunshea, F.R.; Leury, B.J. Impact of rumen fluid storage on in vitro feed fermentation characteristics. Fermentation 2023, 9, 392. [Google Scholar] [CrossRef]
- Menke, K.H.; Steingass, H. Estimation of the energetic feed value obtained from chemical analysis and in vitro gas production using rumen fluid. Anim. Res. Dev. 1988, 28, 7255. [Google Scholar]
- Blümmel, M.; Steingaβ, H.; Becker, K. The relationship between in vitro gas production, in vitro microbial biomass yield and 15N incorporation and its implications for the prediction of voluntary feed intake of roughages. Br. J. Nutr. 1997, 77, 911–921. [Google Scholar] [CrossRef]
- Cobellis, G.; Acuti, G.; Forte, C.; Menghini, L.; De Vincenzi, S.; Orrù, M.; Valiani, A.; Pacetti, D.; Trabalza-Marinucci, M. Use of Rosmarinus officinalis in sheep diet formulations: Effects on ruminal fermentation, microbial numbers and in situ degradability. Small Rumin. Res. 2015, 126, 10–18. [Google Scholar] [CrossRef]
- Weatherburn, M. Phenol-hypochlorite reaction for determination of ammonia. Anal. Chem. 1967, 39, 971–974. [Google Scholar] [CrossRef]
- Moss, A.R.; Jouany, J.-P.; Newbold, J. Methane production by ruminants: Its contribution to global warming. In Proceedings of the Annales De Zootechnie; Institut National de la Recherche Agronomique: Paris, France, 2000; pp. 231–253. [Google Scholar]
- Blümmel, M.; Aiple, K.P.; Steingaβ, H.; Becker, K. A note on the stoichiometrical relationship of short chain fatty acid production and gas formation in vitro in feedstuffs of widely differing quality. Anim. Physiol. Anim. Nutr. 1999, 81, 157–167. [Google Scholar] [CrossRef]
- Maxin, G.; Ouellet, D.; Lapierre, H. Ruminal degradability of dry matter, crude protein, and amino acids in soybean meal, canola meal, corn, and wheat dried distillers grains. J. Dairy Sci. 2013, 96, 5151–5160. [Google Scholar] [CrossRef] [PubMed]
- Jeong, C.-D.; Mamuad, L.L.; Kim, S.-H.; Choi, Y.J.; Soriano, A.P.; Cho, K.K.; Jeon, C.-O.; Lee, S.S.; Lee, S.-S. Effect of soybean meal and soluble starch on biogenic amine production and microbial diversity using in vitro rumen fermentation. Asian-Australas. J. Anim. Sci. 2015, 28, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Das, L.K.; Kundu, S.; Kumar, D.; Datt, C. Metabolizable protein systems in ruminant nutrition: A review. Vet. World. 2014, 7, 622–629. [Google Scholar] [CrossRef]
- Wei, Z.; Zhang, B.; Liu, J. Effects of the dietary nonfiber carbohydrate content on lactation performance, rumen fermentation, and nitrogen utilization in mid-lactation dairy cows receiving corn stover. J. Anim. Sci. Biotechnol. 2018, 9, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Renna, M.; Coppa, M.; Lussiana, C.; Le Morvan, A.; Gasco, L.; Maxin, G. Full-fat insect meals in ruminant nutrition: In vitro rumen fermentation characteristics and lipid biohydrogenation. J. Anim. Sci. Biotechnol. 2022, 13, 138. [Google Scholar] [CrossRef] [PubMed]
- Bach, A.; Calsamiglia, S.; Stern, M.D. Nitrogen metabolism in the rumen. J. Dairy Sci. 2005, 88 (Suppl. 1), E9–E21. [Google Scholar] [CrossRef] [PubMed]
- Astuti, D.; Anggraeny, A.; Khotijah, L.; Suharti, S.; Jayanegara, A. Performance, physiological status, and rumen fermentation profiles of pre-and post-weaning goat kids fed cricket meal as a protein source. Trop. J. Anim. Sci. 2019, 42, 145–151. [Google Scholar] [CrossRef]
- Chaudhari, S.S.; Arakane, Y.; Specht, C.A.; Moussian, B.; Boyle, D.L.; Park, Y.; Kramer, K.J.; Beeman, R.W.; Muthukrishnan, S. Knickkopf protein protects and organizes chitin in the newly synthesized insect exoskeleton. Proc. Natl. Acad. Sci. USA 2011, 108, 17028–17033. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Rodríguez, M.; Barroso, F.G.; Fabrikov, D.; Sánchez-Muros, M.J. In vitro crude protein digestibility of insects: A review. Insects 2022, 13, 682. [Google Scholar] [CrossRef]
- Phesatcha, B.; Phesatcha, K.; Viennaxay, B.; Matra, M.; Totakul, P.; Wanapat, M. Cricket Meal (Gryllus bimaculatus) as a Protein Supplement on In Vitro Fermentation Characteristics and Methane Mitigation. Insects 2022, 13, 129. [Google Scholar] [CrossRef]
- Pinho, R.; Santos, E.M.; de Oliveira, J.S.; de Carvalho, G.G.; Alves, J.P.; Macêdo, A.J.d.S.; Pereira, G.A.; Pereira, D.M.; Perazzo, A.F.; Zanine, A.d.M. Relationship between forage neutral detergent fiber and non-fibrous carbohydrates on ruminal fermentation products and neutral detergent fiber digestibility in goats. Rev. Colomb. Cienc. Pecu. 2019, 32, 126–138. [Google Scholar] [CrossRef]
- Homem Junior, A.C.; Ezequiel, J.M.B.; Perez, H.L.; Almeida, M.T.C.; Paschoaloto, J.R.; Carvalho, V.B.d.; Cremasco, L.F.; da Costa, M.B. In vitro fermentation of corn silage using rumen fluid buffered or not and different sample amounts. Cienc. Rural. 2015, 45, 2229–2232. [Google Scholar] [CrossRef]
- Sung, H.-G.; Kobayashi, Y.; Chang, J.; Ha, A.; Hwang, I.-H.; Ha, J. Low Ruminal pH Reduces Dietary Fiber Digestion via Reduced Microbial Attachment. Asian-Australas. J. Anim. Sci. 2007, 20, 200–207. [Google Scholar] [CrossRef]
- Buccioni, A.; Decandia, M.; Minieri, S.; Molle, G.; Cabiddu, A. Lipid metabolism in the rumen: New insights on lipolysis and biohydrogenation with an emphasis on the role of endogenous plant factors. Anim. Feed Sci. Technol. 2012, 174, 1–25. [Google Scholar] [CrossRef]
- Palmquist, D.L.; Jenkins, T.C. A 100-Year Review: Fat feeding of dairy cows. J. Dairy Sci. 2017, 100, 10061–10077. [Google Scholar] [CrossRef] [PubMed]
- Fischer, H.; Romano, N.; Sinha, A.K. Conversion of spent coffee and donuts by black soldier fly (Hermetia illucens) larvae into potential resources for animal and plant farming. Insects 2021, 12, 332. [Google Scholar] [CrossRef] [PubMed]
- De Boever, J.; Aerts, J.; Vanacker, J.; De Brabander, D. Evaluation of the nutritive value of maize silages using a gas production technique. Anim. Feed Sci. Technol. 2005, 123, 255–265. [Google Scholar] [CrossRef]
- Meneguz, M.; Schiavone, A.; Gai, F.; Dama, A.; Lussiana, C.; Renna, M.; Gasco, L. Effect of rearing substrate on growth performance, waste reduction efficiency and chemical composition of black soldier fly (Hermetia illucens) larvae. J. Sci. Food Agric. 2018, 98, 5776–5784. [Google Scholar] [CrossRef]
- Pasini, G.; Cullere, M.; Vegro, M.; Simonato, B.; Dalle Zotte, A. Potentiality of protein fractions from the house cricket (Acheta domesticus) and yellow mealworm (Tenebrio molitor) for pasta formulation. LWT-Food Sci. Technol. 2022, 164, 113638. [Google Scholar] [CrossRef]


{kind=link}
| Diets | |||
|---|---|---|---|
| Item | TMR0 | TMR20 | TMR40 |
| Ingredients (% DM) | |||
| Corn silage | 30 | 30 | 30 |
| Wheat straw | 15 | 15 | 15 |
| Alfalfa hay | 15 | 15 | 15 |
| Barley, crushed | 24 | 24 | 24 |
| Black soldier fly larvae meal | 0 | 3.2 | 6.4 |
| Soybean meal | 16 | 12.6 | 9.6 |
| Chemical composition (DM basis) | |||
| TDN, % * | 70.13 | 71.09 | 71.89 |
| NEL, Mcal/kg * | 1.71 | 1.8 | 1.87 |
| NEM, Mcal/kg * | 1.83 | 1.95 | 2.03 |
| NEG, Mcal/kg * | 1.19 | 1.29 | 1.36 |
| Dry matter, % | 50.28 | 53 | 52.68 |
| Crude protein, % | 16.32 | 17.2 | 16.76 |
| Ether extraction, % | 3.19 | 3.74 | 3.97 |
| NDF, % | 36.39 | 36.54 | 36.45 |
| ADF, % | 23.57 | 23.06 | 23.32 |
| NFC, % | 27.93 | 26.33 | 26.74 |
| Starch, % | 19.15 | 22.17 | 23.80 |
| Ash% | 6.76 | 6.67 | 6.01 |
| A fraction, % CP | 20.66 | 18.55 | 18.56 |
| Level of Black Soldier Fly Larvae | |||||
|---|---|---|---|---|---|
| Parameters | TMR0 | TMR20 | TMR40 | Linear | Quadratic |
| pH | 6.51 ± 0.02 | 6.56 ± 0.01 | 6.59 ± 0.02 | 0.356 | 0.256 |
| IVDMD,% | 48.85 ± 2.03 | 50.47 ± 0.55 | 48.07 ± 0.57 | 0.407 | 0.312 |
| IVNDFD, % | 33.47 ± 1.13 b | 38.63 ± 1.18 a | 34.62 ± 1.11 b | 0.012 | 0.024 |
| Acetate, mmol/L | 49.57 ± 7.37 | 45.46 ± 7.37 | 46.11 ± 4.47 | 0.653 | 0.221 |
| Propionate, mmol/L | 24.46 ± 2.73 | 21.92 ± 3.17 | 22.95 ± 2.28 | 0.454 | 0.224 |
| Butyrate, mmol/L | 0.44 ± 0.04 | 0.43 ± 0.01 | 0.46 ± 0.12 | 0.667 | 0.517 |
| Isobutyric, mmol/L | 8.96 ± 8.75 | 8.79 ± 0.66 | 9.13 ± 1.13 | 0.684 | 0.452 |
| Isovaleric, mmol/L | 0.69 ± 0.08 | 0.77 ± 0.11 | 0.78 ± 0.02 | 0.305 | 0.711 |
| Valeric, mmol/L | 1.39 ± 0.12 | 1.44 ± 0.13 | 1.50 ± 0.31 | 0.277 | 0.951 |
| Hexanoic, mmol/L | 0.27 ± 0.04 | 0.28 ± 0.02 | 0.28 ± 0.05 | 0.447 | 0.274 |
| Total VFA, mmol/L | 85.80 ± 3.38 a | 79.80 ± 3.85 b | 81.27 ± 2.40 b | 0.042 | 0.045 |
| CH4 mmol/L | 19.44 ± 2.84 | 17.94 ± 3.52 | 19.01 ± 2.67 | 0.720 | 0.222 |
| CO2 mmol/L | 44.10 ± 5.36 | 41.39 ± 5.9 | 43.52 ± 3.42 | 0.803 | 0.236 |
| Acet/Prop | 2.11 ± 0.44 | 2.06 ± 0.13 | 2.09 ± 0.04 | 0.893 | 0.362 |
| NH3-N, mg/dL | 3.04 ± 0.68 a | 2.20 ± 0.64 b | 2.94 ± 0.79 a | 0.017 | 0.019 |
| Level of Black Soldier Fly Larvae | |||||
|---|---|---|---|---|---|
| Parameters | TMR0 | TMR20 | TMR40 | Linear | Quadratic |
| pH | 6.45 ± 0.03 | 6.41 ± 0.02 | 6.42 ± 0.03 | 0.152 | 0.102 |
| IVDMD,% | 57.13 ± 0.59 b | 63.44 ± 0.44 a | 64.54 ± 0.43 a | <0.001 | <0.001 |
| IVNDFD, % | 49.83 ± 0.76 c | 54.47 ± 0.83 b | 57.40 ± 0.68 a | <0.001 | <0.001 |
| Acetate, mmol/L | 54.29 ± 8.54 | 51.64 ± 6.58 | 48.46 ± 9.22 | 0.104 | 0.457 |
| Propionate, mmol/L | 25.02 ± 0.82 | 25.03 ± 3.56 | 22.73 ± 4.10 | 0.199 | 0.363 |
| Butyrate, mmol/L | 0.67 ± 0.1 | 0.66 ± 0.12 | 0.63 ± 0.15 | 0.428 | 0.516 |
| Isobutyric, mmol/L | 9.87 ± 8,69 | 9.97 ± 1.44 | 9.42 ± 1.53 | 0.513 | 0.808 |
| Isovaleric, mmol/L | 1.17 ± 0.17 | 1.15 ± 0.23 | 1.11 ± 0.26 | 0.589 | 0.893 |
| Valeric, mmol/L | 1.83 ± 0.28 | 1.82 ± 0.26 | 1.78 ± 0.29 | 0.685 | 0.911 |
| Hexanoic, mmol/L | 0.32 ± 0.03 | 0.33 ± 0.01 | 0.31 ± 0.05 | 0.864 | 0.998 |
| Total VFA, mmol/L | 93.18 ± 1.24 | 90.58 ± 1.47 | 92.23 ± 1.95 | 0.826 | 0.961 |
| CH4 mmol/L | 21.50 ± 1.58 | 21.24 ± 2.54 | 19.32 ± 3.52 | 0.129 | 0.495 |
| CO2 mmol/L | 48.21 ± 2.03 | 48.08 ± 6.11 | 44.04 ± 8.85 | 0.152 | 0.446 |
| Acet/Prop | 2.17 ± 0.41 | 2.15 ± 0.1 | 2.11 ± 0.16 | 0.326 | 0.828 |
| NH3-N, mg/dL | 7.99 ± 1.66 a | 7.54 ± 1.61 b | 7.16 ± 1.39 b | 0.041 | 0.037 |
| Parameters | Level of Black Soldier Fly Larvae | ||||
|---|---|---|---|---|---|
| TMR0 | TMR20 | TMR40 | Linear | Quadratic | |
| ME | 9.51 ± 0.11 | 9.61 ± 0.15 | 9.59 ± 0.19 | 0.412 | 0.322 |
| NEL | 5.21 ± 0.03 | 5.32 ± 0.02 | 5.36 ± 0.02 | 0.075 | 0.062 |
| Lag time, h | 4.34 ± 2.5 | 4.71 ± 2.5 | 4.25 ± 2.0 | 0.917 | 0.592 |
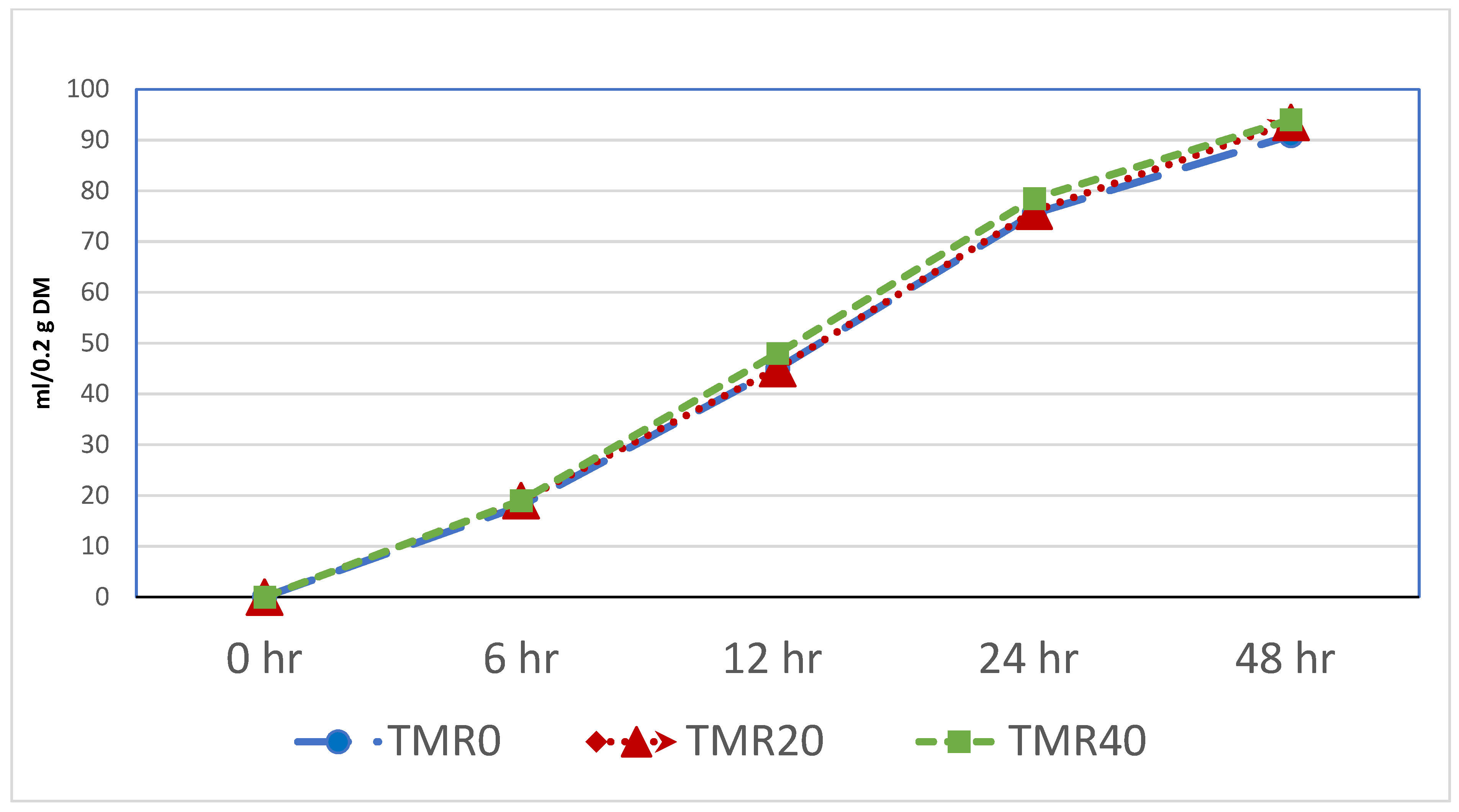
| 6 h gas, mL | 18.78 ± 1.5 | 19.44 ± 1.7 | 19.38 ± 0.9 | 0.764 | 0.837 |
| 12 h gas, mL | 45.33 ± 1.5 | 45.52 ± 1.8 | 48.09 ± 0.9 | 0.334 | 0.464 |
| 24 h gas, mL | 72.79 ± 1.5 | 73.97 ± 2.7 | 74.50 ± 1.1 | 0.132 | 0.125 |
| 48 h gas, mL | 83.58 ± 2.8 b | 92.82 ± 3.1 a | 90.66 ± 3.8 a | 0.041 | 0.023 |
| MBP 24 h, mg/g | 102.66 ± 5.7 | 109.60 ± 6.6 | 114.93 ± 7.9 | 0.457 | 0.924 |
| MBP 48 h, mg/g | 192.53 ± 12.3 | 201.21 ± 21.4 | 203.34 ± 16.8 | 0.354 | 0.402 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kahraman, O.; Gülşen, N.; İnal, F.; Alataş, M.S.; İnanç, Z.S.; Ahmed, İ.; Şişman, D.; Küçük, A.E. Comparative Analysis of In Vitro Fermentation Parameters in Total Mixed Rations of Dairy Cows with Varied Levels of Defatted Black Soldier Fly Larvae (Hermetia illucens) as a Substitute for Soybean Meal. Fermentation 2023, 9, 652. https://doi.org/10.3390/fermentation9070652
Kahraman O, Gülşen N, İnal F, Alataş MS, İnanç ZS, Ahmed İ, Şişman D, Küçük AE. Comparative Analysis of In Vitro Fermentation Parameters in Total Mixed Rations of Dairy Cows with Varied Levels of Defatted Black Soldier Fly Larvae (Hermetia illucens) as a Substitute for Soybean Meal. Fermentation. 2023; 9(7):652. https://doi.org/10.3390/fermentation9070652
Chicago/Turabian StyleKahraman, Oğuzhan, Nurettin Gülşen, Fatma İnal, Mustafa Selçuk Alataş, Zekeriya Safa İnanç, İbrar Ahmed, Deniz Şişman, and Atalay Enes Küçük. 2023. "Comparative Analysis of In Vitro Fermentation Parameters in Total Mixed Rations of Dairy Cows with Varied Levels of Defatted Black Soldier Fly Larvae (Hermetia illucens) as a Substitute for Soybean Meal" Fermentation 9, no. 7: 652. https://doi.org/10.3390/fermentation9070652
APA StyleKahraman, O., Gülşen, N., İnal, F., Alataş, M. S., İnanç, Z. S., Ahmed, İ., Şişman, D., & Küçük, A. E. (2023). Comparative Analysis of In Vitro Fermentation Parameters in Total Mixed Rations of Dairy Cows with Varied Levels of Defatted Black Soldier Fly Larvae (Hermetia illucens) as a Substitute for Soybean Meal. Fermentation, 9(7), 652. https://doi.org/10.3390/fermentation9070652