
Bioprotective Effect of a Torulaspora delbrueckii/Lachancea thermotolerans-Mixed Inoculum in Red Winemaking

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms
2.2. Vinifications
- -
- Initial spontaneous fermentation without addition of SO2 and inoculation after 72 h with the commercial yeast S. cerevisiae Uvaferm VRB at a dose of 4 × 106 cells/mL (S).
- -
- Initial spontaneous fermentation with addition of SO2 at a dose of 50 mg/L and inoculation after 72 h with the commercial yeast S. cerevisiae Uvaferm VRB (4 × 106 cells/mL) (SS).
- -
- Inoculation of the NS-mixed starter at vatting at 2 × 106 cells/mL without addition of SO2 and inoculation of S. cerevisiae Uvaferm VRB (4 × 106 cells/mL) at 72 h (TL).
2.3. Microorganism Count
2.4. Microorganism Identification
2.5. Physicochemical Analysis
2.6. Statistical Analysis
3. Results and Discussion
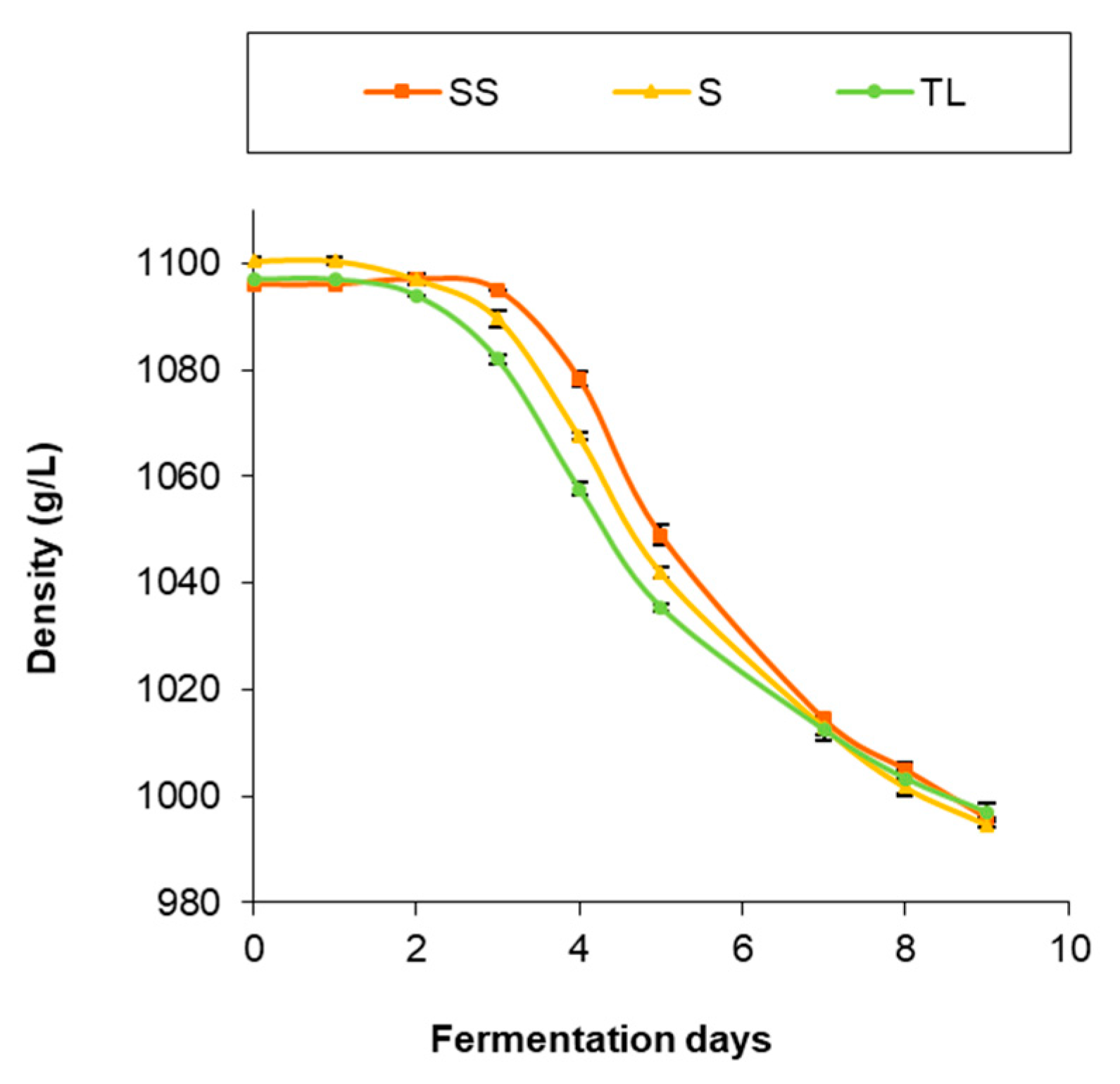
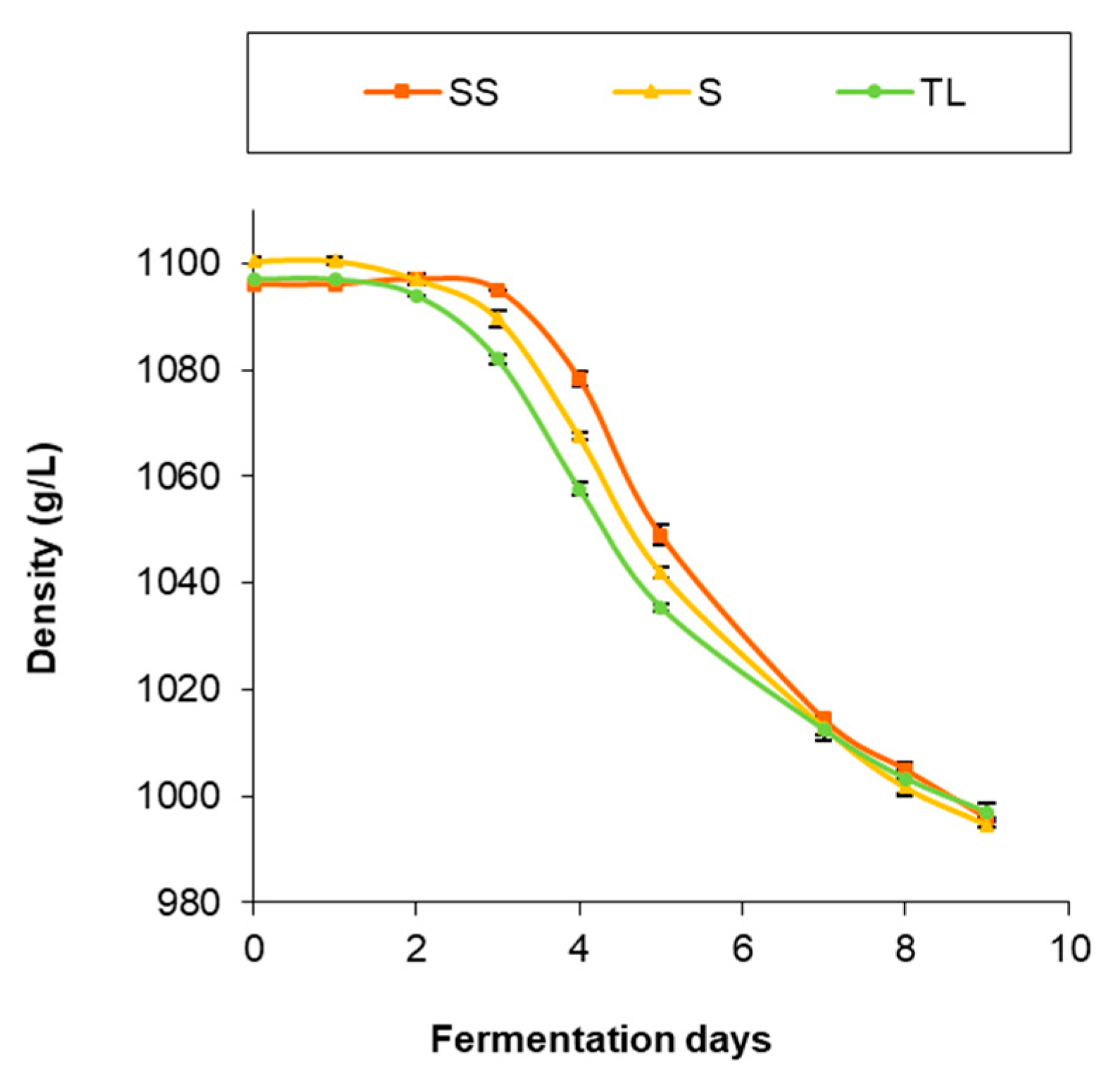
3.1. AF Kinetics
3.2. Yeasts
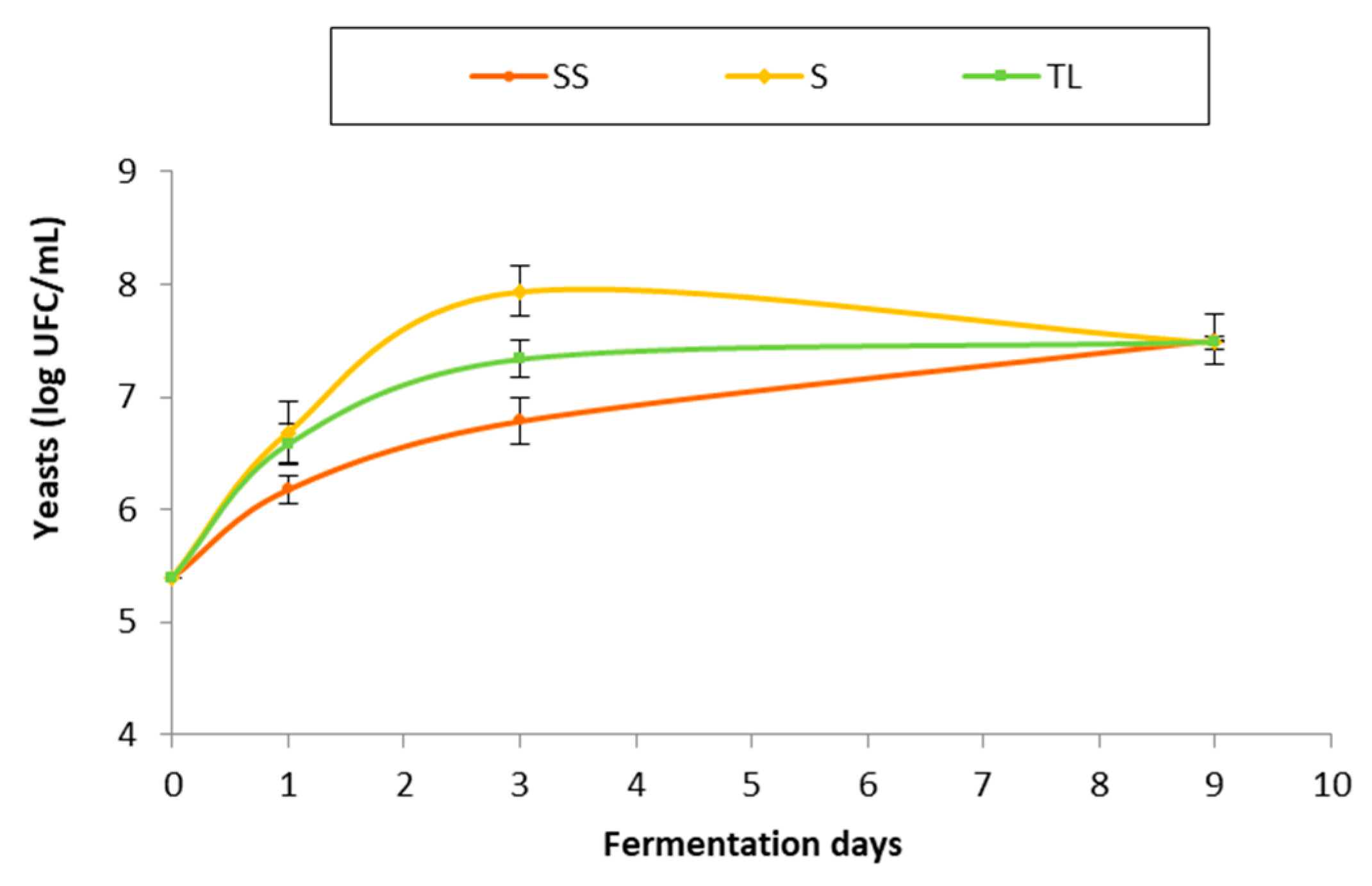
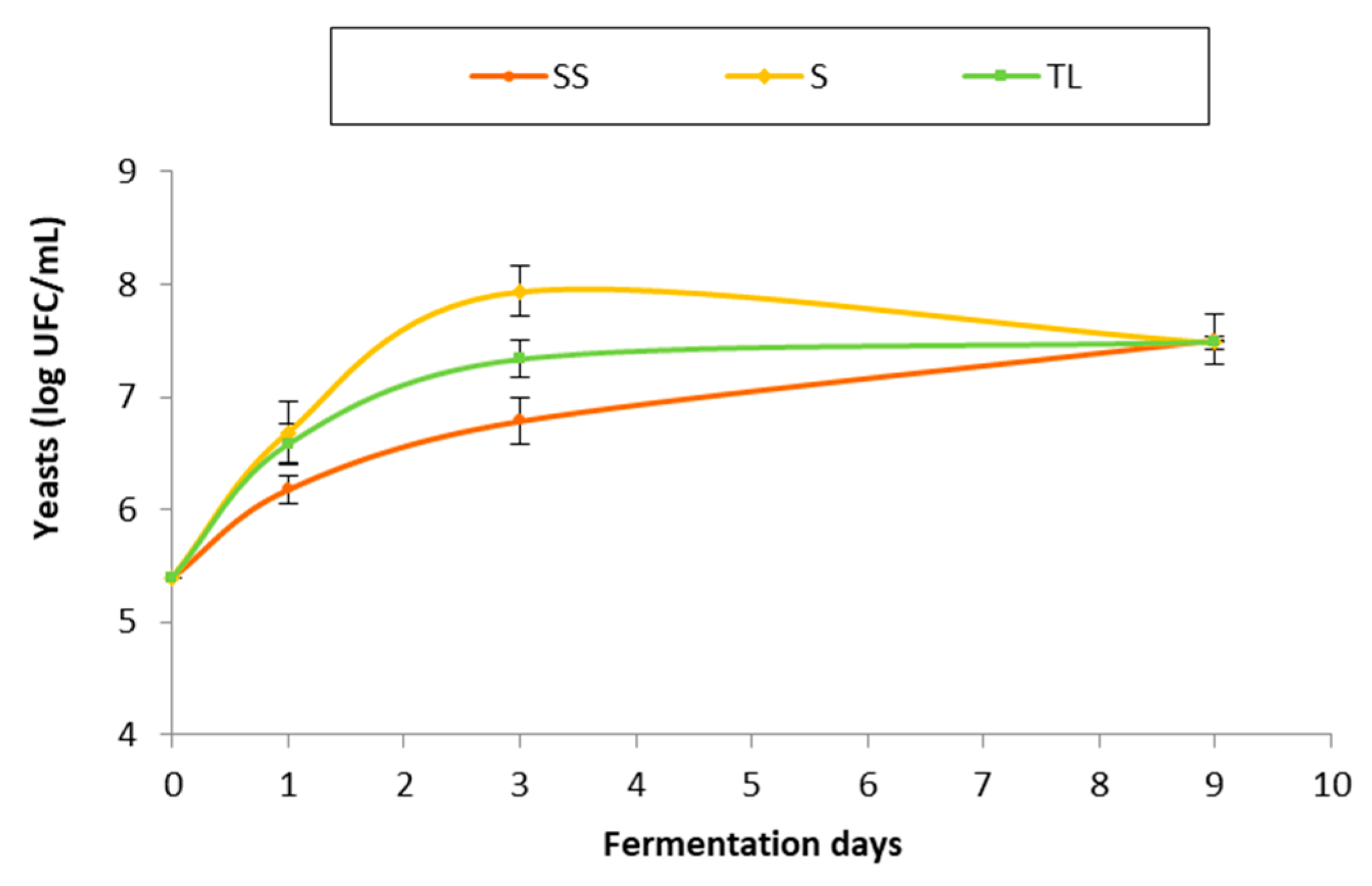
3.2.1. Evolution of the Yeast Population during Fermentation
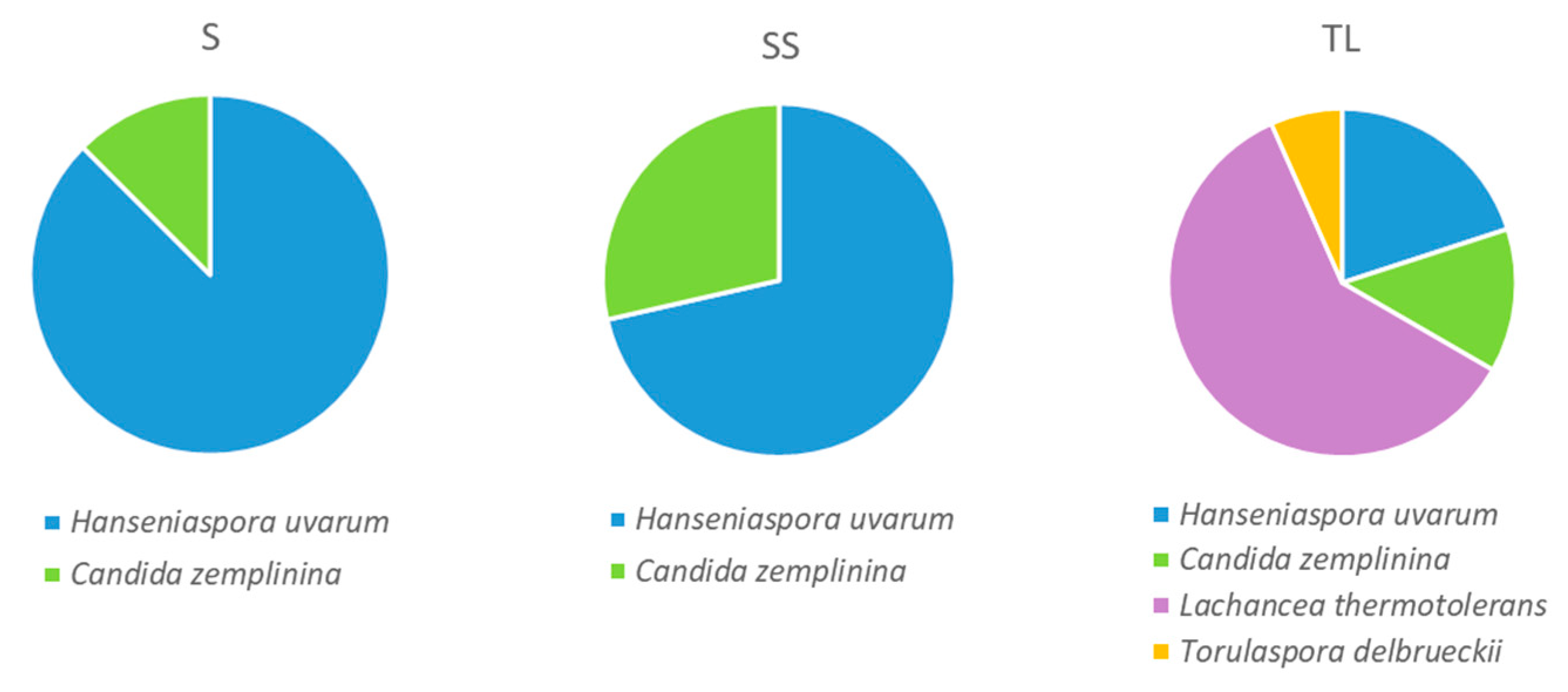
3.2.2. Yeast Species Present in the Tanks 24 h after Vatting
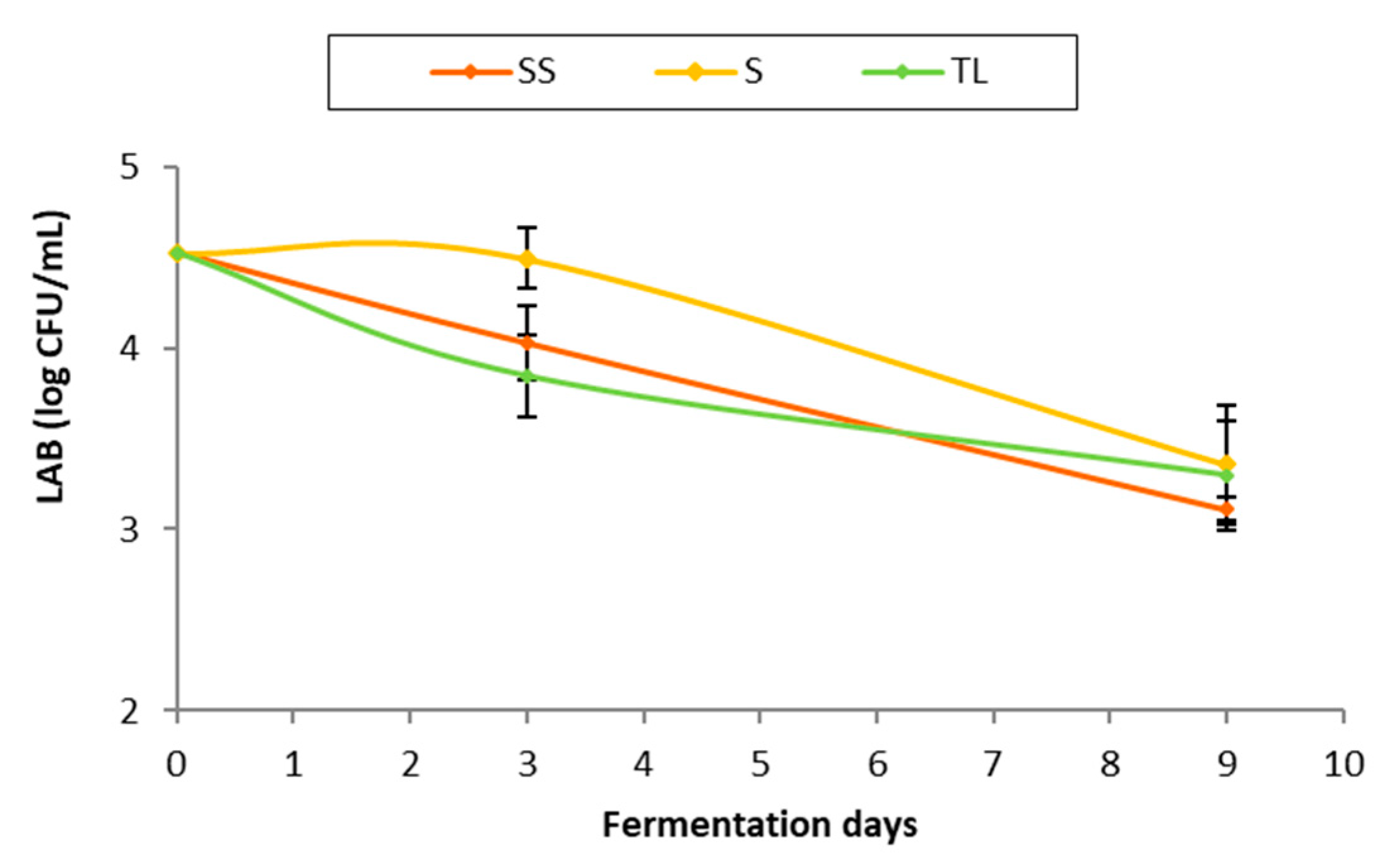
3.3. Lactic Acid Bacteria
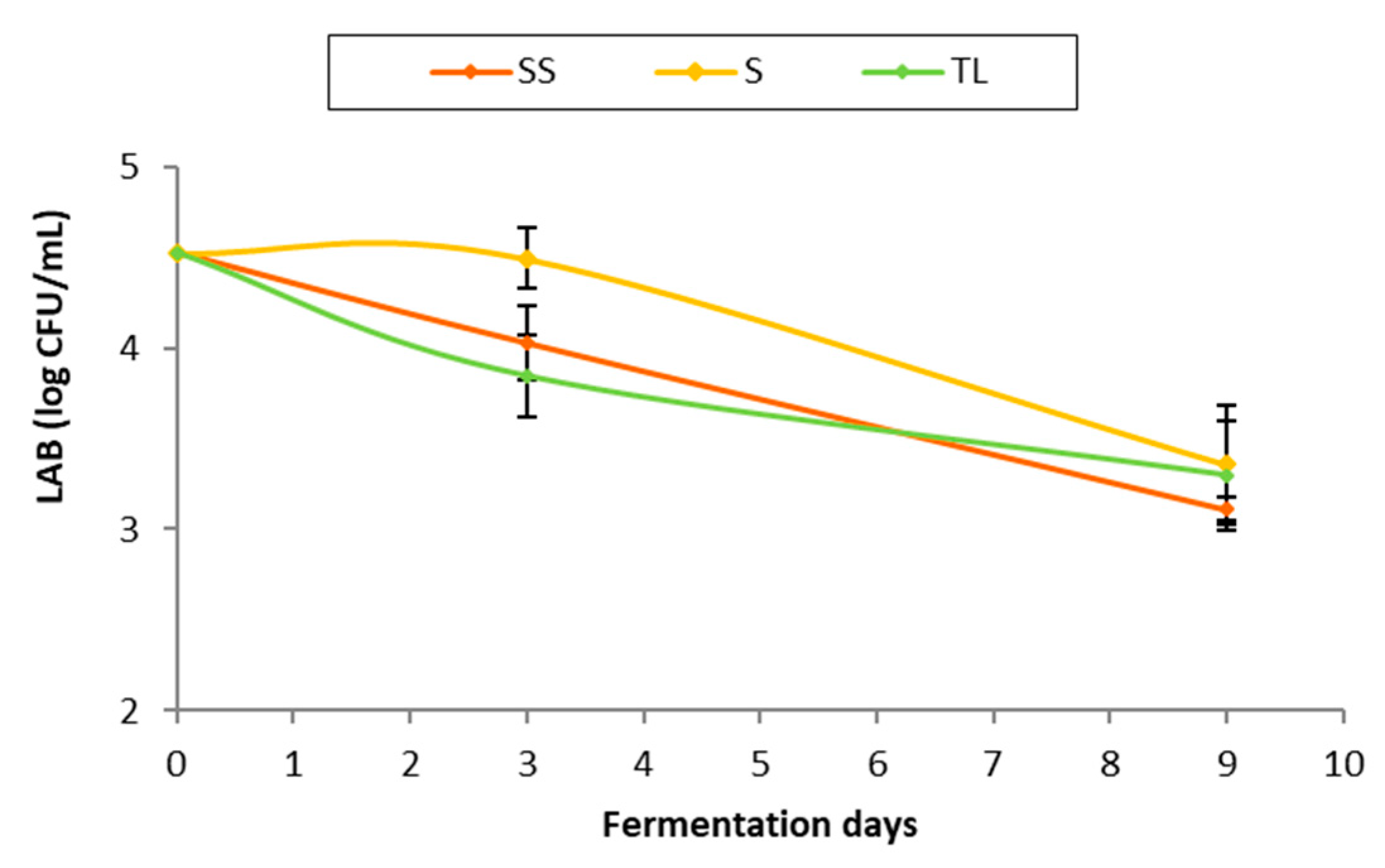
3.3.1. Evolution of the Lactic Acid Bacteria Population during Fermentation
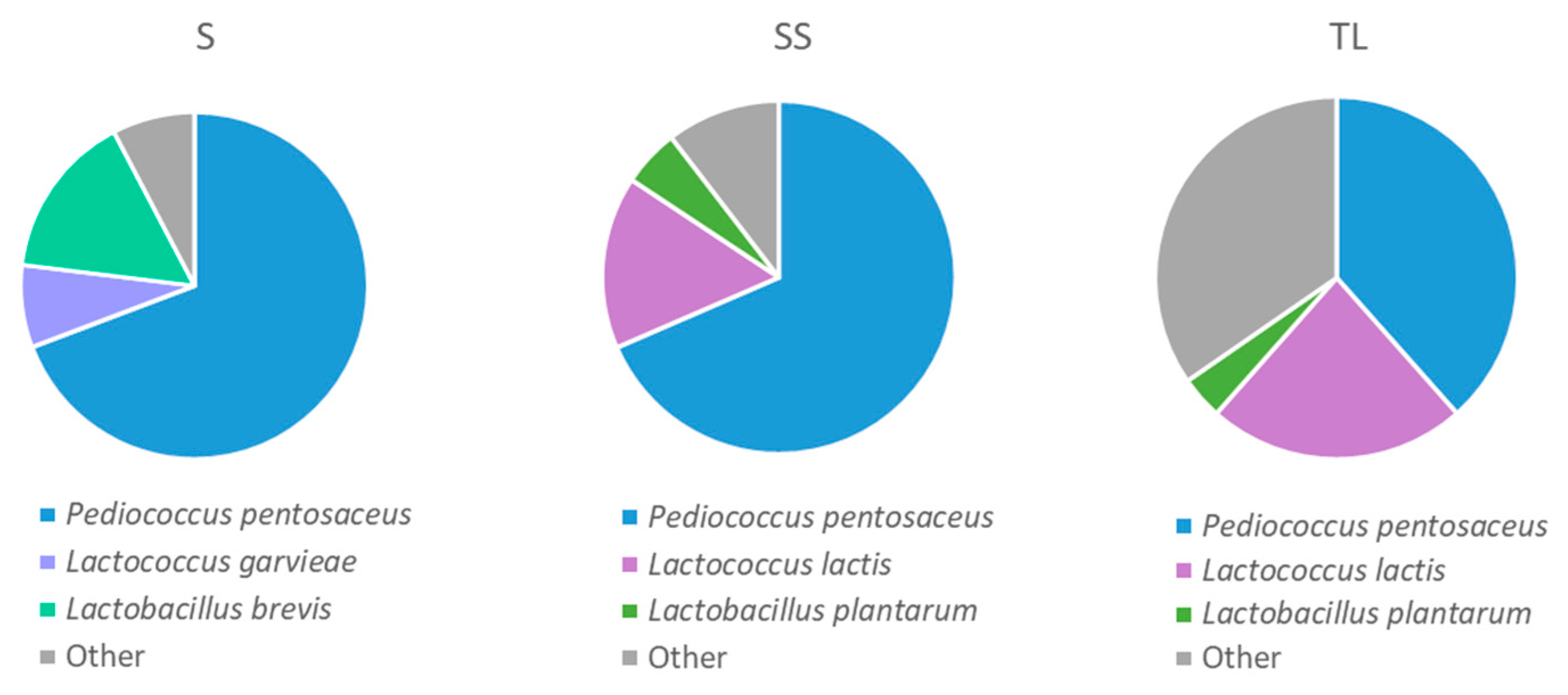
3.3.2. Lactic Acid Bacteria Present in the Tanks 24 h after Vatting
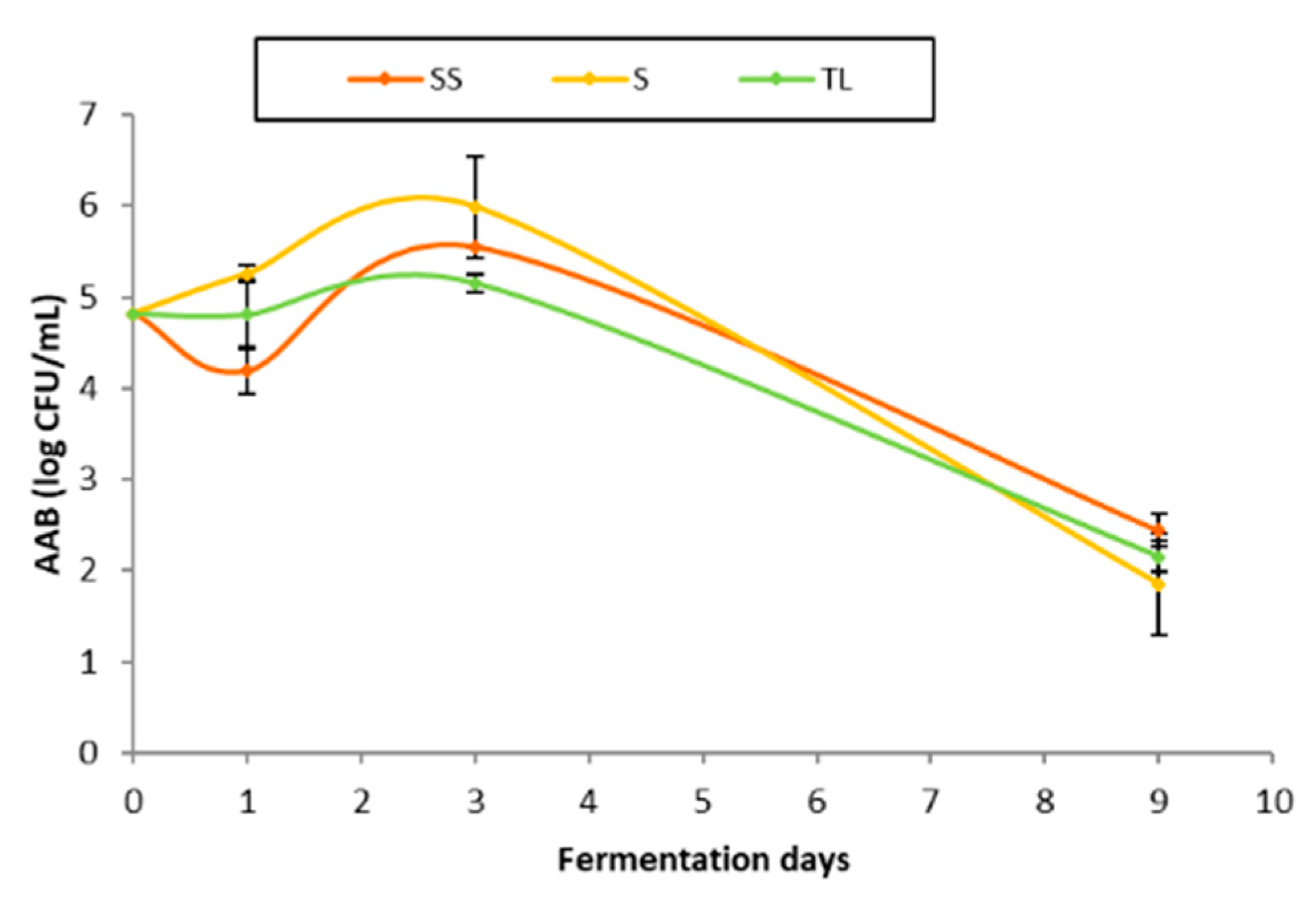
3.4. Acetic Acid Bacteria
3.4.1. Evolution of the Acetic Acid Bacteria Population during Fermentation
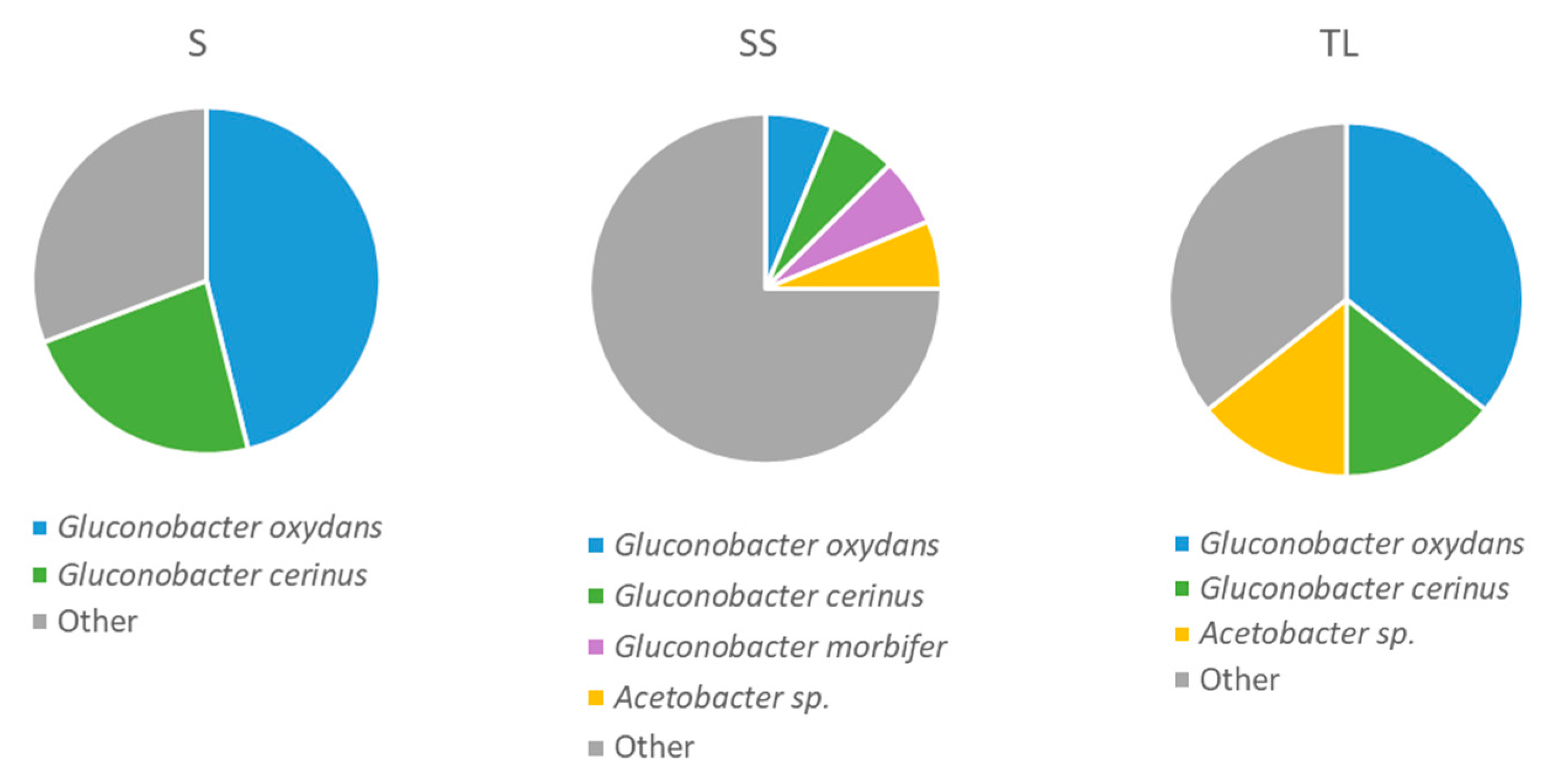
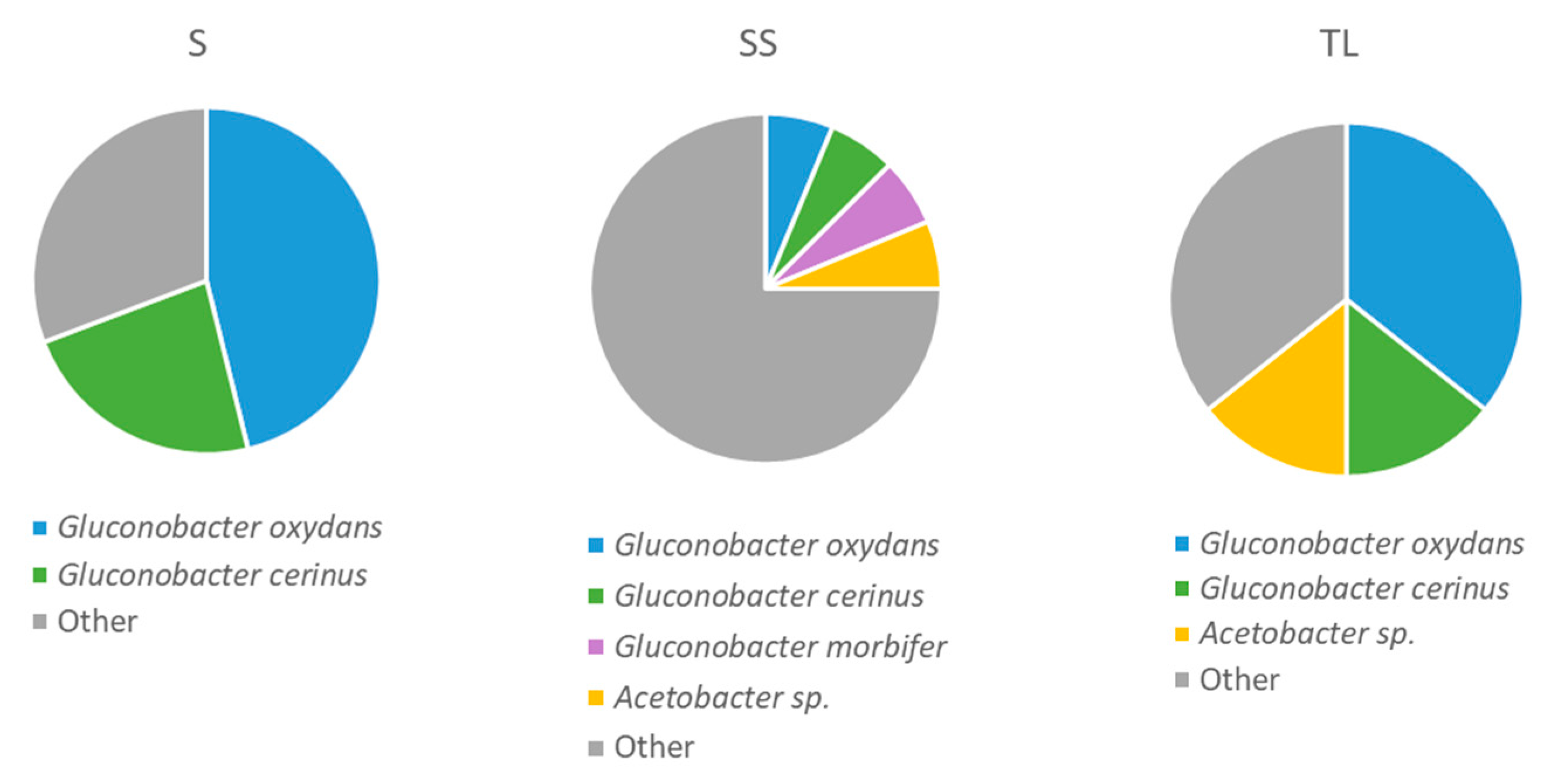
3.4.2. Acetic Acid Bacteria Present in the Tanks 24 h after Vatting
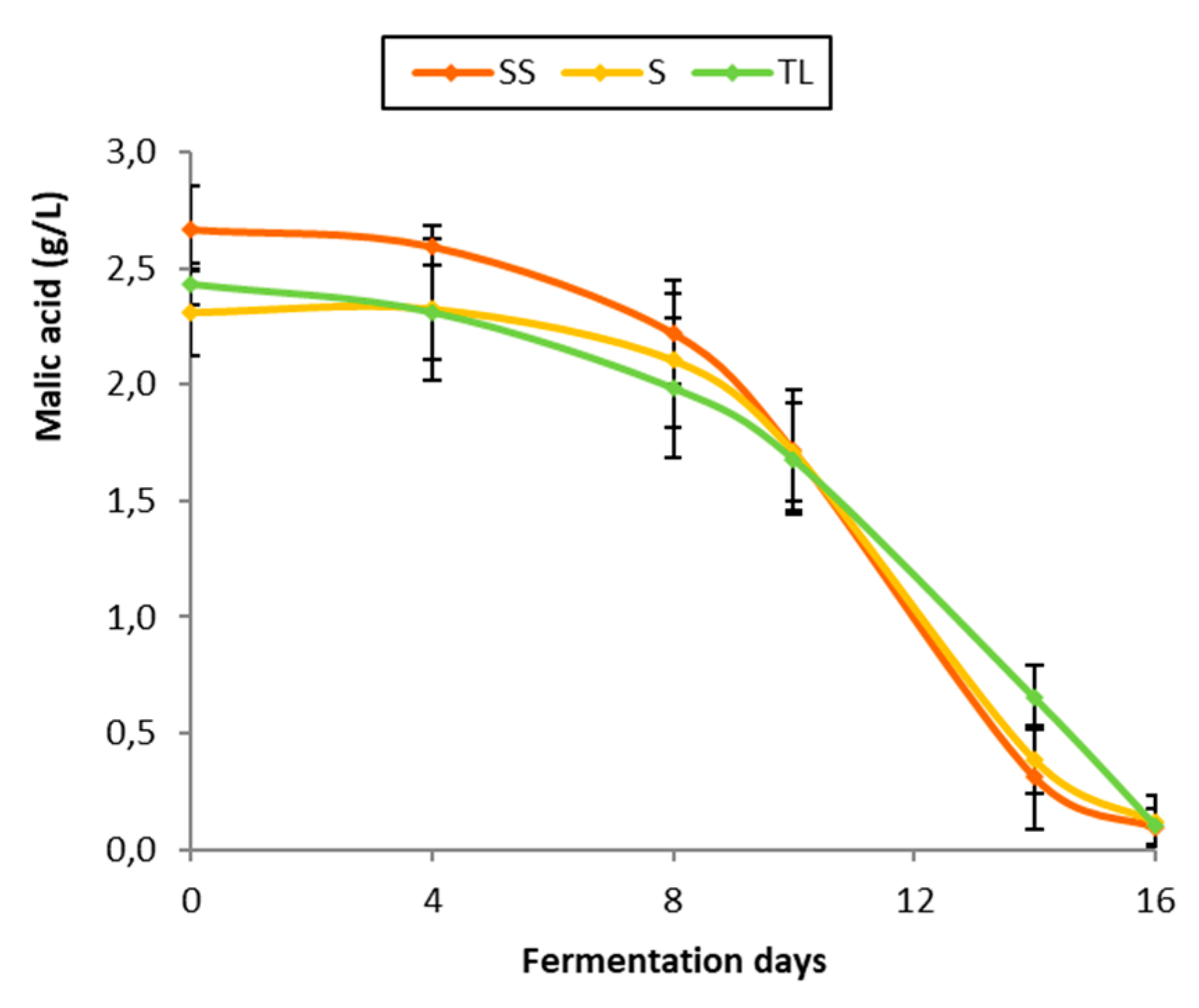
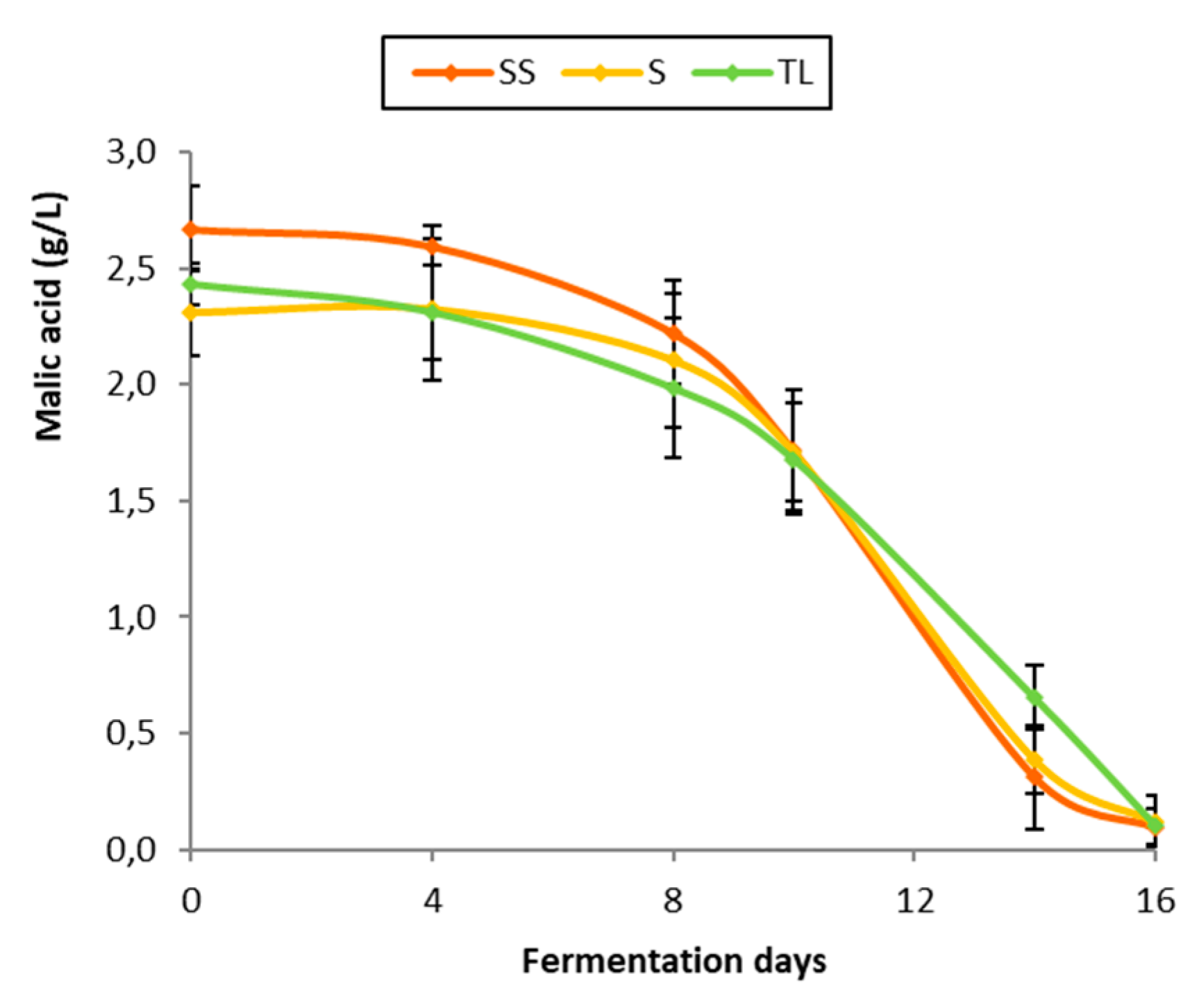
3.5. Malolactic Fermentation
3.6. Characteristics of the Wines
3.6.1. Physical Chemical Parameters
3.6.2. Color Parameters
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Hidalgo, J. Tratado de Enología, 2nd ed.; Mundi Prensa: Madrid, España, 2011. [Google Scholar]
- Carrascón, V.; Vallverdú-queralt, A.; Meudec, E.; Sommerer, N. The kinetics of oxygen and SO2 consumption by red wines. What do they tell about oxidation mechanisms and about changes in wine composition? Food Chem. 2018, 241, 206–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vally, H.; Misso, N.L.A.; Madan, V. Clinical effects of sulphite additives. Clin. Exp. Allergy 2009, 39, 1643–1651. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.; Gama, J.; Pinto, N.; Pivi, G.; Brancal, H.; Carvalho, L.; Loureiro, V.; Vaz Patto, M. Sulfite concentration and the occurrence of headache in young adults: A prospective study. Eur. J. Clin. Nutr. 2019, 73, 1316–1322. [Google Scholar] [CrossRef] [PubMed]
- Bauer, M.A.; Kainz, K.; Carmona-Gutiérrez, D.; Madeo, F. Microbial wars: Competition in ecological niches and within the microbiome. Microbial Cell. 2018, 5, 215–219. [Google Scholar] [CrossRef]
- Ghoul, M.; Mitri, S. The Ecology and Evolution of Microbial Competition. Trends Microbiol. 2016, 24, 833–845. [Google Scholar] [CrossRef]
- Di Gianvito, P.; Englezos, V.; Rantsiou, K.; Cocolin, L. Bioprotection strategies in winemaking. Int. J. Food Microbiol. 2022, 364, 109532. [Google Scholar] [CrossRef]
- Simonin, S.; Alexandre, H.; Nikolantonaki, M.; Coelho, C.; Tourdot-Maréchal, R. Inoculation of Torulaspora delbrueckii as a bio-protection agent in winemaking. Food Res. Int. 2018, 107, 451–461. [Google Scholar] [CrossRef]
- Simonin, S.; Roullier-Gall, C.; Ballester, J.; Schmitt-Kopplin, P.; Quintanilla-Casas, B.; Vichi, S.; Peyron, D.; Alexandre, H.; Tourdot-Maréchal, R. Bio-Protection as an Alternative to Sulphites: Impact on Chemical and Microbial Characteristics of Red Wines. Front. Microbiol. 2020, 11, 1308. [Google Scholar] [CrossRef]
- Escribano-Viana, R.; González-Arenzana, L.; Garijo, P.; López, R.; Santamaría, P.; Gutiérrez, A.R. Selection process of a mixed inoculum of non-Saccharomyces yeasts isolated in the D.O.Ca. Rioja. Fermentation 2021, 7, 148. [Google Scholar] [CrossRef]
- Gutiérrez, A.R. Selección de Levaduras Vínicas en la Denominación de Origen Rioja; Universidad del País Vasco: Bilbao, Spain, 1994. [Google Scholar]
- González-Arenzana, L.; Santamaría, P.; López, R.; López-Alfaro, I. Indigenous lactic acid bacteria communities in alcoholic and malolactic fermentations of Tempranillo wines elaborated in ten wineries of La Rioja (Spain). Food Res. Int. 2013, 50, 438–445. [Google Scholar] [CrossRef]
- Cocolin, L.; Bisson, L.F.; Mills, D.A. Direct profiling of the yeast dynamics in wine fermentations. FEMS Microbiol. Lett. 2000, 189, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Lopez, I.; Ruiz-Larrea, F.; Cocolin, L.; Orr, E.; Phister, T.; Marshall, M.; VanderGheynst, J.; Mills, D.A. Design and Evaluation of PCR Primers for Analysis of Bacterial Populations in Wine by Denaturing Gradient Gel Electrophoresis. Appl. Environ. Microbiol. 2003, 69, 6801–6807. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Commission Regulation (EEC) No. 2676/90 determining Community methods for the analysis of wines. Off. J. Eur. Union 1990, 272, 1–192.
- Lipka, Z.; Tanner, H. Une nouvelle méthode de dosage rapide de l’acide tartrique dans les moûts, les vins et autres boissons (selon Rebelein). Rev. Suisse Vitic Arboric Hortic. 1974, 6, 5–10. [Google Scholar]
- Somers, T.C.; Evans, M.E. Wine quality: Correlations with colour density and anthocyanin equilibria in a group of young red wines. J. Sci. Food Agric. 1974, 25, 1369–1379. [Google Scholar] [CrossRef]
- Glories, Y. Recherches sur la Matière Colorantes des Vins Rouges; Université de Bourdeaux II: Bordeaux, France, 1978. [Google Scholar]
- Ruiz, M. La Crianza del Vino Tinto Desde la Perspectiva Vitícola; AMV Ediciones: Madrid, Spain, 1999. [Google Scholar]
- Grangeteau, C.; Gerhards, D.; Von Wallbrunn, C.; Alexandre, H.; Rousseaux, S.; Guilloux-Benatier, M. Persistence of two non-Saccharomyces yeasts (Hanseniaspora and Starmerella) in the cellar. Front. Microbiol. 2016, 7, 268. [Google Scholar] [CrossRef]
- Plata, C.; Millán, C.; Mauricio, J.C.; Ortega, J.M. Formation of ethyl acetate and isoamyl acetate by various species of wine yeasts. Food Microbiol. 2003, 20, 217–224. [Google Scholar] [CrossRef]
- Romano, P.; Suzzi, G.; Comi, G.; Zironi, R. Higher alcohol and acetic acid production by apiculate wine yeasts. J. Appl. Bacteriol. 1992, 73, 126–130. [Google Scholar] [CrossRef]
- Escribano-Viana, R.; López-Alfaro, I.; López, R.; Santamaría, P.; Gutiérrez, A.R.; González-Arenzana, L. Impact of Chemical and Biological Fungicides Applied to Grapevine on Grape Biofilm, Must, and Wine Microbial Diversity. Front. Microbiol. 2018, 9, 59. [Google Scholar] [CrossRef]
- Muñoz, R.; Moreno-Arribas, M.V.; De Las Rivas, B. Bacterias lácticas. In Microbiología del Vino; Carrascosa, A.V., Muñoz, R., González, R., Eds.; AMV Ediciones: Madrid, Spain, 2005. [Google Scholar]
- Costello, P.J.; Henschke, P.A.; Markides, A.J. Standardised methodology for testing malolactic bacteria and wine yeast compatibility. Aust. J. Grape Wine Res. 2003, 9, 127–137. [Google Scholar] [CrossRef]
- Wade, M.E.; Strickland, M.T.; Osborne, J.P.; Edwards, C.G. Role of Pediococcus in winemaking. Aust. J. Grape Wine Res. 2018, 25, 7–24. [Google Scholar] [CrossRef] [Green Version]
- Edwards, C.G.; Peterson, J.C.; Boylston, T.D.; Vasile, T.D. Interactions Between Leuconostoc oenos and Pediococcus spp. During Vinification of Red Wines. Am. J. Enol. Vitic. 1994, 45, 49 LP–55. [Google Scholar]
- Juega, M.; Costantini, A.; Bonello, F.; Cravero, M.-C.; Martinez-Rodriguez, A.; Carrascosa, A.; Garcia-Moruno, E. Effect of malolactic fermentation by Pediococcus damnosus on the composition and sensory profile of Albariño and Caiño white wines. J. Appl. Microbiol. 2014, 116, 586–595. [Google Scholar] [CrossRef] [PubMed]
- Costello, P.J.; Henschke, P.A. Mousy off-flavor of wine: Precursors and biosynthesis of the causative N-Heterocycles 2-ethyltetrahydropyridine, 2-acetyltetrahydropyridine, and 2-acetyl-1-pyrroline by Lactobacillus hilgardii DSM 20176. J. Agric. Food Chem. 2002, 50, 7079–7087. [Google Scholar] [CrossRef] [PubMed]
- Costello, P.J.; Lee, T.H.; Henschke, P. Ability of lactic acid bacteria to produce N-heterocycles causing mousy off-flavour in wine. Aust. J. Grape Wine Res. 2001, 7, 160–167. [Google Scholar] [CrossRef]
- Capozzi, V.; Russo, P.; Ladero, V.; Fernández, M.; Fiocco, D.; Alvarez, M.A.; Grieco, F.; Spano, G. Biogenic amines degradation by Lactobacillus plantarum: Toward a potential application in wine. Front. Microbiol. 2012, 3, 122. [Google Scholar] [CrossRef] [Green Version]
- Iorizzo, M.; Testa, B.; Lombardi, S.J.; García-Ruiz, A.; Muñoz-González, C.; Bartolomé, B.; Moreno-Arribas, M.V. Selection and technological potential of Lactobacillus plantarum bacteria suitable for wine malolactic fermentation and grape aroma release. LWT Food Sci. Technol. 2016, 73, 557–566. [Google Scholar] [CrossRef] [Green Version]
- Krieger-Weber, S.; Heras, J.M.; Suarez, C. Lactobacillus plantarum, a new biological tool to control malolactic fermentation: A review and an outlook. Beverages 2020, 6, 23. [Google Scholar] [CrossRef] [Green Version]
- Guillamón, J.M.; Mas, A. Bacterias acéticas. In Microbiología del Vino; Carrascosa, A.V., Muñoz, R., González, R., Eds.; AMV Ediciones: Madrid, Spain, 2005. [Google Scholar]
- Maragkoudakis, P.A.; Nardi, T.; Bovo, B.; D’Andrea, M.; Howell, K.S.; Giacomini, A.; Corich, V. Biodiversity, dynamics and ecology of bacterial community during grape marc storage for the production of grappa. Int. J. Food Microbiol. 2013, 162, 143–151. [Google Scholar] [CrossRef]
- Takahashi, M.; Ohta, T.; Masaki, K.; Mizuno, A.; Goto-Yamamoto, N. Evaluation of microbial diversity in sulfite-added and sulfite-free wine by culture-dependent and -independent methods. J. Biosci. Bioeng. 2014, 117, 569–575. [Google Scholar] [CrossRef] [PubMed]
- Shelef, L.A. Antimicrobial Effects of Lactates: A Review. J. Food Prot. 1994, 57, 445–450. [Google Scholar] [CrossRef] [PubMed]
- Morata, A.; Loira, I.; Tesfaye, W.; Bañuelos, M.A.; González, C.; Suárez Lepe, J.A. Lachancea thermotolerans applications in wine technology. Fermentation 2018, 4, 53. [Google Scholar] [CrossRef] [Green Version]
- Escribano-Viana, R.; González-Arenzana, L.; Portu, J.; Garijo, P.; López-Alfaro, I.; López, R.; Santamaría, P.; Gutiérrez, A.R. Wine aroma evolution throughout alcoholic fermentation sequentially inoculated with non-Saccharomyces/Saccharomyces yeasts. Food Res. Int. 2018, 112, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Belda, I.; Navascués, E.; Marquina, D.; Santos, A.; Calderon, F.; Benito, S. Dynamic analysis of physiological properties of Torulaspora delbrueckii in wine fermentations and its incidence on wine quality. Appl. Microbiol. Biotechnol. 2015, 99, 1911–1922. [Google Scholar] [CrossRef]
- Ochando, T.; Mouret, J.R.; Humbert-Goffard, A.; Aguera, E.; Sablayrolles, J.M.; Farines, V. Comprehensive study of the dynamic interaction between SO2 and acetaldehyde during alcoholic fermentation. Food Res. Int. 2020, 136, 109607. [Google Scholar] [CrossRef]
- Glories, Y. La couleur des vins rouges. 2e partie: Mesure, origine et interprétation. OENO One 1984, 18, 253. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SS | S | TL | |
|---|---|---|---|
| Ethanol (% v/v) | 12.6 | 13.07 | 12.87 |
| pH | 4.14 b | 4.13 b | 3.98 a |
| Total acidity | 4.47 a | 4.74 a | 5.82 b |
| Volatile acidity | 0.49 | 0.50 | 0.58 |
| Tartaric acid (g/L) | 1.49 | 1.66 | 1.65 |
| Malic acid (g/L) | 0.10 | 0.08 | 0.13 |
| Lactic acid (g/L) | 2.27 a | 2.33 a | 3.63 b |
| Free SO2 | 40.28 | <5 | <5 |
| Total SO2 | 96.50 | <20 | <20 |
| Glycerol | 9.0 a | 9.87 b | 9.87 b |
| Acetaldehyde | 0.49 a | 3.59 b | 3.00 b |
| SS | S | TL | |
|---|---|---|---|
| Anthocyanins (mg/L) | 743.2 c | 673.06 bc | 631.69 ab |
| CI | 5.89 a | 7.68 b | 8.04 b |
| TPI | 46.27 b | 41.85 ab | 38.83 a |
| Hue | 0.792 b | 0.762 b | 0.686 a |
| Ionization index (%) | 7.62 a | 12.52 b | 14.81 c |
| Polymerization index | 1.24 | 1.30 | 1.30 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Escribano-Viana, R.; González-Arenzana, L.; Garijo, P.; Fernández, L.; López, R.; Santamaría, P.; Gutiérrez, A.R. Bioprotective Effect of a Torulaspora delbrueckii/Lachancea thermotolerans-Mixed Inoculum in Red Winemaking. Fermentation 2022, 8, 337. https://doi.org/10.3390/fermentation8070337
Escribano-Viana R, González-Arenzana L, Garijo P, Fernández L, López R, Santamaría P, Gutiérrez AR. Bioprotective Effect of a Torulaspora delbrueckii/Lachancea thermotolerans-Mixed Inoculum in Red Winemaking. Fermentation. 2022; 8(7):337. https://doi.org/10.3390/fermentation8070337
Chicago/Turabian StyleEscribano-Viana, Rocío, Lucía González-Arenzana, Patrocinio Garijo, Laura Fernández, Rosa López, Pilar Santamaría, and Ana Rosa Gutiérrez. 2022. "Bioprotective Effect of a Torulaspora delbrueckii/Lachancea thermotolerans-Mixed Inoculum in Red Winemaking" Fermentation 8, no. 7: 337. https://doi.org/10.3390/fermentation8070337
APA StyleEscribano-Viana, R., González-Arenzana, L., Garijo, P., Fernández, L., López, R., Santamaría, P., & Gutiérrez, A. R. (2022). Bioprotective Effect of a Torulaspora delbrueckii/Lachancea thermotolerans-Mixed Inoculum in Red Winemaking. Fermentation, 8(7), 337. https://doi.org/10.3390/fermentation8070337