
Establishment of Flavonoid Fingerprint of TMR Diet and Optimization Factor Analysis Strategy and In Vitro Fermentation Parameters Based on Spectrum–Effect Relationship

Abstract
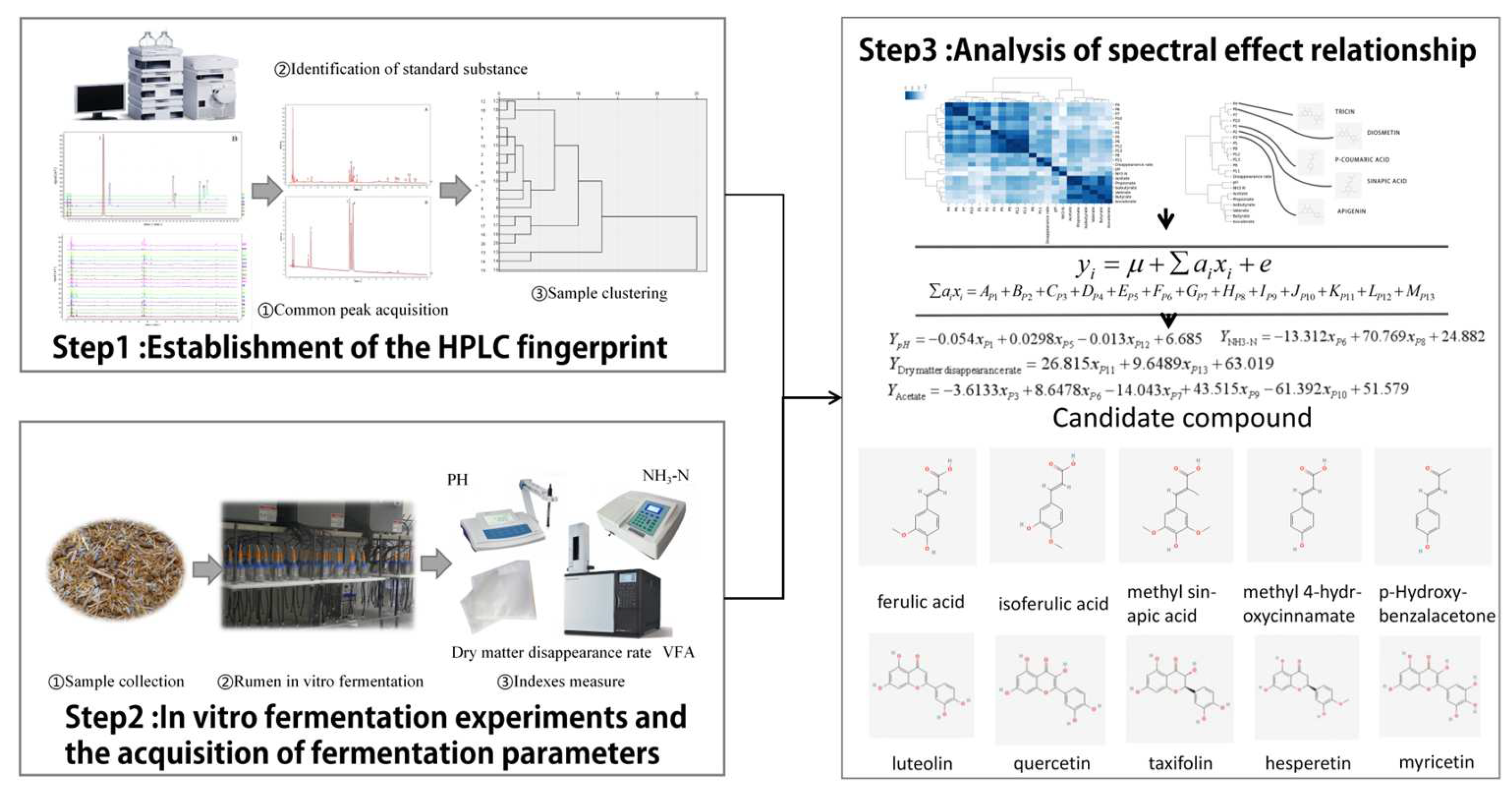
1. Introduction
2. Results
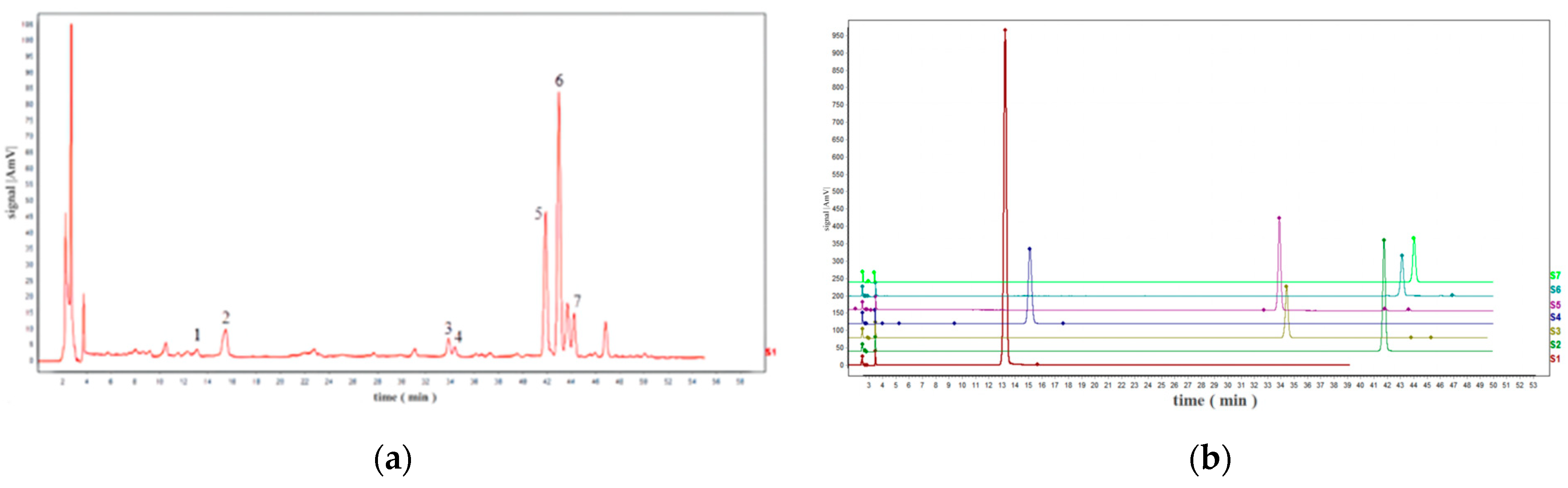
2.1. HPLC–UV Identification of Seven Standard Compounds from the TMR
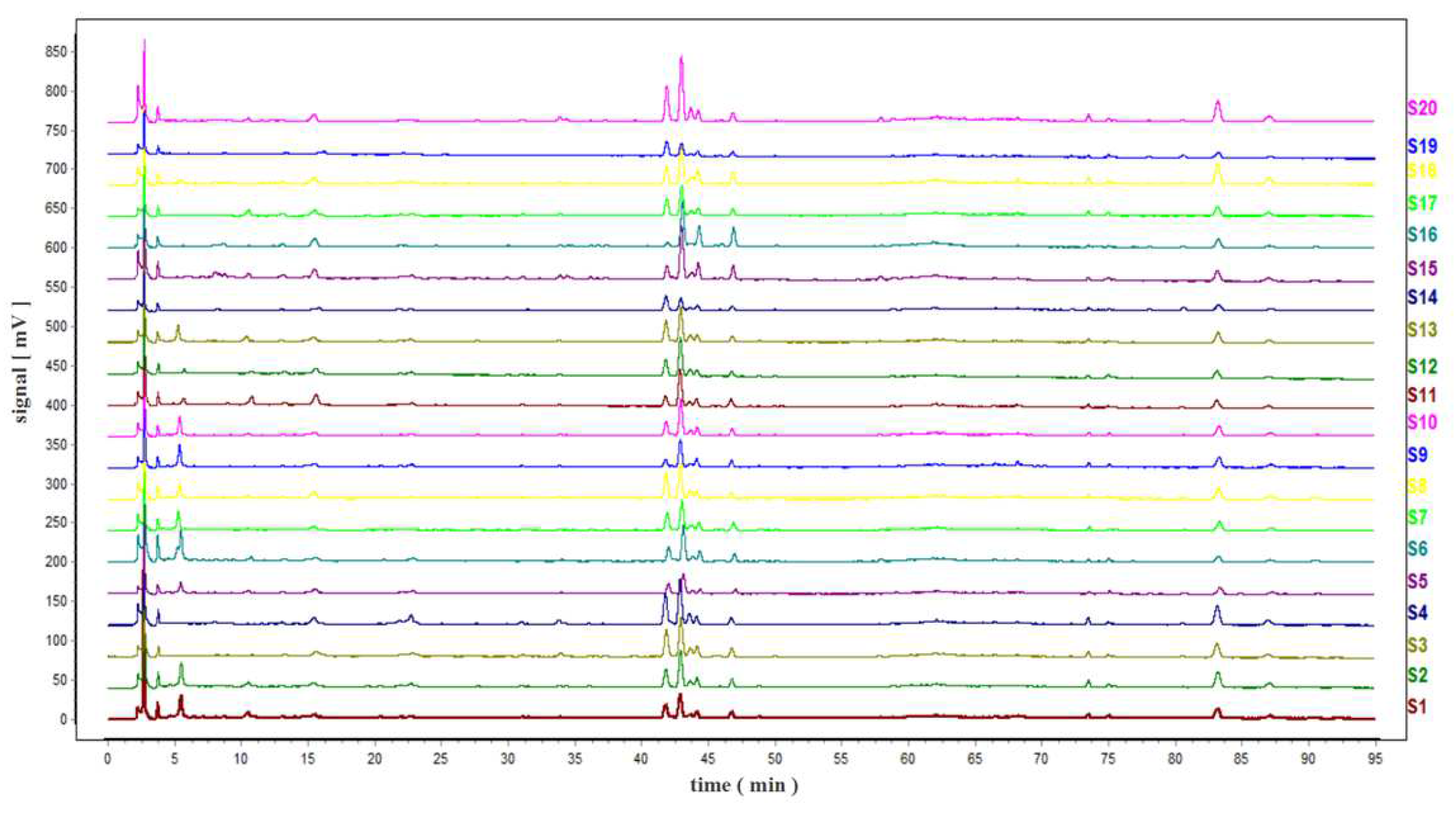
2.2. Construction of TMR Fingerprint
2.3. Confirmation of Reference Peak and Common Peak of Fingerprint
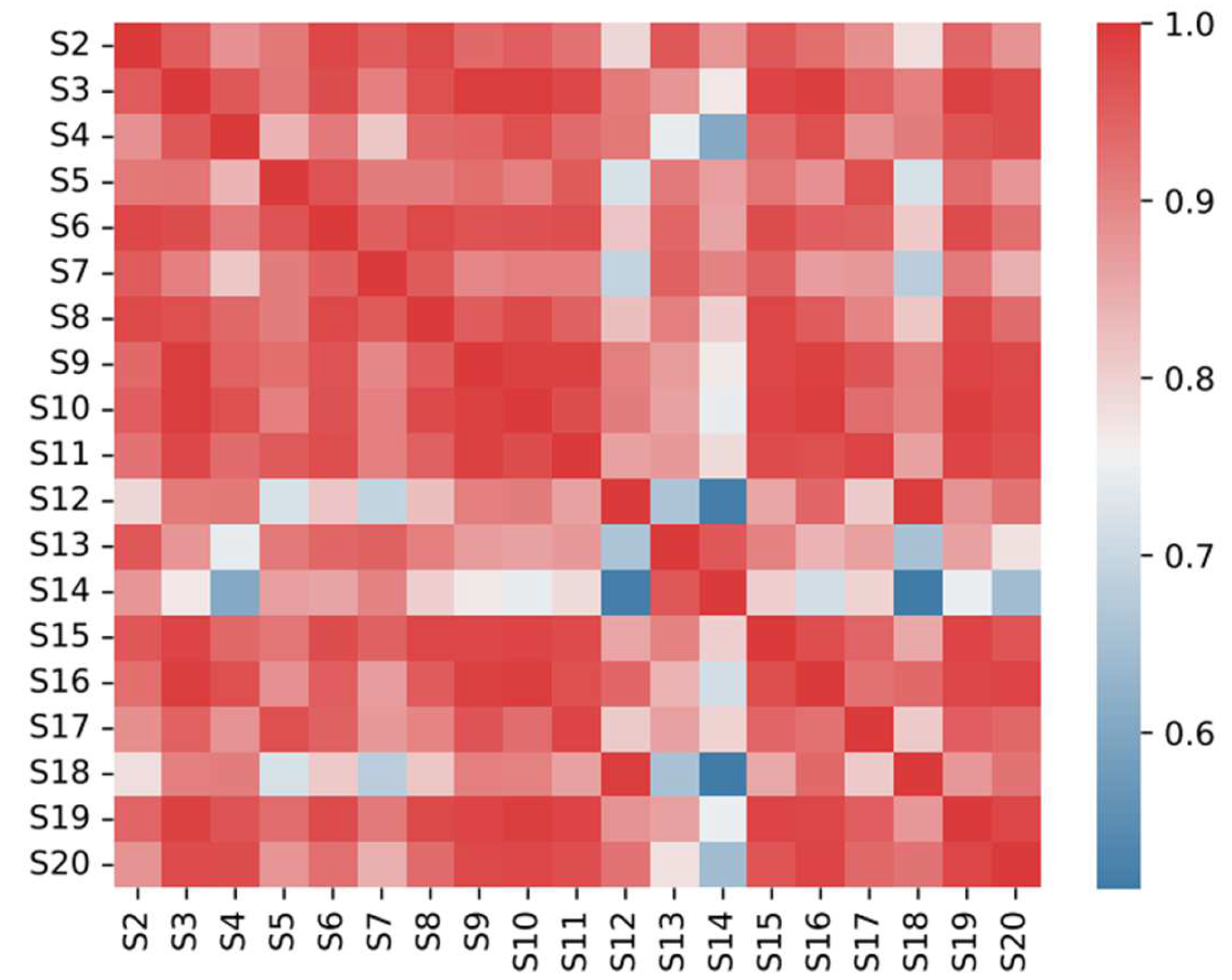
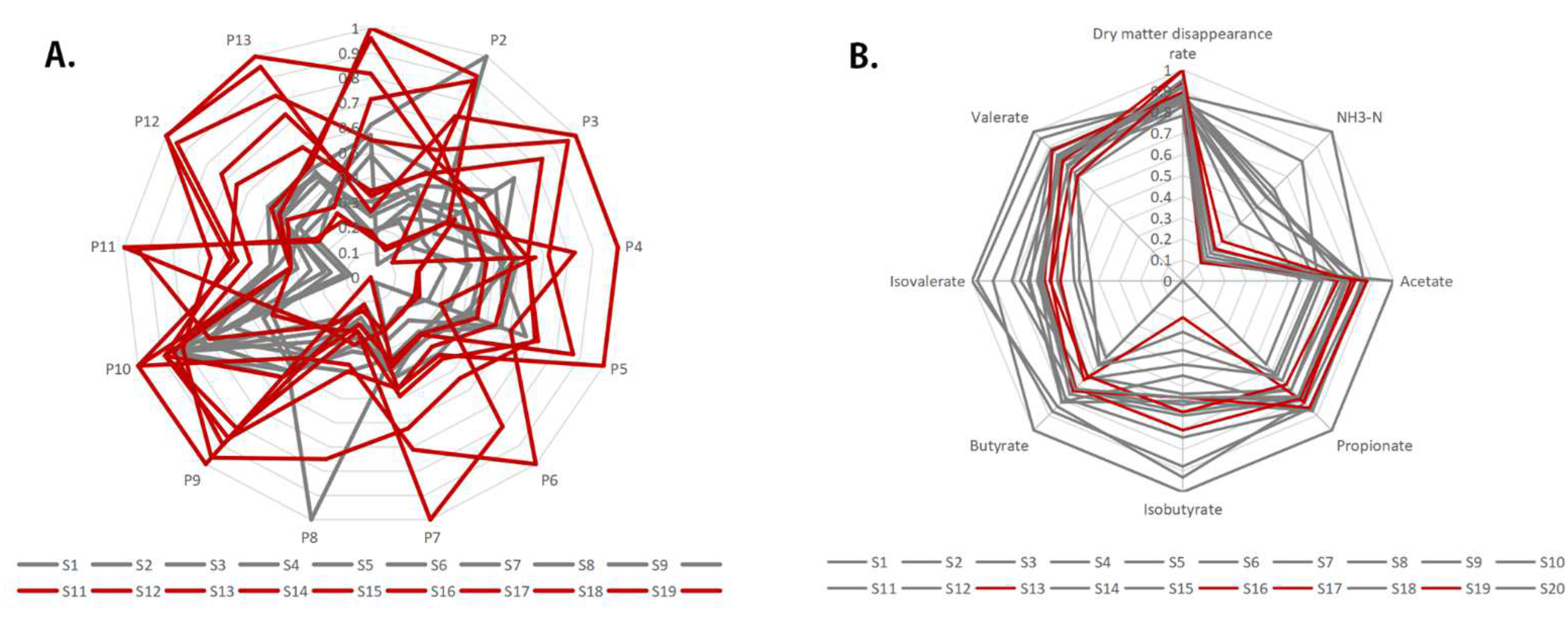
2.4. Similarity Evaluation of the HPLC Fingerprint
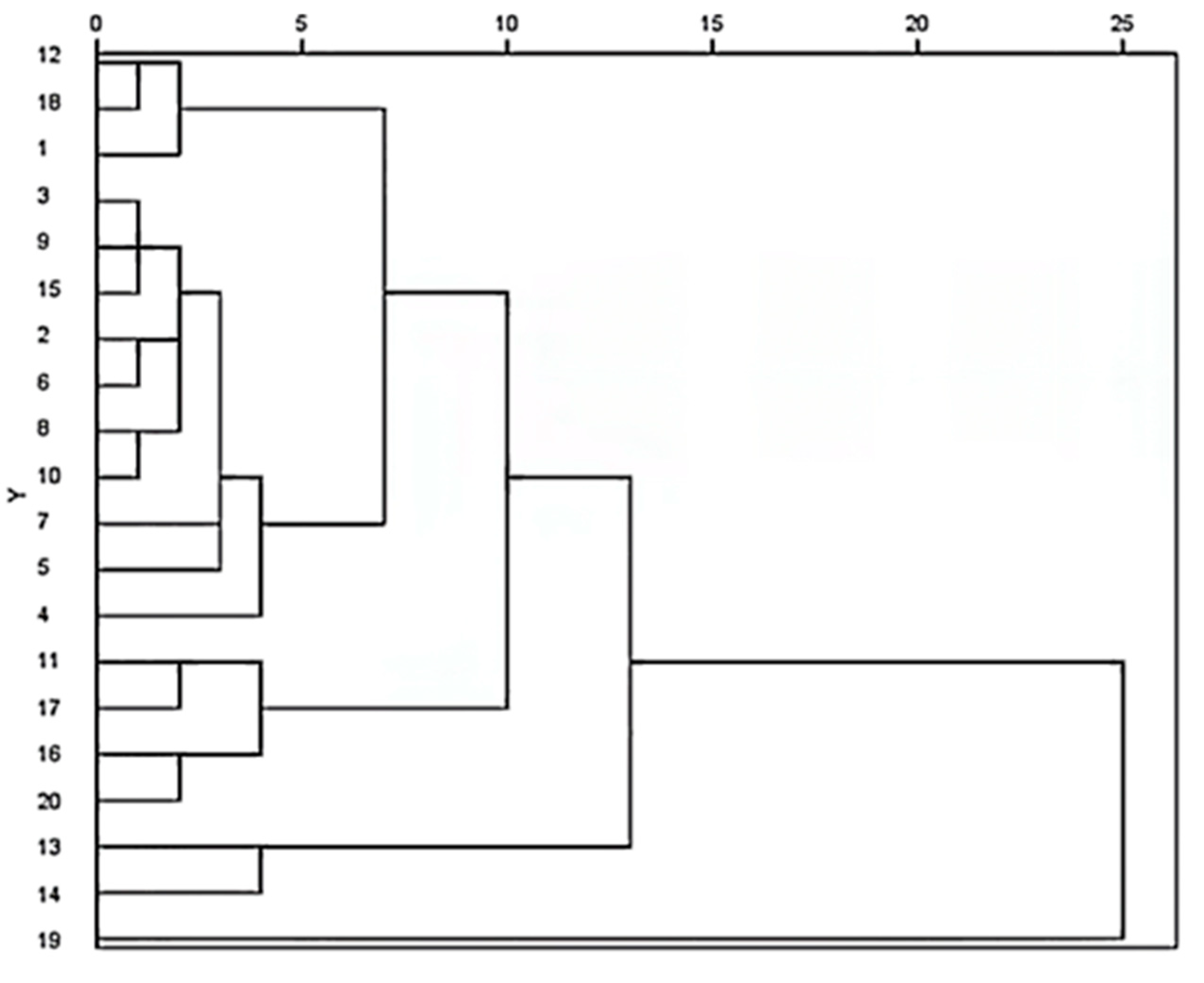
2.5. Sample Clustering Analysis
2.6. In Vitro Rumen Fermentation Results
2.6.1. pH, NH3-N Concentration, and Dry Matter Disappearance Rate of Fermentation Fluid under Different TMR
2.6.2. VFA Concentration in Fermentation Fluid under Different TMR
2.6.3. Content of Total Flavonoids in TMR
2.7. Construction of Fingerprint–Effect Relationship
2.7.1. Correlation Analysis between Total Flavonoids Content in TMR and Fermentation Parameters In Vitro
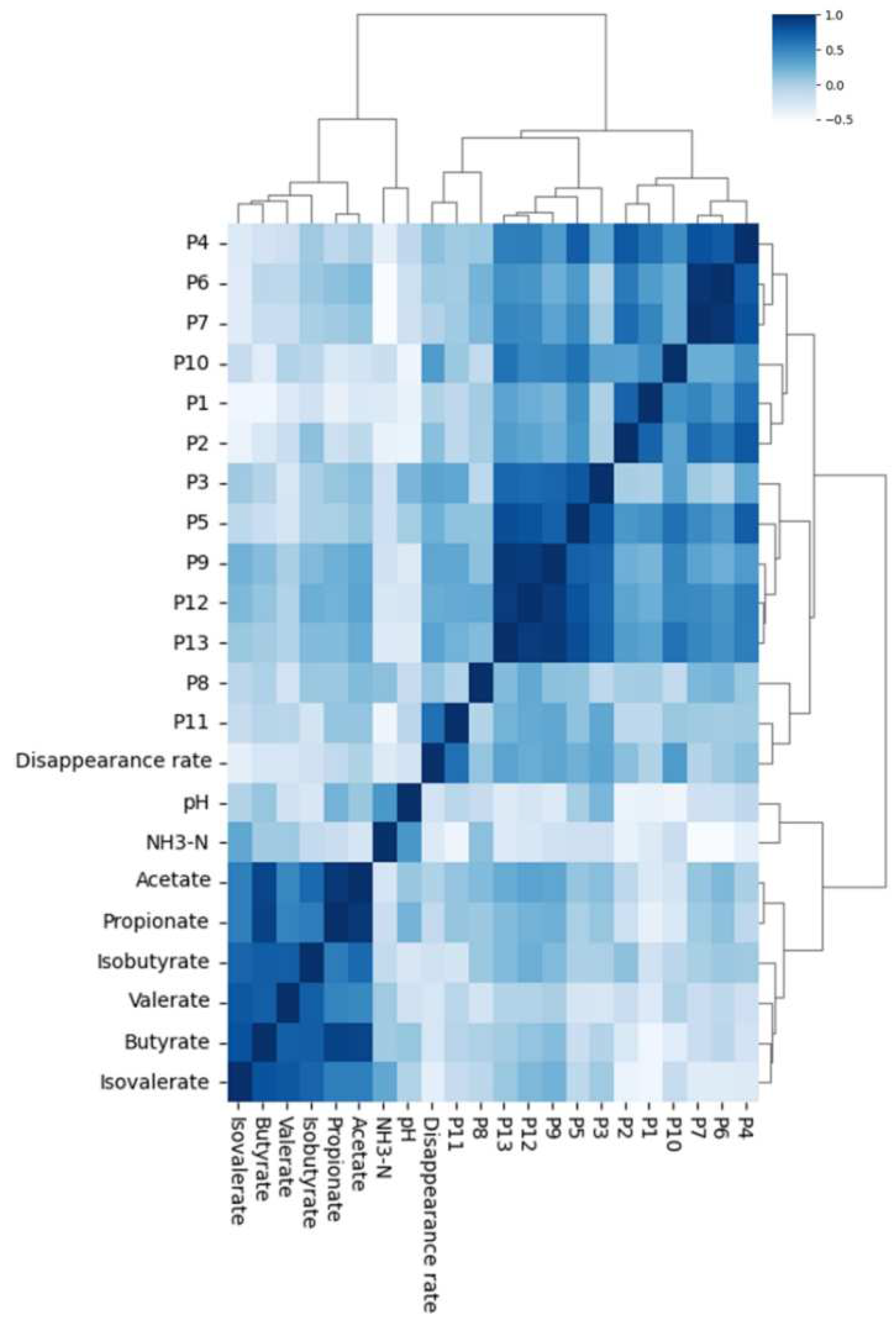
2.7.2. Hierarchical Clustering Analysis Based on Fingerprint–Effect Relationship
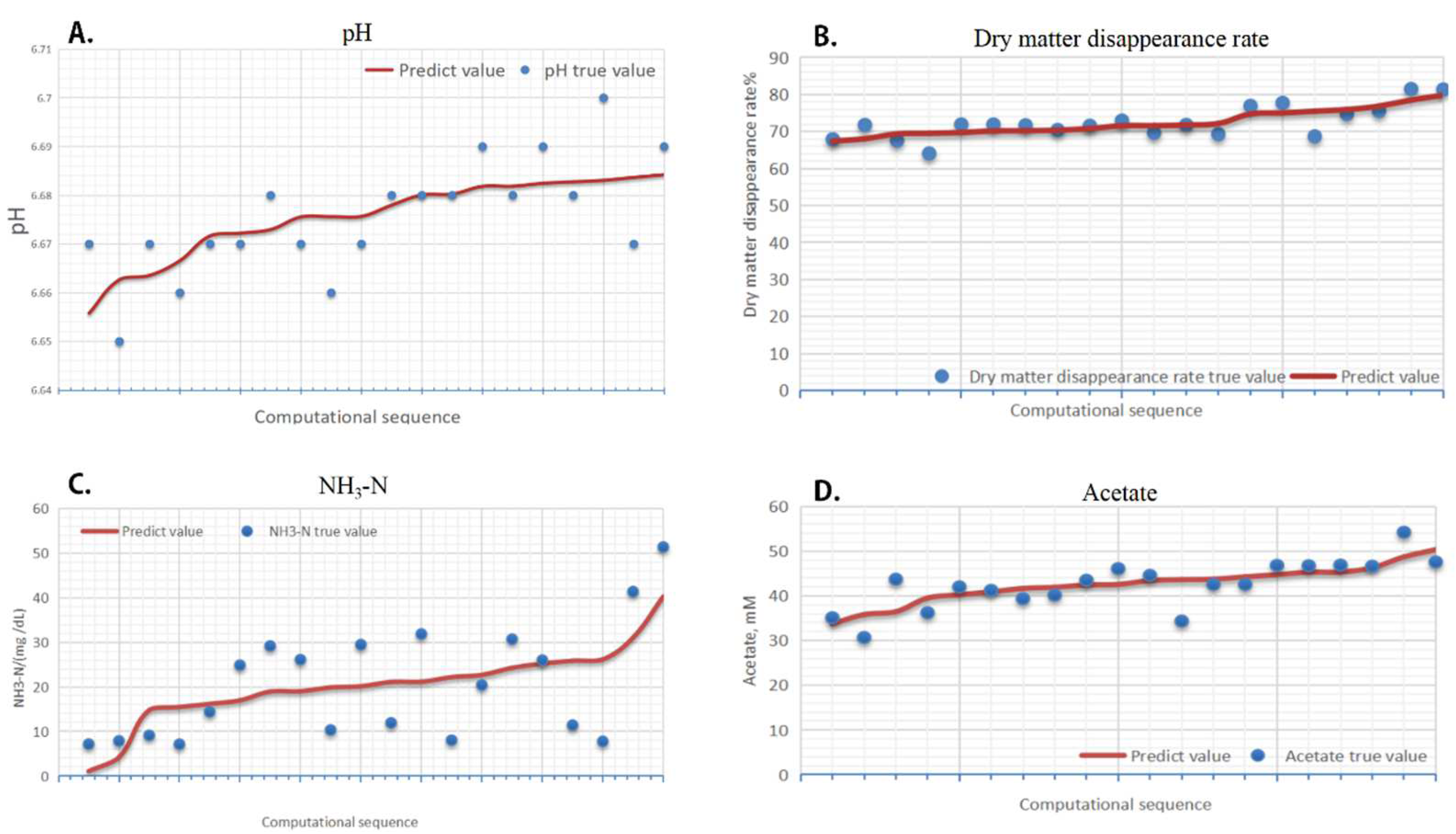
2.7.3. Principal Effect Factor Based on Optimization Fixed Model Strategy
2.7.4. Analysis of Fingerprint–Effect Relationship
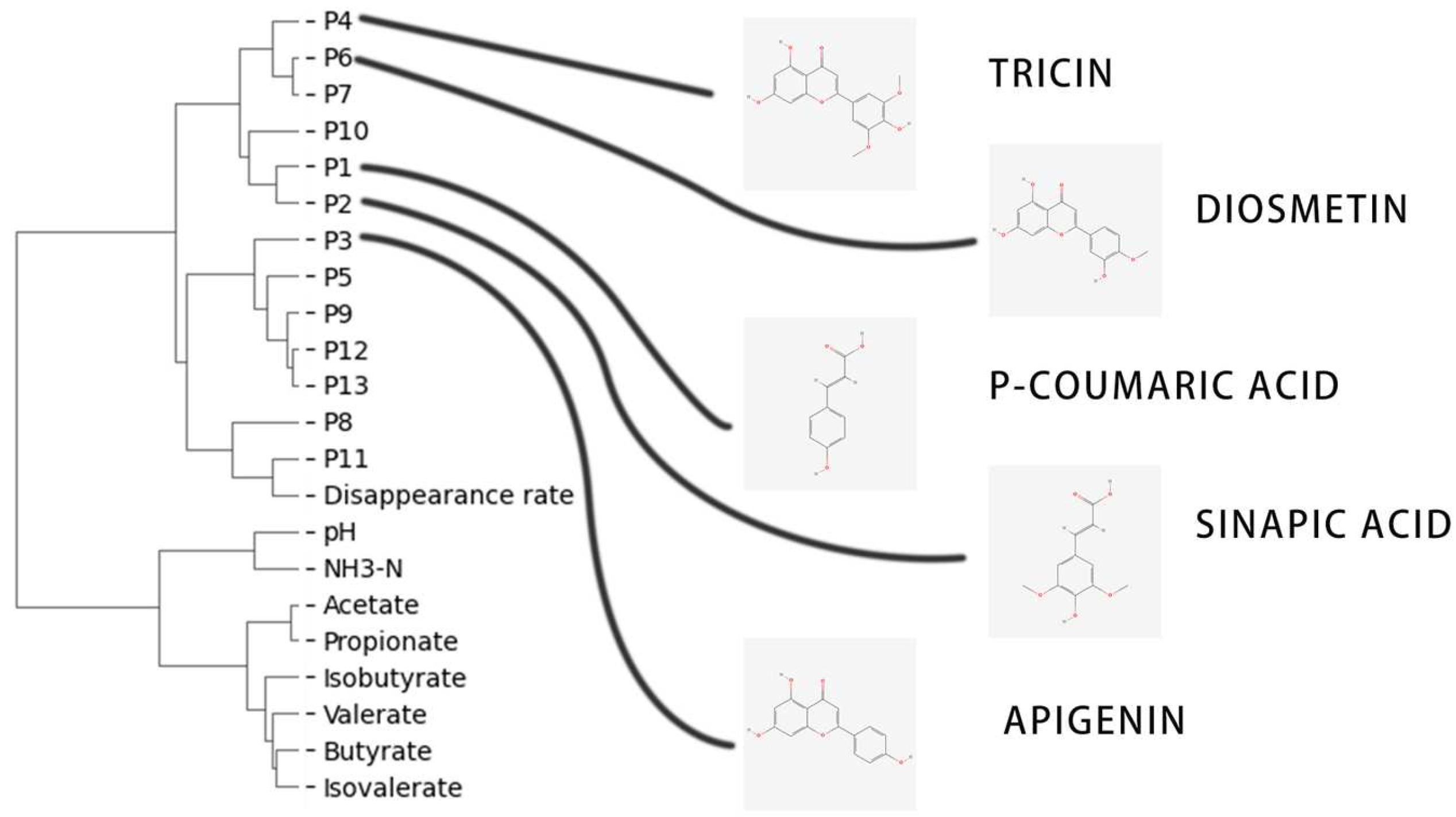
2.8. Structure–Activity Analysis Based on the Hierarchical Clustering Results
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. TMR Sample Collection
4.3. Preparation of Standard Solution and Sample Solution
4.4. Apparatus and Chromatographic Conditions
4.5. Method Validation for Quantitative Analysis
4.6. Fingerprint Similarity Evaluation and Sample Correlation Clustering Analysis
4.7. Experimental Design of Rumen Fermentation In Vitro
4.8. Sample Collection and Determination of Rumen Fermentation In Vitro
4.9. Hierarchical Clustering Analysis of Spectrum–Effect Relationships
4.10. Factor Analysis of Spectral Relationship Based on Optimization Fixed Model Strategy
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clay, N.; Garnett, T.; Lorimer, J. Dairy intensification: Drivers, impacts and alternatives. Ambio 2020, 49, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Berding, K.; Vlckova, K.; Marx, W.; Schellekens, H.; Stanton, C.; Clarke, G.; Jacka, F.; Dinan, T.G.; Cryan, J.F. Diet and the Microbiota-Gut-Brain Axis: Sowing the Seeds of Good Mental Health. Adv. Nutr. 2021, 12, 1239–1285. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Cheng, H. Health risk from veterinary antimicrobial use in China’s food animal production and its reduction. Environ. Pollut. 2016, 219, 993–997. [Google Scholar] [CrossRef]
- Nisbet, D.J.; Callaway, T.R.; Edrington, T.S.; Anderson, R.C.; Krueger, N. Effects of the dicarboxylic acids malate and fumarate on E. coli O157:H7 and Salmonella enterica typhimurium populations in pure culture and in mixed ruminal microorganism fermentations. Curr. Microbiol. 2009, 58, 488–492. [Google Scholar] [CrossRef]
- El-Kafrawy, S.A.; Abbas, A.T.; Oelkrug, C.; Tahoon, M.; Ezzat, S.; Zumla, A.; Azhar, E.I. IgY antibodies: The promising potential to overcome antibiotic resistance. Front. Immunol. 2023, 14, 1065353. [Google Scholar] [CrossRef]
- Batiha, G.E.; Alkazmi, L.M.; Wasef, L.G.; Beshbishy, A.M.; Nadwa, E.H.; Rashwan, E.K. Syzygium aromaticum L. (Myrtaceae): Traditional Uses, Bioactive Chemical Constituents, Pharmacological and Toxicological Activities. Biomolecules 2020, 10, 202. [Google Scholar] [CrossRef]
- Hume, M.E. Food Safety Symposium: Potential Impact of Reduced Antibiotic Use and the Roles of Prebiotics, Probiotics, and Other Alternatives in Antibiotic-Free Broiler Production. Poult. Sci. 2011, 90, 2663–2669. [Google Scholar] [CrossRef]
- Arsène, M.M.J.; Davares, A.K.L.; Andreevna, S.L.; Vladimirovich, E.A.; Carime, B.Z.; Marouf, R.; Khelifi, I. The use of probiotics in animal feeding for safe production and as potential alternatives to antibiotics. Vet. World 2021, 14, 319–328. [Google Scholar] [CrossRef]
- Adams, C.A. Nutrition-based health in animal production. Nutr. Res. Rev. 2006, 19, 79–89. [Google Scholar] [CrossRef]
- Bitsie, B.; Osorio, A.M.; Henry, D.D.; Silva, B.C.; Godoi, L.A.; Supapong, C.; Brand, T.; Schoonmaker, J.P. Enteric methane emissions, growth, and carcass characteristics of feedlot steers fed a garlic- and citrus-based feed additive in diets with three different forage concentrations. J. Anim. Sci. 2022, 100, skac139. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.; Harper, M.; Giallongo, F.; Bravo, D.M.; Wall, E.H.; Hristov, A.N. Effects of rumen-protected Capsicum oleoresin on immune responses in dairy cows intravenously challenged with lipopolysaccharide. J. Dairy. Sci. 2017, 100, 1902–1913. [Google Scholar] [CrossRef] [PubMed]
- Patra, A.K.; Amasheh, S.; Aschenbach, J.R. Modulation of gastrointestinal barrier and nutrient transport function in farm animals by natural plant bioactive compounds—A comprehensive review. Crit. Rev. Food Sci. Nutr. 2019, 59, 3237–3266. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Kessler, M.G.C.; Marchán-Rivadeneira, M.R.; Han, Y. Combating Antimicrobial Resistance in the Post-Genomic Era: Rapid Antibiotic Discovery. Molecules 2023, 28, 4183. [Google Scholar] [CrossRef]
- Mustafa, A.M.; Abouelenein, D.; Acquaticci, L.; Alessandroni, L.; Angeloni, S.; Borsetta, G.; Caprioli, G.; Nzekoue, F.K.; Sagratini, G.; Vittori, S. Polyphenols, Saponins and Phytosterols in Lentils and Their Health Benefits: An Overview. Pharmaceuticals 2022, 15, 1225. [Google Scholar] [CrossRef] [PubMed]
- Serafini, M.; Peluso, I.; Raguzzini, A. Flavonoids as anti-inflammatory agents. Proc. Nutr. Soc. 2010, 69, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Lu, P.; Li, Z.; Yuan, C.; Liu, H.; Zhao, J.; Lu, W.; Wang, J. Oligomeric Proanthocyanidins and Bamboo Leaf Flavonoids Improve the Quality of Bull Semen Cryopreservation. Molecules 2022, 27, 1144. [Google Scholar] [CrossRef]
- Langhans, W. Food Components in Health Promotion and Disease Prevention. J. Agric. Food Chem. 2018, 66, 2287–2294. [Google Scholar] [CrossRef]
- Liu, K.; Pi, F.; Zhang, H.; Ji, J.; Xia, S.; Cui, F.; Sun, J.; Sun, X. Metabolomics Analysis To Evaluate the Anti-Inflammatory Effects of Polyphenols: Glabridin Reversed Metabolism Change Caused by LPS in RAW 264.7 Cells. J. Agric. Food Chem. 2017, 65, 6070–6079. [Google Scholar] [CrossRef]
- Kong, L.; Yang, C.; Dong, L.; Diao, Q.; Si, B.; Ma, J.; Tu, Y. Rumen Fermentation Characteristics in Pre- and Post-Weaning Calves upon Feeding with Mulberry Leaf Flavonoids and Candida tropicalis Individually or in Combination as a Supplement. Animals 2019, 9, 990. [Google Scholar] [CrossRef]
- Lu, D. A major breakthrough in the strategic direction of the development of animal nutrition: The construction of animal health nutrition theory and technology system and its practical application. Chin. J. Anim. Nutr. 2021, 33, 1–12. [Google Scholar]
- Tedesco, D.; Tava, A.; Galletti, S.; Tameni, M.; Varisco, G.; Costa, A.; Steidler, S. Effects of silymarin, a natural hepatoprotector, in periparturient dairy cows. J. Dairy Sci. 2004, 87, 2239–2247. [Google Scholar] [CrossRef] [PubMed]
- Balcells, J.; Aris, A.; Serrano, A.; Seradj, A.R.; Crespo, J.; Devant, M. Effects of an extract of plant flavonoids (Bioflavex) on rumen fermentation and performance in heifers fed high-concentrate diets. J. Anim. Sci. 2012, 90, 4975–4984. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Dahuja, A.; Tiwari, S.; Punia, S.; Tak, Y.; Amarowicz, R.; Bhoite, A.G.; Singh, S.; Joshi, S.; Panesar, P.S.; et al. Recent trends in extraction of plant bioactives using green technologies: A review. Food Chem. 2021, 353, 129431. [Google Scholar] [CrossRef] [PubMed]
- Shu, G.; Kong, F.; Xu, D.; Yin, L.; He, C.; Lin, J.; Fu, H.; Wang, K.; Tian, Y.; Zhao, X. Bamboo leaf flavone changed the community of cecum microbiota and improved the immune function in broilers. Sci. Rep. 2020, 10, 12324. [Google Scholar] [CrossRef]
- Burkina, V.; Zlabek, V.; Halsne, R.; Ropstad, E.; Zamaratskaia, G. In vitro effects of the citrus flavonoids diosmin, naringenin and naringin on the hepatic drug-metabolizing CYP3A enzyme in human, pig, mouse and fish. Biochem. Pharmacol. 2016, 110–111, 109–116. [Google Scholar] [CrossRef]
- Chen, Y.; Guo, Y.; Song, Z.; Chang, H.; Kuang, Q.; Zheng, Z.; Wang, H.; Zhang, G. Luteolin restricts ASFV replication by regulating the NF-κB/STAT3/ATF6 signaling pathway. Vet. Microbiol. 2022, 273, 109527. [Google Scholar] [CrossRef]
- Formato, M.; Cimmino, G.; Brahmi-Chendouh, N.; Piccolella, S.; Pacifico, S. Polyphenols for Livestock Feed: Sustainable Perspectives for Animal Husbandry? Molecules 2022, 27, 7752. [Google Scholar] [CrossRef]
- Zhan, J.; Liu, M.; Wu, C.; Su, X.; Zhan, K.; Zhao, G.Q. Effects of alfalfa flavonoids extract on the microbial flora of dairy cow rumen. Asian-Australas. J. Anim. Sci. 2017, 30, 1261–1269. [Google Scholar] [CrossRef]
- Van Hung, P. Phenolic Compounds of Cereals and Their Antioxidant Capacity. Crit. Rev. Food Sci. Nutr. 2016, 56, 25–35. [Google Scholar] [CrossRef]
- Ramachandran, S.; Meenatchisundaram, S.; Allu, R.; Thatipelli, S.; Mandal, A.K. Proximate analysis, HPTLC finger print analysis and multi spectrometric analysis of Strychnos nux-vomica nuts. J. Complement. Integr. Med. 2022, 19, 233–242. [Google Scholar] [CrossRef]
- Xie, P.; Chen, S.; Liang, Y.Z.; Wang, X.; Tian, R.; Upton, R. Chromatographic fingerprint analysis--a rational approach for quality assessment of traditional Chinese herbal medicine. J. Chromatogr. A 2006, 1112, 171–180. [Google Scholar] [CrossRef]
- Zhong, X.K.; Li, D.C.; Jiang, J.G. Identification and quality control of Chinese medicine based on the fingerprint techniques. Curr. Med. Chem. 2009, 16, 3064–3075. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.Z.; Xie, P.S.; Chan, K. Perspective of chemical fingerprinting of Chinese herbs. Planta Med. 2010, 76, 1997–2003. [Google Scholar] [CrossRef] [PubMed]
- González, L.A.; Kyriazakis, I.; Tedeschi, L.O. Review: Precision nutrition of ruminants: Approaches, challenges and potential gains. Animal 2018, 12, s246–s261. [Google Scholar] [CrossRef] [PubMed]
- Li, C.Q.; Yao, C.; Zhu, R.Y.; Huang, Y.X.; Kang, W.Y.; Wang, J.M. Spectrum-effect relationship in antioxidant activity of Ligustri Lucidi Fructus based on DPPH, ABTS and FRAP assays. Zhongguo Zhong Yao Za Zhi 2016, 41, 1670–1677. [Google Scholar] [CrossRef] [PubMed]
- Luteolin|C15H10O6|CID 5280445-PubChem. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/5280445 (accessed on 9 May 2023).
- Quercetin|C15H10O7|CID 5280343-PubChem. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/5280343 (accessed on 9 May 2023).
- Taxifolin|C15H12O7|CID 439533-PubChem. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/439533 (accessed on 9 May 2023).
- Hesperetin|C16H14O6|CID 72281-PubChem. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/72281 (accessed on 9 May 2023).
- Myricetin|C15H10O8|CID 5281672-PubChem. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/5281672 (accessed on 9 May 2023).
- Ferulic Acid|C10H10O4|CID 445858-PubChem. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/445858 (accessed on 9 May 2023).
- Isoferulic Acid|C10H10O4|CID 736186-PubChem. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/736186 (accessed on 9 May 2023).
- Methyl Sinapic Acid|C12H14O5|CID 15711802-PubChem. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/15711802 (accessed on 9 May 2023).
- Methyl 4-Hydroxycinnamate|C10H10O3|CID 5319562-PubChem. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/5319562 (accessed on 9 May 2023).
- p-Hydroxybenzalacetone|C10H10O2|CID 796857-PubChem. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/796857 (accessed on 9 May 2023).
- Guan, X.-Q.; Mao, J.-L.; Tang, Y.-X.; Wang, J.-H.; Sun, R. Research progress on pharmacological effects of p-coumaric acid. Chin. Tradit. Herb. Drugs 2018, 49, 4162–4170. [Google Scholar]
- O’Callaghan, T.F.; Hennessy, D.; McAuliffe, S.; Kilcawley, K.N.; O’Donovan, M.; Dillon, P.; Ross, R.P.; Stanton, C. Effect of pasture versus indoor feeding systems on raw milk composition and quality over an entire lactation. J. Dairy Sci. 2016, 99, 9424–9440. [Google Scholar] [CrossRef]
- Oskoueian, E.; Abdullah, N.; Oskoueian, A. Effects of flavonoids on rumen fermentation activity, methane production, and microbial population. Biomed. Res. Int. 2013, 2013, 349129. [Google Scholar] [CrossRef]
- Yan, H.; Zou, C. Progress and prospect of application of traditional Chinese medicine fingerprint (specific chromatogram) in Chinese Pharmacopoeia (2010–2020). Nan Fang Yi Ke Da Xue Xue Bao 2022, 42, 150–155. [Google Scholar] [CrossRef]
- Yang, H.; Yang, T.; Gong, D.; Li, X.; Sun, G.; Guo, P. A trinity fingerprint evaluation system of traditional Chinese medicine. J. Chromatogr. A 2022, 1673, 463118. [Google Scholar] [CrossRef]
- Ma, J.; Li, K.; Shi, S.; Li, J.; Tang, S.; Liu, L. The Application of UHPLC-HRMS for Quality Control of Traditional Chinese Medicine. Front. Pharmacol. 2022, 13, 922488. [Google Scholar] [CrossRef]
- Yuan, X.; Han, B.; Feng, Z.M.; Jiang, J.S.; Yang, Y.N.; Zhang, P.C. Chemical constituents of Ligusticum chuanxiong and their anti-inflammation and hepatoprotective activities. Bioorg Chem. 2020, 101, 104016. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Yang, J.; Hu, X.; Gao, H.; Wang, P.; Wang, X.; Liu, Y.; Cheng, X.; Wei, F.; Ma, S. A stepwise strategy integrating metabolomics and pseudotargeted spectrum-effect relationship to elucidate the potential hepatotoxic components in Polygonum multiflorum. Front. Pharmacol. 2022, 13, 935336. [Google Scholar] [CrossRef] [PubMed]
- Cushnie, T.P.; Lamb, A.J. Recent advances in understanding the antibacterial properties of flavonoids. Int. J. Antimicrob. Agents 2011, 38, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Cushnie, T.P.; Lamb, A.J. Antimicrobial activity of flavonoids. Int. J. Antimicrob. Agents 2005, 26, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Pandi, A.; Kalappan, V.M. Pharmacological and therapeutic applications of Sinapic acid-an updated review. Mol. Biol. Rep. 2021, 48, 3733–3745. [Google Scholar] [CrossRef] [PubMed]
- Peyrot, C.; Mention, M.M.; Brunissen, F.; Allais, F. Sinapic Acid Esters: Octinoxate Substitutes Combining Suitable UV Protection and Antioxidant Activity. Antioxidants 2020, 9, 782. [Google Scholar] [CrossRef]
- Firkins, J.L.; Yu, Z.; Morrison, M. Ruminal nitrogen metabolism: Perspectives for integration of microbiology and nutrition for dairy. J. Dairy Sci. 2007, 90 (Suppl. 1), E1–E16. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wei, W.; Yang, S.; Huang, Z.; Li, C.; Yu, X.; Qi, R.; Liu, W.; Loor, J.J.; Wang, M.; et al. Regulation of Dietary Protein Solubility Improves Ruminal Nitrogen Metabolism In Vitro: Role of Bacteria-Protozoa Interactions. Nutrients 2022, 14, 2972. [Google Scholar] [CrossRef]
- Lee, D.H.; Park, J.K.; Choi, J.; Jang, H.; Seol, J.W. Anti-inflammatory effects of natural flavonoid diosmetin in IL-4 and LPS-induced macrophage activation and atopic dermatitis model. Int. Immunopharmacol. 2020, 89, 107046. [Google Scholar] [CrossRef]
- Formato, M.; Vastolo, A.; Piccolella, S.; Calabrò, S.; Cutrignelli, M.I.; Zidorn, C.; Pacifico, S. Castanea sativa Mill. Leaf: UHPLC-HR MS/MS Analysis and Effects on In Vitro Rumen Fermentation and Methanogenesis. Molecules 2022, 27, 8662. [Google Scholar] [CrossRef] [PubMed]
- Formato, M.; Piccolella, S.; Zidorn, C.; Vastolo, A.; Calabrò, S.; Cutrignelli, M.I.; Pacifico, S. UHPLC-ESI-QqTOF Analysis and In Vitro Rumen Fermentation for Exploiting Fagus sylvatica Leaf in Ruminant Diet. Molecules 2022, 27, 2217. [Google Scholar] [CrossRef]
- Salehi, B.; Venditti, A.; Sharifi-Rad, M.; Kręgiel, D.; Sharifi-Rad, J.; Durazzo, A.; Lucarini, M.; Santini, A.; Souto, E.B.; Novellino, E.; et al. The Therapeutic Potential of Apigenin. Int. J. Mol. Sci. 2019, 20, 1305. [Google Scholar] [CrossRef]
- Su, T.; Huang, C.; Yang, C.; Jiang, T.; Su, J.; Chen, M.; Fatima, S.; Gong, R.; Hu, X.; Bian, Z.; et al. Apigenin inhibits STAT3/CD36 signaling axis and reduces visceral obesity. Pharmacol. Res. 2020, 152, 104586. [Google Scholar] [CrossRef] [PubMed]
- Breinholt, V.M.; Offord, E.A.; Brouwer, C.; Nielsen, S.E.; Brøsen, K.; Friedberg, T. In vitro investigation of cytochrome P450-mediated metabolism of dietary flavonoids. Food Chem. Toxicol. 2002, 40, 609–616. [Google Scholar] [CrossRef] [PubMed]
- Berger, L.M.; Wein, S.; Blank, R.; Metges, C.C.; Wolffram, S. Bioavailability of the flavonol quercetin in cows after intraruminal application of quercetin aglycone and rutin. J. Dairy Sci. 2012, 95, 5047–5055. [Google Scholar] [CrossRef]
- Albert-Vega, C.; Tawfik, D.M.; Trouillet-Assant, S.; Vachot, L.; Mallet, F.; Textoris, J. Immune Functional Assays, From Custom to Standardized Tests for Precision Medicine. Front. Immunol. 2018, 9, 2367. [Google Scholar] [CrossRef]
- Ding, S.; Jiang, H.; Fang, J. Regulation of Immune Function by Polyphenols. J. Immunol. Res. 2018, 2018, 1264074. [Google Scholar] [CrossRef]
- Duraiswamy, A.; Sneha, A.N.; Jebakani, K.S.; Selvaraj, S.; Pramitha, J.L.; Selvaraj, R.; Petchiammal, K.I.; Kather Sheriff, S.; Thinakaran, J.; Rathinamoorthy, S.; et al. Genetic manipulation of anti-nutritional factors in major crops for a sustainable diet in future. Front. Plant. Sci. 2022, 13, 1070398. [Google Scholar] [CrossRef]
- Williams, A.R.; Krych, L.; Fauzan Ahmad, H.; Nejsum, P.; Skovgaard, K.; Nielsen, D.S.; Thamsborg, S.M. A polyphenol-enriched diet and Ascaris suum infection modulate mucosal immune responses and gut microbiota composition in pigs. PLoS ONE 2017, 12, e0186546. [Google Scholar] [CrossRef]
- Xu, G.; Ren, G.; Xu, X.; Yuan, H.; Wang, Z.; Kang, L.; Yu, W.; Tian, K. Combination of curcumin and green tea catechins prevents dimethylhydrazine-induced colon carcinogenesis. Food Chem. Toxicol. 2010, 48, 390–395. [Google Scholar] [CrossRef]
- Pérez-Berezo, T.; Franch, A.; Castellote, C.; Castell, M.; Pérez-Cano, F.J. Mechanisms involved in down-regulation of intestinal IgA in rats by high cocoa intake. J. Nutr. Biochem. 2012, 23, 838–844. [Google Scholar] [CrossRef] [PubMed]
- Camps-Bossacoma, M.; Pérez-Cano, F.J.; Franch, À.; Castell, M. Theobromine Is Responsible for the Effects of Cocoa on the Antibody Immune Status of Rats. J. Nutr. 2018, 148, 464–471. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.W.; Wang, Q.L.; Luo, M.; Zhu, M.D.; Liang, H.M.; Li, W.J.; Cai, H.; Zhou, Z.B.; Wang, H.; Tong, S.Q.; et al. Phytochemistry and pharmacology of natural prenylated flavonoids. Arch. Pharm. Res. 2023, 46, 207–272. [Google Scholar] [CrossRef]
- Menke, K.H.; Steingass, H.; Fritz, D.; Schneider, W.; Raab, L.; Salewski, A. The estimation of the digestibility and metabolizable energy contentof ruminant feedingstuffs from the gas production when they are incubated with rumen liquor in vitro. J. Agric. Sci. 1979, 93, 217–222. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | P1 | P2 | P3 | P4 | P5 | P6 | P7 | P8 | P9 | P10 | P11 | P12 | P13 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Rt min | 13.13 | 15.51 | 41.86 | 42.96 | 43.69 | 44.20 | 46.80 | 68.17 | 73.50 | 75.03 | 80.62 | 83.24 | 87.11 |
| S1 | 0.15 | 0.55 | 0.97 | 1.99 | 0.38 | 0.42 | 0.43 | 0.07 | 0.20 | 0.22 | 0.14 | 0.97 | 0.35 |
| S2 | 0.24 | 0.48 | 1.53 | 3.61 | 0.67 | 1.02 | 0.80 | 0.08 | 0.13 | 0.22 | 0.03 | 0.78 | 0.37 |
| S3 | 0.08 | 0.45 | 1.94 | 3.01 | 0.65 | 0.76 | 0.69 | 0.06 | 0.28 | 0.12 | 0.09 | 1.33 | 0.51 |
| S4 | 0.28 | 0.08 | 2.66 | 3.47 | 1.02 | 0.07 | 0.63 | 0.10 | 0.31 | 0.22 | 0.15 | 1.54 | 0.53 |
| S5 | 0.09 | 0.34 | 0.79 | 2.69 | 0.50 | 0.76 | 0.65 | 0.36 | 0.29 | 0.22 | 0.11 | 1.49 | 0.50 |
| S6 | 0.09 | 0.37 | 1.46 | 3.60 | 0.67 | 0.82 | 0.73 | 0.07 | 0.24 | 0.18 | 0.15 | 1.31 | 0.47 |
| S7 | 0.30 | 1.37 | 1.18 | 3.86 | 0.62 | 0.76 | 0.79 | 0.09 | 0.23 | 0.15 | 0.12 | 1.10 | 0.41 |
| S8 | 0.27 | 0.67 | 1.88 | 3.95 | 0.85 | 0.57 | 0.51 | 0.04 | 0.17 | 0.22 | 0.04 | 1.07 | 0.41 |
| S9 | 0.12 | 0.50 | 1.71 | 2.60 | 0.52 | 0.75 | 0.65 | 0.11 | 0.29 | 0.19 | 0.07 | 1.42 | 0.50 |
| S10 | 0.13 | 0.57 | 2.32 | 3.53 | 0.86 | 0.72 | 0.65 | 0.14 | 0.27 | 0.11 | 0.06 | 1.49 | 0.49 |
| S11 | 0.16 | 0.65 | 2.07 | 3.82 | 0.82 | 0.92 | 0.89 | 0.04 | 0.52 | 0.22 | 0.18 | 2.24 | 0.79 |
| S12 | 0.08 | 0.19 | 1.56 | 1.29 | 0.32 | 0.46 | 0.43 | 0.10 | 0.16 | 0.10 | 0.37 | 0.78 | 0.31 |
| S13 | 0.49 | 1.25 | 1.43 | 5.49 | 0.92 | 2.48 | 1.38 | 0.13 | 0.25 | 0.26 | 0.12 | 1.49 | 0.55 |
| S14 | 0.35 | 1.22 | 0.40 | 4.43 | 0.46 | 1.98 | 1.94 | 0.08 | 0.16 | 0.11 | 0.12 | 1.25 | 0.34 |
| S15 | 0.47 | 0.82 | 1.77 | 3.11 | 0.70 | 0.75 | 0.67 | 0.07 | 0.31 | 0.23 | 0.12 | 1.37 | 0.57 |
| S16 | 0.17 | 0.64 | 3.18 | 4.22 | 1.10 | 1.08 | 0.95 | 0.08 | 0.47 | 0.23 | 0.21 | 2.01 | 0.63 |
| S17 | 0.40 | 0.74 | 2.03 | 4.24 | 1.08 | 1.34 | 1.21 | 0.27 | 0.56 | 0.21 | 0.24 | 3.06 | 1.07 |
| S18 | 0.09 | 0.17 | 1.55 | 1.26 | 0.29 | 0.47 | 0.43 | 0.05 | 0.15 | 0.18 | 0.35 | 0.83 | 0.27 |
| S19 | 0.13 | 1.00 | 3.80 | 6.65 | 1.53 | 1.03 | 0.89 | 0.14 | 0.50 | 0.23 | 0.21 | 3.07 | 1.02 |
| S20 | 0.27 | 0.79 | 3.66 | 4.79 | 1.33 | 0.76 | 0.71 | 0.00 | 0.58 | 0.26 | 0.20 | 2.91 | 0.88 |
| Items | pH | Dry Matter Disappearance Rate/% | NH3-N/(mg/dL) |
|---|---|---|---|
| S1 | 6.66 ± 0.006 | 71.90 ± 2.71 abc | 30.68 ± 9.27 ab |
| S2 | 6.68 ± 0.003 | 67.91 ± 1.73 bc | 24.86 ± 6.79 ab |
| S3 | 6.68 ± 0.007 | 70.42 ± 2.38 bc | 26.10 ± 3.06 ab |
| S4 | 6.68 ± 0.000 | 69.29 ± 0.44 bc | 41.36 ± 19.98 ab |
| S5 | 6.67 ± 0.003 | 71.55 ± 0.84 abc | 51.37 ± 9.31 a |
| S6 | 6.70 ± 0.003 | 69.64 ± 0.68 bc | 29.17 ± 5.07 ab |
| S7 | 6.68 ± 0.003 | 71.68 ± 1.21 abc | 31.85 ± 0.80 ab |
| S8 | 6.69 ± 0.023 | 71.78 ± 2.83 abc | 29.46 ± 13.21 ab |
| S9 | 6.67 ± 0.007 | 71.91 ± 1.95 abc | 20.40 ± 1.15 ab |
| S10 | 6.69 ± 0.09 | 67.53 ± 4.92 bc | 25.97 ± 7.66 ab |
| S11 | 6.67 ± 0.09 | 68.69 ± 2.24 bc | 7.13 ± 0.40 b |
| S12 | 6.68 ± 0.007 | 74.72 ± 1.96 ab | 11.40 ± 2.41 b |
| S13 | 6.66 ± 0.003 | 72.93 ± 0.41 abc | 7.15 ± 0.57 b |
| S14 | 6.67 ± 0.012 | 64.07 ± 0.81 c | 7.87 ± 1.17 b |
| S15 | 6.65 ± 0.010 | 71.78 ± 1.50 abc | 10.32 ± 1.71 b |
| S16 | 6.69 ± 0.020 | 76.92 ± 1.10 ab | 14.39 ± 3.04 b |
| S17 | 6.67 ± 0.009 | 81.34 ± 0.30 a | 7.76 ± 0.88 b |
| S18 | 6.68 ± 0.007 | 77.73 ± 0.58 ab | 8.03 ± 1.39 b |
| S19 | 6.67 ± 0.006 | 81.48 ± 1.12 a | 11.91 ± 3.34 b |
| S20 | 6.67 ± 0.023 | 75.55 ± 2.20 ab | 9.12 ± 2.59 b |
| p | 0.194 | <0.0001 | <0.0001 |
| Items | Acetate, mM | Propionate, mM | Isobutyrate, mM | Butyrate, mM | Isovalerate, mM | Valerate, mM |
|---|---|---|---|---|---|---|
| S1 | 41.21 ± 1.72 ab | 18.52 ± 0.65 ab | 1.33 ± 0.12 | 9.63 ± 0.32 abc | 2.61 ± 0.15 abc | 3.09 ± 0.13 ab |
| S2 | 30.66 ± 0.58 b | 14.45 ± 0.30 b | 0.99 ± 0.4 | 7.06 ± 0.18 c | 1.97 ± 0.09 bc | 2.68 ± 0.05 b |
| S3 | 46.60 ± 1.30 ab | 20.67 ± 0.59 ab | 1.53 ± 0.10 | 10.90 ± 0.40 abc | 2.99 ± 0.13 a | 3.40 ± 0.21 a |
| S4 | 35.08 ± 0.73 b | 16.26 ± 0.34 b | 1.22 ± 0.03 | 8.74 ± 0.20 bc | 2.53 ± 0.05 abc | 2.96 ± 0.03 ab |
| S5 | 46.71 ± 4.60 ab | 21.56 ± 2.19 ab | 1.50 ± 0.15 | 11.29 ± 1.15 ab | 2.94 ± 0.25 a | 3.29 ± 0.21 ab |
| S6 | 46.06 ± 1.52 ab | 21.97 ± 0.75 ab | 1.25 ± 0.07 | 10.53 ± 0.61 abc | 2.44 ± 0.17 abc | 3.11 ± 0.01 ab |
| S7 | 34.34 ± 3.06 b | 15.85 ± 1.44 b | 1.13 ± 0.12 | 7.56 ± 0.64 bc | 1.86 ± 0.12 c | 2.74 ± 0.06 b |
| S8 | 43.68 ± 2.81 ab | 20.26 ± 1.26 ab | 1.36 ± 0.09 | 9.92 ± 056 abc | 2.33 ± 0.11 abc | 3.00 ± 0.10 ab |
| S9 | 42.53 ± 0.71 ab | 20.20 ± 0.35 ab | 1.31 ± 0.05 | 9.55 ± 0.24 abc | 2.33 ± 0.07 abc | 2.84 ± 0.03 ab |
| S10 | 46.85 ± 1.37 ab | 21.57 ± 0.61 ab | 1.42 ± 0.07 | 10.26 ± 0.55 abc | 2.46 ± 0.17 abc | 2.92 ± 0.05 ab |
| S11 | 54.19 ± 2.82 a | 25.07 ± 1.20 a | 1.57 ± 0.10 | 12.77 ± 0.59 a | 2.79 ± 0.12 ab | 3.12 ± 0.10 ab |
| S12 | 46.79 ± 3.79 ab | 21.92 ± 1.72 ab | 1.32 ± 0.12 | 10.56 ± 0.71 abc | 2.35 ± 0.14 abc | 3.02 ± 0.12 ab |
| S13 | 44.55 ± 4.04 ab | 19.92 ± 1.72 ab | 1.40 ± 0.12 | 9.45 ± 0.78 abc | 2.30 ± 0.17 abc | 3.10 ± 0.09 ab |
| S14 | 41.98 ± 7.66 ab | 19.69 ± 3.55 ab | 1.35 ± 0.19 | 9.55 ± 1.52 abc | 2.32 ± 0.26 abc | 2.99 ± 0.17 ab |
| S15 | 39.35 ± 0.65 ab | 18.54 ± 0.36 ab | 1.30 ± 0.06 | 8.55 ± 0.22 bc | 2.18 ± 0.11 abc | 2.98 ± 0.13 ab |
| S16 | 43.47 ± 0.87 ab | 20.52 ± 0.46 ab | 1.09 ± 0.02 | 8.81 ± 0.25 bc | 2.15 ± 0.05 abc | 2.69 ± 0.05 b |
| S17 | 47.54 ± 2.93 ab | 21.45 ± 1.41 ab | 1.31 ± 0.22 | 9.68 ± 0.84 abc | 2.24 ± 0.19 abc | 2.79 ± 0.18 ab |
| S18 | 36.21 ± 2.80 b | 17.06 ± 1.33 b | 1.18 ± 0.10 | 7.65 ± 0.56 bc | 2.02 ± 0.11 bc | 2.86 ± 0.09 ab |
| S19 | 40.13 ± 3.29 ab | 17.70 ± 1.48 ab | 1.35 ± 0.05 | 8.51 ± 0.56 bc | 2.25 ± 0.09 abc | 2.93 ± 0.04 ab |
| S20 | 42.52 ± 5.64 ab | 19.64 ± 2.57 ab | 1.31 ± 0.15 | 9.52 ± 1.39 abc | 2.37 ± 0.30 abc | 2.97 ± 0.19 ab |
| p | <0.0001 | <0.0001 | 0.053 | <0.0001 | <0.0001 | 0.007 |
| NO. | Total Flavonoids Content, mg/g | NO. | Total Flavonoids Content, mg/g |
|---|---|---|---|
| S1 | 13.95 ± 0.19 | S11 | 11.95 ± 0.66 |
| S2 | 10.91 ± 0.62 | S12 | 10.71 ± 0.53 |
| S3 | 10.56 ± 0.50 | S13 | 12.61 ± 1.24 |
| S4 | 12.30 ± 1.20 | S14 | 10.51 ± 0.62 |
| S5 | 12.71 ± 0.60 | S15 | 10.96 ± 0.25 |
| S6 | 11.21 ± 0.43 | S16 | 11.16 ± 0.34 |
| S7 | 9.97 ± 0.41 | S17 | 13.00 ± 0.65 |
| S8 | 11.81 ± 1.08 | S18 | 10.86 ± 0.62 |
| S9 | 13.50 ± 0.57 | S19 | 17.38 ± 0.49 |
| S10 | 10.12 ± 0.53 | S20 | 12.55 ± 0.82 |
| pH | Dry Matter Disappearance Rate/% | NH3-N/(mg/dL) | |
|---|---|---|---|
| Total flavonoids content mg/g | −0.378 | −0.047 | 0.535 * |
| p | 0.100 | 0.845 | 0.015 |
| Acetate, mM | Propionate, mM | Isobutyrate, mM | Butyrate, mM | Isovalerate, mM | Valerate, mM | |
|---|---|---|---|---|---|---|
| Total flavonoids content mg/g | 0.048 | −0.038 | 0.194 | 0.016 | 0.132 | 0.057 |
| p | 0.847 | 0.876 | 0.425 | 0.948 | 0.589 | 0.816 |
| Model | Before Optimization | After Optimization | |||
|---|---|---|---|---|---|
| R- Squared | p- Value | R- Squared | p- Value | Formula | |
| PH | 0.7293 | 0.4159 | 0.4599 | 0.0174 | |
| Dry matter disappearance rate | 0.9235 | 0.0224 | 0.6173 | 0.0003 | |
| NH3-N | 0.9499 | 0.0071 | 0.4309 | 0.0083 | |
| Acetate | 0.7637 | 0.3243 | 0.5549 | 0.0294 | |
| Item | Model Checking | ||||
|---|---|---|---|---|---|
| Predicted Value and True Value | Paired Difference | ||||
| Correlation | p-Value | Average Value | SEM | p-Value | |
| pH | 0.678 | 0.001 | 0.00003 | 0.0875 | 0.988 |
| Dry matter disappearance rate | 0.786 | 0 | −0.00007 | 0.61658 | 1 |
| NH3-N | 0.656 | 0.002 | −0.00058 | 2.16054 | 1 |
| Acetate | 0.745 | 0 | 0.00082 | 0.81847 | 0.999 |
| Classification According to the Cluster | Structure | Putative Structure | Base Feature | |
|---|---|---|---|---|
| P4, P6, P7 |  Tricin |  Diosmetin |  Luteolin [36] | flavonoids with methoxy links in the B ring |
| P4, P6, P7 |  Tricin |  Diosmetin |  Quercetin [37] | flavonoids with methoxy links in the B ring |
| P4, P6, P7 |  Tricin |  Diosmetin |  Taxifolin [38] | flavonoids with methoxy links in the B ring |
| P4, P6, P7 |  Tricin |  Diosmetin |  Hesperetin [39] | flavonoids with methoxy links in the B ring |
| P4, P6, P7 |  Tricin |  Diosmetin |  Myricetin [40] | flavonoids with methoxy links in the B ring |
| P1, P2, P10 |  p-coumaric acid |  sinapic acid |  ferulic acid [41] | 4-hydroxy cassia bark acid |
| P1, P2, P10 |  p-coumaric acid |  sinapic acid |  isoferulic acid [42] | 4-hydroxy cassia bark acid |
| P1, P2, P10 |  p-coumaric acid |  sinapic acid |  methyl sinapic acid [43] | 4-hydroxy cassia bark acid |
| P1, P2, P10 |  p-coumaric acid |  sinapic acid |  Methyl 4-hydroxycinnamate [44] | 4-hydroxy cassia bark acid |
| P1, P2, P10 |  p-coumaric acid |  sinapic acid |  p-Hydroxybenzalacetone [45] | 4-hydroxy cassia bark acid |
| Peak | Moisture% | Dry Matter% | Crude Fat% | Crude Ash% | Crude Protein% | Neutral Detergent Fiber (NDF)% | Acid Detergent Fiber (ADF)% |
|---|---|---|---|---|---|---|---|
| S1 | 44.76 | 54.02 | 2.02 | 5.86 | 10.07 | 14.37 | 6.74 |
| S2 | 54.78 | 44.33 | 1.96 | 4.93 | 8.20 | 15.47 | 7.22 |
| S3 | 50.64 | 48.31 | 2.03 | 5.21 | 9.45 | 16.59 | 6.53 |
| S4 | 46.79 | 51.99 | 2.45 | 3.58 | 9.69 | 17.81 | 7.86 |
| S5 | 46.15 | 52.68 | 2.17 | 5.24 | 9.90 | 14.79 | 7.38 |
| S6 | 49.83 | 49.09 | 1.93 | 4.89 | 8.85 | 15.31 | 6.89 |
| S7 | 56.31 | 42.67 | 1.68 | 3.88 | 7.62 | 12.40 | 4.82 |
| S8 | 52.28 | 46.48 | 1.58 | 4.58 | 8.26 | 13.19 | 5.89 |
| S9 | 50.90 | 47.89 | 1.92 | 3.96 | 9.08 | 13.64 | 5.00 |
| S10 | 41.02 | 57.68 | 2.92 | 5.94 | 11.11 | 17.30 | 7.66 |
| S11 | 46.33 | 52.42 | 2.61 | 5.02 | 9.57 | 19.03 | 6.13 |
| S12 | 49.03 | 49.80 | 2.38 | 4.50 | 9.48 | 14.12 | 6.09 |
| S13 | 51.83 | 46.81 | 0.97 | 3.76 | 8.27 | 20.55 | 12.43 |
| S14 | 64.02 | 34.79 | 1.73 | 2.44 | 5.17 | 19.67 | 9.31 |
| S15 | 46.26 | 52.21 | 2.24 | 4.62 | 9.35 | 17.36 | 10.34 |
| S16 | 43.26 | 55.08 | 1.85 | 4.62 | 10.29 | 20.75 | 11.64 |
| S17 | 53.77 | 44.88 | 0.95 | 3.81 | 7.84 | 17.77 | 8.07 |
| S18 | 48.32 | 50.29 | 2.24 | 5.16 | 9.24 | 18.87 | 8.44 |
| S19 | 51.44 | 46.98 | 1.44 | 4.33 | 9.32 | 15.23 | 11.13 |
| S20 | 51.55 | 46.92 | 1.85 | 3.47 | 8.75 | 20.03 | 9.07 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, X.; Xiong, A.; Yu, S.; Wang, L.; Wang, J.; Zhao, Y.; Jiang, L. Establishment of Flavonoid Fingerprint of TMR Diet and Optimization Factor Analysis Strategy and In Vitro Fermentation Parameters Based on Spectrum–Effect Relationship. Fermentation 2023, 9, 571. https://doi.org/10.3390/fermentation9060571
Zhao X, Xiong A, Yu S, Wang L, Wang J, Zhao Y, Jiang L. Establishment of Flavonoid Fingerprint of TMR Diet and Optimization Factor Analysis Strategy and In Vitro Fermentation Parameters Based on Spectrum–Effect Relationship. Fermentation. 2023; 9(6):571. https://doi.org/10.3390/fermentation9060571
Chicago/Turabian StyleZhao, Xiaobo, Anran Xiong, Shiqiang Yu, Linwei Wang, Jing Wang, Yuchao Zhao, and Linshu Jiang. 2023. "Establishment of Flavonoid Fingerprint of TMR Diet and Optimization Factor Analysis Strategy and In Vitro Fermentation Parameters Based on Spectrum–Effect Relationship" Fermentation 9, no. 6: 571. https://doi.org/10.3390/fermentation9060571
APA StyleZhao, X., Xiong, A., Yu, S., Wang, L., Wang, J., Zhao, Y., & Jiang, L. (2023). Establishment of Flavonoid Fingerprint of TMR Diet and Optimization Factor Analysis Strategy and In Vitro Fermentation Parameters Based on Spectrum–Effect Relationship. Fermentation, 9(6), 571. https://doi.org/10.3390/fermentation9060571