
Dynamic Variations in Rumen Fermentation Characteristics and Bacterial Community Composition during In Vitro Fermentation


Abstract
1. Introduction
2. Materials and Methods
2.1. Composition of Fermentation Substrate
2.2. Rumen Fluid Collection and Experimental Design
2.3. In Vitro Incubation
2.4. Rumen Fermentation Characteristics Determination
2.5. Bacterial Community Analysis
2.6. Statistical Analysis
3. Results
3.1. Total Gas Production and Methane Production
3.2. Rumen Fermentation Characteristics
3.3. Alpha Diversity Metrics
3.4. Rumen Bacterial Community
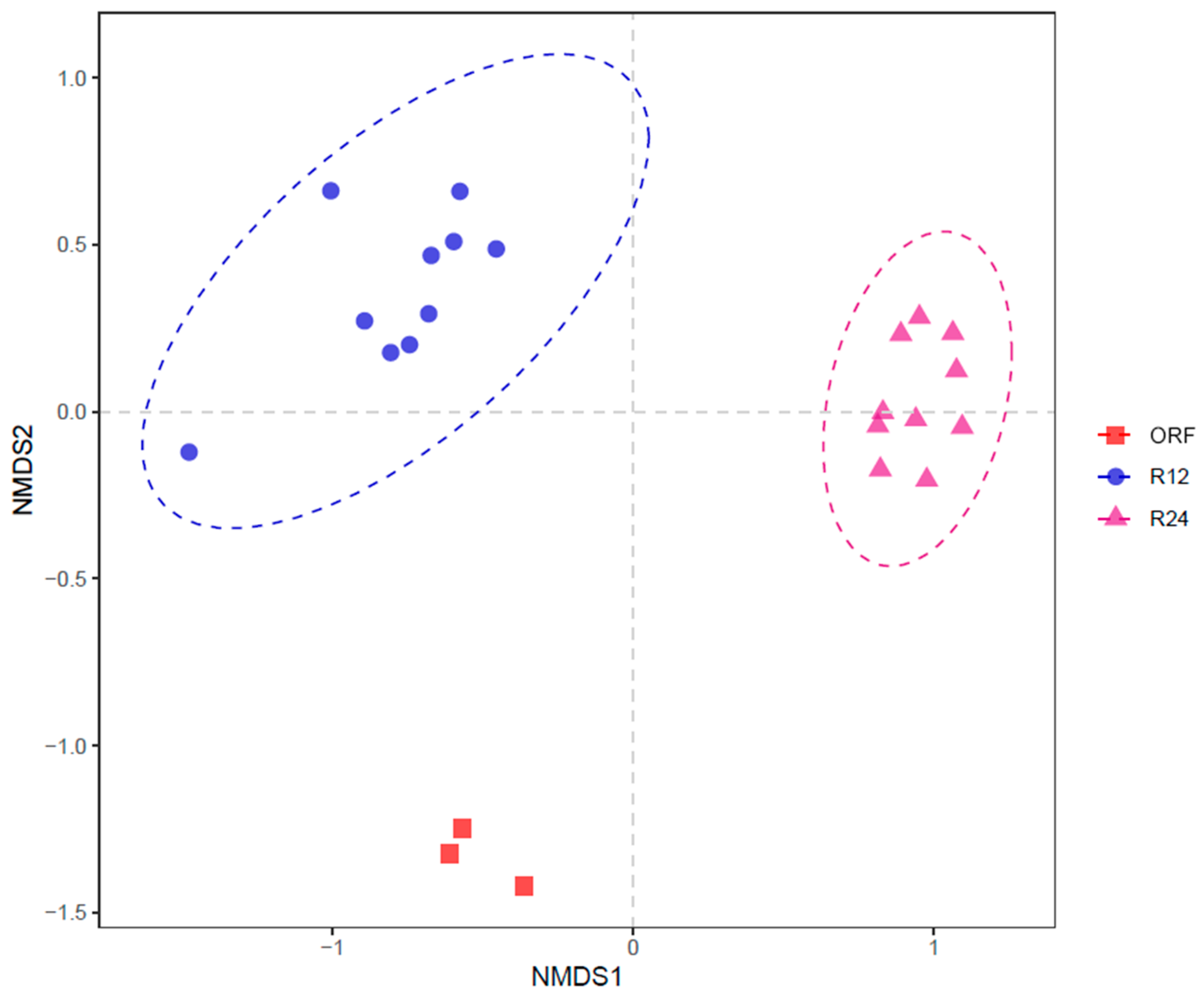
3.5. Beta Diversity
3.6. Correlation Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, E.T.; Hwang, H.S.; Lee, S.M.; Lee, S.J.; Lee, I.D.; Lee, S.K.; Oh, D.S.; Lim, J.H.; Yoon, H.B.; Jeong, H.Y.; et al. Effects of medicinal herb extracts on in vitro ruminal methanogenesis, microbe diversity and fermentation system. Asian-Australas. J. Anim. Sci. 2016, 29, 1280–1286. [Google Scholar] [CrossRef] [PubMed]
- Swift, R.W.; Hershberger, T.V.; Hartsook, E.W. Use of the artificial rumen technique to estimate the nutritive value of forages. J. Anim. Sci. 1959, 18, 1189. [Google Scholar]
- Pigden, W.J.; Bell, J.M. The artificial rumen as a procedure for evaluating forage quality. J. Anim. Sci. 1955, 14, 1239–1240. [Google Scholar]
- Wang, W.; Wu, Q.; Li, W.; Wang, Y.; Zhang, F.; Lv, L.; Li, S.; Yang, H. High-gossypol whole cottonseed exhibited mediocre rumen degradability and less microbial fermentation efficiency than cottonseed hull and cottonseed meal with an in vitro gas production technique. Fermentation 2022, 8, 103. [Google Scholar] [CrossRef]
- Keim, J.P.; Berthiaume, R.; Pacheco, D.; Muetzel, S. Comparison of rumen in vitro fermentation of temperate pastures using different batch culture systems. Anim. Prod. Sci. 2017, 57, 690–696. [Google Scholar] [CrossRef]
- Odenyo, A.A.; McSweeney, C.S.; Palmer, B.; Negassa, D.; Osuji, P.O. In vitro screening of rumen fluid samples from indigenous African ruminants provides evidence for rumen fluid with superior capacities to digest tannin-rich fodders. Aust. J. Agric. Res. 1999, 50, 1147–1157. [Google Scholar] [CrossRef]
- McSweeney, C.S.; Palmer, B.; Bunch, R.; Krause, D.O. In vitro quality assessment of tannin-containing tropical shrub legumes: Protein and fibre digestion. Anim. Feed Sci. Technol. 1999, 82, 227–241. [Google Scholar] [CrossRef]
- Remy, J.; Armstrong, S. Effect of substrate to inoculum ratio on outcomes of in vitro rumen fermentation. In Proceedings of the ADSA Annual Meeting, Cincinnati, OH, USA, 23–26 June 2019. [Google Scholar]
- Gülşen, N.; Arik, H.; Hayirli, A.; Alatas, M.; Aksoy, M. Utilization of cryopreserved ruminal liquor in in vitro gas production technique for evaluating nutritive value of some feedstuffs. Kafkas Univ. Vet. Fak. Derg. 2017, 23, 325–332. [Google Scholar]
- Broudiscou, L.P.; Offner, A.; Sauvant, D. Effects of inoculum source, pH, redox potential and headspace di-hydrogen on rumen in vitro fermentation yields. Animal 2014, 8, 931–937. [Google Scholar] [CrossRef]
- Onime, L.; Zanfi, C.; Agostinis, C.; Bulla, R.; Spanghero, M. The use of quantitative real time polymerase chain reaction to quantify some rumen bacterial strains in an in vitro rumen system. Ital. J. Anim. Sci. 2013, 12, e58. [Google Scholar] [CrossRef]
- Váradyová, Z.; Štyriaková, I.; Kišidayová, S. Effect of natural dolomites on the in vitro fermentation and rumen protozoan population using rumen fluid and fresh faeces inoculum from sheep. Small Rumin. Res. 2007, 73, 58–66. [Google Scholar] [CrossRef]
- Boguhn, J.; Kluth, H.; Rodehutscord, M. Effect of total mixed ration composition on fermentation and efficiency of ruminal microbial crude protein synthesis in vitro. J. Dairy Sci. 2006, 89, 1580–1591. [Google Scholar] [CrossRef]
- Dyne, G.M.V.; Haug, P.T. Variables Affecting In Vitro Rumen Fermentation Studies in Forage Evaluation: An Annotated Bibliography; Oak Ridge National Lab., Tenn.: Oak Ridge, TN, USA, 1968. [Google Scholar] [CrossRef]
- Church, D.C.; Petersen, R.G. Effect of several variables on in vitro rumen fermentation. J. Dairy Sci. 1960, 43, 81–92. [Google Scholar] [CrossRef]
- Cutrignelli, M.I.; Calabrò, S.; Tudisco, R.; Zicarelli, F.; Gazaneo, M.P.; Piccolo, V. Comparison of buffalo rumen liquor and buffalo faeces as inoculum for the in vitro gas production technique. Ital. J. Anim. Sci. 2005, 4, 319–321. [Google Scholar] [CrossRef]
- Paz, H.A.; Anderson, C.L.; Muller, M.J.; Kononoff, P.J.; Fernando, S.C. Rumen bacterial community composition in Holstein and Jersey cows is different under same dietary condition and is not affected by sampling method. Front. Microbiol. 2016, 7, 1206. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Morales, E.; Arco-Pérez, A.; Martín-García, A.I.; Yáñez-Ruiz, D.R.; Frutos, P.; Hervás, G. Use of stomach tubing as an alternative to rumen cannulation to study ruminal fermentation and microbiota in sheep and goats. Anim. Feed Sci. Technol. 2014, 198, 57–66. [Google Scholar] [CrossRef]
- Mlambo, V.; Sikosana, J.L.N.; Mould, F.L.; Smith, T.; Owen, E.; Mueller-Harvey, I. The effectiveness of adapted rumen fluid versus PEG to ferment tannin-containing substrates in vitro. Anim. Feed Sci. Technol. 2007, 136, 128–136. [Google Scholar] [CrossRef]
- Zheng, Y.; He, T.; Xie, T.; Wang, J.; Yang, Z.; Sun, X.; Wang, W.; Li, S. Hydroxy-selenomethionine supplementation promotes the in vitro rumen fermentation of dairy cows by altering the relative abundance of rumen microorganisms. J. Appl. Microbiol. 2022, 132, 2583–2593. [Google Scholar] [CrossRef]
- Zhang, X.; Dong, X.; Wanapat, M.; Shah, A.M.; Luo, X.; Peng, Q.; Kang, K.; Hu, R.; Guan, J.; Wang, Z. Ruminal pH pattern, fermentation characteristics and related bacteria in response to dietary live yeast (Saccharomyces cerevisiae) supplementation in beef cattle. Anim. Biosci. 2022, 35, 184–195. [Google Scholar] [CrossRef]
- Kim, J.N.; Song, J.; Kim, E.J.; Chang, J.; Kim, C.-H.; Seo, S.; Chang, M.B.; Bae, G.-S. Effects of short-term fasting on in vivo rumen microbiota and in vitro rumen fermentation characteristics. Asian-Australas. J. Anim. Sci. 2019, 32, 776–782. [Google Scholar] [CrossRef]
- Ramos, S.C.; Jeong, C.-D.; Mamuad, L.L.; Kim, S.-H.; Son, A.R.; Miguel, M.A.; Islam, M.; Cho, Y.-I.; Lee, S.-S. Enhanced ruminal fermentation parameters and altered rumen bacterial community composition by formulated rumen buffer agents fed to dairy cows with a high-concentrate diet. Agriculture 2021, 11, 554. [Google Scholar] [CrossRef]
- Wang, Y.; Alexander, T.W.; McAllister, T.A. In vitro effects of phlorotannins from Ascophyllum nodosum (brown seaweed) on rumen bacterial populations and fermentation. J. Sci. Food Agric. 2009, 89, 2252–2260. [Google Scholar] [CrossRef]
- Kang, J.; Zeng, B.; Tang, S.; Wang, M.; Han, X.; Zhou, C.; Yan, Q.; Liu, J.; Tan, Z. Effects of Momordica charantia polysaccharide on in vitro ruminal fermentation and cellulolytic bacteria. Ital. J. Anim. Sci. 2017, 16, 226–233. [Google Scholar] [CrossRef]
- Ruiz, O.; Castillo, Y.; Arzola, C.; Burrola, E.; Salinas, J.; Corral, A.; Hume, M.E.; Murillo, M.; Itza, M. Effects of Candida norvegensis live cells on in vitro oat straw rumen fermentation. Asian-Australas. J. Anim. Sci. 2016, 29, 211–218. [Google Scholar] [CrossRef]
- Yarza, P.; Yilmaz, P.; Pruesse, E.; Glöckner, F.O.; Ludwig, W.; Schleifer, K.-H.; Whitman, W.B.; Euzéby, J.; Amann, R.; Rosselló-Móra, R. Uniting the classification of cultured and uncultured bacteria and archaea using 16S rRNA gene sequences. Nat. Rev. Microbiol. 2014, 12, 635–645. [Google Scholar] [CrossRef]
- Huang, X.; Mi, J.; Denman, S.E.; Basangwangdui; Pingcuozhandui; Zhang, Q.; Long, R.; McSweeney, C.S. Changes in rumen microbial community composition in yak in response to seasonal variations. J. Appl. Microbiol. 2022, 132, 1652–1665. [Google Scholar] [CrossRef]
- Denman, S.E.; Martinez Fernandez, G.; Shinkai, T.; Mitsumori, M.; McSweeney, C.S. Metagenomic analysis of the rumen microbial community following inhibition of methane formation by a halogenated methane analog. Front. Microbiol. 2015, 6, 1087. [Google Scholar] [CrossRef]
- Kang, S.H.; Evans, P.; Morrison, M.; McSweeney, C. Identification of metabolically active proteobacterial and archaeal communities in the rumen by DNA- and RNA-derived 16S rRNA gene. J. Appl. Microbiol. 2013, 115, 644–653. [Google Scholar] [CrossRef]
- Krause, D.O.; Smith, W.J.M.; Ryan, F.M.E.; Mackie, R.I.; McSweeney, C.S. Use of 16S-rRNA based techniques to investigate the ecological succession of microbial populations in the immature lamb rumen: Tracking of a specific strain of inoculated ruminococcus and interactions with other microbial populations in vivo. Microb. Ecol. 1999, 38, 365–376. [Google Scholar] [CrossRef]
- Gilbert, R.A.; Netzel, G.; Chandra, K.; Ouwerkerk, D.; Fletcher, M.T. Degradation of the indospicine toxin from Indigofera spicata by a mixed population of rumen bacteria. Toxins 2021, 13, 389. [Google Scholar] [CrossRef]
- Zheng, Y.H.; Zhao, Y.Y.; Xue, S.L.; Wang, W.; Wang, Y.J.; Cao, Z.J.; Yang, H.J.; Li, S.L. Feeding value assessment of substituting cassava (Manihot esculenta) residue for concentrate of dairy cows using an in vitro gas test. Animals 2021, 11, 307. [Google Scholar] [CrossRef] [PubMed]
- Broderick, G.A.; Kang, J.H. Automated simultaneous determination of ammonia and total amino acids in ruminal fluid and in vitro media. J. Dairy Sci. 1980, 63, 64–75. [Google Scholar] [CrossRef]
- Makkar, H.P.; Sharma, O.P.; Dawra, R.K.; Negi, S.S. Simple determination of microbial protein in rumen liquor. J. Dairy Sci. 1982, 65, 2170–2173. [Google Scholar] [CrossRef]
- Qiu, Q.; Wei, X.; Zhang, L.; Li, Y.; Qu, M.; Ouyang, K. Effect of dietary inclusion of tea residue and tea leaves on ruminal fermentation characteristics and methane production. Anim. Biotechnol. 2021; in press. [Google Scholar]
- Moss, A.R.; Jouany, J.P.; Newbold, J. Methane production by ruminants: Its contribution to global warming. Ann. Zootech. 2000, 49, 231–253. [Google Scholar] [CrossRef]
- Qiu, Q.; Gao, C.; Aziz ur Rahman, M.; Cao, B.; Su, H. Digestive ability, physiological characteristics, and rumen bacterial community of Holstein finishing steers in response to three nutrient density diets as fattening phases advanced. Microorganisms 2020, 8, 335. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Qiong, W.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar]
- Dijkstra, J. Production and absorption of volatile fatty-acids in the rumen. Livest. Prod. Sci. 1994, 39, 61–69. [Google Scholar] [CrossRef]
- Kozakai, K.; Nakamura, T.; Kobayashi, Y.; Tanigawa, T.; Osaka, I.; Kawamoto, S.; Hara, S. Effect of mechanical processing of corn silage on in vitro ruminal fermentation, and in situ bacterial colonization and dry matter degradation. Can. J. Anim. Sci. 2007, 87, 259–267. [Google Scholar] [CrossRef]
- Qiu, Q.; Gao, C.; Gao, Z.; Muhammad Aziz ur, R.; He, Y.; Cao, B.; Su, H. Temporal dynamics in rumen bacterial community composition of finishing steers during an adaptation period of three months. Microorganisms 2019, 7, 410. [Google Scholar] [CrossRef] [PubMed]
- Hook, S.E.; Steele, M.A.; Northwood, K.S.; Dijkstra, J.; France, J.; Wright, A.D.G.; McBride, B.W. Impact of subacute ruminal acidosis (SARA) adaptation and recovery on the density and diversity of bacteria in the rumen of dairy cows. FEMS Microbiol. Ecol. 2011, 78, 275–284. [Google Scholar] [CrossRef]
- Dunfield, P.F.; Yuryev, A.; Senin, P.; Smirnova, A.V.; Stott, M.B.; Hou, S.; Ly, B.; Saw, J.H.; Zhou, Z.; Ren, Y.; et al. Methane oxidation by an extremely acidophilic bacterium of the phylum Verrucomicrobia. Nature 2007, 450, 879–882. [Google Scholar] [CrossRef]
- McCabe, M.S.; Cormican, P.; Keogh, K.; O’Connor, A.; O’Hara, E.; Palladino, R.A.; Kenny, D.A.; Waters, S.M. Illumina MiSeq phylogenetic amplicon sequencing shows a large reduction of an uncharacterised succinivibrionaceae and an increase of the Methanobrevibacter gottschalkii clade in feed restricted cattle. PLoS ONE 2015, 10, e0133234. [Google Scholar] [CrossRef]
- Pope, P.B.; Smith, W.; Denman, S.E.; Tringe, S.G.; Barry, K.; Hugenholtz, P.; McSweeney, C.S.; McHardy, A.C.; Morrison, M. Isolation of Succinivibrionaceae implicated in low methane emissions from Tammar wallabies. Science 2011, 333, 646–648. [Google Scholar] [CrossRef] [PubMed]
- Pitta, D.W.; Pinchak, W.E.; Dowd, S.E.; Osterstock, J.; Gontcharova, V.; Youn, E.; Dorton, K.; Yoon, I.; Min, B.R.; Fulford, J.D.; et al. Rumen bacterial diversity dynamics associated with changing from bermudagrass hay to grazed winter wheat diets. Microb. Ecol. 2010, 59, 511–522. [Google Scholar] [CrossRef]
- Braz, G.H.R.; Fernandez-Gonzalez, N.; Lema, J.M.; Carballa, M. Organic overloading affects the microbial interactions during anaerobic digestion in sewage sludge reactors. Chemosphere 2019, 222, 323–332. [Google Scholar] [CrossRef]
- Greses, S.; Gaby, J.C.; Aguado, D.; Ferrer, J.; Seco, A.; Horn, S.J. Microbial community characterization during anaerobic digestion of Scenedesmus spp. under mesophilic and thermophilic conditions. Algal Res. 2017, 27, 121–130. [Google Scholar] [CrossRef]
- Gonzalez-Fernandez, C.; Barreiro-Vescovo, S.; de Godos, I.; Fernandez, M.; Zouhayr, A.; Ballesteros, M. Biochemical methane potential of microalgae biomass using different microbial inocula. Biotechnol. Biofuels 2018, 11, 184. [Google Scholar] [CrossRef]
- Jin, W.; Xue, C.; Liu, J.; Yin, Y.; Zhu, W.; Mao, S. Effects of disodium fumarate on in vitro rumen fermentation, the production of lipopolysaccharide and biogenic amines, and the rumen bacterial community. Curr. Microbiol. 2017, 74, 1337–1342. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.B.; Feng, Y.L. A study on the relationship between VFA production and fermentable organic matter in the rumen. Acta Zoonutrimenta Sin. 1996, 8, 32–36. [Google Scholar]
- Chen, R.; Li, Z.; Feng, J.; Zhao, L.; Yu, J. Effects of digestate recirculation ratios on biogas production and methane yield of continuous dry anaerobic digestion. Bioresour. Technol. 2020, 316, 123963. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Guo, C.; Wang, L.; Chen, F.; Dong, X.; Li, X.; Ni, K.; Yang, F. Effects of paper mulberry silage on the growth performance, rumen microbiota and muscle fatty acid composition in Hu lambs. Fermentation 2021, 7, 286. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fermentation Substrate | DM % |
|---|---|
| Ingredients | |
| Corn silage | 23.87 |
| Alfalfa hay | 18.56 |
| Oat hay | 2.64 |
| Corn | 25.69 |
| Soybean meal | 3.01 |
| wheat | 7.65 |
| wheat bran | 2.67 |
| Beet pulp | 1.81 |
| Molasses | 2.54 |
| Cottonseed | 8.53 |
| Fat-energy powder | 1.29 |
| Dicalcium phosphate | 0.62 |
| Salt | 0.57 |
| Premix 1 | 0.55 |
| Total | 100.00 |
| Nutrient composition | |
| Crude protein (CP) | 16.90 |
| Net energy for lactation (NEL), Mcal/kg | 1.75 |
| Neutral detergent fiber (NDF) | 31.01 |
| Acid detergent fiber (ADF) | 23.10 |
| Non-fibrous carbohydrate (NFC) | 40.27 |
| Crude fat | 5.56 |
| Calcium | 0.85 |
| Phosphorus | 0.42 |
| Item 1 | ORF 2 | R12 2 | R24 2 | SEM 3 | p-Value 4 |
|---|---|---|---|---|---|
| pH value | 6.84 | 7.60 a | 7.32 b | 0.047 | 0.001 |
| Acetate, mM | 37.8 | 29.7 b | 33.7 a | 0.516 | <0.001 |
| Propionate, mM | 10.3 | 11.6 b | 13.3 a | 0.221 | <0.001 |
| Isobutyrate, mM | 0.39 | 0.19 b | 0.27 a | 0.009 | <0.001 |
| Butyrate, mM | 4.10 | 3.64 b | 4.10 a | 0.083 | 0.003 |
| Isovalerate, mM | 0.48 | 0.60 b | 0.85 a | 0.032 | <0.001 |
| Valerate, mM | 0.57 | 0.95 b | 1.08 a | 0.025 | 0.005 |
| Branched-chain volatile fatty acids, mM | 1.43 | 1.73 b | 2.20 a | 0.063 | <0.001 |
| Total volatile fatty acids, mM | 53.6 | 46.7 b | 53.2 a | 0.825 | <0.001 |
| Microbial crude protein, mg/L | ND 5 | 121.8 b | 209.4 a | 10.5 | <0.001 |
| Ammonia nitrogen, mg/dL Lactate, mg/L | ND 5 ND 5 | 2.32 b 8.25 | 7.21 a 8.79 | 0.675 0.432 | <0.001 0.542 |
| Acetate to propionate ratio | 3.66 | 2.56 | 2.54 | 0.027 | 0.699 |
| Non-glucogenic to glucogenic acids ratio | 4.27 | 3.03 | 3.00 | 0.036 | 0.688 |
| Fermentation efficiency | 0.73 | 0.76 | 0.76 | 0.0008 | 0.746 |
| Item | ORF | R12 | R24 | SEM | p-Value |
|---|---|---|---|---|---|
| Chao 1 | 3161 | 2849 | 2972 | 58.5 | 0.305 |
| Observed species | 2333 | 2052 | 2225 | 52.2 | 0.099 |
| PD whole tree 1 | 163.8 | 143.5 b | 154.8 a | 2.82 | 0.042 |
| Shannon index | 8.41 | 7.78 b | 8.26 a | 0.075 | <0.001 |
| Simpson index | 0.990 | 0.980 b | 0.989 a | 0.001 | <0.001 |
| Phylum | ORF | R12 | R24 | SEM | p-Value |
|---|---|---|---|---|---|
| Bacteroidetes | 61.63 | 59.78 b | 66.62 a | 1.299 | 0.005 |
| Firmicutes | 20.86 | 13.61 b | 22.28 a | 1.216 | <0.001 |
| Proteobacteria | 10.66 | 21.34 a | 5.33 b | 1.920 | <0.001 |
| Saccharibacteria | 1.79 | 0.98 | 1.04 | 0.094 | 0.757 |
| Tenericutes | 2.60 | 0.97 | 0.77 | 0.079 | 0.220 |
| Fibrobacteres | 0.95 | 1.16 a | 0.85 b | 0.071 | 0.009 |
| Cyanobacteria | 0.20 | 0.59 b | 0.87 a | 0.067 | 0.028 |
| Spirochaetae | 0.19 | 0.56 | 0.64 | 0.062 | 0.521 |
| Verrucomicrobia | 0.31 | 0.28 b | 0.62 a | 0.086 | 0.042 |
| SR1 Absconditabacteria | 0.40 | 0.34 | 0.31 | 0.039 | 0.737 |
| Lentisphaerae | 0.07 | 0.13 b | 0.25 a | 0.026 | 0.010 |
| Elusimicrobia | 0.04 | 0.13 | 0.17 | 0.015 | 0.283 |
| Synergistetes | 0.05 | 0.07 b | 0.14 a | 0.017 | 0.028 |
| Genus | ORF | R12 | R24 | SEM | p-Value |
|---|---|---|---|---|---|
| Prevotella 1 | 39.40 | 41.91 | 42.51 | 1.244 | 0.817 |
| Succinivibrionaceae UCG-002 | 7.49 | 10.65 a | 2.09 b | 1.005 | <0.001 |
| Rikenellaceae RC9 gut group | 5.05 | 4.22 b | 6.53 a | 0.362 | <0.001 |
| Succiniclasticum | 1.76 | 3.05 b | 5.65 a | 0.547 | 0.013 |
| Ruminobacter | 1.58 | 7.28 a | 0.37 b | 0.842 | <0.001 |
| Prevotellaceae UCG-003 | 3.79 | 2.27 b | 2.74 a | 0.111 | 0.032 |
| Christensenellaceae R-7 group | 3.40 | 1.42 b | 2.83 a | 0.210 | <0.001 |
| Prevotellaceae UCG-001 | 2.30 | 1.26 | 1.2 | 0.114 | 0.987 |
| Candidatus Saccharimonas | 1.79 | 0.98 | 1.04 | 0.094 | 0.757 |
| Fibrobacter | 0.95 | 1.16 a | 0.85 b | 0.071 | 0.009 |
| Ruminococcaceae UCG-014 | 1.50 | 1.04 | 0.73 | 0.098 | 0.119 |
| Succinivibrio | 0.25 | 0.75 | 0.97 | 0.071 | 0.129 |
| Ruminococcaceae UCG-002 | 0.09 | 0.38 b | 1.13 a | 0.095 | <0.001 |
| Veillonellaceae UCG-001 | 0.73 | 0.40 b | 0.84 a | 0.089 | 0.010 |
| Ruminococcaceae NK4A214 group | 0.71 | 0.42 b | 0.70 a | 0.062 | 0.019 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, X.; Ouyang, K.; Long, T.; Liu, Z.; Li, Y.; Qiu, Q. Dynamic Variations in Rumen Fermentation Characteristics and Bacterial Community Composition during In Vitro Fermentation. Fermentation 2022, 8, 276. https://doi.org/10.3390/fermentation8060276
Wei X, Ouyang K, Long T, Liu Z, Li Y, Qiu Q. Dynamic Variations in Rumen Fermentation Characteristics and Bacterial Community Composition during In Vitro Fermentation. Fermentation. 2022; 8(6):276. https://doi.org/10.3390/fermentation8060276
Chicago/Turabian StyleWei, Xiao, Kehui Ouyang, Tanghui Long, Zuogui Liu, Yanjiao Li, and Qinghua Qiu. 2022. "Dynamic Variations in Rumen Fermentation Characteristics and Bacterial Community Composition during In Vitro Fermentation" Fermentation 8, no. 6: 276. https://doi.org/10.3390/fermentation8060276
APA StyleWei, X., Ouyang, K., Long, T., Liu, Z., Li, Y., & Qiu, Q. (2022). Dynamic Variations in Rumen Fermentation Characteristics and Bacterial Community Composition during In Vitro Fermentation. Fermentation, 8(6), 276. https://doi.org/10.3390/fermentation8060276