
Biocontrol and Enzymatic Activity of Non-Saccharomyces Wine Yeasts: Improvements in Winemaking

 , and
, and
Abstract
1. Introduction
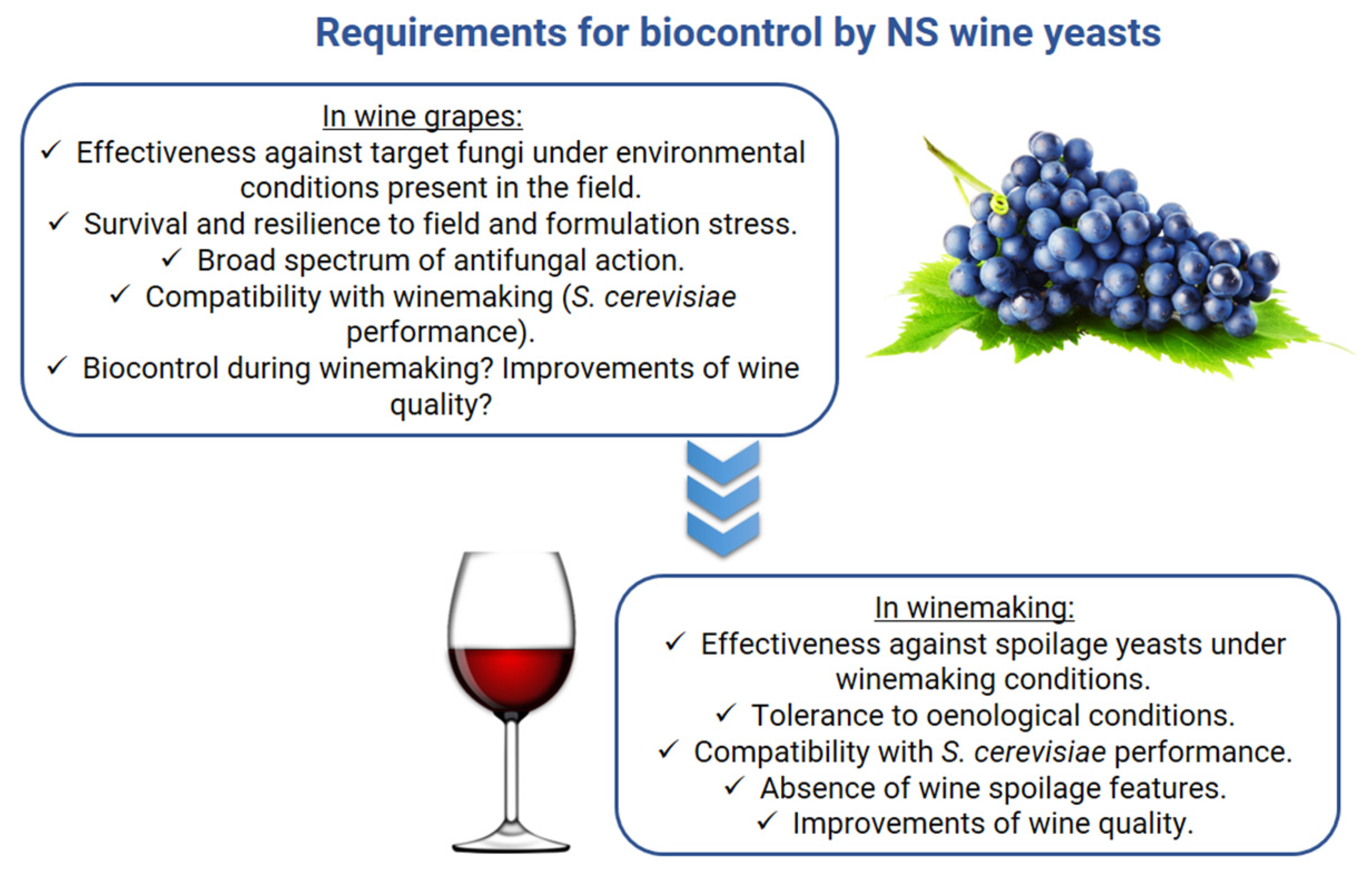
2. Biocontrol Exerted by Non-Saccharomyces Wine Yeasts
2.1. Biocontrol in Wine Grapes
2.1.1. The Importance of Studying Pathogens and the Influencing Environmental Conditions of Non-Saccharomyces BCA Applied to Wine Grapes
2.1.2. Improvements in Non-Saccharomyces BCAs Applied to Wine Grapes
2.2. Biocontrol in Winemaking
2.2.1. Non-Saccharomyces Wine Yeasts as Sources of Antimicrobial Metabolites for Biocontrol in Winemaking
2.2.2. Non-Saccharomyces Wine Yeasts as Selected Strains for Biocontrol in Winemaking
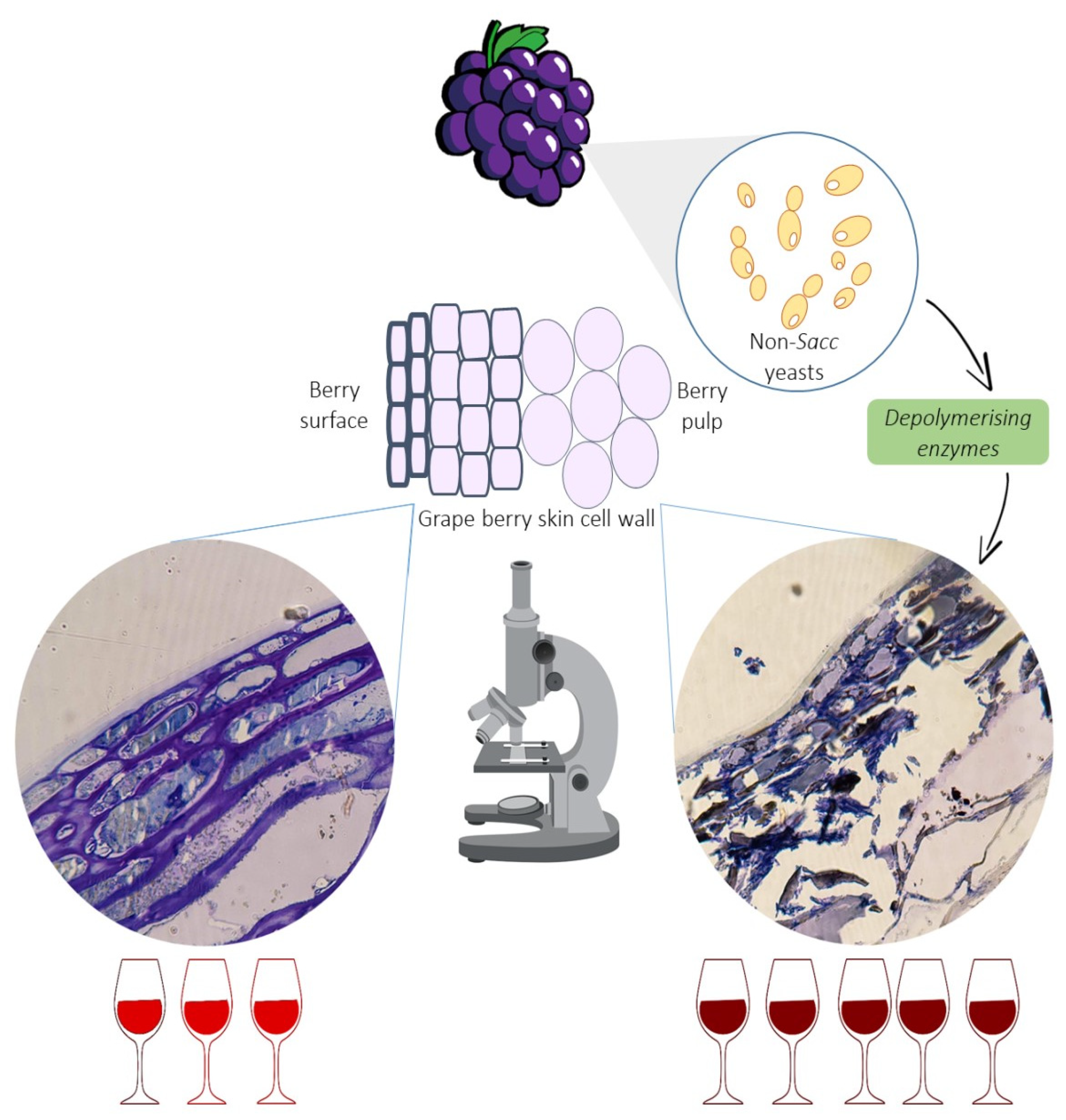
3. Grape Cell Wall Depolymerising Enzymes
3.1. Non-Saccharomyces Wine Yeasts as Alternative Sources of Enzymes
3.1.1. Exogenous Enzymatic Extracts Produced by Non-Saccharomyces Yeasts
3.1.2. Non-Saccharomyces Wine Yeasts for In Situ Enzyme Production in Winemaking
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- OIV International Organisation of Vine and Wine. International Code of Oenological Practices; OIV International Organisation of Vine and Wine: Dijon, France, 2023; ISBN 978-2-85038-071-6. [Google Scholar]
- OIV International Organisation of Vine and Wine. State of the World Vine and Wine Sector in 2022. Available online: https://www.oiv.int/sites/default/files/documents/2023_SWVWS_report_EN.pdf (accessed on 27 January 2024).
- Nardi, T. Microbial Resources as a Tool for Enhancing Sustainability in Winemaking. Microorganisms 2020, 8, 507. [Google Scholar] [CrossRef] [PubMed]
- IPCC. Climate Change 2014: Synthesis Report; IPCC: Geneva, Switzerland, 2014; pp. 1–112. [Google Scholar]
- Fazio, N.A.; Russo, N.; Foti, P.; Pino, A.; Caggia, C.; Randazzo, C.L. Inside Current Winemaking Challenges: Exploiting the Potential of Conventional and Unconventional Yeasts. Microorganisms 2023, 11, 1338. [Google Scholar] [CrossRef] [PubMed]
- Zilelidou, E.A.; Nisiotou, A. Understanding Wine through Yeast Interactions. Microorganisms 2021, 9, 1620. [Google Scholar] [CrossRef] [PubMed]
- Ellis, D.J.; Kerr, E.D.; Schenk, G.; Schulz, B.L. Metabolomics of Non-Saccharomyces Yeasts in Fermented Beverages. Beverages 2022, 8, 41. [Google Scholar] [CrossRef]
- Cordero-Bueso, G.; Mangieri, N.; Maghradze, D.; Foschino, R.; Valdetara, F.; Cantoral, J.M.; Vigentini, I. Wild Grape-Associated Yeasts as Promising Biocontrol Agents against Vitis vinifera Fungal Pathogens. Front. Microbiol. 2017, 8, 2025. [Google Scholar] [CrossRef] [PubMed]
- Marsico, A.D.; Velenosi, M.; Perniola, R.; Bergamini, C.; Sinonin, S.; David-Vaizant, V.; Maggiolini, F.A.M.; Hervè, A.; Cardone, M.F.; Ventura, M. Native Vineyard Non-Saccharomyces Yeasts Used for Biological Control of Botrytis cinerea in Stored Table Grape. Microorganisms 2021, 9, 457. [Google Scholar] [CrossRef] [PubMed]
- Nally, M.C.; Pesce, V.M.; Maturano, Y.P.; Rodriguez Assaf, L.A.; Toro, M.E.; Castellanos de Figueroa, L.I.; Vazquez, F. Antifungal Modes of Action of Saccharomyces and Other Biocontrol Yeasts against Fungi Isolated from Sour and Grey Rots. Int. J. Food Microbiol. 2015, 204, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Prendes, L.P.; Merín, M.G.; Fontana, A.R.; Bottini, R.A.; Ramirez, M.L.; Morata de Ambrosini, V.I. Isolation, Identification and Selection of Antagonistic Yeast against Alternaria alternata Infection and Tenuazonic Acid Production in Wine Grapes from Argentina. Int. J. Food Microbiol. 2018, 266, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Sabaghian, S.; Braschi, G.; Vannini, L.; Patrignani, F.; Samsulrizal, N.H.; Lanciotti, R. Isolation and Identification of Wild Yeast from Malaysian Grapevine and Evaluation of Their Potential Antimicrobial Activity against Grapevine Fungal Pathogens. Microorganisms 2021, 9, 2582. [Google Scholar] [CrossRef]
- Kuchen, B.; Maturano, Y.P.; Mestre, M.V.; Combina, M.; Toro, M.E.; Vazquez, F. Selection of Native Non-Saccharomyces Yeasts with Biocontrol Activity against Spoilage Yeasts in Order to Produce Healthy Regional Wines. Fermentation 2019, 5, 60. [Google Scholar] [CrossRef]
- Mewa-Ngongang, M.; du Plessis, H.W.; Hlangwani, E.; Ntwampe, S.K.O.; Chidi, B.S.; Hutchinson, U.F.; Jolly, N.P. Activity Interactions of Crude Biopreservatives against Spoilage Yeast Consortia. Fermentation 2019, 5, 53. [Google Scholar] [CrossRef]
- Villalba, M.L.; Susana Sáez, J.; del Monaco, S.; Lopes, C.A.; Sangorrín, M.P. TdKT, a New Killer Toxin Produced by Torulaspora delbrueckii Effective against Wine Spoilage Yeasts. Int. J. Food Microbiol. 2016, 217, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Altieri, V.; Rossi, V.; Fedele, G. Efficacy of Preharvest Application of Biocontrol Agents against Gray Mold in Grapevine. Front. Plant Sci. 2023, 14, 1154370. [Google Scholar] [CrossRef] [PubMed]
- Wilson, C.L.; Wisniewski, M.E. Biological Control of Postharvest Diseases of Fruits and Vegetables: An Emerging Technology*. Annu. Rev. Phytopathol. 1989, 27, 425–441. [Google Scholar] [CrossRef]
- Bassim Atta, M.; Ruiz-Larrea, F. Fungal Pectinases in Food Technology. In Pectins—The New-Old Polysaccharides; Masuelli, M.A., Ed.; IntechOpen: London, UK, 2022. [Google Scholar]
- Merín, M.G.; Morata de Ambrosini, V.I. Kinetic and Metabolic Behaviour of the Pectinolytic Strain Aureobasidium pullulans GM-R-22 during Pre-Fermentative Cold Maceration and Its Effect on Red Wine Quality. Int. J. Food Microbiol. 2018, 285, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Merín, M.G.; Morata de Ambrosini, V.I. Application of a Grape Surface Majority Pectinolytic Species, Aureobasidium pullulans, to Low-Temperature Red Winemaking: Development and Stability of Wine Colour. J. Wine Res. 2020, 31, 218–239. [Google Scholar] [CrossRef]
- Espejo, F. Role of Commercial Enzymes in Wine Production: A Critical Review of Recent Research. J. Food Sci. Technol. 2021, 58, 9–21. [Google Scholar] [CrossRef]
- Alimardani-Theuil, P.; Gainvors-Claisse, A.; Duchiron, F. Yeasts: An Attractive Source of Pectinases—From Gene Expression to Potential Applications: A Review. Process Biochem. 2011, 46, 1525–1537. [Google Scholar] [CrossRef]
- Welke, J.E. Fungal and Mycotoxin Problems in Grape Juice and Wine Industries. Curr. Opin. Food Sci. 2019, 29, 7–13. [Google Scholar] [CrossRef]
- Omotayo, O.P.; Omotayo, A.O.; Mwanza, M.; Babalola, O.O. Prevalence of Mycotoxins and Their Consequences on Human Health. Toxicol. Res. 2019, 35, 1–7. [Google Scholar] [CrossRef]
- Guo, W.; Fan, K.; Nie, D.; Meng, J.; Huang, Q.; Yang, J.; Shen, Y.; Tangni, E.K.; Zhao, Z.; Wu, Y.; et al. Development of a QuEChERS-Based UHPLC-MS/MS Method for Simultaneous Determination of Six Alternaria Toxins in Grapes. Toxins 2019, 11, 87. [Google Scholar] [CrossRef] [PubMed]
- Kassemeyer, H.-H. Fungi of Grapes. In Biology of Microorganisms on Grapes, in Must and in Wine; Springer International Publishing: Cham, Switzerland, 2017; pp. 103–132. [Google Scholar]
- Ostry, V.; Malir, F.; Cumova, M.; Kyrova, V.; Toman, J.; Grosse, Y.; Pospichalova, M.; Ruprich, J. Investigation of Patulin and Citrinin in Grape Must and Wine from Grapes Naturally Contaminated by Strains of Penicillium expansum. Food Chem. Toxicol. 2018, 118, 805–811. [Google Scholar] [CrossRef] [PubMed]
- Prendes, L.P.; Fontana, A.R.; Merín, M.G.; D´ Amario Fernández, A.; Bottini, R.; Ramirez, M.L.; Morata de Ambrosini, V.I. Natural Occurrence and Production of Tenuazonic Acid in Wine Grapes in Argentina. Food Sci. Nutr. 2018, 6, 523–531. [Google Scholar] [CrossRef] [PubMed]
- Barata, A.; Malfeito-Ferreira, M.; Loureiro, V. The Microbial Ecology of Wine Grape Berries. Int. J. Food Microbiol. 2012, 153, 243–259. [Google Scholar] [CrossRef] [PubMed]
- Lemos, W.J.; Bovo, B.; Nadai, C.; Crosato, G.; Carlot, M.; Favaron, F.; Giacomini, A.; Corich, V. Biocontrol Ability and Action Mechanism of Starmerella bacillaris (Synonym Candida zemplinina) Isolated from Wine Musts against Gray Mold Disease Agent Botrytis cinerea on Grape and Their Effects on Alcoholic Fermentation. Front. Microbiol. 2016, 7, 1249. [Google Scholar] [CrossRef] [PubMed]
- Freimoser, F.M.; Rueda-Mejia, M.P.; Tilocca, B.; Migheli, Q. Biocontrol Yeasts: Mechanisms and Applications. World J. Microbiol. Biotechnol. 2019, 35, 154. [Google Scholar] [CrossRef] [PubMed]
- Esteves, M.; Lage, P.; Sousa, J.; Centeno, F.; de Fátima Teixeira, M.; Tenreiro, R.; Mendes-Ferreira, A. Biocontrol Potential of Wine Yeasts against Four Grape Phytopathogenic Fungi Disclosed by Time-Course Monitoring of Inhibitory Activities. Front. Microbiol. 2023, 14, 1146065. [Google Scholar] [CrossRef]
- Carbó, A.; Torres, R.; Teixidó, N.; Usall, J.; Magan, N.; Medina, A. Predicted Ecological Niches and Environmental Resilience of Different Formulations of the Biocontrol Yeast Candida sake CPA-1 Using the Bioscreen C. BioControl 2018, 63, 855–866. [Google Scholar] [CrossRef]
- Lahlali, R.; Ezrari, S.; Radouane, N.; Kenfaoui, J.; Esmaeel, Q.; El Hamss, H.; Belabess, Z.; Barka, E.A. Biological Control of Plant Pathogens: A Global Perspective. Microorganisms 2022, 10, 596. [Google Scholar] [CrossRef]
- Solairaj, D.; Yang, Q.; Guillaume Legrand, N.N.; Routledge, M.N.; Zhang, H. Molecular Explication of Grape Berry-Fungal Infections and Their Potential Application in Recent Postharvest Infection Control Strategies. Trends Food Sci. Technol. 2021, 116, 903–917. [Google Scholar] [CrossRef]
- Ponsone, M.L.; Combina, M.; Dalcero, A.; Chulze, S. Ochratoxin A and Ochratoxigenic Aspergillus Species in Argentinean Wine Grapes Cultivated under Organic and Non-Organic Systems. Int. J. Food Microbiol. 2007, 114, 131–135. [Google Scholar] [CrossRef]
- Chiotta, M.L.; Ponsone, M.L.; Combina, M.; Torres, A.M.; Chulze, S.N. Aspergillus Section Nigri Species Isolated from Different Wine-Grape Growing Regions in Argentina. Int. J. Food Microbiol. 2009, 136, 137–141. [Google Scholar] [CrossRef]
- Ponsone, M.L.; Chiotta, M.L.; Combina, M.; Dalcero, A.; Chulze, S. Biocontrol as a Strategy to Reduce the Impact of Ochratoxin A and Aspergillus Section Nigri in Grapes. Int. J. Food Microbiol. 2011, 151, 70–77. [Google Scholar] [CrossRef]
- Bau, M.; Bragulat, M.R.; Abarca, M.L.; Minguez, S.; Cabañes, F.J. Ochratoxigenic Species from Spanish Wine Grapes. Int. J. Food Microbiol. 2005, 98, 125–130. [Google Scholar] [CrossRef]
- Bellí, N.; Bau, M.; Marín, S.; Abarca, M.L.; Ramos, A.J.; Bragulat, M.R. Mycobiota and Ochratoxin A Producing Fungi from Spanish Wine Grapes. Int. J. Food Microbiol. 2006, 111, S40–S45. [Google Scholar] [CrossRef]
- Rosa, C.A.D.R.; Palacios, V.; Combina, M.; Fraga, M.E.; Rekson, A.D.O.; Magnoli, C.E.; Dalcero, A.M. Potential Ochratoxin A Producers from Wine Grapes in Argentina and Brazil. Food Addit. Contam. 2002, 19, 408–414. [Google Scholar] [CrossRef]
- Magnoli, C.; Violante, M.; Combina, M.; Palacio, G.; Dalcero, A. Mycoflora and Ochratoxin-Producing Strains of Aspergillus Section Nigri in Wine Grapes in Argentina. Lett. Appl. Microbiol. 2003, 37, 179–184. [Google Scholar] [CrossRef]
- Medina, A.; Mateo, R.; López-Ocaña, L.; Valle-Algarra, F.M.; Jiménez, M. Study of Spanish Grape Mycobiota and Ochratoxin A Production by Isolates of Aspergillus tubingensis and Other Members of Aspergillus Section Nigri. Appl. Environ. Microbiol. 2005, 71, 4696–4702. [Google Scholar] [CrossRef]
- Prendes, L.P.; Merín, M.G.; Andreoni, M.A.; Ramirez, M.L.; Morata de Ambrosini, V.I. Mycobiota and Toxicogenic Alternaria spp. Strains in Malbec Wine Grapes from DOC San Rafael, Mendoza, Argentina. Food Control 2015, 57, 122–128. [Google Scholar] [CrossRef]
- Sage, L.; Krivobok, S.; Delbos, É.; Seigle-Murandi, F.; Creppy, E.E. Fungal Flora and Ochratoxin a Production in Grapes and Musts from France. J. Agric. Food Chem. 2002, 50, 1306–1311. [Google Scholar] [CrossRef]
- Tančinová, D.; Rybárik, Ä.; Mašková, Z.; Felšöciová, S.; Císarová, M. Endogenal Colonization of Grapes Berries. J. Microbiol. Biotechnol. Food Sci. 2015, 4, 69–73. [Google Scholar] [CrossRef]
- Fontana, A.R.; Prendes, L.P.; Morata, V.I.; Bottini, R. High-Throughput Modified QuEChERS Method for the Determination of the Mycotoxin Tenuazonic Acid in Wine Grapes. RSC Adv. 2016, 6, 95670–95679. [Google Scholar] [CrossRef]
- Alexander, J.; Benford, D.; Boobis, A.; Ceccatelli, S.; Cottrill, B.; Cravedi, J.; Farmer, P. Scientific Opinion on the Risks for Animal and Public Health Related to the Presence of Alternaria Toxins in Feed and Food. EFSA J. 2011, 9, 2407. [Google Scholar] [CrossRef]
- Arcella, D.; Eskola, M.; Gómez Ruiz, J.A. Dietary Exposure Assessment to Alternaria Toxins in the European Population. EFSA J. 2016, 14, e04654. [Google Scholar] [CrossRef]
- Dall’Asta, C.; Cirlini, M.; Falavigna, C. Mycotoxins from Alternaria. Adv. Mol. Toxicol. 2014, 8, 107–121. [Google Scholar]
- EC-European Commission. Commission Recommendation (EU) 2022/553 of 5 April 2022 on Monitoring the Presence of Alternaria Toxins in Food. Off. J. Eur. Communities 2022, 107, 90. [Google Scholar]
- Magan, N. Importance of Ecological Windows for Efficacy of Biocontrol Agents. In How Research Can Stimulate the Development of Commercial Biological Control Against Plant Diseases. Progress in Biological Control; De Cal, A., Melgarejo, P., Magan, N., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 1–14. [Google Scholar]
- Liu, J.; Sui, Y.; Wisniewski, M.; Droby, S.; Liu, Y. Review: Utilization of Antagonistic Yeasts to Manage Postharvest Fungal Diseases of Fruit. Int. J. Food Microbiol. 2013, 167, 153–160. [Google Scholar] [CrossRef]
- Sui, Y.; Wisniewski, M.; Droby, S.; Liu, J. Responses of Yeast Biocontrol Agents to Environmental Stress. Appl. Environ. Microbiol. 2015, 81, 2968–2975. [Google Scholar] [CrossRef]
- Prendes, L.P.; Zachetti, V.G.L.; Pereyra, A.; Morata de Ambrosini, V.I.; Ramirez, M.L. Water Activity and Temperature Effects on Growth and Mycotoxin Production by Alternaria alternata Strains Isolated from Malbec Wine Grapes. J. Appl. Microbiol. 2017, 122, 481–492. [Google Scholar] [CrossRef]
- Prendes, L.P.; Merín, M.G.; Zachetti, V.G.L.; Pereyra, A.; Ramirez, M.L.; Morata de Ambrosini, V.I. Impact of Antagonistic Yeasts from Wine Grapes on Growth and Mycotoxin Production by Alternaria alternata. J. Appl. Microbiol. 2021, 131, 833–843. [Google Scholar] [CrossRef]
- Fedele, G.; Bove, F.; González-Domínguez, E.; Rossi, V. A Generic Model Accounting for the Interactions among Pathogens, Host Plants, Biocontrol Agents, and the Environment, with Parametrization for Botrytis cinerea on Grapevines. Agronomy 2020, 10, 222. [Google Scholar] [CrossRef]
- Fedele, G.; Brischetto, C.; Rossi, V. Biocontrol of Botrytis cinerea on Grape Berries as Influenced by Temperature and Humidity. Front. Plant Sci. 2020, 11, 1232. [Google Scholar] [CrossRef] [PubMed]
- Calvo-Garrido, C.; Roudet, J.; Aveline, N.; Davidou, L.; Dupin, S.; Fermaud, M. Microbial Antagonism Toward Botrytis Bunch Rot of Grapes in Multiple Field Tests Using One Bacillus ginsengihumi Strain and Formulated Biological Control Products. Front. Plant Sci. 2019, 10, 105. [Google Scholar] [CrossRef] [PubMed]
- Viñas, I.; Usall, J.; Teixidó, N.; Sanchis, V. Biological Control of Major Postharvest Pathogens on Apple with Candida Sake. Int. J. Food Microbiol. 1998, 40, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Cañamás, T.P.; Viñas, I.; Torres, R.; Usall, J.; Solsona, C.; Teixidó, N. Field Applications of Improved Formulations of Candida sake CPA-1 for Control of Botrytis cinerea in Grapes. Biol. Control 2011, 56, 150–158. [Google Scholar] [CrossRef]
- Calvo-Garrido, C.; Viñas, I.; Elmer, P.; Usall, J.; Teixidó, N. Candida Sake CPA-1 and Other Biologically Based Products as Potential Control Strategies to Reduce Sour Rot of Grapes. Lett. Appl. Microbiol. 2013, 57, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Calvo-Garrido, C.; Viñas, I.; Usall, J.; Rodríguez-Romera, M.; Ramos, M.C.; Teixidó, N. Survival of the Biological Control Agent Candida sake CPA-1 on Grapes under the Influence of Abiotic Factors. J. Appl. Microbiol. 2014, 117, 800–811. [Google Scholar] [CrossRef] [PubMed]
- Calvo-Garrido, C.; Teixidó, N.; Roudet, J.; Viñas, I.; Usall, J.; Fermaud, M. Biological Control of Botrytis Bunch Rot in Atlantic Climate Vineyards with Candida sake CPA-1 and Its Survival under Limiting Conditions of Temperature and Humidity. Biol. Control 2014, 79, 24–35. [Google Scholar] [CrossRef]
- Pertot, I.; Giovannini, O.; Benanchi, M.; Caffi, T.; Rossi, V.; Mugnai, L. Combining Biocontrol Agents with Different Mechanisms of Action in a Strategy to Control Botrytis cinerea on Grapevine. Crop Prot. 2017, 97, 85–93. [Google Scholar] [CrossRef]
- Galli, V.; Romboli, Y.; Barbato, D.; Mari, E.; Venturi, M.; Guerrini, S.; Granchi, L. Indigenous Aureobasidium pullulans Strains as Biocontrol Agents of Botrytis cinerea on Grape Berries. Sustainability 2021, 13, 9389. [Google Scholar] [CrossRef]
- Ponsone, M.L.; Nally, M.C.; Chiotta, M.L.; Combina, M.; Köhl, J.; Chulze, S.N. Evaluation of the Effectiveness of Potential Biocontrol Yeasts against Black Sur Rot and Ochratoxin a Occurring under Greenhouse and Field Grape Production Conditions. Biol. Control 2016, 103, 78–85. [Google Scholar] [CrossRef]
- de Felice, D.V.; Solfrizzo, M.; De Curtis, F.; Lima, G.; Visconti, A.; Castoria, R. Strains of Aureobasidium pullulans Can Lower Ochratoxin A Contamination in Wine Grapes. Phytopathology 2008, 98, 1261–1270. [Google Scholar] [CrossRef]
- Dimakopoulou, M.; Tjamos, S.E.; Antoniou, P.P.; Pietri, A.; Battilani, P.; Avramidis, N.; Markakis, E.A.; Tjamos, E.C. Phyllosphere Grapevine Yeast Aureobasidium pullulans Reduces Aspergillus Carbonarius (Sour Rot) Incidence in Wine-Producing Vineyards in Greece. Biol. Control 2008, 46, 158–165. [Google Scholar] [CrossRef]
- Pantelides, I.S.; Christou, O.; Tsolakidou, M.-D.; Tsaltas, D.; Ioannou, N. Isolation, Identification and in Vitro Screening of Grapevine Yeasts for the Control of Black Aspergilli on Grapes. Biol. Control 2015, 88, 46–53. [Google Scholar] [CrossRef]
- Carbó, A.; Torres, R.; Usall, J.; Solsona, C.; Teixidó, N. Fluidised-Bed Spray-Drying Formulations of Candida sake CPA-1 by Adding Biodegradable Coatings to Enhance Their Survival under Stress Conditions. Appl. Microbiol. Biotechnol. 2017, 101, 7865–7876. [Google Scholar] [CrossRef] [PubMed]
- Carbó, A.; Torres, R.; Teixidó, N.; Usall, J.; Medina, A.; Magan, N. Impact of Climate Change Environmental Conditions on the Resilience of Different Formulations of the Biocontrol Agent Candida sake CPA-1 on Grapes. Lett. Appl. Microbiol. 2018, 67, 2–8. [Google Scholar] [CrossRef]
- Carbó, A.; Torres, R.; Usall, J.; Marín, A.; Chiralt, A.; Teixidó, N. Novel Film-forming Formulations of the Biocontrol Agent Candida sake CPA-1: Biocontrol Efficacy and Performance at Field Conditions in Organic Wine Grapes. Pest Manag. Sci. 2019, 75, 959–968. [Google Scholar] [CrossRef] [PubMed]
- Carbó, A.; Teixidó, N.; Usall, J.; Torres, R. Verifying the Biocontrol Activity of Novel Film-forming Formulations of Candida Sake CPA-1: Resilience in Relation to Environmental Factors, Rainfall Episodes, and Control of Botrytis cinerea on Different Hosts. J. Sci. Food Agric. 2019, 99, 4969–4976. [Google Scholar] [CrossRef] [PubMed]
- Abadias, M.; Teixido, N.; Usall, J.; Vinas, I. Optimization of Growth Conditions of the Postharvest Biocontrol Agent Candida sake CPA-1 in a Lab-Scale Fermenter. J. Appl. Microbiol. 2003, 95, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Pelinski, R.; Cerrutti, P.; Ponsone, M.L.; Chulze, S.; Galvagno, M. Statistical Optimization of Simple Culture Conditions to Produce Biomass of an Ochratoxigenic Mould Biocontrol Yeast Strain. Lett. Appl. Microbiol. 2012, 54, 377–382. [Google Scholar] [CrossRef]
- Spadaro, D.; Droby, S. Development of Biocontrol Products for Postharvest Diseases of Fruit: The Importance of Elucidating the Mechanisms of Action of Yeast Antagonists. Trends Food Sci. Technol. 2016, 47, 39–49. [Google Scholar] [CrossRef]
- Köhl, J.; Kolnaar, R.; Ravensberg, W.J. Mode of Action of Microbial Biological Control Agents Against Plant Diseases: Relevance Beyond Efficacy. Front. Plant Sci. 2019, 10, 845. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, B.; Zhang, Z.; Chen, Y.; Tian, S. Antagonistic Yeasts: A Promising Alternative to Chemical Fungicides for Controlling Postharvest Decay of Fruit. J. Fungi 2020, 6, 158. [Google Scholar] [CrossRef] [PubMed]
- Nally, M.C.; Pesce, V.M.; Maturano, Y.P.; Toro, M.E.; Combina, M.; Castellanos de Figueroa, L.I.; Vazquez, F. Biocontrol of Fungi Isolated from Sour Rot Infected Table Grapes by Saccharomyces and Other Yeast Species. Postharvest Biol. Technol. 2013, 86, 456–462. [Google Scholar] [CrossRef]
- Fernandez-San Millan, A.; Larraya, L.; Farran, I.; Ancin, M.; Veramendi, J. Successful Biocontrol of Major Postharvest and Soil-Borne Plant Pathogenic Fungi by Antagonistic Yeasts. Biol. Control 2021, 160, 104683. [Google Scholar] [CrossRef]
- Maluleke, E.; Jolly, N.P.; Patterton, H.G.; Setati, M.E. Antifungal Activity of Non-Conventional Yeasts against Botrytis cinerea and Non-Botrytis Grape Bunch Rot Fungi. Front. Microbiol. 2022, 13, 986229. [Google Scholar] [CrossRef] [PubMed]
- Nally, M.C.; Ponsone, M.L.; Pesce, V.M.; Toro, M.E.; Vazquez, F.; Chulze, S. Evaluation of Behaviour of Lachancea thermotolerans Biocontrol Agents on Grape Fermentations. Lett. Appl. Microbiol. 2018, 67, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Alexandre, H.; Puyo, M.; Tourdot-Maréchal, R. Bioprotection in Winemaking. In New Advances in Saccharomyces; IntechOpen: London, UK, 2023. [Google Scholar]
- Ciani, M.; Capece, A.; Comitini, F.; Canonico, L.; Siesto, G.; Romano, P. Yeast Interactions in Inoculated Wine Fermentation. Front. Microbiol. 2016, 7, 555. [Google Scholar] [CrossRef]
- Maicas, S.; Mateo, J.J. The Life of Saccharomyces and Non-Saccharomyces Yeasts in Drinking Wine. Microorganisms 2023, 11, 1178. [Google Scholar] [CrossRef]
- Malfeito-Ferreira, M.; Silva, A.C. Yeasts in the Production of Wine; Romano, P., Ciani, M., Fleet, G.H., Eds.; Springer: New York, NY, USA, 2019; ISBN 978-1-4939-9780-0. [Google Scholar]
- Malfeito-Ferreira, M. Two Decades of “Horse Sweat” Taint and Brettanomyces Yeasts in Wine: Where Do We Stand Now? Beverages 2018, 4, 32. [Google Scholar] [CrossRef]
- Hernández, A.; Pérez-Nevado, F.; Ruiz-Moyano, S.; Serradilla, M.J.; Villalobos, M.C.; Martín, A.; Córdoba, M.G. Spoilage Yeasts: What Are the Sources of Contamination of Foods and Beverages? Int. J. Food Microbiol. 2018, 286, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Sáez, J.S.; Lopes, C.A.; Kirs, V.C.; Sangorrín, M.P. Enhanced Volatile Phenols in Wine Fermented with Saccharomyces cerevisiae and Spoiled with Pichia guilliermondii and Dekkera bruxellensis. Lett. Appl. Microbiol. 2010, 51, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Saez, J.S.; Lopes, C.A.; Kirs, V.E.; Sangorrín, M. Production of Volatile Phenols by Pichia manshurica and Pichia membranifaciens Isolated from Spoiled Wines and Cellar Environment in Patagonia. Food Microbiol. 2011, 28, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Sangorrín, M.P.; Lopes, C.A.; Jofré, V.; Querol, A.; Caballero, A.C. Spoilage Yeasts from Patagonian Cellars: Characterization and Potential Biocontrol Based on Killer Interactions. World J. Microbiol. Biotechnol. 2008, 24, 945–953. [Google Scholar] [CrossRef]
- Pinto, L.; Baruzzi, F.; Cocolin, L.; Malfeito-Ferreira, M. Emerging Technologies to Control Brettanomyces Spp. in Wine: Recent Advances and Future Trends. Trends Food Sci. Technol. 2020, 99, 88–100. [Google Scholar] [CrossRef]
- Lebleux, M.; Abdo, H.; Coelho, C.; Basmaciyan, L.; Albertin, W.; Maupeu, J.; Laurent, J.; Roullier-Gall, C.; Alexandre, H.; Guilloux-Benatier, M.; et al. New Advances on the Brettanomyces bruxellensis Biofilm Mode of Life. Int. J. Food Microbiol. 2020, 318, 108464. [Google Scholar] [CrossRef] [PubMed]
- Cibrario, A.; Miot-Sertier, C.; Paulin, M.; Bullier, B.; Riquier, L.; Perello, M.-C.; de Revel, G.; Albertin, W.; Masneuf-Pomarède, I.; Ballestra, P.; et al. Brettanomyces bruxellensis Phenotypic Diversity, Tolerance to Wine Stress and Wine Spoilage Ability. Food Microbiol. 2020, 87, 103379. [Google Scholar] [CrossRef] [PubMed]
- Ubeda, C.; Hornedo-Ortega, R.; Cerezo, A.B.; Garcia-Parrilla, M.C.; Troncoso, A.M. Chemical Hazards in Grapes and Wine, Climate Change and Challenges to Face. Food Chem. 2020, 314, 126222. [Google Scholar] [CrossRef]
- Aranda, A.; Jiménez-Martí, E.; Orozco, H.; Matallana, E.; del Olmo, M. Sulfur and Adenine Metabolisms Are Linked, and Both Modulate Sulfite Resistance in Wine Yeast. J. Agric. Food Chem. 2006, 54, 5839–5846. [Google Scholar] [CrossRef]
- Guerrero, R.F.; Cantos-Villar, E. Demonstrating the Efficiency of Sulphur Dioxide Replacements in Wine: A Parameter Review. Trends Food Sci. Technol. 2015, 42, 27–43. [Google Scholar] [CrossRef]
- Divol, B.; du Toit, M.; Duckitt, E. Surviving in the Presence of Sulphur Dioxide: Strategies Developed by Wine Yeasts. Appl. Microbiol. Biotechnol. 2012, 95, 601–613. [Google Scholar] [CrossRef] [PubMed]
- Dimopoulou, M.; Renault, M.; Dols-Lafargue, M.; Albertin, W.; Herry, J.-M.; Bellon-Fontaine, M.-N.; Masneuf-Pomarede, I. Microbiological, Biochemical, Physicochemical Surface Properties and Biofilm Forming Ability of Brettanomyces bruxellensis. Ann. Microbiol. 2019, 69, 1217–1225. [Google Scholar] [CrossRef]
- Lisanti, M.T.; Blaiotta, G.; Nioi, C.; Moio, L. Alternative Methods to SO 2 for Microbiological Stabilization of Wine. Compr. Rev. Food Sci. Food Saf. 2019, 18, 455–479. [Google Scholar] [CrossRef] [PubMed]
- Morata, A.; Loira, I.; Vejarano, R.; González, C.; Callejo, M.J.; Suárez-Lepe, J.A. Emerging Preservation Technologies in Grapes for Winemaking. Trends Food Sci. Technol. 2017, 67, 36–43. [Google Scholar] [CrossRef]
- Di Gianvito, P.; Englezos, V.; Rantsiou, K.; Cocolin, L. Bioprotection Strategies in Winemaking. Int. J. Food Microbiol. 2022, 364, 109532. [Google Scholar] [CrossRef] [PubMed]
- Chi, Z.; Wang, F.; Chi, Z.; Yue, L.; Liu, G.; Zhang, T. Bioproducts from Aureobasidium pullulans, a Biotechnologically Important Yeast. Appl. Microbiol. Biotechnol. 2009, 82, 793–804. [Google Scholar] [CrossRef] [PubMed]
- Agarbati, A.; Ciani, M.; Esin, S.; Agnolucci, M.; Marcheggiani, F.; Tiano, L.; Comitini, F. Comparative Zymocidial Effect of Three Different Killer Toxins against Brettanomyces bruxellensis Spoilage Yeasts. Int. J. Mol. Sci. 2023, 24, 1309. [Google Scholar] [CrossRef] [PubMed]
- Comitini, F.; Ciani, M. Kluyveromyces wickerhamii Killer Toxin: Purification and Activity towards Brettanomyces/Dekkera Yeasts in Grape Must. FEMS Microbiol. Lett. 2011, 316, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Berbegal, C.; Spano, G.; Fragasso, M.; Grieco, F.; Russo, P.; Capozzi, V. Starter Cultures as Biocontrol Strategy to Prevent Brettanomyces bruxellensis Proliferation in Wine. Appl. Microbiol. Biotechnol. 2018, 102, 569–576. [Google Scholar] [CrossRef]
- Fernández de Ullivarri, M.; Mendoza, L.M.; Raya, R.R. Characterization of the Killer Toxin KTCf20 from Wickerhamomyces anomalus, a Potential Biocontrol Agent against Wine Spoilage Yeasts. Biol. Control 2018, 121, 223–228. [Google Scholar] [CrossRef]
- Fernández de Ullivarri, M.; Merín, M.G.; Raya, R.R.; Morata de Ambrosini, V.I.; Mendoza, L.M. Killer Yeasts Used as Starter Cultures to Modulate the Behavior of Potential Spoilage Non-Saccharomyces Yeasts during Malbec Wine Fermentation. Food Biosci. 2024, 57, 103424. [Google Scholar] [CrossRef]
- Ghoul, M.; Mitri, S. The Ecology and Evolution of Microbial Competition. Trends Microbiol. 2016, 24, 833–845. [Google Scholar] [CrossRef] [PubMed]
- Mendoza, L.M.; Manca de Nadra, M.C.; Farías, M.E. Antagonistic Interaction between Yeasts and Lactic Acid Bacteria of Oenological Relevance. Food Res. Int. 2010, 43, 1990–1998. [Google Scholar] [CrossRef]
- Albergaria, H.; Arneborg, N. Dominance of Saccharomyces cerevisiae in Alcoholic Fermentation Processes: Role of Physiological Fitness and Microbial Interactions. Appl. Microbiol. Biotechnol. 2016, 100, 2035–2046. [Google Scholar] [CrossRef] [PubMed]
- Peña, R.; Ganga, M.A. Novel Antimicrobial Peptides Produced by Candida intermedia LAMAP1790 Active against the Wine-Spoilage Yeast Brettanomyces bruxellensis. Antonie Van Leeuwenhoek 2019, 112, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Lowes, K.F.; Shearman, C.A.; Payne, J.; MacKenzie, D.; Archer, D.B.; Merry, R.J.; Gasson, M.J. Prevention of Yeast Spoilage in Feed and Food by the Yeast Mycocin HMK. Appl. Environ. Microbiol. 2000, 66, 1066–1076. [Google Scholar] [CrossRef]
- Comitini, F.; Agarbati, A.; Canonico, L.; Galli, E.; Ciani, M. Purification and Characterization of WA18, a New Mycocin Produced by Wickerhamomyces anomalus Active in Wine Against Brettanomyces bruxellensis Spoilage Yeasts. Microorganisms 2020, 9, 56. [Google Scholar] [CrossRef] [PubMed]
- Belda, I.; Ruiz, J.; Alonso, A.; Marquina, D.; Santos, A. The Biology of Pichia membranifaciens Killer Toxins. Toxins 2017, 9, 112. [Google Scholar] [CrossRef] [PubMed]
- Kast, A.; Klassen, R.; Meinhardt, F. RRNA Fragmentation Induced by a Yeast Killer Toxin. Mol. Microbiol. 2014, 91, 606–617. [Google Scholar] [CrossRef]
- Santos, A.; Navascués, E.; Bravo, E.; Marquina, D. Ustilago Maydis Killer Toxin as a New Tool for the Biocontrol of the Wine Spoilage Yeast Brettanomyces bruxellensis. Int. J. Food Microbiol. 2011, 145, 147–154. [Google Scholar] [CrossRef]
- Liu, G.-L.; Chi, Z.; Wang, G.-Y.; Wang, Z.-P.; Li, Y.; Chi, Z.-M. Yeast Killer Toxins, Molecular Mechanisms of Their Action and Their Applications. Crit. Rev. Biotechnol. 2015, 35, 222–234. [Google Scholar] [CrossRef] [PubMed]
- Bevan, E.A.; Makower, M. The Physiological Basis of the Killer Character in Yeast. In Proceedings of the XI International Congress of Genetics, The Hague, The Netherlands, September 1963; 1963; pp. 202–203. [Google Scholar]
- de Ullivarri, M.F.; Mendoza, L.M.; Raya, R.R.; Farías, M.E. Killer Phenotype of Indigenous Yeasts Isolated from Argentinian Wine Cellars and Their Potential Starter Cultures for Winemaking. Biotechnol. Lett. 2011, 33, 2177–2183. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.; San Mauro, M.; Bravo, E.; Marquina, D. PMKT2, a New Killer Toxin from Pichia membranifaciens, and Its Promising Biotechnological Properties for Control of the Spoilage Yeast Brettanomyces bruxellensis. Microbiology 2009, 155, 624–634. [Google Scholar] [CrossRef] [PubMed]
- Vicente, J.; Calderón, F.; Santos, A.; Marquina, D.; Benito, S. High Potential of Pichia kluyveri and Other Pichia Species in Wine Technology. Int. J. Mol. Sci. 2021, 22, 1196. [Google Scholar] [CrossRef] [PubMed]
- Ciani, M.; Fatichenti, F. Killer Toxin of Kluyveromyces phaffii DBVPG 6076 as a Biopreservative Agent To Control Apiculate Wine Yeasts. Appl. Environ. Microbiol. 2001, 67, 3058–3063. [Google Scholar] [CrossRef] [PubMed]
- Comitini, F.; Ingeniis, J.; Pepe, L.; Mannazzu, I.; Ciani, M. Pichia Anomala and Kluyveromyces wickerhamii Killer Toxins as New Tools against Dekkera/Brettanomyces Spoilage Yeasts. FEMS Microbiol. Lett. 2004, 238, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Oro, L.; Ciani, M.; Bizzaro, D.; Comitini, F. Evaluation of Damage Induced by Kwkt and Pikt Zymocins against Brettanomyces/Dekkera Spoilage Yeast, as Compared to Sulphur Dioxide. J. Appl. Microbiol. 2016, 121, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Mehlomakulu, N.N.; Setati, M.E.; Divol, B. Characterization of Novel Killer Toxins Secreted by Wine-Related Non-Saccharomyces Yeasts and Their Action on Brettanomyces Spp. Int. J. Food Microbiol. 2014, 188, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Mehlomakulu, N.N.; Prior, K.J.; Setati, M.E.; Divol, B. Candida pyralidae Killer Toxin Disrupts the Cell Wall of Brettanomyces bruxellensis in Red Grape Juice. J. Appl. Microbiol. 2017, 122, 747–758. [Google Scholar] [CrossRef]
- Labbani, F.-Z.K.; Turchetti, B.; Bennamoun, L.; Dakhmouche, S.; Roberti, R.; Corazzi, L.; Meraihi, Z.; Buzzini, P. A Novel Killer Protein from Pichia kluyveri Isolated from an Algerian Soil: Purification and Characterization of Its in Vitro Activity against Food and Beverage Spoilage Yeasts. Antonie Van Leeuwenhoek 2015, 107, 961–970. [Google Scholar] [CrossRef]
- Buzzini, P.; Turchetti, B.; Vaughan-Martini, A.E. The Use of Killer Sensitivity Patterns for Biotyping Yeast Strains: The State of the Art, Potentialities and Limitations. FEMS Yeast Res. 2007, 7, 749–760. [Google Scholar] [CrossRef] [PubMed]
- Sangorrín, M.P.; Lopes, C.A.; Giraudo, M.R.; Caballero, A.C. Diversity and Killer Behaviour of Indigenous Yeasts Isolated from the Fermentation Vat Surfaces in Four Patagonian Wineries. Int. J. Food Microbiol. 2007, 119, 351–357. [Google Scholar] [CrossRef]
- Ramírez, M.; Velázquez, R.; Maqueda, M.; López-Piñeiro, A.; Ribas, J.C. A New Wine Torulaspora delbrueckii Killer Strain with Broad Antifungal Activity and Its Toxin-Encoding Double-Stranded RNA Virus. Front. Microbiol. 2015, 6, 983. [Google Scholar] [CrossRef]
- Lopes, C.A.; Sáez, J.S.; Sangorrín, M.P. Differential Response of Pichia guilliermondii Spoilage Isolates to Biological and Physico-Chemical Factors Prevailing in Patagonian Wine Fermentations. Can. J. Microbiol. 2009, 55, 801–809. [Google Scholar] [CrossRef]
- Lopes, C.A.; Sangorrín, M.P. Optimization of Killer Assays for Yeast Selection Protocols. Rev. Argent. Microbiol. 2010, 42, 298–306. [Google Scholar] [CrossRef]
- Velázquez, R.; Zamora, E.; Álvarez, M.L.; Hernández, L.M.; Ramírez, M. Effects of New Torulaspora delbrueckii Killer Yeasts on the Must Fermentation Kinetics and Aroma Compounds of White Table Wine. Front. Microbiol. 2015, 6, 1222. [Google Scholar] [CrossRef] [PubMed]
- Vejarano, R.; Gil-Calderón, A. Commercially Available Non-Saccharomyces Yeasts for Winemaking: Current Market, Advantages over Saccharomyces, Biocompatibility, and Safety. Fermentation 2021, 7, 171. [Google Scholar] [CrossRef]
- Comitini, F.; Capece, A.; Ciani, M.; Romano, P. New Insights on the Use of Wine Yeasts. Curr. Opin. Food Sci. 2017, 13, 44–49. [Google Scholar] [CrossRef]
- Csoma, H.; Kállai, Z.; Antunovics, Z.; Czentye, K.; Sipiczki, M. Vinification without Saccharomyces: Interacting Osmotolerant and “Spoilage” Yeast Communities in Fermenting and Ageing Botrytised High-Sugar Wines (Tokaj Essence). Microorganisms 2020, 9, 19. [Google Scholar] [CrossRef]
- Ramírez, M.; Velázquez, R.; Maqueda, M.; Zamora, E.; López-Piñeiro, A.; Hernández, L.M. Influence of the Dominance of Must Fermentation by Torulaspora delbrueckii on the Malolactic Fermentation and Organoleptic Quality of Red Table Wine. Int. J. Food Microbiol. 2016, 238, 311–319. [Google Scholar] [CrossRef]
- Velázquez, R.; Zamora, E.; Álvarez, M.L.; Ramírez, M. Using Torulaspora delbrueckii Killer Yeasts in the Elaboration of Base Wine and Traditional Sparkling Wine. Int. J. Food Microbiol. 2019, 289, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Simonin, S.; Alexandre, H.; Nikolantonaki, M.; Coelho, C.; Tourdot-Maréchal, R. Inoculation of Torulaspora delbrueckii as a Bio-Protection Agent in Winemaking. Food Res. Int. 2018, 107, 451–461. [Google Scholar] [CrossRef] [PubMed]
- Claus, H.; Mojsov, K. Enzymes for Wine Fermentation: Current and Perspective Applications. Fermentation 2018, 4, 52. [Google Scholar] [CrossRef]
- Hüfner, E.; Haßelbeck, G. Application of Microbial Enzymes During Winemaking. In Biology of Microorganisms on Grapes, in Must and in Wine; Springer International Publishing: Cham, Switzerland, 2017; pp. 635–658. [Google Scholar]
- Mojsov, K.; Andronikov, D.; Janevski, A.; Jordeva, S.; Zezova, S. Enzymes and Wine: The Enhanced Quality and Yield. Savrem. Tehnol. 2015, 4, 94–100. [Google Scholar] [CrossRef]
- Osete-Alcaraz, A.; Gómez-Plaza, E.; Pérez-Porras, P.; Bautista-Ortín, A.B. Revisiting the Use of Pectinases in Enology: A Role beyond Facilitating Phenolic Grape Extraction. Food Chem. 2022, 372, 131282. [Google Scholar] [CrossRef]
- Longhi, S.J.; Martín, M.C.; Merín, M.G.; Morata de Ambrosini, V.I. Yeast Multi-Enzymatic Systems for Improving Colour Extraction, Technological Parameters and Antioxidant Activity of Wine. Food Technol. Biotechnol. 2022, 60, 556–570. [Google Scholar] [CrossRef] [PubMed]
- Mojsov, K. Use of Enzymes in Wine Making: A Review. Int. J. Mark. Technol. 2013, 3, 112–127. [Google Scholar]
- John, J.; Kaimal, K.K.S.; Smith, M.L.; Rahman, P.K.S.M.; Chellam, P.V. Advances in Upstream and Downstream Strategies of Pectinase Bioprocessing: A Review. Int. J. Biol. Macromol. 2020, 162, 1086–1099. [Google Scholar] [CrossRef]
- Bautista-Ortín, A.B.; Martínez-Cutillas, A.; Ros-García, J.M.; López-Roca, J.M.; Gómez-Plaza, E. Improving Colour Extraction and Stability in Red Wines: The Use of Maceration Enzymes and Enological Tannins. Int. J. Food Sci. Technol. 2005, 40, 867–878. [Google Scholar] [CrossRef]
- Gómez-Plaza, E.; Romero-Cascales, I.; Bautista-Ortín, A. Use of Enzymes for Wine Production. In Enzymes in Fruit and Vegetable Processing; CRC Press: Boca Raton, FL, USA, 2010; pp. 215–244. [Google Scholar]
- Zimman, A.; Joslin, W.S.; Lyon, M.L.; Meier, J.; Waterhouse, A.L. Maceration Variables Affecting Phenolic Composition in Commercial-Scale Cabernet Sauvignon Winemaking Trials. Am. J. Enol. Vitic. 2002, 53, 93–98. [Google Scholar] [CrossRef]
- Romero-Cascales, I.; Fernández-Fernández, J.I.; Ros-García, J.M.; López-Roca, J.M.; Gómez-Plaza, E. Characterisation of the Main Enzymatic Activities Present in Six Commercial Macerating Enzymes and Their Effects on Extracting Colour during Winemaking of Monastrell Grapes. Int. J. Food Sci. Technol. 2008, 43, 1295–1305. [Google Scholar] [CrossRef]
- Río Segade, S.; Pace, C.; Torchio, F.; Giacosa, S.; Gerbi, V.; Rolle, L. Impact of Maceration Enzymes on Skin Softening and Relationship with Anthocyanin Extraction in Wine Grapes with Different Anthocyanin Profiles. Food Res. Int. 2015, 71, 50–57. [Google Scholar] [CrossRef]
- Martín, M.C.; Morata de Ambrosini, V.I. Effect of a Cold-active Pectinolytic System on Colour Development of Malbec Red Wines Elaborated at Low Temperature. Int. J. Food Sci. Technol. 2014, 49, 1893–1901. [Google Scholar] [CrossRef]
- Dal Magro, L.; Hertz, P.F.; Fernandez-Lafuente, R.; Klein, M.P.; Rodrigues, R.C. Preparation and Characterization of a Combi-CLEAs from Pectinases and Cellulases: A Potential Biocatalyst for Grape Juice Clarification. RSC Adv. 2016, 6, 27242–27251. [Google Scholar] [CrossRef]
- Kanellaki, M.; Bekatorou, A.; Koutinas, A.A. Low-Temperature Production of Wine, Beer, and Distillates Using Cold-Adapted Yeasts. In Cold-Adapted Yeasts; Springer Berlin Heidelberg: Berlin/Heidelberg, Germany, 2014; pp. 417–439. [Google Scholar]
- Nighojkar, A.; Patidar, M.K.; Nighojkar, S. Pectinases: Production and Applications for Fruit Juice Beverages. In Processing and Sustainability of Beverages; Elsevier: Amsterdam, The Netherlands, 2019; pp. 235–273. [Google Scholar]
- Echave, J.; Barral, M.; Fraga-Corral, M.; Prieto, M.A.; Simal-Gandara, J. Bottle Aging and Storage of Wines: A Review. Molecules 2021, 26, 713. [Google Scholar] [CrossRef]
- Li, S.; Bindon, K.; Bastian, S.E.P.; Jiranek, V.; Wilkinson, K.L. Use of Winemaking Supplements To Modify the Composition and Sensory Properties of Shiraz Wine. J. Agric. Food Chem. 2017, 65, 1353–1364. [Google Scholar] [CrossRef]
- Main, G.L.; Morris, J.R. Effect of Macerating Enzymes and Postfermentation Grape-Seed Tannin on the Color of Cynthiana Wines. Am. J. Enol. Vitic. 2007, 58, 365–372. [Google Scholar] [CrossRef]
- Mojsov, K. The Effects of Different Carbon Sources on Biosynthesis of Pectinolytic Enzymes by Aspergillus niger. Appl. Technol. Innov. 2010, 3, 23–29. [Google Scholar] [CrossRef]
- Osete-Alcaraz, A.; Bautista-Ortín, A.B.; Ortega-Regules, A.E.; Gómez-Plaza, E. Combined Use of Pectolytic Enzymes and Ultrasounds for Improving the Extraction of Phenolic Compounds During Vinification. Food Bioprocess Technol. 2019, 12, 1330–1339. [Google Scholar] [CrossRef]
- Romero-Cascales, I.; Ros-García, J.M.; López-Roca, J.M.; Gómez-Plaza, E. The Effect of a Commercial Pectolytic Enzyme on Grape Skin Cell Wall Degradation and Colour Evolution during the Maceration Process. Food Chem. 2012, 130, 626–631. [Google Scholar] [CrossRef]
- Guler, A. Effects of Different Maceration Techniques on the Colour, Polyphenols and Antioxidant Capacity of Grape Juice. Food Chem. 2023, 404, 134603. [Google Scholar] [CrossRef]
- Vejarano, R.; Luján-Corro, M. Red Wine and Health: Approaches to Improve the Phenolic Content During Winemaking. Front. Nutr. 2022, 9, 890066. [Google Scholar] [CrossRef]
- Fernández, M.; Úbeda, J.F.; Briones, A.I. Typing of Non-Saccharomyces Yeasts with Enzymatic Activities of Interest in Wine-Making. Int. J. Food Microbiol. 2000, 59, 29–36. [Google Scholar] [CrossRef]
- Rahman, M.M.; Rahaman, M.S.; Islam, M.R.; Rahman, F.; Mithi, F.M.; Alqahtani, T.; Almikhlafi, M.A.; Alghamdi, S.Q.; Alruwaili, A.S.; Hossain, M.S.; et al. Role of Phenolic Compounds in Human Disease: Current Knowledge and Future Prospects. Molecules 2021, 27, 233. [Google Scholar] [CrossRef]
- Álvarez, I.; Aleixandre, J.L.; García, M.J.; Lizama, V. Impact of Prefermentative Maceration on the Phenolic and Volatile Compounds in Monastrell Red Wines. Anal. Chim. Acta 2006, 563, 109–115. [Google Scholar] [CrossRef]
- Gao, Y.; Fangel, J.U.; Willats, W.G.T.; Vivier, M.A.; Moore, J.P. Dissecting the Polysaccharide-Rich Grape Cell Wall Matrix Using Recombinant Pectinases during Winemaking. Carbohydr. Polym. 2016, 152, 510–519. [Google Scholar] [CrossRef]
- Osete-Alcaraz, A.; Gómez-Plaza, E.; Martínez-Pérez, P.; Weiller, F.; Schückel, J.; Willats, W.G.T.; Moore, J.P.; Ros-García, J.M.; Bautista-Ortín, A.B. The Impact of Carbohydrate-Active Enzymes on Mediating Cell Wall Polysaccharide-Tannin Interactions in a Wine-like Matrix. Food Res. Int. 2020, 129, 108889. [Google Scholar] [CrossRef]
- Osete-Alcaraz, A.; Gómez-Plaza, E.; Martínez-Pérez, P.; Weiller, F.; Schückel, J.; Willats, W.G.T.; Moore, J.P.; Ros-García, J.M.; Bautista-Ortín, A.B. The Influence of Hydrolytic Enzymes on Tannin Adsorption-Desorption onto Grape Cell Walls in a Wine-Like Matrix. Molecules 2021, 26, 770. [Google Scholar] [CrossRef]
- Zietsman, A.J.J.; Moore, J.P.; Fangel, J.U.; Willats, W.G.T.; Trygg, J.; Vivier, M.A. Following the Compositional Changes of Fresh Grape Skin Cell Walls during the Fermentation Process in the Presence and Absence of Maceration Enzymes. J. Agric. Food Chem. 2015, 63, 2798–2810. [Google Scholar] [CrossRef]
- Benucci, I.; Río Segade, S.; Cerreti, M.; Giacosa, S.; Paissoni, M.A.; Liburdi, K.; Bautista-Ortín, A.B.; Gómez-Plaza, E.; Gerbi, V.; Esti, M.; et al. Application of Enzyme Preparations for Extraction of Berry Skin Phenolics in Withered Winegrapes. Food Chem. 2017, 237, 756–765. [Google Scholar] [CrossRef]
- Gao, Y.; Zietsman, A.J.J.; Vivier, M.A.; Moore, J.P. Deconstructing Wine Grape Cell Walls with Enzymes During Winemaking: New Insights from Glycan Microarray Technology. Molecules 2019, 24, 165. [Google Scholar] [CrossRef]
- Apolinar-Valiente, R.; Romero-Cascales, I.; Gómez-Plaza, E.; Ros-García, J.M. Degradation of Monastrell Grape Skins: Effect of Individual Enzymatic Activities and Their Synergic Combination. Eur. Food Res. Technol. 2017, 243, 1933–1942. [Google Scholar] [CrossRef]
- Martínez-Lapuente, L.; Guadalupe, Z.; Ayestarán, B.; Pérez-Porras, P.; Bautista-Ortín, A.B.; Gómez-Plaza, E. Effects of Combining High Power Ultrasound and Enological Enzymes on the Composition of Polysaccharides in Red Wine. LWT 2022, 170, 114060. [Google Scholar] [CrossRef]
- Suhaimi, H.; Dailin, D.J.; Malek, R.A.; Hanapi, S.Z.; Ambehabati, K.K.; Keat, H.C.; Prakasham, S.; Elsayed, E.A.; Misson, M.; El Enshasy, H. Fungal Pectinases: Production and Applications in Food Industries. In Sustainable Food Production. Fungal Biology; Dai, X., Sharma, M., Chen, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2021; pp. 85–115. [Google Scholar]
- Blanco, P.; Sieiro, C.; Villa, T.G. Production of Pectic Enzymes in Yeasts. FEMS Microbiol. Lett. 1999, 175, 1–9. [Google Scholar] [CrossRef]
- Onetto, C.A.; Borneman, A.R.; Schmidt, S.A. Investigating the Effects of Aureobasidium pullulans on Grape Juice Composition and Fermentation. Food Microbiol. 2020, 90, 103451. [Google Scholar] [CrossRef]
- Longhi, S.J.; Martín, M.C.; Fontana, A.; Morata de Ambrosini, V.I. Different Approaches to Supplement Polysaccharide-Degrading Enzymes in Vinification: Effects on Color Extraction, Phenolic Composition, Antioxidant Activity and Sensory Profiles of Malbec Wines. Food Res. Int. 2022, 157, 111447. [Google Scholar] [CrossRef]
- Fernández-González, M.; Úbeda, J.F.; Cordero-Otero, R.R.; Thanvanthri Gururajan, V.; Briones, A.I. Engineering of an Oenological Saccharomyces cerevisiae Strain with Pectinolytic Activity and Its Effect on Wine. Int. J. Food Microbiol. 2005, 102, 173–183. [Google Scholar] [CrossRef]
- Fernández-González, M.; Úbeda, J.; Vasydevan, T.; Otero, R.; Briones, A. Evaluation of Polygalacturonase Activity in Wine Strains. FEMS Microbiol. Lett. 2004, 237, 261–266. [Google Scholar] [CrossRef][Green Version]
- Merín, M.G.; Morata de Ambrosini, V.I. Highly Cold-Active Pectinases under Wine-like Conditions from Non- Saccharomyces Yeasts for Enzymatic Production during Winemaking. Lett. Appl. Microbiol. 2015, 60, 467–474. [Google Scholar] [CrossRef]
- Gainvors, A.; Karam, N.; Lequart, C.; Belarbi, A. Use of Saccharomyces cerevisiae for the Clarification of Fruit Juices. Biotechnol. Lett. 1994, 16, 1329–1334. [Google Scholar] [CrossRef]
- Blanco, P.; Sieiro, C.; Díaz, A.; Reboredo, N.M.; Villa, T.G. Grape Juice Biodegradation by Polygalacturonases from Saccharomyces cerevisiae. Int. Biodeterior. Biodegrad. 1997, 40, 115–118. [Google Scholar] [CrossRef]
- Charoenchai, C.; Fleet, G.H.; Henschke, P.A.; Todd, B.E.N. Screening of Non- Saccharomyces Wine Yeasts for the Presence of Extracellular Hydrolytic Enzymes. Aust. J. Grape Wine Res. 1997, 3, 2–8. [Google Scholar] [CrossRef]
- Strauss, M.L.A.; Jolly, N.P.; Lambrechts, M.G.; van Rensburg, P. Screening for the Production of Extracellular Hydrolytic Enzymes by Non-Saccharomyces Wine Yeasts. J. Appl. Microbiol. 2001, 91, 182–190. [Google Scholar] [CrossRef]
- Belda, I.; Ruiz, J.; Alastruey-Izquierdo, A.; Navascués, E.; Marquina, D.; Santos, A. Unraveling the Enzymatic Basis of Wine “Flavorome”: A Phylo-Functional Study of Wine Related Yeast Species. Front. Microbiol. 2016, 7, 12. [Google Scholar] [CrossRef]
- Rodríguez, M.E.; Lopes, C.; Valles, S.; Giraudo, M.R.; Caballero, A. Selection and Preliminary Characterization of β-Glycosidases Producer Patagonian Wild Yeasts. Enzyme Microb. Technol. 2007, 41, 812–820. [Google Scholar] [CrossRef]
- Sahay, S.; Hamid, B.; Singh, P.; Ranjan, K.; Chauhan, D.; Rana, R.S.; Chaurse, V.K. Evaluation of Pectinolytic Activities for Oenological Uses from Psychrotrophic Yeasts. Lett. Appl. Microbiol. 2013, 57, 115–121. [Google Scholar] [CrossRef]
- Merín, M.G.; Mendoza, L.M.; Farías, M.E.; Morata de Ambrosini, V.I. Isolation and Selection of Yeasts from Wine Grape Ecosystem Secreting Cold-Active Pectinolytic Activity. Int. J. Food Microbiol. 2011, 147, 144–148. [Google Scholar] [CrossRef]
- Blanco, P.; Sieiro, C.; Di´az, A.; Villa, T.G. Differences between Pectic Enzymes Produced by Laboratory and Wild-Type Strains of Saccharomyces cerevisiae. World J. Microbiol. Biotechnol. 1997, 13, 711–712. [Google Scholar] [CrossRef]
- Belda, I.; Conchillo, L.B.; Ruiz, J.; Navascués, E.; Marquina, D.; Santos, A. Selection and Use of Pectinolytic Yeasts for Improving Clarification and Phenolic Extraction in Winemaking. Int. J. Food Microbiol. 2016, 223, 1–8. [Google Scholar] [CrossRef]
- Escribano-Viana, R.; Portu, J.; Garijo, P.; López, R.; Santamaría, P.; López-Alfaro, I.; Gutiérrez, A.R.; González-Arenzana, L. Effect of the Sequential Inoculation of Non-Saccharomyces/Saccharomyces on the Anthocyans and Stilbenes Composition of Tempranillo Wines. Front. Microbiol. 2019, 10, 773. [Google Scholar] [CrossRef]
- Benucci, I.; Cerreti, M.; Liburdi, K.; Nardi, T.; Vagnoli, P.; Ortiz-Julien, A.; Esti, M. Pre-Fermentative Cold Maceration in Presence of Non- Saccharomyces Strains: Evolution of Chromatic Characteristics of Sangiovese Red Wine Elaborated by Sequential Inoculation. Food Res. Int. 2018, 107, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Maturano, Y.P.; Rodríguez Assaf, L.A.; Toro, M.E.; Nally, M.C.; Vallejo, M.; Castellanos de Figueroa, L.I.; Combina, M.; Vazquez, F. Multi-Enzyme Production by Pure and Mixed Cultures of Saccharomyces and Non-Saccharomyces Yeasts during Wine Fermentation. Int. J. Food Microbiol. 2012, 155, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Merín, M.G.; Martín, M.C.; Rantsiou, K.; Cocolin, L.; de Ambrosini, V.I.M. Characterization of Pectinase Activity for Enology from Yeasts Occurring in Argentine Bonarda Grape. Brazilian J. Microbiol. 2015, 46, 815–823. [Google Scholar] [CrossRef] [PubMed]
- Merín, M.G.; Mendoza, L.M.; Morata de Ambrosini, V.I. Pectinolytic Yeasts from Viticultural and Enological Environments: Novel Finding of Filobasidium capsuligenum Producing Pectinases. J. Basic Microbiol. 2014, 54, 835–842. [Google Scholar] [CrossRef] [PubMed]
- Escribano, R.; González-Arenzana, L.; Garijo, P.; Berlanas, C.; López-Alfaro, I.; López, R.; Gutiérrez, A.R.; Santamaría, P. Screening of Enzymatic Activities within Different Enological Non-Saccharomyces Yeasts. J. Food Sci. Technol. 2017, 54, 1555–1564. [Google Scholar] [CrossRef] [PubMed]
- Samoticha, J.; Wojdyło, A.; Chmielewska, J.; Politowicz, J.; Szumny, A. The Effects of Enzymatic Pre-Treatment and Type of Yeast on Chemical Properties of White Wine. LWT-Food Sci. Technol. 2017, 79, 445–453. [Google Scholar] [CrossRef]
- Massera, A.; Assof, M.; Sari, S.; Ciklic, I.; Mercado, L.; Jofré, V.; Combina, M. Effect of Low Temperature Fermentation on the Yeast-Derived Volatile Aroma Composition and Sensory Profile in Merlot Wines. LWT 2021, 142, 111069. [Google Scholar] [CrossRef]
- Jayani, R.S.; Saxena, S.; Gupta, R. Microbial Pectinolytic Enzymes: A Review. Process Biochem. 2005, 40, 2931–2944. [Google Scholar] [CrossRef]
- Obreque-Slier, E.; Cortés-Araya, K.; Medel-Marabolí, M.; López-Solís, R. Different Effectiveness of Protein Fining Agents When Tested for Interaction and Precipitation with Tannic Acid, a Seed Polyphenol Extract and Seven Wines. OENO One 2023, 57, 13–28. [Google Scholar] [CrossRef]
- Oskay, M. Production, Partial Purification, and Characterization of Polygalacturonase from Aureobasidium pullulans P56 under Submerged Fermentation Using Agro-Industrial Wastes. Curr. Microbiol. 2022, 79, 296. [Google Scholar] [CrossRef]
- Sudeep, K.C.; Upadhyaya, J.; Joshi, D.R.; Lekhak, B.; Kumar Chaudhary, D.; Raj Pant, B.; Raj Bajgai, T.; Dhital, R.; Khanal, S.; Koirala, N.; et al. Production, Characterization, and Industrial Application of Pectinase Enzyme Isolated from Fungal Strains. Fermentation 2020, 6, 59. [Google Scholar] [CrossRef]
- Fleet, G.H. Wine Yeasts for the Future. FEMS Yeast Res. 2008, 8, 979–995. [Google Scholar] [CrossRef] [PubMed]
- Sternes, P.R.; Lee, D.; Kutyna, D.R.; Borneman, A.R. A Combined Meta-Barcoding and Shotgun Metagenomic Analysis of Spontaneous Wine Fermentation. Gigascience 2017, 6, gix040. [Google Scholar] [CrossRef] [PubMed]
- Prasongsuk, S.; Lotrakul, P.; Ali, I.; Bankeeree, W.; Punnapayak, H. The Current Status of Aureobasidium pullulans in Biotechnology. Folia Microbiol. 2018, 63, 129–140. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Enzymes | Enzyme-Producing Yeast Strain (Origin) | Grape Variety Used in Winemaking | Biotechnological Impacts | References |
|---|---|---|---|---|
| Exogenous enzymes | ||||
| Multienzymatic extract (mainly carbohydrolases) | Aureobasidium pullulans m11-2 (DO San Rafael-Mendoza wine region, Argentina) | Vitis vinifera L. cv. Malbec | Improvement in clarification and filtration. Increases in colour extraction and antioxidant capacity of wines. Great potential for prefermentative cold maceration. | [180] |
| Cold-active pectinases with side activities (cellulases and xylanases) | Strains of A. pullulans, Filobasidium capsuligenum, Rhodotorula dairenensis, Cryptococcus saitoi and Saccharomyces cerevisiae (DO San Rafael-Mendoza wine region, Argentina) | - | * Enzymes resistant to oenological conditions, with potential application for low-temperature winemaking. | [183] |
| Cold-active pectinolytic activities (pectinmetilesterases, exo- and endo-polygalacturonases) | Rhodotorula mucilaginosa PT1 and Cystofilobasidium capitatum SPY11 (from soil in different regions in India) | - | * Potential application in wine production and juice clarification at low temperature. | [190] |
| Pectinases cold-active | Strains of A. pullulans (DO San Rafael-Mendoza wine region, Argentina) | - | * Potential for winemaking at low temperature. | [191] |
| Polygalacturonase activity (PG) | S. cerevisiae (from viticultural area of Valdepena, Spain) | - | * Potential application for wine production with a strain S. cerevisiae with unusually high PG activity. | [182] |
| Polygalacturonase activity (PG) | S. cerevisiae (C.E.C.T., Colección Española de Cultivos Tipo) | Grape juice (from La Mancha, España) | Reduction in filtration times. | [192] |
| For “in situ” production | ||||
| Pectinases with side activities (cellulases and xylanases) | Torulaspora delbrueckii m7-2 in sequential fermentations with S. cerevisiae (DO San Rafael-Mendoza wine region, Argentina) | V. vinifera L. cv. Malbec | Improvement in clarification and filtration. Increases in colour extraction and antioxidant capacity of wines. | [180] |
| Pectinase activity | A. pullulans GM-R-22 (DO San Rafael-Mendoza wine region, Argentina) in both sequential and simultaneous inoculation with commercial S. cerevisiae | V. vinifera L. cv. Malbec | High efficiency in low-temperature winemaking and prefermentative cold maceration. Increases colour, total polyphenols, and stability. Wines with better tonality and flavour. Improvement in clarification efficiency. | [20] |
| Pectinase activity (with β-glucosidase and tannase side activities) | A. pullulans AWRI4229 and AWRI4231 (Adelaide Hills, South Australia, Australia) | V. vinifera L. cv. Chardonnay (grape juice) | Important implications for wine production and quality. Impacts grape juice composition and modulates fermentation kinetics by competition for trace elements. | [179] |
| Pectinase activity | A. pullulans GM-R-22 (DO San Rafael-Mendoza wine region, Argentina) in sequential inoculation with commercial S. cerevisiae | V. vinifera L. cv. Malbec | Great effectiveness in prefermentative cold maceration. Improvement in chromatic parameters and decreasing of filtration time and turbidity. Enhancement in desirable volatile compounds, such as esters and norisoprenoids. | [19] |
| Polygalacturonase activity | Metschnikowia pulcherrima NS-EM-34 (Spanish Designation of Origin Ribera del Duero) in sequential fermentations combined with commercial S. cerevisiae | V. vinifera L. cv. Tempranillo | Improvement in clarification process and liberation of phenolic compounds. Impacts sensorial aspects of wines such as their aromatic complexity and alcoholic content. | [193] |
| Pectinase, cellulase, xylanase, and glucanase activities (plus other aroma-related enzymes) | M. pulcherrima, Zygosaccharomyces bailii, Candida zeylanoides, and T. delbrueckii in sequential inoculation with S. cerevisiae (the Rioja “Qualified” Designation of Origin, The Rioja, Spain) | V. vinifera L. cv. Tempranillo | Improvement in monomeric anthocyanin and stilbene composition. Wines with better colour and likely healthy properties. | [194] |
| Pectinase activity with glucosidase side activity | M. pulcherrima MP 346 and Metschnikowia fructicola MF 98-3 in sequential inoculation with S. cerevisiae with prefermentative cold maceration (Lallemand SAS, Blagnac, France) | V. vinifera L. cv. Sangiovese | Enhancement in properties and stability of wine colour (higher levels of flavonoids and anthocyanins, and preservation of red tone). | [195] |
| Multiple enzymes (pectinases, β-glucosidases, proteases, amylases, and xylanases) | S. cerevisiae BSc562, Hanseniaspora vineae BHv438, and T. delbrueckii BTd259 in pure and mixed cultures (Culture Collection of Autochthonous Microorganisms at the Biotechnology Institute, Faculty of Engineering-National University of San Juan, Argentina) | V. vinifera L. cv. Pedro Giménez | Impacts aromatic characteristics and final quality of the wine by natural precursor hydrolysis of polymers present in grape juice. | [196] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martín, M.C.; Prendes, L.P.; Morata, V.I.; Merín, M.G. Biocontrol and Enzymatic Activity of Non-Saccharomyces Wine Yeasts: Improvements in Winemaking. Fermentation 2024, 10, 218. https://doi.org/10.3390/fermentation10040218
Martín MC, Prendes LP, Morata VI, Merín MG. Biocontrol and Enzymatic Activity of Non-Saccharomyces Wine Yeasts: Improvements in Winemaking. Fermentation. 2024; 10(4):218. https://doi.org/10.3390/fermentation10040218
Chicago/Turabian StyleMartín, María Carolina, Luciana Paola Prendes, Vilma Inés Morata, and María Gabriela Merín. 2024. "Biocontrol and Enzymatic Activity of Non-Saccharomyces Wine Yeasts: Improvements in Winemaking" Fermentation 10, no. 4: 218. https://doi.org/10.3390/fermentation10040218
APA StyleMartín, M. C., Prendes, L. P., Morata, V. I., & Merín, M. G. (2024). Biocontrol and Enzymatic Activity of Non-Saccharomyces Wine Yeasts: Improvements in Winemaking. Fermentation, 10(4), 218. https://doi.org/10.3390/fermentation10040218