
Bio-Drying of Municipal Wastewater Sludge: Effects of High Temperature, Low Moisture Content and Volatile Compounds on the Microbial Community

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Substrate
2.2. Experimental Setup
2.3. Physicochemical Studies
2.4. Microbial Counting with Real-Time PCR (qPCR-RT)
2.5. Profiling of Prokaryotic and Fungal Communities Based on 16S rRNA Gene and Internal Transcribed Spacer (ITS)
2.6. Statistical Data Analysis
3. Results
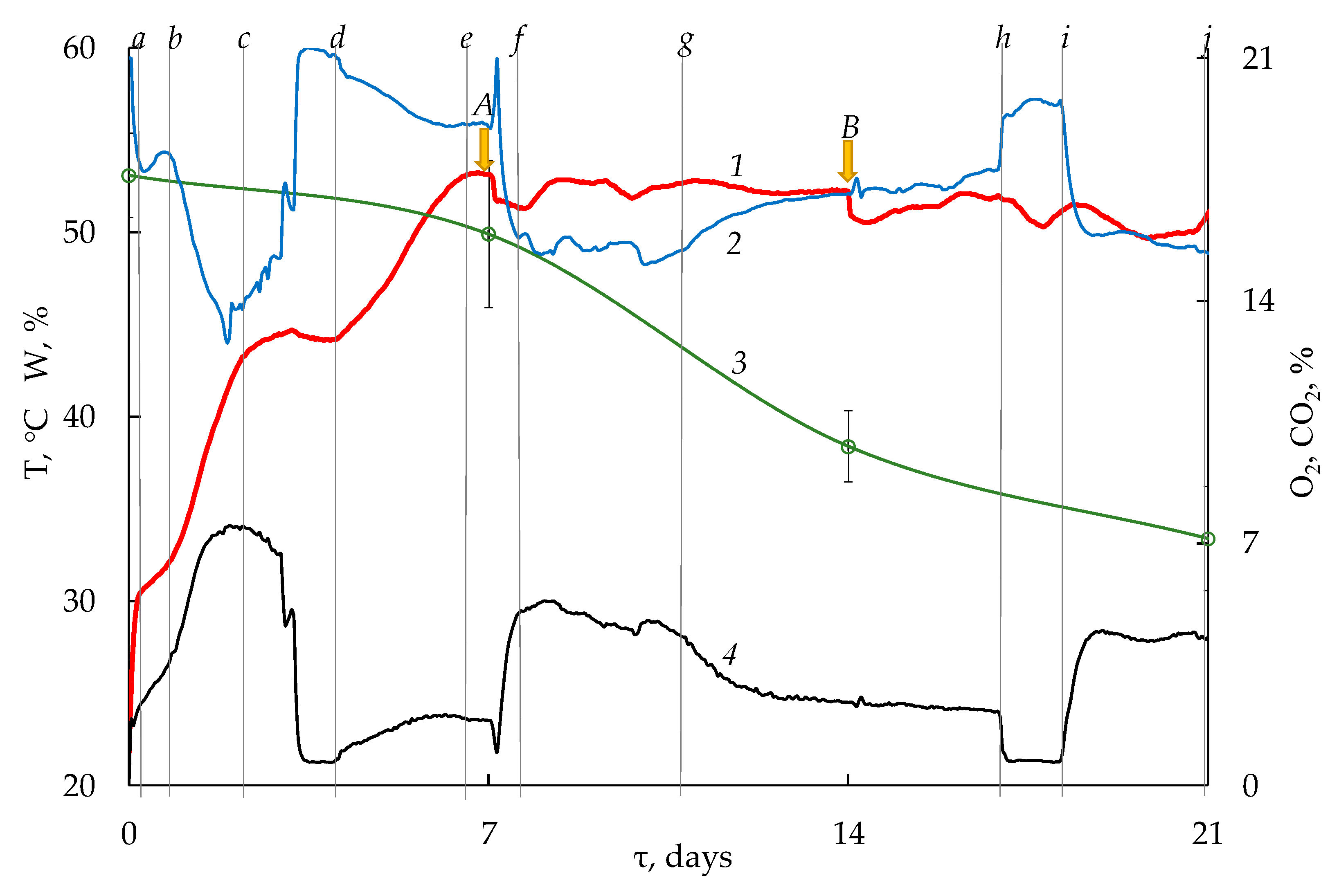
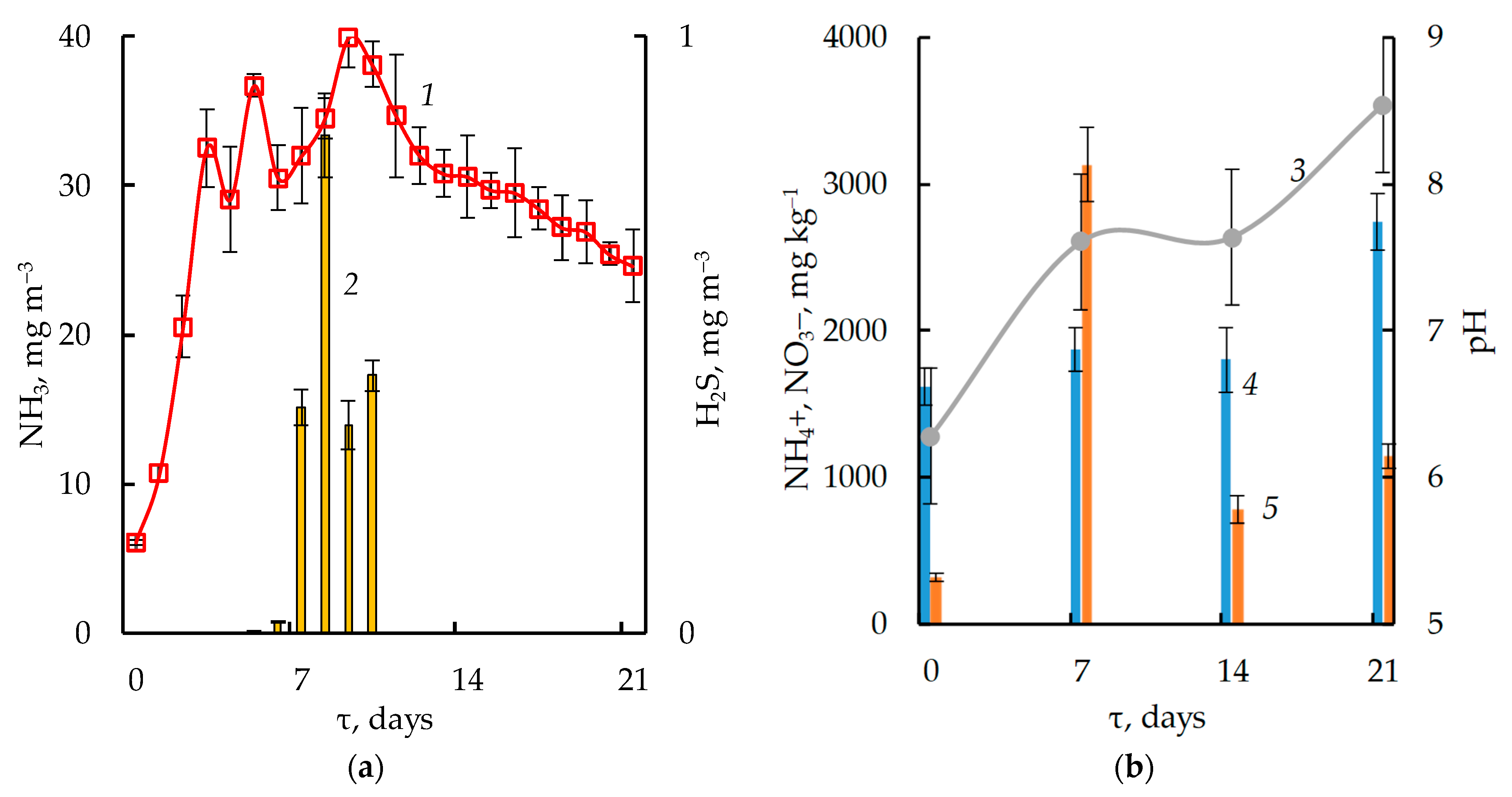
3.1. Dynamics of Physicochemical Parameters
3.2. Molecular Genetic Research on the Taxonomic Diversity of Microbiota of the Substrate during Bio-Drying
3.2.1. General Parameters of the Microbial Community
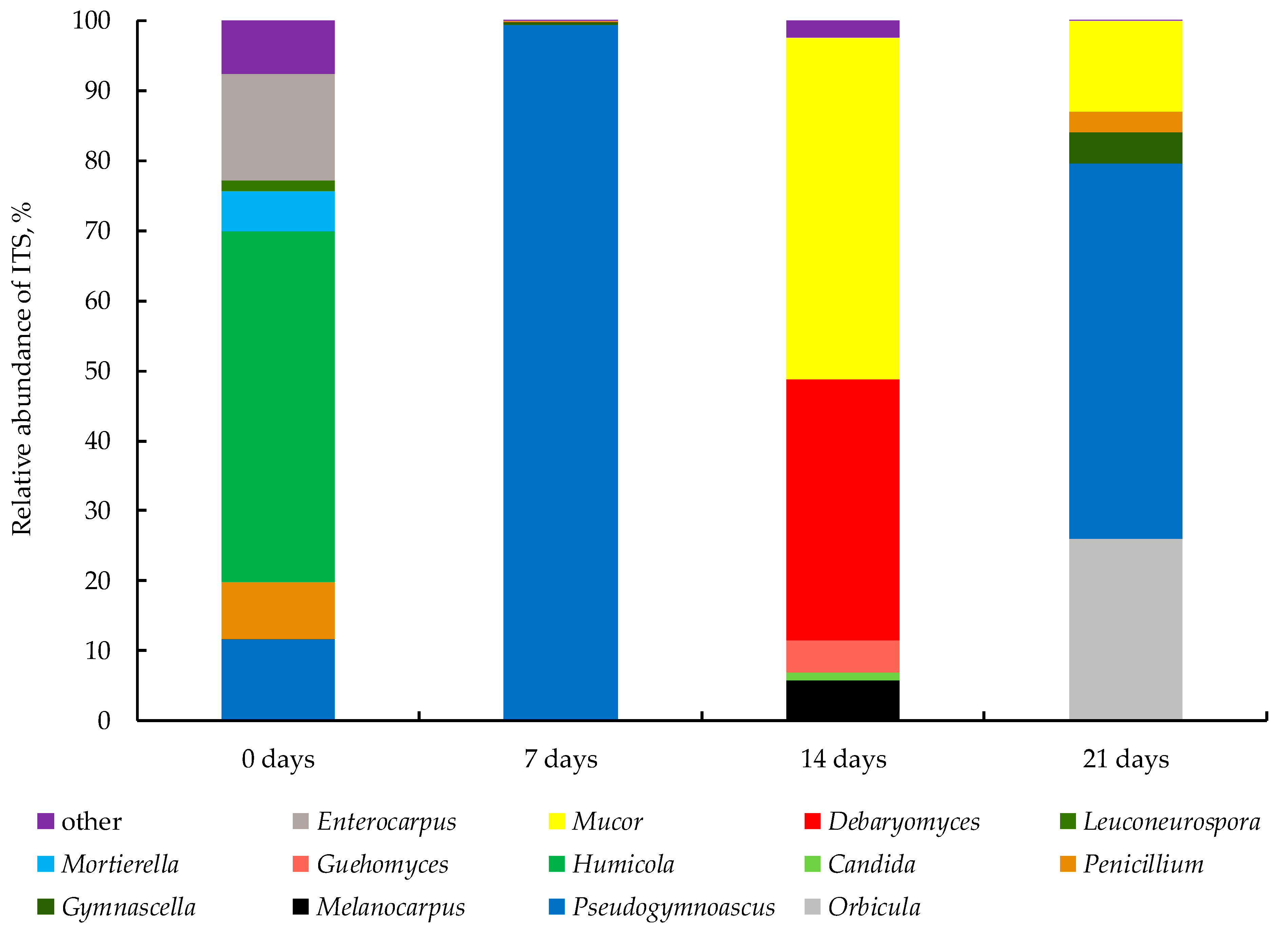
3.2.2. Fungal Community of WS during Bio-Drying
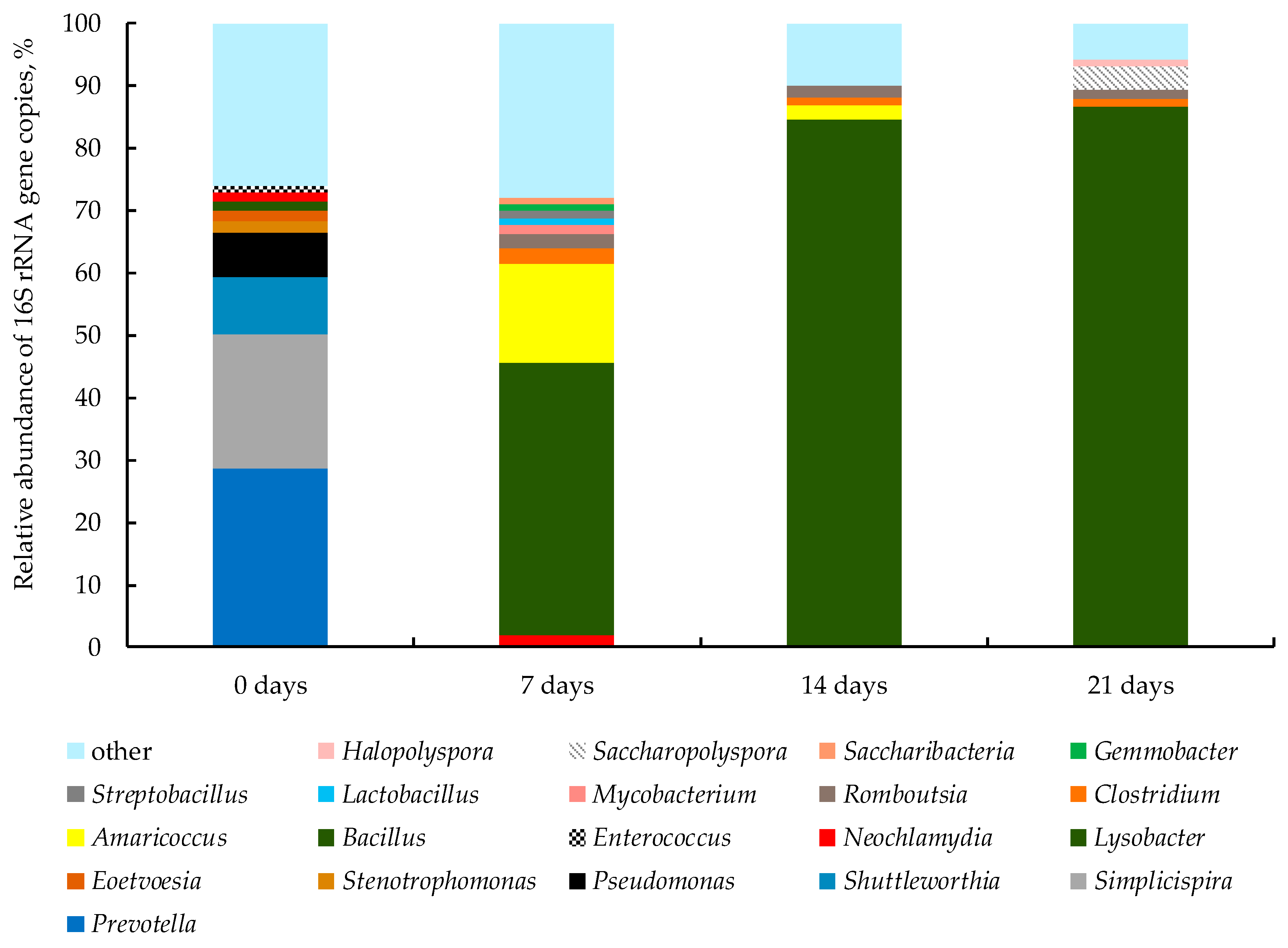
3.2.3. Bacterial Community of WS during Bio-Drying
3.2.4. Taxonomic Diversity of Biofilter Microbiota
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Business Guide. V Budushee bez Osadka 28.09.2018 [To the Future without Sediment. 28.09.2018]. Available online: https://www.kommersant.ru/doc/3745197 (accessed on 12 April 2023).
- Muscarella, S.M.; Badalucco, L.; Armando Laudicina, V.; Wang, Z.; Mannina, G. Chapter 6—Wastewater treatment sludge composting. In Current Developments in Biotechnology and Bioengineering; Mannina, G., Pandey, A., Sirohi, R., Eds.; Elsevier: Amsterdam, The Netherlands, 2023; pp. 115–136. [Google Scholar] [CrossRef]
- González, D.; Guerra, N.; Colón, J.; Gabriel, D.; Ponsá, S.; Sánchez, A. Filling in sewage sludge biodrying gaps: Greenhouse gases, volatile organic compounds and odour emissions. Bioresour. Technol. 2019, 291, 121857. [Google Scholar] [CrossRef] [PubMed]
- Ogwueleka, T.C.; Ofeoshi, C.I.; Ubah, J.I. Application of bio-drying technique for effective moisture reduction and disposal of sewage sludge in the framework of water-energy nexus. Energy Nexus 2021, 4, 100028. [Google Scholar] [CrossRef]
- Cai, L.; Chen, T.-B.; Gao, D.; Yu, J. Bacterial communities and their association with the bio-drying of sewage sludge. Water Res. 2016, 90, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Kiselev, A.; Magaril, E.; Magaril, R.; Panepinto, D.; Ravina, M.; Zanetti, M.C. Towards Circular Economy: Evaluation of Sewage Sludge Biogas Solutions. Resources 2019, 8, 91. [Google Scholar] [CrossRef]
- Interfax. “Rosvodokanal” Vnedryaet v Regionah Tehnologiu Utilizacii OSV 24.12.2021 [Rosvodokanal Introduces Sewage Sludge Disposal Technologies in the Regions 24 December 2021]. Available online: https://www.interfax.ru/presscenter/812437 (accessed on 12 April 2023).
- Ma, J.; Mu, L.; Zhang, Z.; Wang, Z.; Kong, W.; Feng, S.; Li, A.; Shen, B.; Zhang, L. Influence of thermal assistance on the biodegradation of organics during food waste bio-drying: Microbial stimulation and energy assessment. Chemosphere 2021, 272, 129875. [Google Scholar] [CrossRef]
- Adams, J.D.W.; Frostick, L.E. Analysis of bacterial activity, biomass and diversity during windrow composting. Waste Manag. 2009, 29, 598–605. [Google Scholar] [CrossRef]
- Lü, H.; Chen, X.-H.; Mo, C.-H.; Huang, Y.-H.; He, M.-Y.; Li, Y.-W.; Feng, N.-X.; Katsoyiannis, A.; Cai, Q.-Y. Occurrence and dissipation mechanism of organic pollutants during the composting of sewage sludge: A critical review. Bioresour. Technol. 2021, 328, 124847. [Google Scholar] [CrossRef]
- Chen, Z.; Li, Y.; Peng, Y.; Mironov, V.; Chen, J.; Jin, H.; Zhang, S. Feasibility of sewage sludge and food waste aerobic co-composting: Physicochemical properties, microbial community structures, and contradiction between microbial metabolic activity and safety risks. Sci. Total Environ. 2022, 825, 154047. [Google Scholar] [CrossRef]
- Mironov, V.V.; Bochkova, E.A.; Gannesen, A.V.; Vanteeva, A.V.; Russkova, Y.I.; Nozhevnikova, A.N. Dynamics of Biological Processes during Composting of Anaerobically Digested Wastewater Sludge. Microbiology 2020, 89, 470–482. [Google Scholar] [CrossRef]
- Mironov, V.; Vanteeva, A.; Merkel, A. Microbiological Activity during Co-Composting of Food and Agricultural Waste for Soil Amendment. Agronomy 2021, 11, 928. [Google Scholar] [CrossRef]
- Mironov, V.; Vanteeva, A.; Sokolova, D.; Merkel, A.; Nikolaev, Y. Microbiota Dynamics of Mechanically Separated Organic Fraction of Municipal Solid Waste during Composting. Microorganisms 2021, 9, 1877. [Google Scholar] [CrossRef]
- Bonito, G.; Isikhuemhen, O.S.; Vilgalys, R. Identification of fungi associated with municipal compost using DNA-based techniques. Bioresour. Technol. 2010, 101, 1021–1027. [Google Scholar] [CrossRef]
- Yamamoto, N.; Asano, R.; Yoshii, H.; Otawa, K.; Nakai, N. Archael community dynamics and detection of ammonia-oxidizing archaea during composting of cattle manure using culture-independent DNA analysis. Appl. Microbiol. Biotechnol. 2011, 90, 1501–1510. [Google Scholar] [CrossRef]
- Wang, K.; Mao, H.; Li, X. Functional characteristics and influence factors of microbial community in sewage sludge composting with inorganic bulking agent. Bioresour. Technol. 2018, 249, 527–535. [Google Scholar] [CrossRef]
- Galitskaya, P.; Biktasheva, L.; Saveliev, A.; Grigoryeva, T.; Boulygina, E.; Selivanovskaya, S. Fungal and bacterial successions in the process of co-composting of organic wastes as revealed by 454 pyrosequencing. PLoS ONE 2017, 12, e0186051. [Google Scholar] [CrossRef]
- Nakasaki, K.; Hirai, H.; Mimoto, H.; Quyen, T.N.M.; Koyama, M.; Takeda, K. Succession of microbial community during vigorous organic matter degradation in the primary fermentation stage of food waste composting. Sci. Total Environ. 2019, 671, 1237–1244. [Google Scholar] [CrossRef]
- Takaku, H.; Kodaira, S.; Kimoto, A.; Nashimoto, M.; Takagi, M. Microbial communities in the garbage composting with rice hull as an amendment revealed by culture-dependent and -independent approaches. J. Biosci. Bioeng. 2006, 101, 42–50. [Google Scholar] [CrossRef]
- Awasthi, M.K.; Pandey, A.K.; Bundela, P.S.; Khan, J. Co-composting of organic fraction of municipal solid waste mixed with different bulking waste: Characterization of physicochemical parameters and microbial enzymatic dynamic. Bioresour. Technol. 2015, 182, 200–207. [Google Scholar] [CrossRef]
- Voběrková, S.; Vaverková, M.D.; Burešová, A.; Adamcová, D.; Vršanská, M.; Kynický, J.; Brtnický, M.; Adam, V. Effect of inoculation with white-rot fungi and fungal consortium on the composting efficiency of municipal solid waste. Waste Manag. 2017, 61, 157–173. [Google Scholar] [CrossRef]
- Fang, Y.; Jia, X.; Chen, L.; Lin, C.; Zhang, H.; Chen, J. Effect of thermotolerant bacterial inoculation on the microbial community during sludge composting. Can. J. Microbiol. 2019, 65, 750–761. [Google Scholar] [CrossRef]
- Sarkar, S.; Banerjee, R.; Chanda, S.; Das, P.; Ganguly, S.; Pal, S. Effectiveness of inoculation with isolated Geobacillus strains in the thermophilic stage of vegetable waste composting. Bioresour. Technol. 2010, 101, 2892–2895. [Google Scholar] [CrossRef] [PubMed]
- Setlow, P. Spores of Bacillus subtilis: Their resistance to and killing by radiation, heat and chemicals. J. Appl. Microbiol. 2006, 101, 514–525. [Google Scholar] [CrossRef]
- Malinska, K.; Zabochnicka, M. Selection of bulking agents for composting of sewage sludge. Environ. Prot. Eng. 2013, 39, 91–103. [Google Scholar] [CrossRef]
- Palaniveloo, K.; Amran, M.A.; Norhashim, N.A.; Mohamad-Fauzi, N.; Peng-Hui, F.; Hui-Wen, L.; Kai-Lin, Y.; Jiale, L.; Chian-Yee, M.G.; Jing-Yi, L.; et al. Food Waste Composting and Microbial Community Structure Profiling. Processes 2020, 8, 723. [Google Scholar] [CrossRef]
- Martínez-Valdez, F.J.; Martínez-Ramírez, C.; Martínez-Montiel, L.; Favela-Torres, E.; Soto-Cruz, N.O.; Ramírez-Vives, F.; Saucedo-Castañeda, G. Rapid mineralisation of the Organic Fraction of Municipal Solid Waste. Bioresour. Technol. 2015, 180, 112–118. [Google Scholar] [CrossRef] [PubMed]
- SanPiN 1.2.3685-21; Hygienic Standards and Requirements for Safety and (or) Harmlessness for Human Environmental Factors, Approved. Decree of the Chief State Sanitary Doctor of the Russian Federation: Moscow, Russia, 2021.
- Ivankin, A.; Pandya, U.; Saraf, M. Intensification of Aerobic Processing of the Organic Wastes into Compost. In Maheshwari D. Composting for Sustainable Agriculture. Sustainable Development and Biodiversity; Springer International Publishing: Cham, Switzerland, 2014; Volume 3, pp. 23–42. [Google Scholar]
- Navarro, A.; Cegarra, J.; Roig, A.; Garcia, D. Relationships between organic matter and carbon contents of organic wastes. Bioresour. Technol. 1993, 44, 203–207. [Google Scholar] [CrossRef]
- Banegas, V.; Moreno, J.L.; Moreno, J.I.; Garcia, C.; Leon, G.; Hernandez, T. Composting anaerobic and aerobic sewage sludges using two proportions of sawdust. Waste Manag. 2007, 27, 1317–1327. [Google Scholar] [CrossRef]
- Cesaro, A.; Belgiorno, V.; Guida, M. Compost from organic solid waste: Quality assessment and European regulations for its sustainable use. Resour. Conserv. Recycl. 2015, 94, 72–79. [Google Scholar] [CrossRef]
- Zorpas, A.A.; Loizidou, M. Sawdust and natural zeolite as a bulking agent for improving quality of a composting product from anaerobically stabilized sewage sludge. Bioresour. Technol. 2008, 99, 7545–7552. [Google Scholar] [CrossRef]
- Bengtson, P.; Sterngren, A.E.; Rousk, J. Archaeal abundance across a pH gradient in an arable soil and its relationship to bacterial and fungal growth rates. Appl. Environ. Microbiol. 2012, 78, 5906–5911. [Google Scholar] [CrossRef]
- Fierer, N.; Jackson, J.A.; Vilgalys, R.; Jackson, R.B. Assessment of soil microbial community structure by use of taxon-specific quantitative PCR assays. Appl. Environ. Microbiol. 2005, 71, 4117–4120. [Google Scholar] [CrossRef]
- Chemidlin Prevost-Boure, N.; Christen, R.; Dequiedt, S.; Mougel, C.; Lelièvre, M.; Jolivet, C.; Shahbazkia, H.; Guillou, L.; Arrouays, D.; Ranjard, L. Validation and Application of a PCR Primer Set to Quantify Fungal Communities in the Soil Environment by Real-Time Quantitative PCR. PLoS ONE 2011, 6, e24166. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Stephen, F.; Altschul, T.L.; Madden, A.; Schäffer, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Strode, P.; Brokaw, A. Using biointeractive resources to teach Mathematics and Statistics in Biology. P Strode A Brokaw 2015, 2015, 39. [Google Scholar]
- Levasseur, A.; Piumi, F.; Coutinho, P.M.; Rancurel, C.; Asther, M.; Delattre, M.; Henrissat, B.; Pontarotti, P.; Asther, M.; Record, E. FOLy: An integrated database for the classification and functional annotation of fungal oxidoreductases potentially involved in the degradation of lignin and related aromatic compounds. Fungal Genet. Biol. 2008, 45, 638–645. [Google Scholar] [CrossRef]
- Batista-García, R.A.; Kumar, V.V.; Ariste, A.; Tovar-Herrera, O.E.; Savary, O.; Peidro-Guzmán, H.; González-Abradelo, D.; Jackson, S.A.; Dobson, A.D.W.; del Rayo Sánchez-Carbente, M.; et al. Simple Screening Protocol for Identification of Potential Mycoremediation Tools for the Elimination of Polycyclic Aromatic Hydrocarbons and Phenols from Hyperalkalophile Industrial Effluents. J. Environ. Manag. 2017, 198, 1–11. [Google Scholar] [CrossRef]
- Hossein, K.D.; Hossein, S.; Mitra, R. Isolation and identification of some bacterial and fungal isolates based on qualitative test of cellulase and ligninase activity from different sources. J. Soil Manag. Sustain. 2022, 11, 141–159. [Google Scholar] [CrossRef]
- Liu, T.; He, J.; Cui, C.; Tang, J. Exploiting community structure, interactions and functional characteristics of fungi involved in the biodrying of storage sludge and beer lees. J. Environ. Manag. 2019, 232, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Bian, B.; Shen, Y.; Hu, X.; Tian, G.; Zhang, L. Reduction of sludge by a biodrying process: Parameter optimization and mechanism. Chemosphere 2020, 248, 125970. [Google Scholar] [CrossRef] [PubMed]
- Di Piazza, S.; Houbraken, J.; Meijer, M.; Cecchi, G.; Kraak, B.; Rosa, E.; Zotti, M. Thermotolerant and Thermophilic Mycobiota in Different Steps of Compost Maturation. Microorganisms 2020, 8, 880. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Wang, B.; Zhang, D.; Chu, S.; Zhi, Y.; Hayat, K.; Wang, J.; Chen, X.; Hui, N.; Zhou, P. Streptomyces griseorubens JSD-1 promotes rice straw composting efficiency in industrial-scale fermenter: Evaluation of change in physicochemical properties and microbial community. Bioresour. Technol. 2021, 321, 124465. [Google Scholar] [CrossRef] [PubMed]
- Tortosa, G.; Torralbo, F.; Maza-Márquez, P.; Aranda, E.; Calvo, C.; González-Murua, C.; Bedmar, E.J. Assessment of the diversity and abundance of the total and active fungal population and its correlation with humification during two-phase olive mill waste (“alperujo”) composting. Bioresour. Technol. 2020, 295, 122267. [Google Scholar] [CrossRef]
- Campbell, C.; Rodgers, J.D.; Murray, T.D. A psychrophilic Orbicula associated with oat kernels. Mycologist 1991, 5, 113–114. [Google Scholar] [CrossRef]
- Deshmukh, S.K.; Verekar, S.A. Incidence of Keratinophilic Fungi from Selected Soils of Vidarbha Region of Maharashtra State, India. J. Mycol. 2014, 2014, 148970. [Google Scholar] [CrossRef]
- Steger, K.; Jarvis, Å.; Vasara, T.; Romantschuk, M.; Sundh, I. Effects of differing temperature management on development of Actinobacteria populations during composting. Res. Microbiol. 2007, 158, 617–624. [Google Scholar] [CrossRef]
- Mukjang, N.; Chitov, T.; Mhuantong, W.; Champreda, V.; Pathomaree, W.; Sattayawat, P.; Bovonsombut, S. Bacterial Communities Associated with Crude Oil Bioremediation through Composting Approaches with Indigenous Bacterial Isolate. Life 2022, 12, 1712. [Google Scholar] [CrossRef]
- Piceno, Y.M.; Pecora-Black, G.; Kramer, S.; Roy, M.; Reid, F.C.; Dubinsky, E.A.; Andersen, G.L. Bacterial community structure transformed after thermophilically composting human waste in Haiti. PLoS ONE 2017, 12, e0177626. [Google Scholar] [CrossRef]
- Noel, S.J.; Attwood, G.T.; Rakonjac, J.; Moon, C.D.; Waghorn, G.C.; Janssen, P.H. Seasonal changes in the digesta-adherent rumen bacterial communities of dairy cattle grazing pasture. PLoS ONE 2017, 12, e0173819. [Google Scholar] [CrossRef]
- Neubauer, V.; Petri, R.; Humer, E.; Kröger, I.; Mann, E.; Reisinger, N.; Wagner, M.; Zebeli, Q. High-grain diets supplemented with phytogenic compounds or autolyzed yeast modulate ruminal bacterial community and fermentation in dry cows. J. Dairy Sci. 2018, 101, 2335–2349. [Google Scholar] [CrossRef]
- Beauchamp, C.J.; Lévesque, G.; Prévost, D.; Chalifour, F.-P. Isolation of free-living dinitrogen-fixing bacteria and their activity in compost containing de-inking paper sludge. Bioresour. Technol. 2006, 97, 1002–1011. [Google Scholar] [CrossRef]
- Al-Dhabi, N.A.; Esmail, G.A.; Alzeer, A.F.; Arasu, M.V. Removal of nitrogen from wastewater of date processing industries using a Saudi Arabian mesophilic bacterium, Stenotrophomonas maltophilia Al-Dhabi-17 in sequencing batch reactor. Chemosphere 2021, 268, 128636. [Google Scholar] [CrossRef]
- Felfoldi, T.; Vengring, A.; Keki, Z.; Marialigeti, K.; Schumann, P.; Toth, E.M. Eoetvoesia caeni gen. nov., sp. nov., isolated from an activated sludge system treating coke plant effluent. Int. J. Syst. Evol. Microbiol. 2014, 64 Pt 6, 1920–1925. [Google Scholar] [CrossRef]
- Vaz-Moreira, I.; Silva, M.E.; Manaia, C.M.; Nunes, O.C. Diversity of Bacterial Isolates from Commercial and Homemade Composts. Microb. Ecol. 2007, 55, 714–722. [Google Scholar] [CrossRef]
- Zhuang, H.; Hong, X.; Han, H.; Shan, S. Effect of pure oxygen fine bubbles on the organic matter removal and bacterial community evolution treating coal gasification wastewater by membrane bioreactor. Bioresour. Technol. 2016, 221, 262–269. [Google Scholar] [CrossRef]
- Hölzel, C.S.; Schwaiger, K.; Harms, K.; Küchenhoff, H.; Kunz, A.; Meyer, K.; Müller, C.; Bauer, J. Sewage sludge and liquid pig manure as possible sources of antibiotic resistant bacteria. Environ. Res. 2010, 110, 318–326. [Google Scholar] [CrossRef]
- Ghosh, S.; Zhang, P.; Li, Y.-Q.; Setlow, P. Superdormant Spores of Bacillus Species Have Elevated Wet-Heat Resistance and Temperature Requirements for Heat Activation. J. Bacteriol. 2009, 191, 5584–5591. [Google Scholar] [CrossRef]
- Balcom, I.N.; Driscoll, H.; Vincent, J.; Leduc, M. Metagenomic analysis of an ecological wastewater treatment plant’s microbial communities and their potential to metabolize pharmaceuticals. Version 1. F1000Research 2016, 5, 1881. [Google Scholar] [CrossRef]
- Muñoz-Gómez, S.A.; Wideman, J.G.; Roger, A.J.; Slamovits, C.H. The origin of mitochondrial cristae from alphaproteobacteria. Mol. Biol. Evol. 2017, 34, 943–956. [Google Scholar] [CrossRef] [PubMed]
- Maszenan, A.M.; Seviour, R.J.; Patel, B.K.C.; Rees, G.N.; McDougall, B.M. Amaricoccus gen. nov., a Gram-Negative Coccus Occurring in Regular Packages or Tetrads, Isolated from Activated Sludge Biomass, and Descriptions of Amaricoccus veronensis sp. nov., Amaricoccus tamworthensis sp. nov., Amaricoccus macauensis sp. nov., and Amaricoccus kaplicensis sp. nov. Int. J. Syst. Bacteriol. 1997, 47, 727–734. [Google Scholar] [CrossRef]
- Rieke, E.L.; Soupir, M.L.; Moorman, T.B.; Yang, F.; Howe, A.C. Temporal Dynamics of Bacterial Communities in Soil and Leachate Water after Swine Manure Application. Front. Microbiol. 2018, 9, 3197. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Wang, Y.; Wen, Q. Effects of chlortetracycline on the fate of multi-antibiotic resistance genes and the microbial community during swine manure composting. Environ. Pollut. 2018, 237, 977–987. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.-Z.; Li, X.-X.; Zeng, Y.; Wang, S.-P.; Sun, Z.-Y.; Tang, Y.-Q. Dynamic change of bacterial community during dairy manure composting process revealed by high-throughput sequencing and advanced bioinformatics tools. Bioresour. Technol. 2020, 306, 123091. [Google Scholar] [CrossRef]
- Zhan, Y.; Zhang, Z.; Ma, T.; Zhang, X.; Wang, R.; Liu, Y.; Sun, B.; Xu, T.; Ding, G.; Wei, Y.; et al. Phosphorus excess changes rock phosphate solubilization level and bacterial community mediating phosphorus fractions mobilization during composting. Bioresour. Technol. 2021, 337, 125433. [Google Scholar] [CrossRef]
- Robledo-Mahón; Gómez-Silván, C.; Andersen, G.L.; Calvo, C.; Aranda, E. Assessment of bacterial and fungal communities in a full-scale thermophilic sewage sludge composting pile under a semipermeable cover. Bioresour. Technol. 2020, 298, 122550. [Google Scholar] [CrossRef]
- Okunishi, S.; Morita, Y.; Higuchi, T.; Maeda, H.; Nishi, K. Transformation of microflora during degradation of gaseous toluene in a biofilter detected using PCR-DGGE. J. Air Waste Manag. Assoc. 2012, 62, 748–757. [Google Scholar] [CrossRef]
- Yang, K.; Li, L.; Ding, W.; Liu, J.; Xue, S. A full-scale thermophilic biofilter in the treatment of sludge drying exhaust: Performance, microbial characteristics and bioaerosol emission. J. Chem. Technol. Biotechnol. 2018, 93, 2216–2225. [Google Scholar] [CrossRef]
- Li, L.; Han, Y.; Yan, X.; Liu, J. H2S removal and bacterial structure along a full-scale biofilter bed packed with polyurethane foam in a landfill site. Bioresour. Technol. 2013, 147, 52–58. [Google Scholar] [CrossRef]
- Liu, H.; Dai, L.; Yao, J.; Mei, Y.; Hrynsphan, D.; Tatsiana, S.; Chen, J. Efficient biotransformation of sulfide in anaerobic sequencing batch reactor by composite microbial agent: Performance optimization and microbial community analysis. Environ. Sci. Pollut. Res. Int. 2021, 28, 48718–48727. [Google Scholar] [CrossRef]
- Sun, Y.; Men, M.; Xu, B.; Meng, Q.; Bello, A.; Xu, X.; Huang, X. Assessing key microbial communities determining nitrogen transformation in composting of cow manure using illumine high-throughput sequencing. Waste Manag. 2019, 92, 59–67. [Google Scholar] [CrossRef]
- Wu, H.; Guo, C.; Yin, Z.; Quan, Y.; Yin, C. Performance and bacterial diversity of biotrickling filters filled with conductive packing material for the treatment of toluene. Bioresour. Technol. 2018, 257, 201–209. [Google Scholar] [CrossRef]
- Yasuda, T.; Waki, M.; Fukumoto, Y.; Hanajima, D.; Kuroda, K.; Suzuki, K. Characterization of the denitrifying bacterial community in a full-scale rockwool biofilter for compost waste-gas treatment. Appl. Microbiol. Biotechnol. 2017, 101, 6779–6792. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, J.; Qin, Y.; Yang, Z.; Cao, J.; Xing, Y.; Li, J. Performance and microbial community evolution of toluene degradation using a fungi-based bio-trickling filter. J. Hazard. Mater. 2019, 365, 642–649. [Google Scholar] [CrossRef]
- Vergara-Fernandez, A.; Scott, F.; Carreno-Lopez, F.; Aroca, G.; Moreno-Casas, P.; Gonzalez-Sanchez, A.; Munoz, R. A comparative assessment of the performance of fungal-bacterial and fungal biofilters for methane abatement. J. Environ. Chem. Eng. 2020, 8, 104421. [Google Scholar] [CrossRef]
- Vergara-Fernández, A.; Hernández, S.; Muñoz, R.; Revah, S. Influence of the inlet load, EBRT and mineral medium addition on spore emission by Fusarium solani in the fungal biofiltration of hydrophobic VOCs. J. Chem. Technol. Biotechnol. 2012, 87, 778–784. [Google Scholar] [CrossRef]
- Arellano-García, L.; Le Borgne, S.; Revah, S. Simultaneous treatment of dimethyl disulfide and hydrogen sulfide in an alkaline biotrickling filter. Chemosphere 2018, 191, 809–816. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
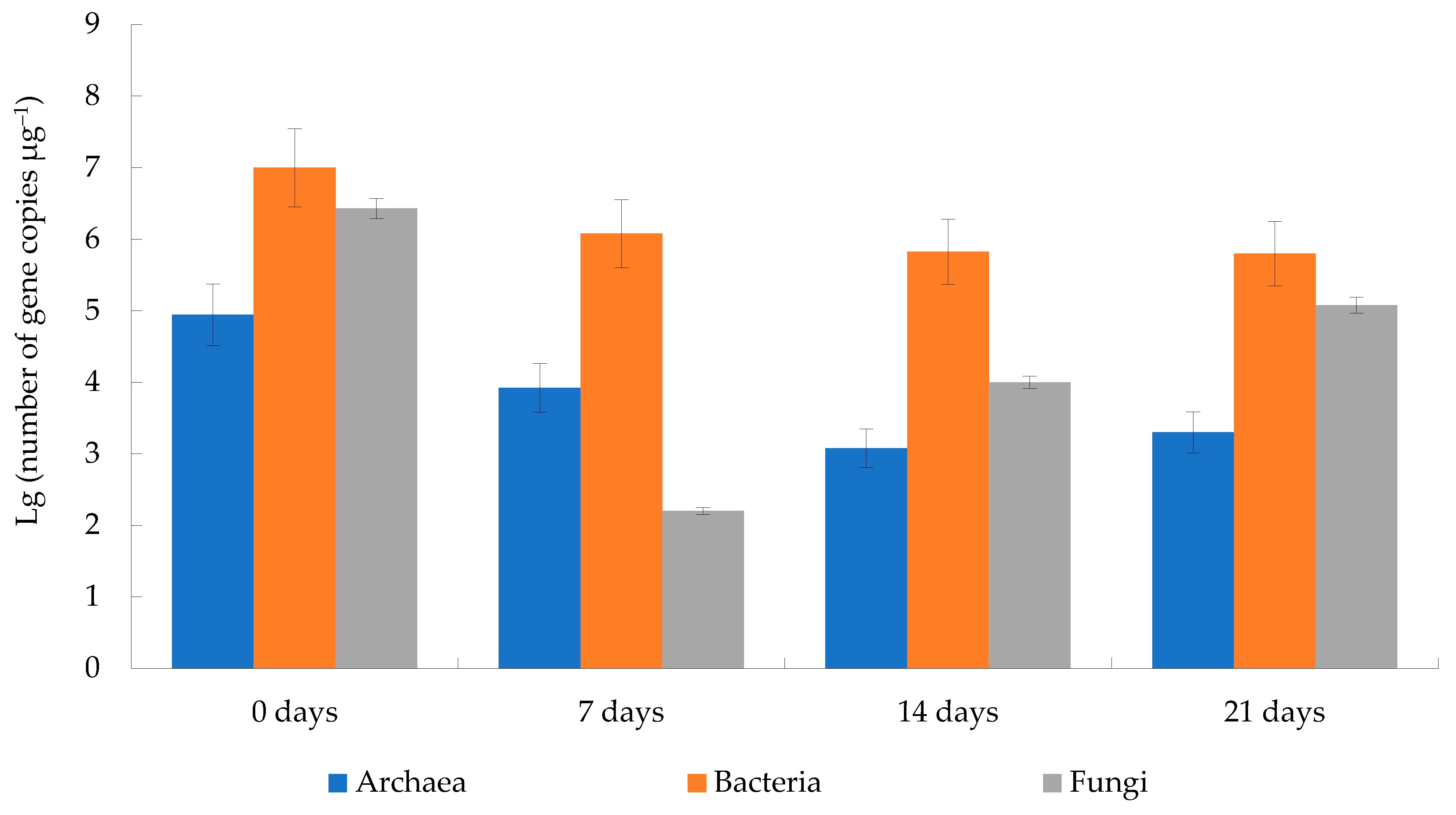{kind=link}
{kind=link}
{kind=link}
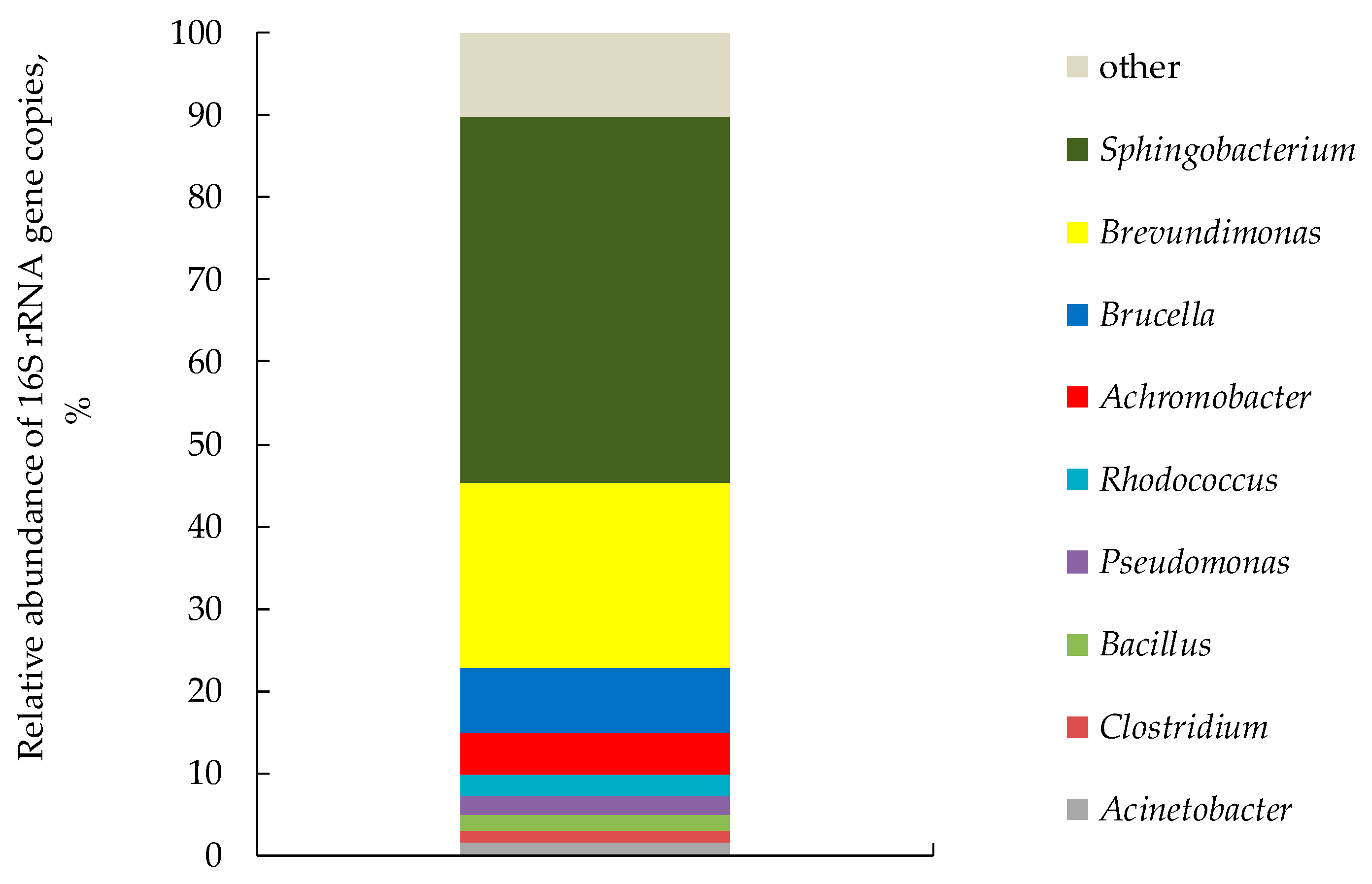{kind=link}
| Parameter | Units | Value * | Optimal Limits |
|---|---|---|---|
| pH | pH units | 6.28 ± 0.50 | 6.5–8.0 [27] |
| Moisture content | % | 53.10 ± 2.12 | 50–60% [26,27] |
| Air-filled porosity | % | 57.1 ± 0.2 | 30–60% [26] |
| Total Kjeldahl nitrogen (N) | % | 1.76 ± 0.14 | |
| NH4+ | mg kg−1 | 1618.21 ± 129.46 | |
| NO3− | mg kg−1 | 319.08 ± 25.5 | |
| Organic matter (OM) | % | 74.14 ± 1.48 | |
| Total organic carbon (C) | % | 37.07 ± 1.48 | |
| C/N | 21.33 ± 1.62 | 20–30 [26,27,28] | |
| Germination index (GI) | % | 59.56 ± 4.76 | |
| Pb | mg kg−1 | 27.30 ± 1.94 | 130 [29] |
| As | 0.0 ± 0.0 | 10 [29] | |
| Cd | 0.0 ± 0.0 | 2 [29] | |
| Ni | 5.85 ± 0.46 | 80 [29] | |
| Cr3+ | 11.31 ± 0.85 | 6 [29] | |
| Mn | 85.29 ± 5.37 | 1500 [29] | |
| Zn | 509.65 ± 25.99 | 220 [29] | |
| Cu | 37.44 ± 1.68 | 132 [29] |
| Parameter | Units | Value * |
|---|---|---|
| pH | pH units | 8.54 ± 0.60 |
| Moisture content | % | 33.40 ± 3.20 |
| Air-filled porosity | % | 70.8 ± 0.5 |
| Total Kjeldahl nitrogen (N) | % | 2.00 ± 0.14 |
| NH4+ | mg kg−1 | 2739.74 ± 191.78 |
| NO3− | mg kg−1 | 1144.59 ± 80.12 |
| Organic matter (OM) | % | 57.4 ± 1.15 |
| Total organic carbon (C) | % | 28.70 ± 1.15 |
| C/N | 14.35 ± 2.44 | |
| Germination index (GI) | % | 70.60 ± 4.94 |
| Pb | mg kg−1 | 31.28 ± 1.81 |
| As | 0.0 ± 0.0 | |
| Cd | 0.0 ± 0.0 | |
| Ni | 6.26 ± 0.36 | |
| Cr3+ | 12.10 ± 0.39 | |
| Mn | 92.80 ± 6.77 | |
| Zn | 487.99 ± 39.53 | |
| Cu | 40.98 ± 1.43 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mironov, V.; Moldon, I.; Shchelushkina, A.; Zhukov, V.; Zagustina, N. Bio-Drying of Municipal Wastewater Sludge: Effects of High Temperature, Low Moisture Content and Volatile Compounds on the Microbial Community. Fermentation 2023, 9, 570. https://doi.org/10.3390/fermentation9060570
Mironov V, Moldon I, Shchelushkina A, Zhukov V, Zagustina N. Bio-Drying of Municipal Wastewater Sludge: Effects of High Temperature, Low Moisture Content and Volatile Compounds on the Microbial Community. Fermentation. 2023; 9(6):570. https://doi.org/10.3390/fermentation9060570
Chicago/Turabian StyleMironov, Vladimir, Ivan Moldon, Anna Shchelushkina, Vitaly Zhukov, and Nataliya Zagustina. 2023. "Bio-Drying of Municipal Wastewater Sludge: Effects of High Temperature, Low Moisture Content and Volatile Compounds on the Microbial Community" Fermentation 9, no. 6: 570. https://doi.org/10.3390/fermentation9060570
APA StyleMironov, V., Moldon, I., Shchelushkina, A., Zhukov, V., & Zagustina, N. (2023). Bio-Drying of Municipal Wastewater Sludge: Effects of High Temperature, Low Moisture Content and Volatile Compounds on the Microbial Community. Fermentation, 9(6), 570. https://doi.org/10.3390/fermentation9060570